HLA-DQ8
Эта статья может быть слишком технической для понимания большинства читателей . ( Май 2017 г. ) |
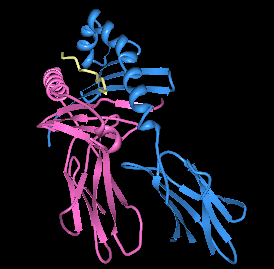 Рендеринг HLA-DQ8 с помощью иммунодоминантного инсулинового пептида в связывающем кармане. | |||||||||||||||||
| Тип полимера | MHC класса II , DQ антиген клеточной поверхности | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||
| Редкие гаплотипы | |||||||||||||||||
| |||||||||||||||||
HLA-DQ8 ( DQ8 ) представляет собой человеческого лейкоцитарного антигена серотип в группе серотипов HLA-DQ (DQ). DQ8 представляет собой расщепленный антиген антигена DQ3 широкого . DQ8 определяется распознаванием антителами β 8 и это обычно обнаруживает генный продукт DQB1 *0302 .
DQ8 обычно связан с аутоиммунными заболеваниями у людей. DQ8 является второй наиболее распространенной изоформой , связанной с целиакией , и DQ, наиболее связанной с диабетом 1 типа . DQ8 увеличивает риск развития ревматоидного артрита и связан с основным локусом риска РА, HLA-DR4 . DR4 также играет важную роль при диабете 1 типа. Хотя гаплотип DQ8.1 связан с заболеванием, связь гаплотипов DQB1*0305, DQ8.4 или DQ8.5 (см. информационный блок) с аутоиммунным заболеванием неизвестна; однако это может быть результатом недостаточного изучения популяций, которые являются их носителями, и очень низкой частоты.
DQ8.1 также отличается от других HLA по популяционным частотам. Обычно для антигенов MHC класса II у человека частоты гаплотипов не превышают 40%. Например, в США самая высокая частота гаплотипа, гаплотипа, кодирующего DQ6.2, составляет около 15%, что соответствует частоте фенотипов менее 30%. Частота атипичных гаплотипов превышает 40%.
Для DQ8 самые высокие частоты гаплотипов достигают 80% в некоторых частях Центральной и Южной Америки, а частоты фенотипов приближаются к 90%. Это самая высокая частота фенотипа, наблюдаемая с большим отрывом для любого фенотипа DR или DQ в человеческой популяции.
Серология
[ редактировать ]| DQB1* | DQ8 | ДК3 | DQ7 | Образец |
| аллель | % | % | % | размер (Н) |
| *0302 | 66 | 23 | 4 | 6687 |
| *0304 | 8 | 35 | 40 | 111 |
| *0305 | 34 | 30 | 70 | |
| Красный цвет указывает на уровень «ложной» реакции у серотипов, отличных от DQ8. | ||||
| Связь аллелей с базой данных IMGT/HLA в EBI | ||||
| частота | ||
| исх. | Население | (%) |
| [ 2 ] | Гватемала Майя | 48.1 |
| Мексика Н.Леон Метисы | 22.5 | |
| Выходцы из Латинской Америки Южного Техаса США | 20.6 | |
| Швеция | 18.7 | |
| Россия Сибирь Негидаль | 18.6 | |
| Россия Мурманск Саоми | 18.5 | |
| Джордан Амман | 17.8 | |
| Самоа | 17.2 | |
| Англия европеоидная | 16.4 | |
| Финляндия | 15.7 | |
| Франция | 14.5 | |
| Центральная Япония | 10.8 | |
| Греция Крит | 9.2 | |
| Испания Баскы Долина Арратия | 6.7 | |
| Алжир Оран | 6.6 | |
| Китай Пекин и Сиань | 6.1 | |
| Эфиопия Амхара | 5.6 | |
| США ЮВ Афроамериканец | 4.9 | |
| США, коренные жители Аляски юпики | 3.8 | |
| Индия Северные индусы | 3.0 | |
| Зимбабве Хараре Шона | 2.2 | |
| Россия Сибирь Эскимосы | 0.9 | |
| Руанда Кигали Хуту и тутси | 0.5 | |
| PNG Восточное Хайлендс Горока | 0.0 | |
| Тунис Джерба Бербер | 0.0 |
Хотя ложная реакция на DQB1*0302 невелика, эффективность положительной реакции невысока, и существует риск ложного обнаружения DQB1*0305, что может привести к несовместимости. Для диагностики и подтверждения заболевания не существует известной связи DQB1*0305 с целиакией или аутоиммунным диабетом. Поэтому разумно использовать набор текста DQB1 с высоким разрешением для DQ8.
Аллели
[ редактировать ]| ДК | ДК | ДК | Частота | |||||
|---|---|---|---|---|---|---|---|---|
| Серотип | цис-изоформа | Подтип | А1 | Б1 | % [ 3 ] | классифицировать | ||
| DQ8 | а 3 - б 8 | 8.1 | 0301 | 0302 | 9. | 62 | 6-е место | |
| 0302 ♠ | 0302 | 0. | 93 | |||||
| ♠ DQA1*0302 и *0303 не решены | ||||||||
DQ8 определяется распознаванием антителами β 8 и осложняется тем, что DQ8 распознает некоторые HLA-DQB1 *03 кодирует изоформы хорошо, частично или совсем плохо (см. серологию ) DQ β 3.2 и б 3.5 лучше всего известны как DQ8. Эти расщепленные антигены являются аллельными продуктами DQB1. *0302 и DQB1 *0305 соответственно.
DQB1*0302
[ редактировать ]| частота | ||
| исх. | Население | (%) |
| [ 2 ] | Ливан у моста Кафар | 5.9 |
| Ливан Ниха-эль-Шуфф | 2.5 | |
| Лакота Сиу | 2.1 | |
| Тунис | 2.0 | |
| Ливан Юхмур | 1.8 | |
| Гватемала Майя | 1.5 | |
| Марокко | 1.0 | |
| Пакистан | 1.0 | |
| Саудовская Аравия Гураят и Хайль | 0.8 | |
| Китай Лицзян Наси | 0.7 | |
| Russia Chuvash | 0.6 | |
| Тунис Берберский вопрос | 0.6 | |
| Чешская Республика | 0.5 | |
| Индия Дели | 0.5 |
DQB1 *0302 и чаще всего встречается в гаплотипе DQA1. *0301 :DQB1 *0302 , около 10% случаев встречается в гаплотипе DQA1. *0302 :DQB1 *0302 . DQB1*0302 почти всегда связаны с DR4, DRB1. *0401 , *0402 и *0404 у кавказцев. Первый и третий DRB1 наиболее тесно связаны с ревматоидным артритом.
