Миелиногенез
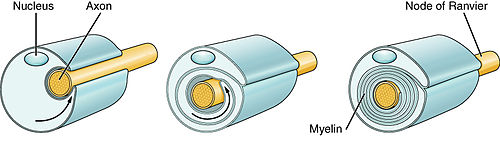
Миелиногенез — это образование и развитие миелиновых оболочек в нервной системе , которое обычно начинается на поздних стадиях пренатального развития нервной системы и продолжается на протяжении всего постнатального развития. [1] Миелиногенез продолжается на протяжении всей жизни, поддерживая обучение и память посредством пластичности нейронных цепей , а также ремиелинизации после травмы. [2] Успешная миелинизация аксонов увеличивает скорость потенциала действия , обеспечивая скачкообразную проводимость , которая необходима для своевременной передачи сигнала между пространственно разделенными областями мозга, а также обеспечивает метаболическую поддержку нейронов. [3]
Этапы
[ редактировать ]Миелин образован олигодендроцитами центральной нервной системы и шванновскими клетками периферической нервной системы . Таким образом, первую стадию миелиногенеза часто определяют как дифференцировку ( клеток-предшественников олигодендроцитов OPC) или предшественников шванновских клеток в их зрелые аналоги. [4] с последующим образованием миелина вокруг аксонов. [5]
Линию олигодендроцитов можно разделить на четыре стадии в зависимости от их связи с началом миелинизации: [6]
- Дифференциация: OPC выходят из своего пролиферативного, самообновляющегося состояния и начинают экспрессировать гены и белки, связанные с детерминацией судьбы олигодендроцитов.
- Преолигодендроциты: эти клетки экспрессируют антиген O4 и развивают множественные отростки, которые распространяются радиально без какой-либо конкретной организации. [7]
- Незрелые олигодендроциты: эти клетки иногда называют премиелинизирующими олигодендроцитами. Эти клетки расширяют «первопроходческие процессы», которые контактируют с аксонами и прикрепляют премиелинизирующие олигодендроциты к нейронам, так что они готовы начать миелиногенез в ответ на аксональные сигналы. Эти пионерские отростки растут в продольном направлении вдоль аксонов-мишеней. [7]
- Зрелые олигодендроциты: после миелиногенеза зрелые олигодендроциты окружают аксоны организованными многослойными миелиновыми оболочками, которые содержат основной белок миелина (MBP) и протеолипидный белок миелина (PLP).
Таким образом, миелиногенез включает в себя процесс перехода между фазами 3 и 4. [6] После инициации миелиногенеза каждый пионерский процесс образует пластинчатые отростки, которые распространяются и развиваются по окружности вокруг аксона-мишени. Это формирует первый виток миелиновой оболочки. [7] Оболочка продолжает расширяться по длине аксона-мишени, в то время как новая мембрана синтезируется на переднем крае внутреннего язычка развивающейся миелиновой оболочки, которая начинает приобретать спиральную структуру поперечного сечения.

Чтобы обеспечить правильную сборку мембранных слоев, PLP вводится в мембрану для стабилизации взаимодействия между внешними листками миелиновых мембран; MBP локально транслируется и вставляется в листки цитоплазматической мембраны для внутреннего укрепления миелиновых мембран. [8] Одновременно с образованием аксональных узлов Ранвье края миелиновой оболочки образуют паранодальные петли. [9]
Механизм
[ редактировать ]

Основной спираль-петля-спираль фактор транскрипции OLIG1 играет важную роль в процессе миелиногенеза олигодендроцитов, регулируя экспрессию генов, связанных с миелином. OLIG1 необходим для инициации миелинизации олигодендроцитами в головном мозге, но в некоторой степени необязателен в спинном мозге. [10]
Сигналы, полученные от аксонов, регулируют начало миелиногенеза. Исследователи изучали регенерацию аксонов ПНС в течение 28 недель, чтобы выяснить, стимулируют ли периферические аксоны олигодендроциты к началу миелинизации. Экспериментальная индукция миелинизации путем регенерации периферических аксонов продемонстрировала, что шванновские клетки и олигодендроциты имеют общий механизм стимуляции миелинизации. [11] Аналогичное исследование, направленное на предоставление доказательств нейрональной регуляции миелиногенеза, показало, что образование миелина происходит за счет шванновских клеток, которые контролируются неопределенным свойством соответствующего аксона. [11]
Недавние исследования на крысах показали, что апотрансферрин и гормон щитовидной железы действуют как по отдельности, так и синергически, способствуя миелиногенезу, поскольку апотрансферрин способствует экспрессии рецептора гормона щитовидной железы альфа . [12]
Периферический миелиногенез
[ редактировать ]
Периферический миелиногенез контролируется синтезом белков Р1, Р2 и Р0. [13] С помощью SDS-PAGE исследователи выявили отдельные полосы с размерами 27 000 дальтон (P1), 19 000 дальтон (P2) и 14 000 дальтон (P0). Исследования также показали, что P1 и P2 активны раньше P0, поскольку этот белок поступает из периферической нервной системы. [13] В процессе регенерации шванновские клетки повторно синтезируют белки, связанные со специфичными для миелина белками, когда присутствие аксонов восстанавливается. Синтез миелин-специфичных белков происходит только в шванновских клетках, связанных с аксонами. [13] Более того, мембранно-мембранные взаимодействия между аксонами могут потребоваться для стимулирования синтеза P1, P2 и P0.
