Улучшенный оливарный комплекс
| Улучшенный оливарный комплекс | |
|---|---|
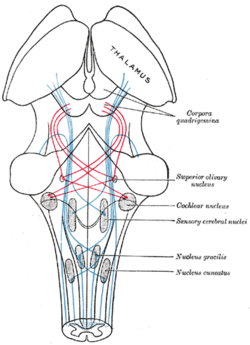 Схема, показывающая ход волокон лемниска ; медиальная петля синего цвета, латеральная - красного. (Верхнее оливковое ядро отмечено в центре справа.) | |
| Подробности | |
| Идентификаторы | |
| латинский | Верхнее оливарисное ядро |
| МеШ | D065832 |
| Нейроимена | 569 |
| НейроЛекс ID | бирнлекс_1307 |
| ТА98 | A14.1.05.415 |
| ТА2 | 5937 |
| ФМА | 72247 |
| Анатомические термины нейроанатомии | |
Верхний оливковый комплекс ( SOC ) или верхняя олива представляет собой совокупность ядер ствола мозга, которая расположена в мосту , функционирует во многих аспектах слуха и является важным компонентом восходящих и нисходящих слуховых путей слуховой системы . SOC тесно связан с телом трапеции : большинство групп клеток SOC расположены дорсально (сзади у приматов) по отношению к этому пучку аксонов, в то время как ряд групп клеток встроен в тело трапеции. В целом SOC демонстрирует значительную межвидовую изменчивость: самый большой у летучих мышей и грызунов и меньший у приматов.
Физиология
[ редактировать ]Верхнее оливковое ядро играет ряд ролей в слухе. Медиальная верхняя олива (MSO) представляет собой специализированное ядро, которое, как полагают, измеряет разницу во времени прибытия звуков между ушами ( межушную разницу во времени или ITD). ИТД является основным признаком определения азимута звуков, т. е. локализации их в азимутальной плоскости – их градуса влево или вправо.
Считается, что верхняя латеральная олива (LSO) участвует в измерении разницы в интенсивности звука между ушами ( межушная разница уровней или ILD). ILD является вторым важным сигналом при определении азимута высокочастотных звуков.
Связь со слуховой системой
[ редактировать ]Верхний оливковый комплекс обычно расположен в мосту , но у человека простирается от рострального продолговатого мозга до середины моста. [1] и получает проекции преимущественно от передне-вентрального ядра улитки (AVCN) через трапециевидное тело, хотя задне-вентральное ядро проецируется в SOC через промежуточную акустическую полоску. SOC — это первое крупное место схождения слуховой информации из левого и правого уха. [2]
Первичные ядра
[ редактировать ]Верхний оливарный комплекс разделен на три первичных ядра: MSO, LSO и медиальное ядро трапециевидного тела, а также несколько более мелких периоливарных ядер. [3] Эти три ядра наиболее изучены и, следовательно, лучше всего понятны. Обычно их рассматривают как образующие восходящий путь азимутальной локализации.
Медиальная верхняя олива (MSO)
[ редактировать ]Считается, что медиальная верхняя олива помогает определить азимут звука, то есть угол влево или вправо, где расположен источник звука. Звуковые сигналы возвышения не обрабатываются в оливарном комплексе. Веретенообразные клетки дорсального ядра улитки (DCN), которые, как полагают, способствуют локализации при возвышении, обходят SOC и проецируются непосредственно на нижние холмики . Присутствуют только горизонтальные данные, но они поступают из двух разных ушных источников, что помогает локализовать звук по азимутальной оси. [4] Способ, которым верхняя олива делает это, заключается в измерении разницы во времени между двумя ушными сигналами, записывающими один и тот же стимул. Путешествие вокруг головы занимает около 700 мкс, а медиальная верхняя олива способна различать гораздо меньшие разницы во времени. Фактически замечено, что люди могут обнаруживать межушные различия длительностью до 10 мкс. [4] Ядро тонотопически организовано, но азимутальная проекция рецептивного поля представляет собой «скорее всего, сложную нелинейную карту». [5]
Проекции медиальной верхней оливы плотно оканчиваются в ипсилатеральном центральном ядре нижнего холмика (CNIC). Считается, что большинство этих аксонов имеют «круглую форму» или тип R. Эти аксоны R в основном являются глутаматергическими, содержат круглые синаптические пузырьки и образуют асимметричные синаптические соединения. [2]
- Это самое большое из ядер, у человека оно содержит около 15 500 нейронов. [1]
- Каждый MSO получает двусторонние входные данные от правого и левого AVCN.