DQB1*0305
[ редактировать ]DQB1 * Продукт гена 0305 несколько интенсивнее реагирует с DQ8, чем с DQ7, в Европе он обычно редок. Северная Америка, за исключением нескольких коренных народов. Уровни DQB1 *0305 , вероятно, выше, поскольку более ранние тесты не позволяли хорошо различать разные *03.
Гаплотипы
[ редактировать ]β-цепи DQ8 объединяются с α-цепями, кодируемыми генетически связанными аллелями HLA-DQA1 , образуя изоформы цис - гаплотипа . Существует только одна общая цис-изоформа DQ8, поскольку связанный DQA1 *03 аллели(2) встречаются у большей части населения, DQ8.1 является подавляющим большинством цис-изоформы DQ8. Редкий гаплотип DQA1. *0503 :DQB1 *0302 обнаруживается ниже 1% всех гаплотипов DQ8 в Азии и Мезоамерике. Еще один более редкий гаплотип, DQA1. *0401 :DQB1 *0302
ДК8.1
[ редактировать ]| Ссылка | DQA1 | DQB1 | Оцененный | |
|---|---|---|---|---|
| Население | *03 | *0302 | ДК8.1 | |
| [ 4 ] | Лакандон Майя (Мексика) |
79.0 | 77.9 | 77.9 |
| [ 5 ] | Перри-Юкпа (Венесуэла) |
74.0 | 75.0 | 74.9 |
| [ 6 ] | майя (Гватемала) |
48.1 | 47.6 | |
| [ 7 ] | Масатекцы (Мексика) |
48.5 | 48.5 | 47.5 |
| [ 8 ] | Ламы (Перу) |
45.2 | 44.7 | |
| [ 9 ] | Дакота Сиу (С.Дакота) |
52.1 | 45.0 | 44.5 |
| [ 10 ] | Микстек (Оахака, Мексика) |
40.0 | 35.9 | 35.4 |
| [ 11 ] | Лакота Сиу (С.Дакота) |
25.7 | 25.5 | |
| [ 12 ] | Поле (Бразилия) |
17.5 | 17.0 | |
| [ 13 ] | Коук., Сан-Антонио (США) |
11.7 | 11.7 | |
| [ 14 ] | Кавказский (США) |
18.5 | 10.5 | 10.5 |
| [ 15 ] | Афроамериканец (ЮВ. США) |
4.9 | 4.5 | |
| [ 16 ] | Тлинглет (Аляска, США) |
14.0 | 8.5 | 8.5 |
| [ 17 ] | эскимосский (Аляска, США) |
3.8 | 3.8 | |
| [ 18 ] | Навахо Канончито (Нью-Мексико, США) |
6.3 | 3.5 | 3.5 |
| [ 19 ] | эскимосский (Э. Гренландия) |
0.0 | 0.0 | 0.0 |
DQA1 *0301 :DQB1 *0302 (DQ8.1) — наиболее распространенный подтип DQ8, представляющий более 98% популяции носителей DQ8. Нечасто, DQA1 *0302 :DQB1 *0302 , но это замена альфа-цепи, DQA1* *0301 по сравнению с *0302 , находится за пределами переплетной щели и, по всей видимости, не влияет на функцию DQ8. DQ8.1 встречается почти повсеместно в каждой региональной популяции человека, но из-за своего уникального распространения он становится объектом молекулярной антропологии. Есть 3 места, где частота гаплотипов повышена: Центральная и Южная Америка, северо-восточная часть Тихоокеанского региона и Северная Европа.
Высокие частоты в Америке
[ редактировать ]Глобальный узел DQ8 находится в Центральной Америке и на севере Южной Америки, где он достигает самой высокой частоты для любого отдельного серотипа DQ, близкой к 90% частоте фенотипа (частота гаплотипа 77%), и относительно высокой частоте встречается у коренного населения Северной Америки. и прибрежные районы Мексиканского залива и долины Миссисипи. Высокая частота DQ8 в северо-восточных регионах Южной Америки. [ 5 ] и низкая частота у коренных американцев более недавнего азиатского происхождения. [ 16 ] [ 19 ] или сибирского происхождения [ 20 ] предполагают, что DQ8 встречался с высокой частотой у самых ранних америндов. Характер распределения согласуется с недавними результатами мтДНК, предполагающими, что первые мигранты в Новый Свет селились в низменных прибрежных районах, речных долинах и медленно продвигались вглубь страны, последующие поселенцы переместились в высокогорные районы. DQ8 и DQ2.5 имеют много аналогичных функциональных сходств, и эта предвзятость первого поселенца может быть причиной сходства. Исследования вируса Эпштейна-Барра [ 21 ] и другие белки предполагают, что оба белка являются кислыми (то есть пептиды с повышенным отрицательным зарядом) пептидными презентаторами (иллюстрацию процесса презентации см. в DQ8) и, возможно, были адаптивными к определенному образу жизни охоты и собирательства, возможно, прибрежным собирателям.
