Морская звезда с терновым венцом
| Морская звезда с терновым венцом | |
|---|---|
 | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Иглокожие |
| Сорт: | астероид |
| Заказ: | охранять |
| Семья: | Акантастериды |
| Род: | Акантастер |
| Разновидность: | А. плакать |
| Биномиальное имя | |
| Акантастер плачи | |
Морская звезда с терновым венцом (часто сокращенно COTS), [1] Acanthaster planci — крупная морская звезда , питающаяся твердыми или каменистыми коралловыми полипами ( Scleractinia ). Морская звезда с терновым венцом получила свое название из-за ядовитых шиповидных шипов, покрывающих ее верхнюю поверхность, напоминающих библейский терновый венец . Это одна из крупнейших морских звезд в мире.
A. planci имеет очень широкое распространение в Индо-Тихоокеанском регионе . Вероятно, он наиболее распространен в Австралии, но может встречаться в тропических и субтропических широтах от Красного моря и восточноафриканского побережья через Индийский океан и через Тихий океан до западного побережья Центральной Америки. Это происходит там, где в регионе встречаются коралловые рифы или сообщества твердых кораллов.
Описание
[ редактировать ]
Форма тела морской звезды с терновым венцом в основном такая же, как у типичной морской звезды , с центральным диском и расходящимися лучами. Однако к его особым чертам относятся дискообразная форма, многорукость, гибкость, цепкость , сильные шипы и большое соотношение поверхности желудка к массе тела. [2] Его цепкая способность обусловлена двумя рядами многочисленных трубчатых ножек , доходящих до кончиков каждой руки. Будучи многоруким, он утратил пятикратную симметрию ( пентамерию ), типичную для морских звезд, хотя и начинает свой жизненный цикл с этой симметрией . Животное обладает настоящим образообразующим зрением. [3]
Размер взрослой морской звезды с терновым венцом обычно составляет от 25 до 35 см (от 10 до 14 дюймов). [4] У них до 21 руки. [3] [5] Хотя тело тернового венца имеет жесткий вид, оно способно сгибаться и поворачиваться, повторяя контуры кораллов, которыми оно питается. На нижней стороне каждой руки имеется ряд плотно прилегающих друг к другу пластинок, которые образуют бороздку и рядами доходят до рта. [6] В зависимости от диеты или географического региона особи могут быть фиолетовыми, пурпурно-синими, красновато-серыми или коричневыми с красными кончиками шипов или зелеными с желтыми кончиками шипов. [7]
Длинные острые шипы по бокам рук морской звезды и верхней (аборальной) поверхности напоминают шипы и создают форму короны, что и дало существу такое название. Колючки могут иметь длину от 4 до 5 см, они жесткие, очень острые и легко прокалывают мягкие поверхности. [8] Несмотря на совокупность острых шипов на аборальной поверхности и тупых шипов на ротовой поверхности, общая поверхность тела морской звезды с терновым венцом перепончатая и мягкая. Когда морскую звезду вынимают из воды, поверхность тела разрывается, и жидкость организма вытекает, поэтому тело сжимается и сплющивается. Колючки также изгибаются и уплощаются. Они восстанавливают свою форму при повторном погружении, если они еще живы. [9]
Онлайн-база данных модельных организмов
[ редактировать ]Echinobase — это база данных модельных организмов A. planci и ряда других иглокожих.
Таксономия
[ редактировать ]Семья
[ редактировать ]Семейство Acanthasteridae монородно ; его положение среди астероидов неустойчиво. Обычно он считается четко изолированным таксоном . Недавно палеонтолог Дэниел Блейк на основе сравнительных морфологических исследований A. planci пришел к выводу , что он имеет большое сходство с различными представителями Oreasteridae. Он перенес Acanthasteridae из Spinulosida в Valvatida и определил для них положение рядом с Oreasteridae, от которых они, по-видимому, произошли. [10] Он предположил, что акантастра морфология , возможно, развивалась в связи с его передвижением по неровным коралловым поверхностям в средах с более высокой энергией. Однако существует сложность в том, что Acanthaster не является моновидовым родом, и любое рассмотрение этого рода должно также учитывать другой вид, Acanthaster brevispinus , который обитает в совершенно другой среде. A. brevispinus живет на мягких субстратах, иногда, возможно, зарытых в субстрате, как и другие морские звезды, обитающие в мягком субстрате, на умеренных глубинах, где предположительно поверхность ровная и волнение незначительное.