- Выход осуществляется через ипсилатеральную латеральную петлю к нижнему холмику . [6]
- MSO лучше реагирует на бинауральные стимулы.
- Основная функция MSO - обнаружение сигналов межушной разницы во времени (ITD), указывающих на бинауральную латерализацию.
- В мозге аутиста серьезно нарушена MSO. [7]
Боковая верхняя олива (LSO)
[ редактировать ]Эта олива имеет те же функции, что и медиальная верхняя олива, но использует интенсивность для локализации источника звука. [8] LSO получает возбуждающий глутаматергический вход от сферических кустистых клеток ипсилатерального ядра улитки и тормозной глицинергический вход от медиального ядра трапециевидного тела (MNTB). MNTB приводится в действие возбуждающим сигналом от глобулярных кустистых клеток контралатерального ядра улитки. Таким образом, LSO получает возбуждающий сигнал от ипсилатерального уха и тормозящий сигнал от контралатерального уха. Это основа чувствительности к ИЗЛ. Проекции от обоих ядер улитки преимущественно высокочастотные, и эти частоты впоследствии представлены большинством нейронов LSO (>2/3 на частоте более 2–3 кГц у кошки). LSO на самом деле кодирует частоту во всем слышимом животным диапазоне (а не только «высокую» частоту). Дополнительные входные сигналы поступают от ипсилатерального LNTB (глицинэргического, см. ниже), который передает тормозную информацию от ипсилатерального ядра улитки. [9] Другой возможный ингибирующий вклад происходит от ипсилатеральных несферических клеток AVCN. Эти клетки либо шаровидные, кустистые, либо мультиполярные (звездчатые). Любой из этих двух входных сигналов может обеспечить основу для ипсилатерального торможения, наблюдаемого на картах ответов, окружающих первичное возбуждение, что повышает точность настройки частоты устройства. [10] [11]
LSO билатерально проецируется в центральное ядро нижнего холмика (ICC). Ипсилатеральные проекции являются преимущественно тормозными (глицинэргическими), а контралатеральные – возбуждающими. Дополнительные цели проекции включают дорсальное и вентральное ядра латеральной петли (DNLL и VNLL) . ГАМКергические проекции DNLL образуют основной источник ГАМК в слуховом стволе мозга и двусторонне проецируются на ICC и на контралатеральный DNLL. Эти сходящиеся возбуждающие и тормозящие связи могут снижать зависимость чувствительности ILD от уровня в ICC по сравнению с LSO.
Дополнительные выступы образуют латеральный оливокохлеарный пучок (LOC), который иннервирует внутренние волосковые клетки улитки. Считается, что эти проекции имеют большую постоянную времени и нормализуют уровень звука, воспринимаемый каждым ухом, чтобы помочь в локализации звука . [12] Существуют значительные видовые различия: проекционные нейроны LOC распределены внутри LSO у грызунов и окружают LSO у хищников (например, кошек).
Медиальное ядро трапециевидного тела (МНТБ)
[ редактировать ]- MNTB в трапециевидном теле состоит в основном из нейронов с круглыми клеточными телами, которые используют глицин в качестве нейромедиатора.
- Размер MNTB у приматов уменьшен. [13] [14] [15]
- Каждый нейрон MNTB имеет большое окончание типа «чашечки», чашечку Хелда, возникающую из шаровидных кустистых клеток в контралатеральном AVCN.