| Ссылка | DQA1 | DQB1 | Оцененный | |
|---|---|---|---|---|
| Население | *03 | *0302 | ДК8.1 | |
| [ 20 ] | Нивхи (ССВ. Дроссель И.) |
0.0 | 0 | |
| [ 22 ] | Зорастра, Ядз (Иран) |
20.8 | 0.8 | 0.8 |
| [ 23 ] | Хотон (Монголия) |
24.4 | 1.2 | 1.2 |
| [ 24 ] | Маданг (Папуа-Новая Гвинея) |
1.5 | 1.5 | 1.5 |
| [ 25 ] | Их (Китай) |
2.6 | 2.6 | |
| [ 26 ] | Наси (Лицзян, Китай) |
2.7 | 2.7 | |
| [ 27 ] | Айны (Япония) |
3.0 | 3.0 | |
| [ 28 ] | Шаньдун Хан (Китай) |
3.1 | 3.1 | |
| [ 29 ] | Халха (Монголия) |
22.3 | 6.1 | 6.1 |
| [ 30 ] | Тайцы | 21.5 | 7.4 | 7.4 |
| [ 31 ] | Ket, Yenisey (Сибирь) |
14.7 | 8.4 | 8.4 |
| [ 32 ] | Южная Корея | 10.3 | 10.3 | |
| [ 33 ] | японский | 10.8 | 10.8 | |
| [ 34 ] | Нганасан (Сибирь) |
39.6 | 11.4 | 11.4 |
| [ 35 ] | Уйгуры, Урумчи (Китай) |
21.9 | 11.4 | 11.4 |
| [ 36 ] | казахский | 11.4 | 11.4 | |
| [ 37 ] | Негидаль (Сибирь) |
50.0 | 18.6 | 18.6 |
| [ 38 ] | Остров Кука. (Тихий океан) |
25.0 | 25.0 |
Изобилие в Азии
[ редактировать ]Перерыв DQ8 в северо-восточной части Сибирской Арктики, повышенные уровни в Амурской области и восточных тюрках
Уровни DQ8 на юго-западе и в западной части Тихоокеанского региона находятся на различных частотах гаплотипов, от 2 до 30%, и стабилизируются примерно на 10% для Рюкюаня, японцев, корейцев, Амурских регионов, а в северо-западной части Тихоокеанского региона падают до менее 1% в нивхи. В настоящее время в регионе Аляска-Восточная Сибирь наблюдается перерыв в работе DQ8, и неясно, связано ли это с заменой, отбором или способом прибытия первых американцев (т.е. строго морским маршрутом). Типы DR, связанные с DQ8: DRB1*0403, *0404, *0406, *0407, *0408 и *0401 разделены между многими гаплотипами DQA1:B1. DQB1*0405 обычно ассоциируется с DQA1*0303:DQB1*04 и поэтому не включается в DRB1*0401 при оценках с высоким разрешением. У острова Кука DQ8 был только один ассоциированный гаплотип DR, что позволяет предположить, что разнообразие ограничивает интродукцию в регион либо по маршруту TW- (Япония/Корея/Китай), либо через запад, например, у Бунуна высокий DRB1*0403. Большая часть DRB1*04, по-видимому, перераспределилась из Восточной Азии из неизвестного источника, возможно, в Центральной Азии или Индии. Распределение можно сравнить с группами коренных жителей, такими как жители Южной Америки. Три группы с высокими уровнями, когуи, сикуни и юкпа, имеют около 75% DQ8, доминантным аллелем DRB1* в 2 из 3 является *0411 (Северный Китай = 0), но *0407 (рюкю, японцы, манси -Восточный Урал, китайцы Наси) и *0403 (нганасанцы, буряты, негидальцы, тунисцы, Рюкю, Корея, айны) также встречаются. В Северной Америке DRB1*0404 и *0407 более распространены, чем *0403, а у сиу лакота B1*0411 встречается редко. Гаплотип DRB1*0404-DQ8 чаще встречается в Северо-Западной Азии и Северной Европе.
| Ссылка | DQA1 | DQB1 | Оцененный | |
|---|---|---|---|---|
| Население | *03 | *0302 | ДК8.1 | |
| [ 39 ] | Ненецкий (Северная Россия) |
40.9 | 20.9 | 20.0 |
| [ 40 ] | Мурманский саоми (Россия) |
38.3 | 18.5 | 17.2 |
| [ 41 ] | Arkhange Pomors (Россия) |
22.6 | 17.1 | 16.4 |
| [ 42 ] | Шведский | 24.2 | 18.7 | 16.1 |
| [ 43 ] | Кавказский (Англия) |
23.7 | 16.4 | 15.9 |
| [ 44 ] | Финляндия | 15.7 | 15.2 | |
| [ 45 ] | CEPH Франции | 21.3 | 14.5 | 13.5 |
| [ 46 ] | датчанин | 20.9 | 13.2 | 13.2 |
| [ 47 ] | Голландский | 11.2 | 11.0 | |
| ирландский | 10.6 | |||
| [ 48 ] | Северо-западный славянский (Россия) |
16.0 | 11.0 | 10.5 |
| [ 49 ] | немецкий | 10.5 | 9.9 | |
| [ 50 ] | Русский | 8.9 | 7.2 | |
| [ 51 ] | Они будут петь (Испания) |
19.3 | 8.4 | 7.0 |
| [ 49 ] | испанский | 8.9 | 6.5 | |
| [ 49 ] | Сардинский | 4.9 | 4.9 | |
| [ 49 ] | итальянский | 4.6 | 4.5 | |
| [ 52 ] | Объединенные Арабские Эмираты | 0.83 | 0.8 | |
| [ 53 ] | Иордания | 24.1 | 17.9 | 17.9 |
| Африка | ||||
| [ 49 ] | !Если | 36.7 | 36.7 | |
| [ 49 ] | Хой | 14.9 | 13.9 | |
| [ 54 ] | Оромо | 17.5 | 9.0 | 9.0 |
| [ 55 ] | Марокко | 17.2 | 8.9 | 8.6 |
| [ 56 ] | Тунис | 10.31 | 10.0 | 8.0 |
| [ 54 ] | Амхара | 11.2 | 5.6 | 5.6 |
| [ 57 ] | Они же пигмеи | 3.2 | 3.0 | |
| [ 49 ] | негр (Северная Африка) |
3.1 | 2.7 | |
| [ 58 ] | Кэмерон | 11.5 | 1.5 | 1.2 |
| [ 57 ] | Банту Конго | 0.6 | 0.5 | |
| [ 59 ] | Габонский | 7.5 | 0.5 | |
| [ 60 ] | Плохо | 17.5 | 0.0 | 0.0 |
Высокие уровни DQ8 в Северной Европе
[ редактировать ]DQ8 также широко распространен в Северной Европе и часто встречается у немецко-скандинавско-уральского населения к северу от Швейцарии. Гаплотипы HLA AB позволяют предположить, что за DQ8 ответственна миграция людей к востоку от Урала, возможно, даже из западной части Тихоокеанского региона. Высокий уровень DQ8 и DQ2,5 представляет большой интерес для DQ-опосредованных заболеваний Скандинавии и Северной Европы. DQ8 также обнаружен в Иберии и местах, где поток генов с востока на запад с помощью других генетических маркеров не может быть подтвержден, а уровни внутри африканской или ближневосточной популяции являются возможными источниками. Иберия имеет значительное равновесие A1 / B1, что позволяет предположить независимые источники из Африки.
