Глия лимитанс
| Глия лимитанс | |
|---|---|
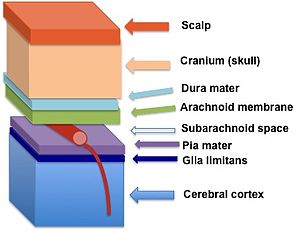 Ограниченная глия (темно-синего цвета) расположена между мягкой мозговой оболочкой и корой головного мозга. | |
| Подробности | |
| Части | Астроцит , базальная пластинка |
| Идентификаторы | |
| латинский | Глия лимитанс |
| НейроЛекс ID | nlx_subcell_100209 |
| Анатомические термины нейроанатомии | |
Glia limitans , или глиальная ограничивающая мембрана , представляет собой тонкий барьер отростков астроцитов ножек , связанный с паренхиматозной базальной пластинкой, окружающей головной и спинной мозг . Это самый внешний слой нервной ткани , и среди его обязанностей — предотвращение чрезмерной миграции нейронов и нейроглии , поддерживающих клеток нервной системы, в мозговые оболочки . Ограниченная глия также играет важную роль в регуляции движения небольших молекул и клеток в ткань головного мозга , работая совместно с другими компонентами центральной нервной системы (ЦНС), такими как гематоэнцефалический барьер (ГЭБ) . [1]
Расположение и структура
[ редактировать ]Периваскулярные ножки астроцитов образуют тесную связь с базальной пластинкой паренхимы головного мозга. [2] для создания лимитирующей глии. Эта мембрана лежит глубоко в мягкой мозговой оболочке и субпиальном пространстве и окружает периваскулярные пространства (пространства Вирхова-Робена). Любое вещество, попадающее в центральную нервную систему из крови или спинномозговой жидкости (СМЖ), должно пересечь пределы глии.
Две разные классификации глиальных ограничивающих мембран, периваскулярная и поверхностная глиальные ограничения, имеют почти идентичную структуру, однако их можно отличить друг от друга по расположению в мозге. Glia limitans периваскулярная примыкает к периваскулярному пространству, окружающему паренхиматозные кровеносные сосуды, и функционирует как вспомогательный компонент гематоэнцефалического барьера. Напротив, непаренхиматозные кровеносные сосуды, присутствующие в субарахноидальном пространстве, не покрыты ограничивающей глией. Вместо этого все субарахноидальное пространство закрыто от нервной ткани поверхностной глией. [3] Эти две части глии лимитанс непрерывны; однако общепринятое мнение гласит, что часть, которая покрывает поверхность мозга, называется поверхностной, а часть, которая окружает кровеносные сосуды внутри мозга, называется периваскулярной.
Функция
[ редактировать ]Физический барьер
[ редактировать ]
Основная роль лимитирующей глии заключается в том, чтобы действовать как физический барьер против нежелательных клеток или молекул, пытающихся проникнуть в ЦНС. Ограниченная глия разделяет мозг на отдельные отсеки, изолируя паренхиму от сосудистых и субарахноидальных отделов. [4] В головном мозге глиальная ограничивающая мембрана является важной составляющей гематоэнцефалического барьера. Эксперименты с использованием электронно-плотных маркеров показали, что функциональными компонентами гематоэнцефалического барьера являются эндотелиальные клетки , из которых состоит сам сосуд. Эти эндотелиальные клетки содержат очень непроницаемые плотные соединения , благодаря которым кровеносные сосуды головного мозга не имеют «протечек», присущих артериям и венам в других частях тела. [5] В ходе экспериментов как in vivo, так и in vitro было показано, что астроцитарные ножки ограничительной глии индуцируют образование плотных соединений эндотелиальных клеток во время развития мозга. [6] В эксперименте in vivo были собраны астроциты крысы, которые были помещены в переднюю камеру куриного глаза или на хориоаллантоис . Проницаемые кровеносные сосуды радужной оболочки или хориоаллантоиса становились непроницаемыми для синего альбумина после того, как они попадали в трансплантированный болюс астроцитов. В эксперименте in vitro эндотелиальные клетки сначала культивировали отдельно, и в репликах замораживания-излома плотные соединения были прерывистыми и пронизаны щелевыми соединениями . Затем эндотелиальные клетки головного мозга культивировали с астроцитами, что привело к усилению плотных контактов и снижению частоты щелевых соединений.
