Претектальная зона
| Претектальная зона | |
|---|---|
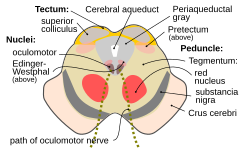 Поперечный срез среднего мозга на уровне верхних холмиков . Претектальная область обозначена (претектум) справа. | |
| Подробности | |
| Часть | Средний мозг |
| Части | переднее претектальное ядро, медиальное претектальное ядро, ядро зрительного тракта, оливковое претектальное ядро, заднее претектальное ядро, задние пределы, комиссуральная претектальная область |
| Идентификаторы | |
| латинский | претектальная зона |
| МеШ | Д066250 |
| Нейроимена | 467 |
| НейроЛекс ID | nlx_59721 |
| ТА98 | A14.1.08.505 A14.1.08.506 |
| ТА2 | 5739 |
| ФМА | 62402 |
| Анатомические термины нейроанатомии | |
В нейроанатомии , претектальная область или претектум , представляет собой структуру среднего мозга, состоящую из семи ядер и включающую часть подкорковой зрительной системы . Посредством реципрокных двусторонних проекций сетчатки он участвует в первую очередь в поведенческих реакциях на острые изменения окружающего света, таких как зрачковый световой рефлекс , оптокинетический рефлекс и временные изменения циркадного ритма . [1] [2] [3] [4] [5] Было обнаружено, что в дополнение к роли претектума в зрительной системе переднее претектальное ядро передает соматосенсорную и ноцицептивную информацию. [6] [7]
Расположение и структура [ править ]
Претектум представляет собой двустороннюю группу сильно связанных между собой ядер, расположенных вблизи соединения среднего и переднего мозга . [8] Претектум обычно классифицируется как структура среднего мозга, хотя из-за его близости к переднему мозгу его иногда классифицируют как часть каудального промежуточного мозга (переднего мозга). [9] У позвоночных претектум расположен непосредственно перед верхним холмиком и сзади от таламуса . Он расположен над околоводопроводной серой и ядром задней комиссуры . [10]
Внутри претектума было идентифицировано несколько ядер, хотя их границы может быть трудно определить, и ведутся споры о том, какие регионы следует включить и их точные названия. [1] [10] [11] Пять первичных ядер: оливковое претектальное ядро (ON), ядро зрительного тракта (NOT), переднее (NPA), медиальное (NPM) и заднее (NPP) претектальные ядра. НОТ состоит из относительно крупных клеток и расположен между верхними холмиками. ON расположен медиальнее NOT и имеет хвост, простирающийся между NOT и NPP, который вентральнее ON. [10] Также были идентифицированы два дополнительных ядра: задние пределы (PLi) и коммизуральная претектальная область (CPA). [12] Хотя эти две области не были изучены в такой же степени, как пять основных ядер, исследования показали, что и PLi, и CPA получают входные данные сетчатки, что предполагает их роль в обработке зрительной информации. [13]
Входы [ править ]
Претектум получает значительную бинокулярную информацию от светочувствительных ганглиозных клеток сетчатки. У приматов эти афференты двусторонние. [14] у грызунов они выступают из контралатеральной сетчатки. Большинство этих ретино-претектальных проекций направлены в положение «ВКЛ» и «НЕ». [14] в то время как другие претектальные ядра получают незначительный входной сигнал сетчатки у млекопитающих, включая задние, медиальные и передние претектальные ядра. [1] [10] [15] [16]
НЕ получает данные из нескольких регионов. Из таламуса НОТ получает тормозные проекции от ГАМК -продуцирующих нейронов ипсилатерального латерального коленчатого ядра и двусторонних межколенных листков. Ипсилатеральное поверхностное супрахиазматическое ядро , а также медиальное, дорсальное и латеральное терминальные ядра среднего мозга проецируются на НОТ. Волокна также проецируются в НОТ из гипоязычного ядра продолговатого мозга, контрлатерального НОТ и из различных областей коры. [1] [17]
Выходы [ править ]
Многие претектальные ядра имеют общие мишени эфферентных проекций. Все претектальные ядра, за исключением НА, проецируются на ядра таламуса, субталамуса, верхних холмиков , ретикулярной формации , моста и нижней оливы . [10] И ON, и CPA имеют эфферентные проекции на ядро Эдингера-Вестфаля . NPP и NPA проецируются на пульвинар, латеральное заднее ядро таламуса и несколько прецеребеллярных ядер. [1]
НОТ имеет эфферентные проекции на неясную зону субталамуса, несколько ядер моста, продолговатого мозга, внутриламинарные ядра, средний мозг, а также дорсальные и вентральные ядра таламуса. Его двусторонние тормозные проекции на добавочную оптическую систему включают соединения с латеральными и медиальными терминальными ядрами. Проекции на субталамус направлены в сторону латерального коленчатого ядра и пульвинарной области . НОТ двусторонне проецируется к верхнему холмику , хотя ипсилатеральные соединения кажутся более доминирующими. В дополнение к этим проекциям НЕ проецируется на вестибулярные и вестибуло-мозжечковые релейные ядра. [1]
Функция [ править ]