Род и вид
[ редактировать ]
A. planci имеет долгую историю в научной литературе с большой путаницей в родовых и видовых названиях с самого начала и длинным списком сложных синонимов . [11] Георг Эберхард Румфиус впервые описал его в 1705 году, назвав Stella marina quindecium radiotorum . Позже Карл Линней описал его как Asterias planci на основе иллюстрации Планка и Гуальтьери (1743), когда он представил свою систему биномиальной номенклатуры . Типовые экземпляры не известны; экземпляр, описанный Планком и Гуальтьери (1743 г.), больше не сохранился.
Последующие родовые названия, используемые для морских звезд с терновым венцом, включали Stellonia , Echinaster и Echinites , прежде чем они обосновались на Акантастере ( Gervais 1841). Конкретные названия включали echintes , Solaris , mauritensis , ellisii и ellisii pseudoplanci (с подвидами). Большинство этих названий возникли из-за путаницы в исторической литературе, но Acanthaster ellisii стал использоваться для обозначения характерной морской звезды в восточной части Тихого океана в Калифорнийском заливе .
восточной части Тихого океана Акантастер очень характерен (см. изображение справа) своим довольно «пухлым» телом, большим соотношением диаметра диска к общему диаметру и короткими тупыми шипами.
Генетические исследования
[ редактировать ]Нишида и Лукас исследовали генетические вариации в 14 аллозимных локусах из 10 популяционных образцов A. planci с помощью электрофореза в крахмальном геле. Образцы были взяты из мест по всему Тихому океану: архипелаг Рюкю (четыре места), Микронезия (два места), а также образцы из одного места на Большом Барьерном рифе, Фиджи, Гавайях и Калифорнийском заливе. Для сравнения была использована выборка из 10 экземпляров A. brevispinus из района Большого Барьерного рифа. Значительная генетическая дифференциация наблюдалась между популяциями A. brevispinus и A. planci (D = 0,20 +/- 0,02) (D — генетическое расстояние ). генетические различия между географическими популяциями A. planci Однако были небольшими (D = 0,03 +/- 0,00; Fsr = 0,07 + 0,02) (Fsr — стандартизированная генетическая дисперсия для каждого полиморфного локуса), несмотря на большие расстояния, разделяющие их. Наблюдалась положительная корреляция между степенью генетической дифференциации и географическим расстоянием, что позволяет предположить, что генетическая однородность среди популяций A. planci обусловлена поток генов путем рассеивания планктонных личинок. Влияние расстояния на генетическую дифференциацию, скорее всего, отражает снижение уровня успешного расселения личинок на большие расстояния. Учитывая уровень макрогеографической однородности, между соседними популяциями, разделенными примерно 10 км, наблюдались значительные различия в частоте аллелей. Население Гавайев наиболее отличалось от других популяций. Рассмотрение морфологически отличительного восточно-тихоокеанского Acanthaster как отдельного вида, A. ellisii , не подтверждается этими данными. Отсутствие уникальных аллелей в популяциях центральной (Гавайи) и восточной части Тихого океана (Калифорнийский залив) позволяет предположить, что они произошли от популяций западной части Тихого океана.
Более подробная информация о генетическом родстве A. planci и A. brevispinus представлена в статье о последнем виде. Это явно родственные виды , и A. planci , специализированный вид, питающийся кораллами, предположительно произошел от A. brevispinus , менее специализированного обитателя мягкого дна. [12]
В очень обширном географическом исследовании Бензи изучил вариации аллозимных локусов в 20 популяциях A. planci в Тихом и Индийском океанах. [13] Самым поразительным результатом стал весьма заметный разрыв между популяциями Индийского и Тихого океана. Однако те, что находились у северной части Западной Австралии, имели сильное родство с Тихим океаном. За исключением очень сильной связи популяций южной Японии с популяциями Большого Барьерного рифа, закономерности вариаций внутри регионов соответствовали изоляции на расстоянии. Опять же, очевидна тенденция снижения уровня успешного распространения личинок на большие расстояния. Бензи предполагает, что расхождение между популяциями Индийского и Тихого океанов началось как минимум 1,6 миллиона лет назад и, вероятно, отражает реакцию на изменения климата и уровня моря.