Миелиногенез в зрительном нерве
[ редактировать ]Процесс и механистическую функцию миелиногенеза традиционно изучают с использованием ультраструктурных и биохимических методов в зрительных нервах крыс . Реализация этого метода исследования уже давно позволила экспериментально наблюдать миелиногенез в модельном нерве организма , полностью состоящем из немиелинизированных аксонов. Кроме того, использование зрительного нерва крысы помогло ранним исследователям миелиногенеза понять неправильное и атипичное течение миелиногенеза. [14]
Одно раннее исследование показало, что в развивающихся зрительных нервах крыс образование олигодендроцитов и последующая миелинизация происходят постнатально . В зрительном нерве клетки олигодендроцитов в последний раз разделились через пять дней, при этом начало образования миелина произошло примерно на 6 или 7 день или примерно на 6 или 7 день. Однако точный процесс, посредством которого олигодендроциты стимулировались к выработке миелина, еще не был полностью изучен. Понятно, но ранняя миелинизация зрительного нерва связана с увеличением выработки различных липидов – холестерина, цереброзида и сульфатида. [14]
Когда исследователи начали проводить постнатальные исследования, они обнаружили, что миелиногенез в зрительном нерве крысы первоначально начинается с аксонов самого большого диаметра, а затем переходит к оставшимся аксонам меньшего размера. На второй неделе после рождения образование олигодендроцитов замедлилось – к этому моменту 15% аксонов миелинизировались – однако миелиногенез продолжал быстро увеличиваться. В течение четвертой недели после рождения почти 85% аксонов зрительных нервов крысы были миелинизированы. [14] В течение пятой недели и далее к шестнадцатой неделе миелинизация замедлялась, и оставшиеся безмиелинизированные аксоны были покрыты миелином. [15] Ранние исследования зрительного нерва крысы внесли значительный вклад в знания в области миелиногенеза.
Роль сульфатидов
[ редактировать ]Исследования развивающегося зрительного нерва показали, что галактоцереброзид (который образует сульфатид) появляется на 9-й день после рождения и достигает пика на 15-й день после рождения. [14] Это выражение было похоже на период, когда зрительный нерв демонстрировал максимальный период миелинизации аксона. Поскольку активность миелинизации аксонов снижалась, можно было заключить, что активность фермента идет параллельно с включением сульфата ([35S]) в сульфатид in vivo.
Исследования зрительного нерва крыс показали, что через 15 дней после рождения наблюдается увеличение миелинизации. До этого периода времени большинство аксонов, примерно 70%, не миелинизированы. В это время [35S]сульфат включался в сульфатид и активность цереброзида , сульфотрансферазы достигала пика активности фермента. Согласно биохимическим данным, этот временной интервал также показал период максимальной миелинизации. [14]
В ЦНС сульфатиды , сульфатированные гликопротеины и сульфатированные мукополисахариды, по-видимому, связаны с нейронами, а не с миелином. При построении графика количества сульфатида, полученного из [35S] и активности сульфотрансферазы, мы получаем четко выраженные пики. [14] Пики приходятся на 15-й послеродовой день. Эти пики соответствовали максимальному периоду миелинизации зрительного нерва, наблюдавшемуся на протяжении всего эксперимента. [14]
В заключение следует отметить, что ранняя фаза миелинизации коррелирует с увеличением синтеза липидов, холестерина, цереброзида и сульфатида. [14] Вероятно, эти соединения синтезируются и упаковываются в аппарате Гольджи олигодендроглии. [14] Хотя транспорт этих липидов неизвестен, по-видимому, миелинизация задерживается без их синтеза.