Являясь частью подкорковой зрительной системы, нейроны претектальных ядер реагируют на различную интенсивность освещения и в первую очередь участвуют в реализации бессознательных поведенческих реакций на острые изменения света. В целом эти реакции включают в себя инициацию оптокинетических рефлексов, хотя претектум также может регулировать ноцицепцию и быстрый сон. [12]
Зрачковый световой рефлекс [ править ]
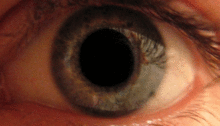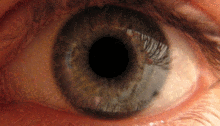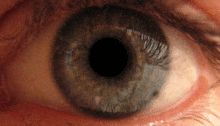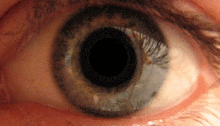
Зрачковый рефлекс на свет осуществляется претектумом. [2] Этот рефлекс отвечает за сужение зрачков при попадании света в глаз. Несколько претектальных ядер, в частности ON, получают информацию об освещенности с ипсилатеральной стороны сетчатки обоих глаз через зрительный тракт. Известно, что ядра в ON постепенно увеличивают активацию в ответ на повышение уровня освещенности. Затем эта информация передается непосредственно в ядро Эдингера-Вестфаля, которое передает команду на сужение зрачков зрачковому сфинктеру через цилиарный ганглий. [4] [18]
Плавное преследование [ править ]
Претектальные ядра, в частности НОТ, участвуют в координации движений глаз при плавном преследовании. Эти движения позволяют глазу внимательно следить за движущимся объектом и догонять объект после неожиданного изменения направления или скорости. Чувствительные к направлению нейроны скольжения сетчатки внутри НЕ передают в кору ипсиверсивную горизонтальную информацию об ошибках сетчатки через нижнюю оливу . В течение дня эта информация воспринимается и передается нейронами с большими рецептивными полями, тогда как парафовеальные нейроны с маленькими рецептивными полями делают это в темноте. Именно по этому пути НЕ может предоставлять информацию об ошибках сетчатки, чтобы направлять движения глаз. [1] [17] [19] Помимо своей роли в поддержании плавного преследования, претектум активируется во время оптокинетического нистагма, при котором глаз возвращается в центральное, обращенное вперед положение после того, как объект, за которым он следовал, выходит из поля зрения. [20]
Рефлекс аккомодации [ править ]
Часть претектума, особенно НОТ и НПП, участвуют в рефлексе аккомодации, с помощью которого глаз сохраняет фокус. [21] Проприоцептивная информация от сетчатки достигает претектума через глазодвигательный нерв и тройничный нерв. С этого момента механизм, с помощью которого глаз сохраняет фокус посредством мышечных сокращений сетчатки, аналогичен механизму зрачкового светового рефлекса. [4]
Антиноцицепция [ править ]
НПА участвует в активном уменьшении восприятия болевых раздражителей (антиноцицепции). [7] Хотя механизм, с помощью которого NPA изменяет реакцию организма на болевые раздражители, до конца не известен, исследования показали, что активность вентрального NPA запускает холинергические и серотонинергические нейроны. Эти нейроны активируют нисходящие пути, которые образуют синапсы в спинном мозге и ингибируют ноцицептивные клетки в задних рогах . [22] Помимо прямого антиноцицептивного механизма, НПА проецируется на области мозга, которые через связи с соматосенсорной корой регулируют восприятие болевых стимулов. Двумя из этих областей, на которые, как известно, проецируется NPA, являются неясная зона и заднее ядро таламуса. Области NPA могут быть специализированы для реагирования на различные типы боли. Исследования показали, что дорсальный NPA лучше всего снижает восприятие кратковременной боли, тогда как вентральный NPA снижает восприятие хронической боли. [23] Из-за своей роли в уменьшении хронической боли считается, что аномальная активность NPA связана с центральным болевым синдромом . [24]
Быстрый сон [ править ]
Множественные претектальные ядра могут участвовать в регуляции быстрого сна и поведения во время сна. Исследования показали, что претектум в сочетании с верхними бугорками могут быть ответственны за нециркадные изменения в поведении в фазе быстрого сна. [25] Было показано, что претектальные ядра, получающие сигналы сетчатки, в частности NOT и NPP, частично ответственны за инициацию быстрого сна у крыс-альбиносов. [5] Открытие проекций от претектума к нескольким ядрам таламуса, участвующим в активации коры во время быстрого сна, в частности проекции на суперхиазматическое ядро, которое является частью известного механизма регуляции быстрого сна, подтверждает эту гипотезу. [12]
См. также [ править ]
- Глазодвигательный нерв
- Средний мозг
- Список регионов человеческого мозга
- Средний мозг#Тектум
- тегментум
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с д и ж г Гамлин П.Д. (2006). «Претектум: связи и глазодвигательные роли». Нейроанатомия глазодвигательной системы . Прогресс в исследованиях мозга. Том. 151. стр. 379–405. дои : 10.1016/S0079-6123(05)51012-4 . ISBN 9780444516961 . ПМИД 16221595 .