Глобальное распространение DQ8
[ редактировать ]DQ8 вместе с несколькими другими гаплотипами, по-видимому, разделен на северо-запад и юго-восток Евразии, а данные о DQ2.5 и других гаплотипах позволяют предположить, что древняя популяция Центральной Азии была вытеснена более поздней африканской миграцией. Есть много общих маркеров, обнаруженных у французов, немцев, датчан, шведов, тибетцев, жителей реки Амур, японцев и корейцев, которые являются потенциальными индикаторами этого двустороннего распространения. Гаплотипы DQ8 встречаются с высокими частотами в !Кунге , хотя можно ожидать большего количества DQ8 в Австронезии, он распространен повсеместно, иногда с низкими частотами, иногда с более высокими частотами (Тайский). Путь распространения DQ8 в Новый Свет загадочен, конечно, Япония и река Амур являются мощными источниками, но нельзя исключать и другие перемещенные группы населения. Если способ передвижения осуществлялся через Берингийский коридор, как предполагает археолог, то очень низкая частота DQ8 в настоящее время является очень необычной находкой с точки зрения свидетельств полного перемещения в других частях мира. Маркеры, общие для японцев и аборигенов TW, имеют тенденцию к снижению частоты по мере приближения к Сибири, маркеры мтДНК снижаются в Курильской цепи. Похоже, что в период Дзёмон в Японии произошло перемещение предков Нинвет/айнов и депрессия DQ8 по всей северной Японии, но упадок во всем регионе несколько необъясним, если не считать катастрофического климатического события между заселением Нового Света и текущее время.
Альтернативная модель состоит в том, что у населения Северо-Восточной Азии было несколько источников DQ8, некоторые источники были из Центральной Азии, а некоторые из Индокитая, некоторые из DQ8, обнаруженных в Северо-Западной Евразии, могли быть из смеси Западного Тихоокеанского региона и Центральной Азии. азиатские источники и были перемещены из более центральных регионов, но не из более восточных регионов.
DQ8 и выбор
[ редактировать ]Как и DQ2.5, DQ8 мог отбираться для морских и прибрежных народов, добывающих пищу, и, в частности, для народов, адаптированных к климату/ситуации среды обитания на северной оконечности обитаемой западной части Тихого океана во время последнего ледникового максимума. При культивировании Triticeae можно применять отрицательный отбор по DQ8. существовало множество представителей видов Triticeae, похожих на дикие средневосточные Triticeae Несмотря на то, что в Америке , а также большое количество одомашненных растений в Новом Свете, ни один вид Triticeae, по-видимому, не был одомашнен в Новом Свете, и нет четких примеров. в близкородственных трибах трав. Среди новых видов трав в мире после Колумбии один вид Elymus был одомашнен для употребления в пищу человеком, а другой - как пастушеский сорт. Это можно интерпретировать двояко. Во-первых, уровни DQ8 отрицательно тормозили одомашнивание штаммов Triticeae . Во-вторых, отсутствие таких сортов, более подходящих, чем уже разработанные сорта, позволило DQ8 подняться или остаться высоким, в то время как уровни DQ2,5 в NW при гораздо более долгосрочной селекции упали, или немного и то, и другое. Большинство американских сортов были одомашнены к югу от Рио-Гранде (исключением являются рис Каддо, техасская тыква и т. д.). Пшеница, особенно ячмень и рожь, являются предпочтительными сортами в более прохладном климате, тогда как Зеа более адаптивна в тропическом климате, а некоторые сорта относительно устойчивы к засухе. Однако Зеа не имеет определенных аминокислот, которые необходимо дополнять другими продуктами, чтобы предотвратить недоедание. Близость неолитизации к экватору в Новом Свете, возможно, во многом связана с неявным негативным отбором DQ8 относительно неолитизации Западной Евразии.

DQ8 и болезнь
[ редактировать ]В Европе DQ8 связан с диабетом 1 типа и целиакией (также известной как целиакия). Самым высоким фактором риска развития диабета 1 типа является фенотип HLA DQ8/DQ2.5. В некоторых частях восточной Скандинавии как DQ2.5, так и DQ8 значительно повышают частоту позднего начала диабета I типа и неоднозначного диабета I/II типа. DQ8 также обнаружен у многих коренных народов Азии . Он был обнаружен на ранних стадиях у бедуинского населения Аравии, где DQ2.5 часто отсутствует, и в этих случаях DQ8 ассоциирован исключительно с HLA при целиакии.
Однако в Соединенных Штатах , похоже, наблюдается сдвиг в риске аутоиммунных заболеваний у иммигрантов из Мексики. Повышенная иммунореактивность латиноамериканцев в Хьюстоне, по-видимому, связана с DR4-DQ8. Гаплотип может нести самый высокий риск развития ревматоидного артрита.
В Японии DQ3 (DQ7, DQ8, DQ9) связан с миастенией гравис у женщин с ранним началом, хотя не похоже, что DQ8 играет большую роль. Есть сходство между миастенией гравис в Японии и миастенией, обнаруженной у латиноамериканцев Хьюстона. с DQ8, связанным с более молодыми женщинами, по сравнению с ассоциациями всех других типов HLA DQ. В Японии растет число случаев целиакии, и очевидно, что причиной являются изменения в питании, но в Японии также нет DQ2,5, а уровни DQ8 умеренные.
Связь DR4
[ редактировать ]Многие заболевания, связанные с DQ8, имеют двойную связь с DR4, а некоторые DR4 (*0405) имеют независимую и зависимую связь риска с DQ8, например, с диабетом 1 типа. [ 61 ] [ 62 ]
Ссылки
[ редактировать ]- ^ Форма запроса аллелей IMGT/HLA - Европейский институт биоинформатики
- ^ Перейти обратно: а б Миддлтон Д., Менчака Л., Руд Х., Комерофски Р. (2003). «Новая база данных частот аллелей: http://www.allerefrequency.net» . Тканевые антигены . 61 (5): 403–7. дои : 10.1034/j.1399-0039.2003.00062.x . ПМИД 12753660 .