Более недавнее комплексное географическое исследование A. planci, проведенное Vogler et al. , используя анализ ДНК (один митохондриальный ген ), предполагает, что на самом деле это видовой комплекс, состоящий из четырех видов или клад . [14] Четыре загадочных вида / клады определены географически: Северный Индийский океан, южная часть Индийского океана, Красное море и Тихий океан. Эти молекулярные данные позволяют предположить, что виды/клады разошлись 1,95 и 3,65 миллиона лет назад. (Дивергенция A. planci и A. brevispinus не включена в эту временную шкалу.) Авторы предполагают, что различия между четырьмя предполагаемыми видами в поведении, диете и среде обитания могут быть важны для разработки соответствующих стратегий сохранения рифов. [15]
Однако существуют проблемы с этим предложением загадочного видообразования ( загадочные виды ). Основание этих данных на данных одного митохондриального гена ( мтДНК ), однако, является только одним источником информации о статусе таксонов, и использование одного гена мтДНК в качестве единственного критерия для идентификации вида оспаривается. [16] [17] Следует также учитывать данные по аллозимам. Три местности, отобранные Vogler et al. представляют особый интерес; Было обнаружено, что в Палау-Себибу, UEA и Омане есть симпатичные две клады/родственные виды . Это важно для изучения природы сосуществования и барьеров на пути проникновения генетического материала. A. planci как таксон является универсальным, являясь одним из наиболее распространенных крупных коралловых хищников на коралловых рифах, питаясь практически всеми видами твердых кораллов, размножаясь летом без закономерного нереста и часто участвуя в массовых нерестах нескольких видов. [18] и высвобождение огромного количества гамет, которые вызывают нерест у других особей. Зарождение двух видов/клад A. planci в симпатрии без конкуренции местообитаний и интрогрессии генетического материала, особенно последнего, очень сложно.
Биология
[ редактировать ]Токсины
[ редактировать ] Сломанные и регенерирующие шипы
Сломанные и регенерирующие шипы Опухла правая рука после прокола.
Опухла правая рука после прокола. Вспенивание воды, содержащей A. planci
Вспенивание воды, содержащей A. planci С морской звездой обращаются так, чтобы не повредить ее (шипы на нижней стороне тупые).
С морской звездой обращаются так, чтобы не повредить ее (шипы на нижней стороне тупые).




Для морских звезд характерно наличие сапонинов, в тканях известных как астеросапонины. Они содержат смесь этих сапонинов, и было проведено не менее 15 химических исследований с целью охарактеризовать эти сапонины. [2] Они обладают моющими свойствами, а содержание морских звезд в ограниченном объеме воды с аэрацией приводит к образованию большого количества пены на поверхности.
У A. planci нет механизма для введения токсина, но когда шипы прокалывают ткани хищника или неосторожного человека, ткань, содержащая сапонины, теряется в ране. У людей это немедленно вызывает острую, жгучую боль, которая может длиться несколько часов, постоянное кровотечение из-за гемолитического действия сапонинов, а также тошноту и отек тканей, которые могут сохраняться в течение недели и более. [19] Хрупкие шипы также могут сломаться и застрять в тканях, где их придется удалить хирургическим путем.
Сапонины, по-видимому, присутствуют на протяжении всего жизненного цикла морской звезды с терновым венцом. Сапонины в яйцах аналогичны сапонинам в тканях взрослых особей и, предположительно, передаются личинкам. [20] Ротовое поведение хищников молоди морских звезд с отторжением позволяет предположить, что молодь содержит сапонины.
Поведение
[ редактировать ] Молодые особи спрятаны под коралловыми обломками
Молодые особи спрятаны под коралловыми обломками Две морские звезды питаются кораллом, оставляя белые шрамы от еды
Две морские звезды питаются кораллом, оставляя белые шрамы от еды Питается ветвящимся Acropora. кораллом
Питается ветвящимся Acropora. кораллом Морские звезды «конкурируют» за оставшиеся живые кораллы
Морские звезды «конкурируют» за оставшиеся живые кораллы




Взрослая особь тернового венца — кораллоядный хищник , который обычно охотится на полипы рифовых кораллов . [21] Он забирается на участок живой колонии кораллов, используя большое количество трубчатых ножек, которые лежат в отчетливых амбулакральных бороздках на поверхности рта. [22] Он плотно прилегает к поверхности коралла, даже к сложным поверхностям ветвящихся кораллов. Затем он вытягивает желудок через рот на поверхность практически до своего диаметра. Поверхность желудка выделяет пищеварительные ферменты, которые позволяют морской звезде усваивать питательные вещества из разжиженной ткани коралла. В результате остается белый шрам на скелете коралла, который быстро заражается нитчатыми водорослями. [23] Отдельная морская звезда может потреблять до 6 квадратных метров (65 квадратных футов) живого кораллового рифа в год. [24] В ходе исследования скорости питания на двух коралловых рифах в центральной части Большого Барьерного рифа крупные морские звезды (диаметром 40 см (16 дюймов) и больше) убили около 61 см. 2 (9 дюймов 2 )/день зимой и от 357 до 478 см. 2 (от 55 до 74 дюймов 2 ) в день летом. Меньшая морская звезда, 20–39 см (8–15 дюймов), убитая от 155 до 234 см. 2 (от 24 до 36 дюймов 2 ) в день в соответствующие сезоны. Площадь гибели крупной морской звезды эквивалентна примерно 10 м2. 2 (108 кв. футов) по этим наблюдениям. [25] Различия в скорости питания и передвижения между летом и зимой отражают тот факт, что терновый венец, как и все морские беспозвоночные , является пойкилотермным животным , температура тела которого и скорость обмена веществ напрямую зависят от температуры окружающей воды. В тропических коралловых рифах особи тернового венца достигают средней скорости передвижения 35 см/мин (14 дюймов/мин). [26] это объясняет, как вспышки могут повредить большие площади рифов за относительно короткие периоды времени.