Клиническое значение
[ редактировать ]Поскольку миелин образует электроизолирующий слой, окружающий аксоны некоторых нервных клеток , любое демиелинизирующее заболевание может повлиять на функционирование нервной системы . Одним из таких заболеваний является рассеянный склероз (РС), при котором демиелинизация происходит в центральной нервной системе (ЦНС). [16] Хотя проводятся исследования по защите олигодендроцитов и стимулированию ремиелинизации при рассеянном склерозе, [17] Современные методы лечения в основном направлены на роль иммунной системы в демиелинизации. [18]
История исследований
[ редактировать ]
Другой исследователь, Пол Флексиг, большую часть своей карьеры изучал и публиковал детали процесса в коре головного мозга человека. Это происходит в основном между двумя месяцами до и после рождения . Он идентифицировал 45 отдельных областей коры и фактически нанес на карту кору головного мозга по схеме миелинизации. Первая область коры, подвергающаяся миелинизированию, находится в моторной коре (часть поля Бродмана 4), вторая — обонятельная кора , а третья — часть соматосенсорной коры (BA 3,1,2).
Последними областями миелинизации являются передняя поясная извилина (F#43), нижняя височная кора (F#44) и дорсолатеральная префронтальная кора (F#45).
В извилинах головного мозга, как и во всех других отделах центральной нервной системы, нервные волокна развиваются не везде одновременно, а шаг за шагом в определенной последовательности, причем этот порядок событий особенно сохраняется в отношении появления мозговых вещество. В извилинах головного мозга облечение мозговым веществом (миелинизация) начинается в некоторых местах уже за три месяца до половозрелости плода, тогда как в других местах многочисленные волокна лишены мозгового вещества даже через три месяца после рождения. Порядок последовательности извилин регулируется законом, идентичным закону, который, как я показал, действует для спинного мозга , продолговатого мозга и среднего мозга и который можно сформулировать примерно так, а именно: одинаково важные нервные волокна развиваются одновременно, а волокна разной важности развиваются одно за другим в последовательности, определяемой императивным законом (основным законом миелогенеза). В некоторых извилинах образование мозгового вещества почти завершается, в то время как в некоторых оно еще даже не началось, а в других продвинулось лишь незначительно. [19]
Ссылки
[ редактировать ]- ^ Эйлам, Р.; Бар-Лев, Д.Д.; Левин-Зайдман, С.; Цури, М.; ЛоПрести, П.; Села, М.; Арнон, Р.; Ахарони, Р. (2014). «Олигодендрогенез и миелиногенез при постнатальном развитии, влияние глатирамера ацетата». Глия . 62 (4): 649–665. дои : 10.1002/glia.22632 . ПМИД 24481644 . S2CID 25559134 .
- ^ Уильямсон Дж. М., Лайонс Д. А. (2018). «Динамика миелина на протяжении всей жизни: постоянно меняющийся ландшафт?» . Переднеклеточные нейроны . 12 : 424. doi : 10.3389/fncel.2018.00424 . ПМК 6252314 . ПМИД 30510502 .
- ^ Филипс Т., Ротштейн Дж. Д. (2017). «Олигодендроглия: метаболические опоры нейронов» . Джей Клин Инвест . 127 (9): 3271–3280. дои : 10.1172/JCI90610 . ПМЦ 5669561 . ПМИД 28862639 .