- ^ Клитц В., Майерс М., Спеллман С. и др. (октябрь 2003 г.). «Новые эталонные стандарты частоты гаплотипов HLA: типирование гаплотипов HLA DR-DQ с высоким разрешением и большой выборкой в выборке американцев европейского происхождения». Тканевые антигены . 62 (4): 296–307. дои : 10.1034/j.1399-0039.2003.00103.x . ПМИД 12974796 .
- ^ «Обзор демографического исследования
(Материалы семинара IHW, Сиэтл, 2002 г.) - Мексика Майя Лакандон» . Частоты аллелей в популяции во всем мире . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. - ^ Перейти обратно: а б Лайрис З, Гедес И, Домингес Э, Пас Н, Монтаньани С, Матос М, Эррера Ф, Огандо В, Бальбас О, Родригес-Ларральде А (2001). «Расширенные гаплотипы HLA у индейцев Карибского бассейна: юкпа хребта Перия». Хум Иммунол . 62 (9): 992–1000. дои : 10.1016/S0198-8859(01)00297-X . ПМИД 11543901 .
- ^ Гомес-Касадо Э, Мартинес-Ласо Х, Москосо Х, Самора Дж, Мартин-Вилла М, Перес-Блас М, Лопес-Санталла М, Лукас Грамахо П, Сильвера С, Лоуи Э, Арнаис-Виллена А (2003). «Происхождение майя по генам HLA и уникальность индейцев». Тканевые антигены . 61 (6): 425–36. дои : 10.1034/j.1399-0039.2003.00040.x . ПМИД 12823766 .
- ^ Арнаис-Виллена А, Варгас-Аларкон Г, Гранадос Х, Гомес-Касадо Э, Лонгас Х, Гонсалес-Хевилла М, Суньига Х, Сальгадо Н, Эрнандес-Пачеко Г, Гильен Х, Мартинес-Ласо Х (2000). «Гены HLA у мексиканских масатеканцев» Тканевые антигены . 56 (5): 405–16. дои : 10.1034/j.1399-0039.2000.560503.x . ПМИД 11144288 .
- ^ Москосо Дж., Секлен С., Серрано-Вела Дж., Вильена А., Мартинес-Ласо Дж., Самора Дж., Морено А., Ира-Кашафейро Дж., Арнаис-Виллена А. (2006). «Гены HLA у перуано-амазонских индейских лам» Мол Иммунол . 43 (11): 1881–9. дои : 10.1016/j.molimm.2005.10.013 . ПМИД 16337001 .
- ^ «Обзор демографического исследования
(Материалы семинара IHW, Сиэтл, 2002 г.) - США, Южная Дакота Сиу» . Частоты аллелей в мировой популяции . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. - ^ Холленбах Дж., Томсон Г., Као К., Фернандес-Вина М., Эрлих Х., Бугаван Т., Винклер С., Винтер М., Клитц В. (2001). «Разнообразие, дифференциация и эволюция гаплотипов HLA у коренных жителей Мезоамерики». Хум Иммунол . 62 (4): 378–90. дои : 10.1016/S0198-8859(01)00212-9 . ПМИД 11295471 .
- ^ Леффелл М., Фаллин М., Хильдебранд В., Каветт Дж., Иглхарт Б., Закари А. (2004). «Аллели и гаплотипы HLA среди сиу лакота: отчет о семинарах меньшинств ASHI, часть III». Хум Иммунол . 65 (1): 78–89. дои : 10.1016/j.humimm.2003.10.001 . ПМИД 14700599 .
- ^ Ласаро А., Мораес М., Маркос С., Мораес Х., Фернандес-Винья М., Стастный П. (1999). «Эволюция HLA-класса I по сравнению с полиморфизмом HLA-класса II у Терены, южноамериканского индейского племени». Хум Иммунол . 60 (11): 1138–49. дои : 10.1016/S0198-8859(99)00092-0 . ПМИД 10600013 .
- ^ «Обзор демографического исследования - кавказцы США, Сан-Антонио» . Частоты аллелей в популяции мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г.
- ^ Мацудзака Ю, Макино С, Накадзима К, Томизава М, Ока А, Бахрам С, Кульски Дж, Тамия Г, Иноко Х (2001). «Новые полиморфные микросателлитные маркеры в области III класса MHC человека». Тканевые антигены . 57 (5): 397–404. дои : 10.1034/j.1399-0039.2001.057005397.x . ПМИД 11556964 .
- ^ Каффнер Т., Уитворт В., Джайрам М., МакНиколл Дж. (2003). «Гены HLA класса II и TNF у афроамериканцев из юго-востока США: региональные различия в частотах аллелей». Хум Иммунол . 64 (6): 639–47. дои : 10.1016/S0198-8859(03)00056-9 . ПМИД 12770797 .
- ^ Перейти обратно: а б Нельсон Дж., Бойер Г., Темплин Д., Ланье А., Баррингтон Р., Нисперос Б., Смит А., Микельсон Э., Хансен Дж. (1992). «HLA-антигены у индейцев тлинкитов, больных ревматоидным артритом» Тканевые антигены . 40 (2): 57–63. дои : 10.1111/j.1399-0039.1992.tb01960.x . ПМИД 1412417 .
- ^ Леффелл М., Фаллин М., Эрлих Х., Фернандес-Вина М., Хильдебранд В., Мак С., Закари А. (2002). «Антигены, аллели и гаплотипы HLA среди юпиков, коренных жителей Аляски: отчет о семинарах меньшинств ASHI, Часть II». Хум Иммунол . 63 (7): 614–25. дои : 10.1016/S0198-8859(02)00415-9 . ПМИД 12072196 .
- ^ Перейти обратно: а б Велиндер Л., Граугард Б., Мэдсен М. (2000). «Частоты антигена и генов HLA у эскимосов Восточной Гренландии». Европейский журнал иммуногенетики . 27 (2): 93–7. дои : 10.1046/j.1365-2370.2000.00209.x . ПМИД 10792425 .