Морские звезды отдают предпочтение твердым кораллам, которыми они питаются. Они, как правило, питаются ветвящимися и табловидными кораллами, такими как виды Acropora , а не более округлыми кораллами с меньшей открытой площадью поверхности, такими как виды Porites . [27] Избегание поритов двустворчатых моллюсков и многощетинковых червей, которые отпугивают морских звезд. и некоторых других кораллов может также быть связано с присутствием на поверхности кораллов [28] Точно так же некоторые симбионты , такие как маленькие крабы, живущие внутри сложных структур ветвящихся кораллов, могут отпугивать морскую звезду, когда она пытается распространить свой желудок по поверхности коралла. [29]
В рифовых районах с низкой плотностью твердых кораллов, что отражает характер рифового сообщества или из-за питания терновым венцом с высокой плотностью, морские звезды могут питаться мягкими кораллами ( Alcyonacea ). [30]
В течение первых двух лет жизни морские звезды ведут себя загадочно, выходя ночью на корм. Обычно они остаются такими и взрослыми, когда остаются в одиночестве. Единственным свидетельством присутствия спрятавшейся особи могут быть белые шрамы от питания на соседних кораллах. Однако их поведение меняется при двух обстоятельствах:
- Во время сезона размножения, который обычно приходится на начало и середину лета, морские звезды могут собираться высоко на рифе и синхронно выпускать гаметы, чтобы достичь высокого уровня оплодотворения икры . [31] Этот образец синхронного нереста вовсе не уникален, но он очень распространен среди морских беспозвоночных, которые не совокупляются . Одиночный нерест не дает возможности для оплодотворения яиц и отходов гамет , и существуют доказательства существования нерестового феромона , который заставляет морскую звезду синхронно агрегировать и высвобождать гаметы. [32]
- Когда морские звезды имеют высокую плотность, они могут перемещаться днем и ночью, конкурируя за живые кораллы.
Хищники
[ редактировать ]
Удлиненные острые шипы, покрывающие почти всю верхнюю поверхность тернового венца, служат механической защитой от крупных хищников. У него также есть химическая защита. Сапонины предположительно служат раздражителем при прокалывании хищника точно так же, как они действуют при прокалывании кожи человека. Сапонины имеют неприятный вкус. Исследование по проверке уровня хищничества молоди Acanthaster spp. с помощью соответствующих видов рыб обнаружили, что морскую звезду часто пробовали в рот, пробовали на вкус и отвергали. [33] Такая защита делает его непривлекательной целью для хищников кораллового сообщества. Однако, несмотря на это, популяции акантастера обычно состоят из определенной части особей с регенерирующими руками.