- Обнаружено два типа ответа: «тип измельчителя», подобный веретенообразным клеткам в AVCN, и первичный тип, который аналогичен ответам кустистых клеток в AVCN.
Периоливарные ядра
[ редактировать ]SOC состоит из шести-девяти периоливарных ядер, в зависимости от цитируемого исследователя, которые обычно называются в зависимости от их расположения относительно первичных ядер. Эти ядра окружают каждое из первичных ядер и вносят вклад как в восходящую, так и в нисходящую слуховую систему. Эти ядра также образуют источник оливокохлеарного пучка, иннервирующего улитку. [16] У морской свинки восходящие проекции на нижние холмики преимущественно ипсилатеральные (>80%), при этом наибольший одиночный источник исходит из SPON. Кроме того, вентральные ядра (RPO, VMPO, AVPO и VNTB) почти полностью ипсилатеральны, тогда как остальные ядра выступают билатерально. [17]
| Имя | Кот | Морская свинка | Крыса | Мышь |
|---|---|---|---|---|
| ЛСО | Х | Х | Х | Х |
| МСО | Х | Х | Х | Х* |
| МНТБ | Х | Х | Х | Х |
| ЛНТБ | Х | Х | "ЛВПО" | Х |
| АЛПО | Х | Х | ||
| ПВПО | Х | Х | ||
| ППО | Х | Х | "ЦПО" | |
| ВЛПО | Х | |||
| ДПО | Х | Х | Х | |
| ДЛПО | Х | Х | ||
| ВТБ | Х | Х | «МВПО» | Х |
| АВПО | Х | |||
| ВМПО | Х | Х | ||
| РПО | Х | Х | ||
| СПН | "ДМПО" | Х | Х | Х |
, [17] * У мышей MSO кажется меньшим и дезорганизованным. [18]
Вентральное ядро трапециевидного тела (ВНТБ)
[ редактировать ]- VNTB представляет собой небольшое ядро, расположенное латерально от MNTB и вентральнее MSO. [19]
- Это ядро, состоящее из гетерогенной популяции клеток, проецируется на многие слуховые ядра и образует медиальный оливокохлеарный пучок (МОС), который иннервирует наружные волосковые клетки улитки. [20] Эти клетки содержат электродвижущие волокна и действуют как механические усилители/аттенюаторы внутри улитки.
- Ядро проецируется на оба IC, при этом ни одна клетка не проецируется с двух сторон. [21]
Латеральное ядро трапециевидного тела (ЛНТБ)
[ редактировать ]- Расположен вентральнее LSO. [19]
- Сферические кустистые клетки AVCN проецируют коллатерали билатерально, а глобулярные кустистые клетки проецируют коллатерали ипсилатерально по отношению к нейронам LNTB. [22]
- Клетки иммунореактивны к глицину, [23] и помечены ретроградно после инъекции тритированного глицина в LSO. [9]
- Ядро проецируется на оба IC, при этом несколько клеток проецируются двусторонне, [21] а также ипсилатеральный LSO. [9]
- Большие мультиполярные клетки проецируются в ядро улитки, но не в IC, как у кошек, так и у морских свинок. [21] [24]
- Входы часто подаются через концевые лампочки Held , что обеспечивает очень быструю передачу сигнала.