- ^ Перейти обратно: а б Лу Х, Ли Х, Куваяма М, Яшики С, Фудзиеси Т, Суэхара М, Осаме М, Ямасита М, Хаями М, Гурцевич В, Баллас М, Иманиши Т, Сонода С (1998). «HLA класса I и класса II у нивхов, коренного населения, несущего HTLV-I, на Сахалине, Дальний Восток России». Тканевые антигены . 52 (5): 444–51. дои : 10.1111/j.1399-0039.1998.tb03071.x . ПМИД 9864034 .
- ^ Сидни Дж., дель Гуэрсио М.Ф., Саутвуд С. и Сетте А. (2002). «Молекулы HLA DQA1*0501/B1*0201 и DQA1*0301/B1*0302 имеют сильное перекрытие в специфичности связывания пептидов» . Журнал иммунологии . 169 (9): 5098–5108. дои : 10.4049/jimmunol.169.9.5098 . ПМИД 12391226 .
- ^ «Обзор демографического исследования - зороастрийцы Ирана Йезда» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Фарджадиан С., Мокадам Ф., Гадери А. (2006). «Полиморфизм гена HLA класса II у парсов и зороастрийцев Ирана». Международный журнал иммуногенетики . 33 (3): 185–91. дои : 10.1111/j.1744-313X.2006.00594.x . ПМИД 16712649 . S2CID 12667253 .
- ^ «Обзор демографического исследования - Монголия Хотон Тариалан» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Мачулла Х., Батнасан Д., Стейнборн Ф., Уяр Ф., Сарухан-Дирескенели Г., Огуз Ф., Карин М., Дорак М. (2003). «Генетическое сходство монгольских этнических групп и их отношение к тюркам». Тканевые антигены . 61 (4): 292–9. дои : 10.1034/j.1399-0039.2003.00043.x . ПМИД 12753667 .
- ^ «Обзор демографического исследования - Папуа-Новая Гвинея, Маданг» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г.
- ^ «Обзор демографического исследования - Китай, провинция Юньнань, Яо» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Лю Ю, Лю З, Фу Ю, Цзя З, Чен С, Сюй А (2006). «Полиморфизм генов HLA класса II у народностей мяо и яо Юго-Западного Китая». Тканевые антигены . 67 (2): 157–9. дои : 10.1111/j.1399-0039.2006.00510.x . ПМИД 16441488 .
- ^ «Обзор демографического исследования - Китай Лицзян Наси» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Фу Ю, Лю З, Линь Дж, Цзя З, Чен В, Пань Д, Лю Ю, Чжу Ю, Чен Р, Сюй А (2003). «Полиморфизм HLA-DRB1, DQB1 и DPB1 у этнической группы наси на юго-западе Китая». Тканевые антигены . 61 (2): 179–83. дои : 10.1034/j.1399-0039.2003.00012.x . ПМИД 12694588 .
- ^ Баннаи М., Токунага К., Иманиши Т., Харихара С., Фудзисава К., Джуджи Т., Омото К. (1996). «Аллели HLA класса II у айнов, живущих в районе Хидака, Хоккайдо, северная Япония». Am J Phys Anthropol . 101 (1): 1–9. doi : 10.1002/(SICI)1096-8644(199609)101:1<1::AID-AJPA1>3.0.CO;2-Z . PMID 8876810 .
- ^ «Обзор демографического исследования - Китай, Шаньдун Хань» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Чжоу Л, Линь Б, Се Ю, Лю З, Ян В, Сюй А (2005). «Полиморфизм генов человеческого лейкоцитарного антигена-DRB1, -DQB1 и -DPB1 популяции Шаньдун-Хань в Китае». Тканевые антигены . 66 (1): 37–43. дои : 10.1111/j.1399-0039.2005.00418.x . ПМИД 15982255 .
- ^ Четыре ссылки: 1. Мачулла Х., Батнасан Д., Стейнборн Ф., Уяр Ф., Сарухан-Дирескенели Г., Огуз Ф., Карин М., Дорак М. (2003). «Генетическое сходство монгольских этнических групп и их отношение к тюркам». Тканевые антигены . 61 (4): 292–9. дои : 10.1034/j.1399-0039.2003.00043.x . ПМИД 12753667 . , 2. Чимге Н., Танака Х., Касивасе К., Аюш Д., Токунага К., Саджи Х., Аказа Т., Бацуури Дж., Джуджи Т. (1997). «Система HLA у населения Монголии». Тканевые антигены . 49 (5): 477–83. дои : 10.1111/j.1399-0039.1997.tb02782.x . ПМИД 9174140 . , 3. «Обзор демографического исследования - Монголия Халха» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 4. «Обзор демографического исследования - Монголия, Халх, Улан-Батор» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г.
- ^ «Обзор демографического исследования - Таиланд» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г.
- ^ «Обзор демографического исследования - Россия Сибирь Кет Нижний Енисей» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Уинук-Оол Т, Такезаки Н, Дербенева О, Володько Н, Сукерник Р (2004). «Вариации генов HLA класса II у нганасан и кетов, двух аборигенных сибирских популяций». Европейский журнал иммуногенетики . 31 (1): 43–51. дои : 10.1111/j.1365-2370.2004.00443.x . ПМИД 15009181 .
- ^ Ли К, О Д, Ли С, Ян С (2005). «Аллельное и гаплотипическое разнообразие генов HLA-A, -B, -C, -DRB1 и -DQB1 в корейской популяции». Тканевые антигены . 65 (5): 437–47. дои : 10.1111/j.1399-0039.2005.00386.x . ПМИД 15853898 .
- ^ Сайто С., Ота С., Ямада Э., Иноко Х., Ота М. (2000). «Частоты аллелей и гаплотипические ассоциации, определяемые типированием аллельной ДНК в локусах HLA класса I и класса II в японской популяции» . Тканевые антигены . 56 (6): 522–9. дои : 10.1034/j.1399-0039.2000.560606.x . ПМИД 11169242 .
- ^ «Обзор демографического исследования - Россия Сибирь Нганасан Дудинка» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Уинук-Оол Т, Такезаки Н, Дербенева О, Володько Н, Сукерник Р (2004). «Вариации генов HLA класса II у нганасан и кетов, двух аборигенных сибирских популяций». Европейский журнал иммуногенетики . 31 (1): 43–51. дои : 10.1111/j.1365-2370.2004.00443.x . ПМИД 15009181 .