Сообщается, что около 11 видов время от времени охотятся на неповрежденных и здоровых взрослых особей A. planci . Все они являются универсальными кормушками, но ни один из них, похоже, не отдает предпочтение морской звезде в качестве источника пищи. [34] Однако это число, вероятно, меньше, поскольку некоторые из этих предполагаемых хищников достоверно не наблюдались в полевых условиях. Некоторые из свидетелей:
- Было замечено, что один вид иглобрюха и два спинорога питаются морскими звездами с терновым венцом в Красном море, и хотя они могут оказывать некоторое влияние на популяцию A. planci , свидетельств систематического хищничества не существует. [35] В водах Индо-Тихоокеанского региона белопятнистые иглобрюхи и спинорог Титан поедают эту морскую звезду. также было обнаружено, что [36]
- Труба Тритона , очень крупный брюхоногий моллюск, является известным хищником акантастера в некоторых частях ареала морской звезды. Тритон описывался как разрывающий морскую звезду на куски своей радулой, похожей на напильник . [37]
- маленькая окрашенная креветка Hymenocera picta Было обнаружено, что , основной хищник морских звезд, охотится на A. planci . в некоторых местах [38] червь Многощетинковый striata Pherecardia . питался морскими звездами вместе с креветками на коралловом рифе в восточной части Тихого океана [39] Около 0,6% морских звезд в популяции рифа подверглись нападению как креветок, так и многощетинковых червей, в результате чего морские звезды погибли примерно за неделю. Глинн предположил, что это привело к балансу между смертностью и пополнением в этой популяции, что привело к относительно стабильной популяции морских звезд. [38]
- Поскольку P. striata может атаковать только поврежденный A. planci и вызвать его смерть, его можно рассматривать как «нетерпеливого падальщика», а не хищника. [40] В отличие от хищников, мертвые и изувеченные взрослые особи A. planci привлекают внимание ряда падальщиков . Глинн перечисляет двух многощетинковых червей, рака-отшельника , морского ежа и семь видов мелких рифовых рыб. [39] Судя по всему, они способны переносить неприятные сапонины для легкой еды.
- Было замечено , что большое, похожее на полипа существо из рода книдарий Pseudocorynactis напало, а затем полностью проглотило морскую звезду с терновым венцом такого же размера. [41] Дальнейшие исследования показали, что этот полип способен полностью проглотить образец тернового венца диаметром до 34 см (13 дюймов). [42]
Жизненный цикл
[ редактировать ]Гаметы и эмбрионы
[ редактировать ] Окрашенный поперечный разрез зрелой завязи, полной яйцеклеток.
Окрашенный поперечный разрез зрелой завязи, полной яйцеклеток. Окрашенный поперечный срез яичка (сперматозоиды синие)
Окрашенный поперечный срез яичка (сперматозоиды синие) Нерест
Нерест Первые деления клеток внутри оплодотворенной яйцеклетки диаметром около 0,3 мм.
Первые деления клеток внутри оплодотворенной яйцеклетки диаметром около 0,3 мм. Свободноживущая стадия гаструлы, длина около 0,5 мм.
Свободноживущая стадия гаструлы, длина около 0,5 мм.





Гонады увеличиваются в размерах по мере того, как животные становятся половозрелыми, а по мере взросления заполняют руки и доходят до области диска. Спелые яичники и семенники легко различимы: первые более желтые и имеют более крупные доли. В разрезе они очень разные: яичники плотно заполнены яйцеклетками, богатыми питательными веществами (см. яйцеклетку и фотографию), а семенники густо заполнены сперматозоидами , которые состоят лишь из ядра и жгутика. Плодовитость самок морской звезды с терновым венцом зависит от размера: крупные морские звезды тратят пропорционально больше энергии на производство яйцеклеток, так что: [43]
- Самка диаметром 200 мм производит 0,5–2,5 миллиона яиц, что составляет 2–8% ее влажного веса.
- Самка диаметром 300 мм производит 6,5–14 миллионов яиц, что составляет 9–14% ее влажного веса.
- Самка диаметром 400 мм производит 47–53 миллиона яиц, что составляет 20–25% ее влажного веса.
В коралловых рифах на Филиппинах были обнаружены самки с гонадосоматическим индексом (отношение массы гонад к массе тела) до 22%. [44] что подчеркивает высокую плодовитость этой морской звезды. Бэбкок и др. (1993) [45] отслеживали изменения в плодовитости и плодовитости (коэффициенте оплодотворения) в течение сезона нереста морской звезды с терновым венцом на рифе Дэвис, центральная часть Большого Барьерного рифа, с 1990 по 1992 год. Нерест морских звезд (фотография) наблюдался с декабря по январь ( с начала до середины лета) в этом регионе, причем большинство наблюдений приходится на январь. Однако как гонадосоматический индекс, так и плодовитость достигли пика рано и снизились до низкого уровня к концу января, что указывает на то, что наиболее успешные репродуктивные события произошли в начале сезона нереста. Однако в коралловых рифах Северного полушария популяции тернового венца размножаются в апреле и мае. [44] а также наблюдался нерест в Сиамском заливе в сентябре. [46] Высокие показатели оплодотворения икры могут быть достигнуты за счет быстрого и синхронизированного нереста (см. выше в разделе «Поведение»).