Верхнее периоливарное ядро (SPON) (дорсомедиальное периоливарное ядро (ДМПО))
[ редактировать ]- Расположен непосредственно дорсально к MNTB. [19]
- У крыс СПОН представляет собой гомогенное ГАМКергическое ядро. Эти тонотопически организованные нейроны получают возбуждающие сигналы от осьминогов и мультиполярных клеток контралатерального вентрального ядра улитки . [25] глицинергический (ингибирующий) вход от ипсилатерального MNTB, неизвестный ГАМКергический (ингибирующий) вход и проекция на ипсилатеральный ICC. [26] Большинство нейронов реагируют только на смещение стимула, могут синхронизироваться по фазе с AM-стимулами частотой до 200 Гц и могут составлять основу селективности длительности ICC. [27] Примечательно, что нейроны SPON не получают нисходящие входные сигналы от IC и не проецируются в улитку или улитковое ядро, как это делают многие периоливарные ядра. [28]
- Напротив, глицинергические проекции на ипсилатеральную ICC наблюдаются у морских свинок и шиншилл, что указывает на видовые различия в нейротрансмиттерах. [29]
- У морских свинок мультиполярные клетки от круглой до овальной формы проецируются на оба IC, при этом многие клетки проецируются билатерально. Более удлиненные клетки, которые проецируются в ядро улитки, не проецируются в ICC. По-видимому, существуют популяции клеток, одна из которых проецируется ипсилатерально, а другая - двусторонне. [21]
- Большая часть информации была получена от SPON грызунов из-за значительного размера ядра у этих видов, и очень мало исследований было проведено на кошачьем ДМПО. [30] ни один из которых не был обширным.
Дорсальное периоливарное ядро (ДПО)
[ редактировать ]- Расположен дорсально и медиально от LSO. [19]
- Содержит блоки EE (возбуждаются обоими ушами) и E0 (возбуждаются только контралатеральным ухом). [31]
- Нейроны тонотопически организованы и обладают высокой частотой.
- Может принадлежать одному ядру вместе с ДЛПО. [32]
- Ядро проецируется на оба IC, при этом ни одна клетка не проецируется с двух сторон. [21]
Дорсолатеральное периоливарное ядро (DLPO)
[ редактировать ]- Расположен дорсально и латерально от LSO. [19]
- Содержит блоки EE (возбуждаются обоими ушами) и E0 (возбуждаются только контралатеральным ухом).
- Нейроны тонотопически организованы и имеют низкую частоту.
- Может принадлежать одному ядру вместе с ДПО.
- Ядро проецируется на оба IC, при этом несколько клеток проецируются двусторонне. [21]
Вентролатеральное периоливарное ядро (ВЛПО)
[ редактировать ]- Расположен вентрально и внутри вентрального холма LSO. [19]
- Содержит единицы EI (возбуждается контралатеральным ухом и подавляется ипсилатеральным ухом) и E0 (возбуждается только контралатеральным ухом).
- Нейроны тонотопически организованы и обладают высокой частотой.
- Подразделяется на LNTB, PPO и ALPO. [33]
Переднелатеральное периоливарное ядро (ALPO)
[ редактировать ]- Ядро проецируется на оба IC, при этом ни одна клетка не проецируется с двух сторон. [21]
- Большие мультиполярные клетки проецируются в ядро улитки, но не в IC, как у кошек, так и у морских свинок. [21] [24]
Вентромедиальное периоливарное ядро (ВМПО)
[ редактировать ]- Расположен между МСО и МНТБ. [19]
- Отправляет прогнозы в ICC на двусторонней основе. [21]
- Ядро проецируется на оба IC, при этом ни одна клетка не проецируется с двух сторон. [21]
Ростральное периоливарное ядро (РПО) (переднее периоливарное ядро (АРО))
[ редактировать ]- Расположен между ростральным полюсом MSO и VNLL. [19]
- Иногда его называют вентральным ядром трапециевидного тела (VNTB). [19]
Каудальное периоливарное ядро (CPO) (заднее периоливарное ядро (PPO))
[ редактировать ]- Расположен между каудальным полюсом МСО и лицевым ядром (7N). [19]
Постеровентральное периоливарное ядро (ПВПО)
[ редактировать ]- Ядро проецируется на оба IC, при этом ни одна клетка не проецируется с двух сторон. [21]
См. также
[ редактировать ]Ссылки
[ редактировать ]![]() Эта статья включает общедоступный текст со страницы 787 20-го издания «Анатомии Грея» (1918 г.).
Эта статья включает общедоступный текст со страницы 787 20-го издания «Анатомии Грея» (1918 г.).