- ^ «Обзор демографического исследования - Китай, Урумчи-Уйгур» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Мизуки Н, Оно С, Андо Х, Сато Т, Иманиши Т, Годобори Т, Исихара М, Гото К, Ота М, Гэн З, Гэн Л, Ли Г, Иноко Х (1998). «Аллели класса II главного комплекса гистосовместимости у уйгурского населения на Шелковом пути Северо-Западного Китая». Тканевые антигены . 51 (3): 287–92. дои : 10.1111/j.1399-0039.1998.tb03104.x . ПМИД 9550330 .
- ^ Мизуки М, Оно С, Андо Х, Сато Т, Иманиши Т, Годобори Т, Исихара М, Ота М, Гэн З, Гэн Л, Ли Г, Кимура М, Иноко Х (1997). «Аллели класса II главного комплекса гистосовместимости у казахов и ханьцев на Шелковом пути северо-западного Китая». Тканевые антигены . 50 (5): 527–34. дои : 10.1111/j.1399-0039.1997.tb02909.x . ПМИД 9389328 .
- ^ «Обзор демографического исследования - Россия, Сибирь, Негидаль» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Уинук-Оол Т, Такезаки Н, Сукерник Р, Нагл С, Кляйн Дж (2002). «Происхождение и родство коренных сибирских популяций, выявленное по частотам генов HLA класса II». Хум Жене . 110 (3): 209–26. дои : 10.1007/s00439-001-0668-0 . ПМИД 11935333 . S2CID 20940785 .
- ^ Великович З., Делахант Б., Картер Дж. (2002). «Полиморфизмы HLA-DRB1 и HLA-DQB1 в популяциях островов Тихого океана». Тканевые антигены . 59 (5): 397–406. дои : 10.1034/j.1399-0039.2002.590506.x . ПМИД 12144623 .
- ^ «Обзор демографического исследования – Ненцы России» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Евсеева И, Спуркланд А, Торсби Е, Смердел А, Транебьерг Л, Болдырева М, Грудакова Е, Гускова И, Алексеев Л (2002). «HLA-профиль трех этносов, проживающих в Северо-Западном регионе России». Тканевые антигены . 59 (1): 38–43. дои : 10.1034/j.1399-0039.2002.590107.x . ПМИД 11972877 .
- ^ «Обзор демографического исследования - Россия, Мурманский саоми» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Евсеева И, Спуркланд А, Торсби Е, Смердел А, Транебьерг Л, Болдырева М, Грудакова Е, Гускова И, Алексеев Л (2002). «HLA-профиль трех этносов, проживающих в Северо-Западном регионе России». Тканевые антигены . 59 (1): 38–43. дои : 10.1034/j.1399-0039.2002.590107.x . ПМИД 11972877 .
- ^ «Обзор народонаселения - Архангельские поморы России» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Евсеева И, Спуркланд А, Торсби Е, Смердел А, Транебьерг Л, Болдырева М, Грудакова Е, Гускова И, Алексеев Л (2002). «HLA-профиль трех этносов, проживающих в Северо-Западном регионе России». Тканевые антигены . 59 (1): 38–43. дои : 10.1034/j.1399-0039.2002.590107.x . ПМИД 11972877 .
- ^ «Обзор демографического исследования - Швеция» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Кимиёси, Цудзи; Айзава М; Сасадзуки Т (1992). Материалы одиннадцатого международного семинара и конференции по гистосовместимости, состоявшихся в Иокохоме, Япония, 6–13 ноября 1991 г. Оксфорд: Издательство Оксфордского университета. ISBN 0-19-262390-7 .
- ^ «Обзор демографического исследования - европеоидная Англия» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Доэрти Д., Воган Р., Дональдсон П., Моват А. (1992). «Генотипирование HLA DQA, DQB и DRB с помощью анализа олигонуклеотидов: распределение аллелей и гаплотипов у британских европеоидов». Хум Иммунол . 34 (1): 53–63. дои : 10.1016/0198-8859(92)90085-2 . ПМИД 1399722 .
- ^ «Обзор демографического исследования – Финляндия» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. «12-я Международная конференция по гистосовместимости. Генетическое разнообразие HLA: функциональное и медицинское значение. Париж, Франция, 9–12 июня 1996 г. Тезисы». Хум Иммунол . 47 (1–2): 1–184. 1996. ПМИД 8909580 .
- ^ Бугаван Т., Клитц В., Блэр А., Эрлих Х. (2000). «Типирование HLA класса I с высоким разрешением в семьях CEPH: анализ неравновесия по сцеплению между локусами HLA». Тканевые антигены . 56 (5): 392–404. дои : 10.1034/j.1399-0039.2000.560502.x . ПМИД 11144287 .
- ^ «Обзор демографического исследования - Дания» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Кимиёси, Цудзи; Айзава М; Сасадзуки Т (1992). Материалы одиннадцатого международного семинара и конференции по гистосовместимости, состоявшихся в Иокохоме, Япония, 6–13 ноября 1991 г. Оксфорд: Издательство Оксфордского университета. ISBN 0-19-262390-7 .
- ^ Шиппер Р., Шредер Г., Д'Амаро Дж., Оудсхорн М. (1996). «Частоты гена и гаплотипов HLA у голландских доноров крови». Тканевые антигены . 48 (5): 562–74. дои : 10.1111/j.1399-0039.1996.tb02670.x . ПМИД 8988539 .
- ^ «Обзор демографического исследования - Северо-Западная Славянская Россия» . Частоты аллелей среди населения мира . Архивировано из оригинала 27 сентября 2007 г. Проверено 17 августа 2008 г. и 2. Капустин С, Лыщов А, Александрова Ю, Имянитов Е, Блинов М (1999). «Молекулярный полиморфизм HLA класса II у здоровых славян Северо-Запада России». Тканевые антигены . 54 (5): 517–20. дои : 10.1034/j.1399-0039.1999.540509.x . ПМИД 10599891 .
- ^ Перейти обратно: а б с д и ж г Кимиёси, Цудзи; Айзава М; Сасадзуки Т (1992). Материалы одиннадцатого международного семинара и конференции по гистосовместимости, состоявшихся в Иокохоме, Япония, 6–13 ноября 1991 г. Оксфорд: Издательство Оксфордского университета. ISBN 0-19-262390-7 .