- ^ Серрано-Регал, член парламента, Луэнгас-Эскуса I, Байон-Кордеро Л, Ибарра-Аиспуруа Н, Альберди Э, Перес-Самартин А; и др. (2020). «Дифференцировка олигодендроцитов и миелинизация усиливаются за счет активации рецептора GABAB» . Нейронаука . 439 : 163–180. doi : 10.1016/j.neuroscience.2019.07.014 . ПМИД 31349008 . S2CID 198934117 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Уоткинс, Т., Мулиньяве, С., Эмери, Б., Баррес, Б. (2008). Различные стадии миелинизации, регулируемые Y-секретазой и астроцитами в быстро миелинизирующейся системе совместной культуры ЦНС. 555-569
- ^ Jump up to: а б Кинни, ХК, и Вольпе, Джей-Джей (2018). События миелинизации. Неврология новорожденных Вольпе, 176–188. doi:10.1016/b978-0-323-42876-7.00008-9
- ^ Jump up to: а б с Фридрих В.Л., Харди Р.Дж. (1996). Прогрессивная ремоделация олигодендроцитарного отростка в процессе миелиногенеза. 243-54.
- ^ Барон В., Хоекстра Д. (2010). «О биогенезе миелиновых мембран: сортировка, транспортировка и полярность клеток» . ФЭБС Летт . 584 (9): 1760–70. дои : 10.1016/j.febslet.2009.10.085 . ПМИД 19896485 .
- ^ Снайдеро Н, Мёбиус В, Чопка Т, Хеккинг ЛХ, Матисен С, Верклей Д; и др. (2014). «Миелиновая мембрана обертывает аксоны ЦНС за счет PI (3,4,5) P3-зависимого поляризованного роста на внутреннем языке» . Клетка . 156 (1–2): 277–90. дои : 10.1016/j.cell.2013.11.044 . ПМЦ 4862569 . ПМИД 24439382 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Синь, М. (2005). Миелиногенез и распознавание аксонов олигодендроцитами головного мозга не связаны у мышей с Olig1-Null. Журнал неврологии, 25 (6), 1354–1365. doi:10.1523/jneurosci.3034-04.2005
- ^ Jump up to: а б Вайнберг Э. и Спенсер П. (1979). Исследования по контролю миелиногенеза. 3. Передача сигнала о миелинизации олигодендроцитов путем регенерации периферических аксонов. Исследования мозга, 162(2), 273-279. doi:10.1016/0006-8993(79)90289-0
- ^ Марциали, Л.Н., Гарсия, CI, Пасквини, JM (2015). Трансферрин и гормон щитовидной железы сходятся в контроле миелиногенеза. Экспериментальная неврология. Том 265. 129–141.
- ^ Jump up to: а б с Политис, М.Дж., Н. Штернбергер, Кэти Эдерле и Питер С. Спенсер. «Исследования по контролю миелиногенеза». Журнал неврологии 2.9 (1982): 1252–266.
- ^ Jump up to: а б с д и ж г час я Теннекун, Г.И., Коэн, С.Р., Прайс, Д.Л., МакХанн, Г.М. (1977). Миелиногенез в зрительном нерве. Морфологический, авторадиографический и биохимический анализ. Журнал клеточной биологии, 72 (3), 604–616.
- ^ Дангата, Ю., Кауфман, М. (1997). Миелиногенез в зрительном нерве гибридных мышей F1 (C57BL x CBA): морфометрический анализ. Европейский журнал морфологии, 35 (1), 3-18.
- ^ «Информационная страница NINDS по рассеянному склерозу» . Национальный институт неврологических расстройств и инсульта . 19 ноября 2015 года. Архивировано из оригинала 13 февраля 2016 года . Проверено 6 марта 2016 г.
- ^ Роджерс, Джейн М.; Робинсон, Эндрю П.; Миллер, Стивен Д. (2013). «Стратегии защиты олигодендроцитов и усиления ремиелинизации при рассеянном склерозе» . Медицина открытий . 86 (86): 53–63. ПМЦ 3970909 . ПМИД 23911232 .
- ^ Коэн Дж. А. (июль 2009 г.). «Новые методы лечения рецидивирующего рассеянного склероза». Арх. Нейрол . 66 (7): 821–8. дои : 10.1001/archneurol.2009.104 . ПМИД 19597083 .
- ^ Флексиг, Пауль (19 октября 1901 г.). «Эволюционная (миелогенетическая) локализация коры головного мозга у человека» . Ланцет . 158 (4077): 1028. дои : 10.1016/s0140-6736(01)01429-5 .