Ограниченная глия также действует как вторая линия защиты от всего, что проходит через гематоэнцефалический барьер. Однако, поскольку астроциты, окружающие сосуды, соединены щелевыми соединениями , они не считаются частью ГЭБ, и материал может легко проходить между отростками ножки.
Иммунологический барьер
[ редактировать ]Астроциты предельной глии ответственны за разделение мозга на два основных отдела. Первый отдел представляет собой иммунопривилегированную паренхиму головного и спинного мозга. Этот компартмент содержит множество иммуносупрессивных белков клеточной поверхности, таких как CD200 и CD95L, и обеспечивает высвобождение противовоспалительных факторов. Второй компартмент представляет собой субарахноидальное, субпиальное и периваскулярное пространство, не привилегированное иммунитетом. Эта область заполнена провоспалительными факторами, такими как антитела , белки комплемента , цитокины и хемокины . Считается, что астроциты предельной глии являются компонентом мозга, секретирующим про- и противовоспалительные факторы. [1]
Разработка
[ редактировать ]Развитие длинных клеточных отростков астроцитов, которые являются неотъемлемой частью структуры ограничивающей глии, связано с наличием менингеальных клеток в мягкой мозговой оболочке. [7] Менингеальные клетки представляют собой специализированные фибробластоподобные клетки, окружающие ЦНС и крупные кровеносные сосуды. Было обнаружено, что они взаимодействуют с астроцитами при начальном формировании лимитирующей глии во время развития и участвуют в ее дальнейшем поддержании на протяжении всей жизни. Было обнаружено, что искусственно вызванное разрушение менингеальных клеток во время развития ЦНС приводит к изменению субпиального внеклеточного матрикса и разрушению ограничивающей глии. [8]
Ограниченная глия также оказалась важной для восстановления ЦНС после травм. Когда повреждения возникают на поверхности мозга, менингеальные клетки делятся и мигрируют в очаг поражения, в конечном итоге выстилая всю полость повреждения. Если травма значительно снизила плотность астроцитов и создала пространство внутри ткани, менингеальные клетки будут инвазироваться еще более диффузно. Когда вторгающиеся менингеальные клетки вступают в контакт с астроцитами, они могут индуцировать образование новой функциональной ограниченной глии. Новые лимитанты глии, образующиеся после повреждения ЦНС, обычно представляют собой барьер для регенерации аксонов. [9]
Клиническая значимость
[ редактировать ]Существует ряд заболеваний, связанных с проблемами или аномалиями лимитирующей глии. Многие заболевания могут возникнуть в результате нарушения предельной глии, при котором она больше не сможет выполнять свою функциональную роль барьера. Ниже описаны два наиболее распространенных заболевания, возникающих в результате нарушения предельной глии.