- ^ Евсеева И, Спуркланд А, Торсби Е, Смердел А, Транебьерг Л, Болдырева М, Грудакова Е, Гускова И, Алексеев Л (2002). «HLA-профиль трех этносов, проживающих в Северо-Западном регионе России». Тканевые антигены . 59 (1): 38–43. дои : 10.1034/j.1399-0039.2002.590107.x . ПМИД 11972877 .
- ^ Санчес-Веласко П., Гомес-Касадо Э., Мартинес-Ласо Дж., Москосо Дж., Самора Дж., Лоуи Э., Сильвера С., Семборайн А., Лейва-Кобиан Ф., Арнаис-Виллена А. (2003). «Аллели HLA в изолированных популяциях Северной Испании: происхождение басков и древних иберийцев». Тканевые антигены . 61 (5): 384–92. дои : 10.1034/j.1399-0039.2003.00041.x . ПМИД 12753657 .
- ^ Валлури В., Валлуэй В., Мустафа М., Сантош А., Миддлтон Д., Альварес М., Алвалес М., Эль Хадж Э., Гумама О., Абдель-Варет Л., Абдель-Вайет Л. (2005). «Частоты фенотипов HLA-A, HLA-B, HLA-DR и HLA-DQ в населении Объединенных Арабских Эмиратов». Тканевые антигены . 66 (2): 107–13. дои : 10.1111/j.1399-0039.2005.00441.x . ПМИД 16029430 .
- ^ Санчес-Веласко П., Карадше Н., Гарсиа-Мартин А., Руис де Алегрия К., Лейва-Кобиан Ф. (2001). «Молекулярный анализ частот и гаплотипов аллелей HLA у иорданцев и сравнение с другими родственными популяциями». Хум Иммунол . 62 (9): 901–9. дои : 10.1016/S0198-8859(01)00289-0 . ПМИД 11543892 .
- ^ Перейти обратно: а б Форт М., де Стефано Дж., Камбон-Томсен А., Хиральдо-Альварес П., Дюгужон Дж., Охайон Э., Скано Г., Аббал М. (1998). «Частоты аллелей и гаплотипов HLA класса II в эфиопских популяциях амхара и оромо» Тканевые антигены . 51 (4 Пт. 1): 327–36. дои : 10.1111/j.1399-0039.1998.tb02971.x . ПМИД 9583804 .
- ^ Гомес-Касадо Э, дель Мораль П, Мартинес-Ласо Х, Гарсия-Гомес А, Альенде Л, Сильвера-Редондо С, Лонгас Х, Гонсалес-Хевилла М, Кандил М, Самора Х, Арнаис-Вильена А (2000). «Гены HLA у арабоязычных марокканцев: тесное родство с берберами и иберийцами». Тканевые антигены . 55 (3): 239–49. дои : 10.1034/j.1399-0039.2000.550307.x . ПМИД 10777099 .
- ^ Айед К., Айед-Жендуби С., Сфар И., Лабонн М., Гебурер Л. (2004). «Фенотипические, генные и гаплотипические частоты HLA класса I и HLA класса II у тунисцев с использованием данных молекулярного типирования». Тканевые антигены . 64 (4): 520–32. дои : 10.1111/j.1399-0039.2004.00313.x . ПМИД 15361135 .
- ^ Перейти обратно: а б Ренкин Дж., Санчес-Масас А., Галле Л., Ривалланд С., Джагер Г., Мбайо К., Бьянки Ф., Каплан С. (2001). «Полиморфизм HLA класса II у ака-пигмеев и конголезских банту и переоценка африканского разнообразия HLA-DRB1». Тканевые антигены . 58 (4): 211–22. дои : 10.1034/j.1399-0039.2001.580401.x . ПМИД 11782272 .
- ^ Пимтанотаи Н., Херли С., Леке Р., Клитц В., Джонсон А. (2001). «Полиморфизм HLA-DR и -DQ в Камеруне». Тканевые антигены . 58 (1): 1–8. дои : 10.1034/j.1399-0039.2001.580101.x . ПМИД 11580849 .
- ^ Шнитгер Л., Мэй Дж., Лелигер С., Галлин М., Эрттманн К., Бинцле У., Кремснер П., Мейер С. (1997). «Разнообразие гаплотипов HLA DRB1-DQA1-DQB1 в двух африканских популяциях». Тканевые антигены . 50 (5): 546–51. дои : 10.1111/j.1399-0039.1997.tb02911.x . ПМИД 9389330 .
- ^ де Поль Р., Гарсиа-Пачеко Х., Вилчес К., Морено М., Санс Л., Рементари М., Бридж С., Крейслер М. (1997). «Распределение аллелей HLA класса I и класса II в популяции буби с острова Биоко».
(Экваториальная Гвинея).". Тканевые антигены . 50 (6): 593–601. doi : 10.1111/j.1399-0039.1997.tb02917.x . PMID 9458112 . - ^ Пак Ю, Тейт Б, Кавасаки Э, Роули М, Маккей I (2004). «Более тесная связь гуморальной аутореактивности IA-2 с HLA DR3/4, чем DQB1*0201/*0302, у корейских пациентов с СД1». Анналы Нью-Йоркской академии наук . 1037 (1): 104–9. Бибкод : 2004NYASA1037..104P . дои : 10.1196/анналы.1337.015 . ПМИД 15699500 . S2CID 38178367 .
- ^ Герман Р., Турпейнен Х., Лайне А., Вейола Р., Книп М., Симелл О., Сипиля И., Акерблом Х., Илонен Дж. (2003). «Генетические детерминанты детского диабета 1 типа, кодируемые HLA DR-DQ, в Финляндии: анализ 622 нуклеарных семей». Тканевые антигены . 62 (2): 162–9. дои : 10.1034/j.1399-0039.2003.00071.x . ПМИД 12889996 .
Внешние ссылки
[ редактировать ]Целиакия
- Целиакия в Великобритании. Архивировано 27 мая 2006 г. в Wayback Machine (благотворительная организация).
- Фонд целиакии. Архивировано 23 октября 2006 г. в Wayback Machine (США).
- Ассоциация целиакии (США)
- Национальный информационный центр по пищеварительным заболеваниям - страница о целиакии
- Национальный фонд осведомленности о целиакии (США)
- Центр исследований целиакии Университета Мэриленда
Диабет 1 типа