- ^ Перейти обратно: а б Кулеша Р.Дж. (март 2007 г.). «Цитоархитектура верхнего оливарного комплекса человека: медиальная и латеральная верхняя олива». Исследование слуха . 225 (1–2): 80–90. дои : 10.1016/j.heares.2006.12.006 . ПМИД 17250984 . S2CID 19696622 .
- ^ Перейти обратно: а б Оливер Д.Л., Бекиус Г.Е., Шнейдерман А. (сентябрь 1995 г.). «Аксональные проекции от латеральной и медиальной верхней оливы до нижнего холмика кошки: исследование с использованием электронно-микроскопической авторадиографии». Журнал сравнительной неврологии . 360 (1): 17–32. дои : 10.1002/cne.903600103 . ПМИД 7499562 . S2CID 22997698 .
- ^ Кахаль, SRY и Л. Азуле (1909). Гистология нервной системы человека и позвоночных животных. Париж, Малоин.
- ^ Перейти обратно: а б Кандель Э.Р., Шварц Дж.Х., Джесселл Т.М. (2000). Принципы нейронауки . Нью-Йорк: МакГроу-Хилл. стр. 591–624 . ISBN 978-0-8385-7701-1 . OCLC 249318861 .
- ^ Оливер Д.Л., Бекиус Г.Е., Бишоп Д.К., Лофтус В.К., Батра Р. (август 2003 г.). «Топография кодирования межушного височного неравенства в проекциях медиальной верхней оливы на нижний холмик» . Журнал неврологии . 23 (19): 7438–7449. doi : 10.1523/JNEUROSCI.23-19-07438.2003 . ПМК 6740450 . ПМИД 12917380 .
- ^ Ринкон, Гектор; Гомес-Мартинес, Марио; Гомес-Альварес, Марсело; Салданья, Энрике (август 2024 г.). «Медиальная верхняя олива у крысы: анатомия, источники поступления и аксональные проекции» . Исследование слуха . 449 : 109036. doi : 10.1016/j.heares.2024.109036 . ISSN 0378-5955 . ПМИД 38797037 .
- ^ Кулеша Р.Дж., Лукозе Р., Стивенс Л.В. (январь 2011 г.). «Порок развития верхней оливы человека при расстройствах аутистического спектра». Исследования мозга . 1367 : 360–371. дои : 10.1016/j.brainres.2010.10.015 . ПМИД 20946889 . S2CID 39753895 .
- ^ Цучитани С., Будро Дж.К. (октябрь 1967 г.). «Кодирование частоты и интенсивности стимула клетками S-сегмента верхней оливы кошки». Журнал Акустического общества Америки . 42 (4): 794–805. дои : 10.1121/1.1910651 . ПМИД 6075565 .
- ^ Перейти обратно: а б с Гленденнинг К.К., Мастертон Р.Б., Бейкер Б.Н., Вентхолд Р.Дж. (август 1991 г.). «Акустический хиазм. III: Природа, распределение и источники афферентов к боковой верхней оливе у кошки». Журнал сравнительной неврологии . 310 (3): 377–400. дои : 10.1002/cne.903100308 . ПМИД 1723989 . S2CID 41964072 .
- ^ Ву Ш., Келли Дж. Б. (февраль 1994 г.). «Физиологические доказательства ипсилатерального торможения в боковой верхней оливе: синаптические реакции в срезе мозга мыши». Исследование слуха . 73 (1): 57–64. дои : 10.1016/0378-5955(94)90282-8 . ПМИД 8157506 . S2CID 34851384 .
- ^ Браунелл В.Е., Манис П.Б., Ритц Л.А. (ноябрь 1979 г.). «Ипсилатеральные тормозные реакции у кошки на боковой верхней оливе» . Исследования мозга . 177 (1): 189–193. дои : 10.1016/0006-8993(79)90930-2 . ПМК 2776055 . ПМИД 497821 .
- ^ Дэрроу К.Н., Maison SF, Liberman MC (декабрь 2006 г.). «Эфферентная обратная связь улитки уравновешивает межушную чувствительность» . Природная неврология . 9 (12): 1474–1476. дои : 10.1038/nn1807 . ПМК 1806686 . ПМИД 17115038 .