Врожденная мышечная дистрофия типа Фукуямы (ВМД)
[ редактировать ]Нарушения в комплексе ограничивающей глии и базальной пластинки связаны с врожденной мышечной дистрофией Фукуямы (ВМД) , которая, как полагают, является результатом микрополигирий или небольших выпячиваний нервной ткани. [10] Хотя основной механизм формирования этих нарушений в значительной степени неизвестен, недавние исследования показали, что белок фукутин напрямую связан с развивающимися поражениями. Мутации белка фукутина приводят к снижению уровня его экспрессии в головном и спинном мозге новорожденных, что, в свою очередь, способствует ослаблению структурной целостности пограничной глии. Нейрональные и глиальные клетки мигрируют через ослабленный барьер, что приводит к скоплению нервной ткани в субарахноидальном пространстве. Предполагается, что эта аномальная миграция, известная как кортикальная дисплазия , является одной из основных причин ФКМД. [11]
Экспериментальный аутоиммунный энцефаломиелит (ЭАЭ)
[ редактировать ]Показано, что клинические признаки экспериментального аутоиммунного энцефаломиелита (ЭАЭ) проявляются только после проникновения воспалительных клеток через пределы глии и при входе в паренхиму ЦНС. Активность матриксных металлопротеиназ , в частности ММП-2 и ММП-9, необходима для проникновения воспалительных клеток в пределы глии. Скорее всего, это связано с биохимией базальной мембраны паренхимы и астроцитарными отростками ножки. ММП-2 и ММП-9 продуцируются миелоидными клетками , которые окружают Т-клетки в периваскулярном пространстве. Эти металлопротеиназы позволяют иммунным клеткам преодолевать пределы глии и достигать паренхимы ЦНС, чтобы атаковать паренхиматозные клетки ЦНС. Как только иммунные клетки достигают паренхимы ЦНС и начинается иммунная атака, паренхиматозные клетки ЦНС приносят в жертву для борьбы с инфекцией. Аутоиммунный ответ на ЭАЭ приводит к хронической атаке олигодендроцитов и нейронов, что способствует демиелинизация и потеря аксонов. В конечном итоге это может привести к потере нейронов ЦНС. [3]
Сравнительная анатомия
[ редактировать ]Поскольку предельная глия выполняет такую важную структурную и физиологическую функцию у человека, неудивительно, что эволюционные предшественники ограничивающей глии мембраны могут быть обнаружены у многих других животных.
насекомых У открытая система кровообращения нет кровеносных сосудов , поэтому внутри их ганглиев . Однако у них есть оболочка из периневральных глиальных клеток , которая окутывает нервную систему и имеет такие же плотные перекрывающие соединения, которые индуцируются ограничивающей глией у людей. Эти клетки действуют как барьер и отвечают за создание градиентов проницаемости.
У некоторых моллюсков наблюдается глиально-интерстициальный жидкий барьер без наличия плотных контактов. Головоногие моллюски, в частности, имеют церебральные ганглии, обладающие микроциркуляцией , часто встречающейся в составе высших организмов. Часто глиальные клетки образуют бесшовную оболочку, полностью окружающую кровеносное пространство. Барьер состоит из зональных межклеточных соединений , а не плотных соединений, с щелями, образованными внеклеточными фибриллами . Считается, что помимо защиты от крови эти барьеры обеспечивают локальный контроль микроокружения вокруг определенных групп нейронов — функцию, необходимую для сложных нервных систем. [6]
Было обнаружено, что у обезьян и других приматов глиальная ограничивающая мембрана очень похожа на человеческую. Исследования на этих животных показали, что толщина лимитирующей глии сильно различается не только у разных видов, но и в разных областях центральной нервной системы одного и того же организма. Дальнейшие наблюдения за молодыми и старыми обезьянами доказали, что у более молодых особей мембраны тоньше с меньшим количеством слоев астроцитарных отростков, тогда как у старших обезьян мембраны гораздо толще. [12]
Текущие исследования
[ редактировать ]По состоянию на 2011 год исследования сосредоточены на двусторонней связи между нейронами и глиальными клетками. Связь между этими двумя типами клеток обеспечивает аксональную проводимость, синаптическую передачу, а также обработку информации для регулирования и лучшего контроля процессов центральной нервной системы. Различные формы коммуникации включают нейротрансмиссию , потоки ионов и сигнальные молекулы . Совсем недавно, в 2002 году, новая информация о процессе взаимодействия нейронов и глии была опубликована Р. Дугласом Филдсом и Бет Стивенс-Грэм. Они использовали передовые методы визуализации, чтобы объяснить, что ионные каналы, наблюдаемые в глиальных клетках, не вносят вклад в потенциалы действия , а скорее позволяют глии определять уровень активности нейронов в непосредственной близости. Было установлено, что глиальные клетки взаимодействуют друг с другом исключительно с помощью химических сигналов и даже имеют специализированные глиально-глиальные и нейрон-глиальные сигнальные системы нейротрансмиттеров. Кроме того, было обнаружено, что нейроны выделяют химические посланники во внесинаптических областях, что позволяет предположить, что нейронно-глиальные отношения включают в себя функции, выходящие за рамки синаптической передачи. Известно, что глия помогает в формирование синапсов , регулирование силы синапсов и обработка информации, как упоминалось выше. Процесс высвобождения аденозинтрифосфата (АТФ), глутамата и других химических веществ-посредников из глии обсуждается и рассматривается как направление будущих исследований. [13]
Ссылки
[ редактировать ]- ^ Jump up to: а б Хельмут Кеттенманн; Брюс Р. Рэнсом (2005). Нейроглия . Издательство Оксфордского университета, США. стр. 303–305. ISBN 978-0-19-515222-7 . Проверено 20 марта 2011 г.