- ^ Jump up to: Перейти обратно: а б Магун Х.В., Рэнсон С.В. (май 1935 г.). «Центральный путь светового рефлекса: исследование влияния поражений». Архив офтальмологии . 13 (5): 791–811. doi : 10.1001/archopht.1935.00840050069006 .
- ^ Нойхубер В., Шредль Ф. (ноябрь 2011 г.). «Вегетативный контроль глаза и радужной оболочки». Автономная нейронаука . 165 (1): 67–79. дои : 10.1016/j.autneu.2010.10.004 . ПМИД 21071284 . S2CID 35330212 .
- ^ Jump up to: Перейти обратно: а б с Донкелаар Х (2012). Клиническая нейроанатомия головного мозга и ее нарушения (1-е изд.). Берлин: Шпрингер. п. 343. ИСБН 978-3642191336 .
- ^ Jump up to: Перейти обратно: а б Миллер А.М., Миллер Р.Б., Обермайер В.Х., Бехан М., Бенка Р.М. (август 1999 г.). «Претектум обеспечивает быструю регуляцию сна при движении глаз с помощью света». Поведенческая нейронаука . 113 (4): 755–65. дои : 10.1037/0735-7044.113.4.755 . ПМИД 10495083 .
- ^ Босман Л.В., Хоувелинг А.Р., Оуэнс CB, Танке Н., Шевчук ОТ, Рахмати Н., Теуниссен WH, Джу К., Гонг В., Куккук СК, Де Зеув CI (1 января 2011 г.). «Анатомические пути, участвующие в создании и восприятии ритмических движений усов» . Границы интегративной нейронауки . 5 : 53. дои : 10.3389/fnint.2011.00053 . ПМК 3207327 . ПМИД 22065951 .
- ^ Jump up to: Перейти обратно: а б Рейс ГМ, Россанейс АС, Сильвейра Дж.В., Прадо В.А. (июнь 2012 г.). «μ1- и 5-НТ1-зависимые механизмы в переднем претектальном ядре опосредуют антиноцицептивные эффекты стимуляции ретросплениальной коры у крыс» . Науки о жизни . 90 (23–24): 950–5. дои : 10.1016/j.lfs.2012.04.023 . ПМИД 22575824 .
- ^ Миллодот М (2009). Словарь оптометрии и визуальных наук (7-е изд.). Эдинбург: Эльзевир/Баттерворт-Хайнеманн. ISBN 978-0-7020-2958-5 .
- ^ Рамачандран В.С. (2002). Энциклопедия человеческого мозга . Амстердам: Акад. Нажимать. ISBN 978-0122272103 .
- ^ Jump up to: Перейти обратно: а б с д и Ньювенхейс Р., тен Донкелаар Х.Дж., Николсон С. (1998). Центральная нервная система позвоночных . Берлин [ua]: Шпрингер. стр. 1812–1817. ISBN 978-3540560135 .
- ^ Боростянкой-Балдауф З., Герцег Л. (1 марта 2002 г.). «Парцелляция человеческого претектального комплекса: хемоархитектоническая переоценка». Нейронаука . 110 (3): 527–40. дои : 10.1016/S0306-4522(01)00462-6 . ПМИД 11906791 . S2CID 45807167 .
- ^ Jump up to: Перейти обратно: а б с Причард Дж.Р., Стоффель Р.Т., Куимби Д.Л., Обермайер В.Х., Бенка Р.М., Бехан М. (1 октября 2002 г.). «Иммунореактивность Fos в подкорковой зрительной оболочке крысы в ответ на изменение освещенности». Нейронаука . 114 (3): 781–93. дои : 10.1016/S0306-4522(02)00293-2 . ПМИД 12220578 . S2CID 32888470 .