- ^ Базвински И., Бидмон Х.Дж., Зиллес К., Хилбиг Х. (декабрь 2005 г.). «Характеристика верхнего оливарного комплекса макаки-резуса с помощью кальцийсвязывающих белков и синаптофизина» . Журнал анатомии . 207 (6): 745–761. дои : 10.1111/j.1469-7580.2005.00491.x . ПМЦ 1571589 . ПМИД 16367802 .
- ^ Базвинский И., Хильбиг Х., Бидмон Х.Дж., Рубсамен Р. (февраль 2003 г.). «Характеристика верхнего оливарного комплекса человека с помощью кальцийсвязывающих белков и нейрофиламента H (SMI-32)». Журнал сравнительной неврологии . 456 (3): 292–303. дои : 10.1002/cne.10526 . ПМИД 12528193 . S2CID 22237348 .
- ^ Кулеша Р.Дж. (июль 2008 г.). «Цитоархитектура верхнего оливкового комплекса человека: ядра трапециевидного тела и заднего яруса». Исследование слуха . 241 (1–2): 52–63. дои : 10.1016/j.heares.2008.04.010 . ПМИД 18547760 . S2CID 44342075 .
- ^ Уорр В.Б., Гинан Дж.Дж. (сентябрь 1979 г.). «Эфферентная иннервация кортиевого органа: две отдельные системы». Исследования мозга . 173 (1): 152–155. дои : 10.1016/0006-8993(79)91104-1 . ПМИД 487078 . S2CID 44556309 .
- ^ Перейти обратно: а б Шофилд Б.Р., Cant NB (декабрь 1991 г.). «Организация верхнего оливкового комплекса у морской свинки. I. Цитоархитектура, гистохимия цитохромоксидазы и морфология дендритов». Журнал сравнительной неврологии . 314 (4): 645–670. дои : 10.1002/cne.903140403 . ПМИД 1726174 . S2CID 2030167 .
- ^ Фишл М.Дж., Бургер Р.М., Шмидт-Паули М., Александрова О., Синклер Дж.Л., Гроте Б. и др. (декабрь 2016 г.). «Физиология и анатомия нейронов медиальной верхней оливы мыши» . Журнал нейрофизиологии . 116 (6): 2676–2688. дои : 10.1152/jn.00523.2016 . ПМК 5133312 . ПМИД 27655966 .
- ^ Перейти обратно: а б с д и ж г час я дж Иллинг Р.Б., Краус К.С., Михлер С.А. (ноябрь 2000 г.). «Пластичность верхнего оливарного комплекса» . Микроскопические исследования и техника . 51 (4): 364–381. doi : 10.1002/1097-0029(20001115)51:4<364::AID-JEMT6>3.0.CO;2-E . ПМИД 11071720 .
- ^ Уорр В.Б., Бек Дж.Э. (апрель 1996 г.). «Множественные проекции вентрального ядра трапециевидного тела крысы». Исследование слуха . 93 (1–2): 83–101. дои : 10.1016/0378-5955(95)00198-0 . ПМИД 8735070 . S2CID 4721400 .
- ^ Перейти обратно: а б с д и ж г час я дж к Шофилд Б.Р., Cant NB (март 1992 г.). «Организация верхнего оливкового комплекса у морской свинки: II. Паттерны проекции периоливарных ядер на нижние холмики». Журнал сравнительной неврологии . 317 (4): 438–455. дои : 10.1002/cne.903170409 . ПМИД 1578006 . S2CID 21120946 .
- ^ Смит П.Х., Джорис П.С., Инь Т.С. (май 1993 г.). «Проекции физиологически охарактеризованных сферических аксонов кустистых клеток из ядра улитки кошки: свидетельства наличия линий задержки к медиальной верхней оливе». Журнал сравнительной неврологии . 331 (2): 245–260. дои : 10.1002/cne.903310208 . ПМИД 8509501 . S2CID 43136339 .