- ^ Саладин, Кеннет С. (2011). Анатомия человека . МакГроу-Хилл. п. 358. ИСБН 9780071222075 .
- ^ Jump up to: а б Энгельхардт Б., Куан С. (2011). «Жидкости и барьеры ЦНС устанавливают иммунные привилегии, ограничивая иммунный надзор рвом с двумя стенами, окружающим замок ЦНС» . Жидкостные барьеры ЦНС . 8 (1): 4. дои : 10.1186/2045-8118-8-4 . ПМК 3039833 . ПМИД 21349152 .
- ^ Алексей Несторович Верхрацкий; Артур Батт (2007). Глиальная нейробиология: учебник . Джон Уайли и сыновья. п. 24. ISBN 978-0-470-01564-3 . Проверено 20 марта 2011 г.
- ^ Алан Питерс; Сэнфорд Л. Палай; Генри деФ. Вебстер (1991). Тонкое строение нервной системы: нейроны и поддерживающие их клетки . Издательство Оксфордского университета. стр. 292–293. ISBN 978-0-19-506571-8 . Проверено 25 марта 2011 г.
- ^ Jump up to: а б Брайтман, Милтон (1991). «Влияние астроглии на гематоэнцефалический барьер». В Эбботе, штат Нью-Джерси (ред.). Глиально-нейрональное взаимодействие . Нью-Йоркская академия наук. п. 633. ИСБН 0-89766-680-1 .
- ^ Штрукхофф, Гернот (1995). «Совместные культуры менингеальных и астроцитарных клеток - способ формирования глиально-ограничивающей мембраны». Межд. Дж. Девл Нейронауки . 13 (6): 595–606. дои : 10.1016/0736-5748(95)00040-N . ПМИД 8553894 . S2CID 29140815 .
- ^ Б. Кастеллано Лопес; Бернар Кастеллано; Мануэль Грансон-Сампедро (15 сентября 2003 г.). Функция глиальных клеток . Профессиональное издательство Персидского залива. п. 18. ISBN 978-0-444-51486-8 . Проверено 25 марта 2011 г.
- ^ Матиас Бэр (2006). Ремонт мозга . Профессиональное издательство Персидского залива. п. 19. ISBN 978-0-306-47859-8 . Проверено 25 марта 2011 г.
- ^ Сайто Ю., Мураяма С., Каваи М., Накано И. (октябрь 1999 г.). «Нарушение комплекса церебральной глии-лимитанс-базальной пластинки при врожденной мышечной дистрофии типа Фукуямы». Акта Нейропатол . 98 (4): 330–6. дои : 10.1007/s004010051089 . ПМИД 10502035 . S2CID 35614560 .
- ^ Накано, Имахару; Фунахаси, М; Такада, К; Тода, Т (1996). «Являются ли нарушения в ограниченной глии основной причиной микрополигирии при врожденной мышечной дистрофии Фукуямы (ВМД)? - Патологическое исследование коры головного мозга плода с ВМД». Акта Нейропатологика . 91 (3): 313–321. дои : 10.1007/s004010050431 . ПМИД 8834545 . S2CID 967866 .
- ^ Эннио Паннесе (1994). Нейроцитология: тонкая структура нейронов, нервных отростков и нейроглиальных клеток . Тиме. стр. 173–175. ISBN 978-0-86577-456-8 . Проверено 25 марта 2011 г.
- ^ Филдс, Дуглас; Стивенс-Грэм, Б. (2002). «Новый взгляд на связь нейронов и глии» . Наука . 298 (5593): 556–562. Бибкод : 2002Sci...298..556F . дои : 10.1126/science.298.5593.556 . ПМЦ 1226318 . ПМИД 12386325 .