- ^ Морен Л.П., Бланшар Дж.Х. (июль – август 1997 г.). «Нейропептид Y и иммунореактивность энкефалина в ретинореципиентных ядрах претектума и таламуса хомяка». Визуальная нейронаука . 14 (4): 765–77. дои : 10.1017/s0952523800012712 . ПМИД 9279004 . S2CID 25125769 .
- ^ Jump up to: Перейти обратно: а б Хатчинс Б., Вебер Дж.Т. (февраль 1985 г.). «Претектальный комплекс обезьяны: повторное исследование морфологии и окончаний сетчатки». Журнал сравнительной неврологии . 232 (4): 425–42. дои : 10.1002/cne.902320402 . ПМИД 3980762 . S2CID 25656241 .
- ^ Хатчинс Б. (1991). «Доказательства прямой проекции сетчатки на переднее претектальное ядро у кошки». Исследования мозга . 561 (1): 169–173. дои : 10.1016/0006-8993(91)90764-м . ПМИД 1797344 . S2CID 2584102 .
- ^ Вебер Дж.Т., Хатчинс Б. (1982). «Демонстрация проекции сетчатки на медиальное претектальное ядро у домашней кошки и беличьей обезьяны: анауторадиографический анализ». Исследования мозга . 232 (1): 181–186. дои : 10.1016/0006-8993(82)90622-9 . ПМИД 6173098 . S2CID 8118675 .
- ^ Jump up to: Перейти обратно: а б Оно С., Мустари MJ (май 2010 г.). «Сигналы зрительных ошибок из претектального ядра зрительного тракта направляют обучение моторике для плавного преследования» . Журнал нейрофизиологии . 103 (5): 2889–99. дои : 10.1152/Jn.01024.2009 . ПМЦ 2867559 . ПМИД 20457849 .
- ^ Гамлин П.Д., Чжан Х., Кларк Р.Дж. (1995). «Светящиеся нейроны в претектальном оливковом ядре опосредуют зрачковый световой рефлекс у макаки-резус». Экспериментальное исследование мозга . 106 (1): 169–76. дои : 10.1007/bf00241367 . ПМИД 8542972 . S2CID 24936336 .
- ^ Коллевейн Х. (январь 1975 г.). «Глазомоторные зоны среднего мозга и претектума кроликов». Журнал нейробиологии . 6 (1): 3–22. дои : 10.1002/neu.480060106 . ПМИД 1185174 .
- ^ Дитрих М., Шлиндвайн П., Януш Б., Бауэрманн Т., Стотер П., Бенсе С. (1 декабря 2007 г.). «Ствол мозга и активация мозжечка во время оптокинетической стимуляции». Клиническая нейрофизиология . 118 (12): 2811–2812. дои : 10.1016/j.clinph.2007.09.019 . S2CID 53198768 .
- ^ Конно С., Оцука К. (январь – февраль 1997 г.). «Области аккомодации и сужения зрачков в среднем мозге кошки». Японский журнал офтальмологии . 41 (1): 43–8. дои : 10.1016/s0021-5155(96)00010-x . ПМИД 9147188 .
- ^ Вильярреал, Дель Бель Е.А., Прадо, Вашингтон (май 2003 г.). «Участие переднего претектального ядра в контроле постоянной боли: исследование поведения и экспрессии c-Fos у крыс». Боль . 103 (1–2): 163–74. дои : 10.1016/S0304-3959(02)00449-9 . ПМИД 12749971 . S2CID 22753046 .
- ^ Вильярреал, Кина В.А., Прадо В.А. (сентябрь 2004 г.). «Антиноцицепция, вызванная стимуляцией переднего претектального ядра в двух моделях боли у крыс». Клиническая и экспериментальная фармакология и физиология . 31 (9): 608–13. дои : 10.1111/j.1440-1681.2004.04057.x . ПМИД 15479168 . S2CID 30378909 .
- ^ Мюррей П.Д., Масри Р., Келлер А. (июнь 2010 г.). «Аномальная активность переднего претектального ядра способствует развитию центрального болевого синдрома» . Журнал нейрофизиологии . 103 (6): 3044–53. дои : 10.1152/jn.01070.2009 . ПМЦ 2888237 . ПМИД 20357063 .
- ^ Миллер А.М., Обермайер В.Х., Бехан М., Бенка Р.М. (июль 1998 г.). «Верхние холмики-претектумы опосредуют прямое влияние света на сон» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (15): 8957–62. Бибкод : 1998PNAS...95.8957M . дои : 10.1073/pnas.95.15.8957 . ПМК 21184 . ПМИД 9671786 .