- ^ Вентольд Р.Дж., Хьюи Д., Альтшулер Р.А., Рикс К.А. (сентябрь 1987 г.). «Иммунореактивность глицина, локализованная в улитковом ядре и верхнем оливарном комплексе». Нейронаука . 22 (3): 897–912. дои : 10.1016/0306-4522(87)92968-X . ПМИД 3683855 . S2CID 22344089 .
- ^ Перейти обратно: а б Адамс Дж. К. (апрель 1983 г.). «Цитология периоливарных клеток и организация их отростков у кошки». Журнал сравнительной неврологии . 215 (3): 275–289. дои : 10.1002/cne.902150304 . ПМИД 6304156 . S2CID 45147218 .
- ^ Фриауф Э., Оствальд Дж (1988). «Дивергентные проекции физиологически охарактеризованных нейронов вентрального ядра улитки крысы, как показано с помощью внутриаксональной инъекции пероксидазы хрена». Экспериментальное исследование мозга . 73 (2): 263–284. дои : 10.1007/BF00248219 . ПМИД 3215304 . S2CID 15155852 .
- ^ Кулеша Р.Дж., Берреби А.С. (декабрь 2000 г.). «Верхнее параоливарное ядро крысы представляет собой ГАМКергическое ядро» . Журнал Ассоциации исследований в области отоларингологии . 1 (4): 255–269. дои : 10.1007/s101620010054 . ПМК 2957197 . ПМИД 11547806 .
- ^ Кулеша Р.Дж., Спироу Г.А., Берреби А.С. (апрель 2003 г.). «Физиологические свойства реакции нейронов верхнего параоливарного ядра крысы». Журнал нейрофизиологии . 89 (4): 2299–2312. дои : 10.1152/jn.00547.2002 . ПМИД 12612016 .
- ^ Уайт Дж.С., Уорр В.Б. (сентябрь 1983 г.). «Двойное происхождение оливокохлеарного пучка у крысы-альбиноса». Журнал сравнительной неврологии . 219 (2): 203–214. дои : 10.1002/cne.902190206 . ПМИД 6619338 . S2CID 44291925 .
- ^ Сент-Мари Р.Л., Бейкер Р.А. (август 1990 г.). «Специфическое для нейротрансмиттера поглощение и ретроградный транспорт [3H] глицина из нижнего холмика ипсилатеральными проекциями верхнего оливкового комплекса и ядрами латеральной петли». Исследования мозга . 524 (2): 244–253. дои : 10.1016/0006-8993(90)90698-B . ПМИД 1705464 . S2CID 21264622 .
- ^ Гинан Джей-Джей, Норрис Б.Е., Гинан СС (1972). «Отдельные слуховые единицы в верхнем оливочном комплексе: II: Расположение категорий единиц и тонотопическая организация». Международный журнал неврологии . 4 (4): 147–166. дои : 10.3109/00207457209164756 .
- ^ Дэвис К.А., Рамачандран Р., Мэй Б.Дж. (июль 1999 г.). «Одноединичные реакции в нижних холмиках децеребрированных кошек. II. Чувствительность к различиям в межушном уровне». Журнал нейрофизиологии . 82 (1): 164–175. дои : 10.1152/jn.1999.82.1.164 . ПМИД 10400945 .
- ^ Цучитани С (март 1977 г.). «Функциональная организация боковых групп клеток верхнего оливкового комплекса кошки». Журнал нейрофизиологии . 40 (2): 296–318. дои : 10.1152/jn.1977.40.2.296 . ПМИД 845625 .
- ^ Спироу Г.А., Берреби А.С. (апрель 1996 г.). «Организация вентролатеральных периоливарных клеток верхней оливы кошки, выявленная с помощью иммуноцитохимии PEP-19 и окраски по Нисслю» . Журнал сравнительной неврологии . 368 (1): 100–120. doi : 10.1002/(SICI)1096-9861(19960422)368:1<100::AID-CNE7>3.0.CO;2-7 . ПМИД 8725296 .