Эмбриональное развитие начинается примерно через 1,5 часа после оплодотворения, с ранних делений клеток ( дробления ) (фотография). К 8–9 часам он достигает стадии 64 клеток.
Некоторые молекулярные и гистологические данные позволяют предположить наличие гермафродитизма у Acanthaster cf. Солярис . [47]
Личиночные стадии
[ редактировать ]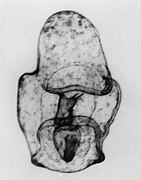 Личинка бипиннарии
Личинка бипиннарии СЭМ личинки бипиннарии
СЭМ личинки бипиннарии Личинка брахиолярии
Личинка брахиолярии Поздние брахиолярии с зачатком морской звезды
Поздние брахиолярии с зачатком морской звезды СЭМ брахиолярные руки
СЭМ брахиолярные руки





К первому дню эмбрион вылупился на стадии мерцательной гаструлы (фотография). Ко второму дню кишечник готов, и личинка теперь известна как бипиннария . Он имеет ресничные полосы вдоль тела и использует их для плавания и фильтрации микроскопических частиц, особенно одноклеточных зеленых жгутиковых ( фитопланктона ). На сканирующей электронной микрофотографии (СЭМ) четко видны сложные ресничные полосы бипиннаральной личинки. К 5-му дню это ранняя брахиолярная личинка. Руки бипиннарии еще больше удлинились, спереди появились два культевидных выступа (на фотографии не видны), а в задней части личинки развиваются структуры. У поздней брахиолярной личинки (11-й день) личиночные руки удлинены, и спереди встречаются три характерных плеча с небольшими структурами на внутренней поверхности. На этом этапе личинка была практически прозрачной, но задняя часть теперь непрозрачна из-за начального развития морской звезды. Поздняя брахиолярия 1,0-1,5 мм. Он имеет тенденцию опускаться на дно и тестировать субстрат своими брахиолярными руками, в том числе сгибать переднюю часть тела, чтобы сориентировать брахиолярные плечи против субстрата.
Это описание и оценка оптимальной скорости развития основаны на ранних исследованиях в лаборатории в оптимальных условиях. [48] [49] [50] Однако неудивительно, что существуют большие различия в скорости роста и выживаемости в различных условиях окружающей среды (см. Причины популяционных вспышек).
Метаморфоза, развитие и рост
[ редактировать ] Расселение брахиолярной личинки
Расселение брахиолярной личинки Молодь пятирукой морской звезды сразу после метаморфоза.
Молодь пятирукой морской звезды сразу после метаморфоза. Ранняя молодь морских звезд, питающаяся коралловыми водорослями (оставляющая после себя белые шрамы от питания)
Ранняя молодь морских звезд, питающаяся коралловыми водорослями (оставляющая после себя белые шрамы от питания) Очень молодая молодь, питающаяся кораллами, с полным набором оружия и мадрепоритами.
Очень молодая молодь, питающаяся кораллами, с полным набором оружия и мадрепоритами. Молодая молодь, питающаяся кораллами
Молодая молодь, питающаяся кораллами





Поздние брахиолярии ищут субстрат своими руками и, когда им предлагают выбор субстрата, имеют тенденцию селиться на кораллиновых водорослях, которыми они впоследствии питаются. В классической модели иглокожих двусторонне-симметричная личинка заменяется пентамерно-симметричной стадией метаморфоза, причем ось тела последней не имеет никакого отношения к оси личинки. Таким образом, новометаморфизованные морские звезды пятирукие и имеют диаметр 0,4–1,0 мм. (Обратите внимание на размер ножек трубки относительно размера животного.) Они питаются тонкими слоями покрытия из твердых, покрывающих корку водорослей (кораллиновых водорослей) на нижней стороне мертвых коралловых обломков и других скрытых поверхностях. Они вытягивают желудок над поверхностью покрывающих корку водорослей и переваривают ткани, как при питании более крупных морских звезд с терновым венцом на твердых кораллах. Живая ткань покрывающих водорослей имеет цвет примерно от розового до темно-красного, и питание этой ранней молодью приводит к образованию белых рубцов на поверхности водорослей. В течение следующих месяцев молодые особи растут и добавляют руки и связанные с ними мадрепориты по схеме, описанной Ямагучи. [50] до достижения численности взрослых особей через 5–7 месяцев после метаморфоза. Два твердых коралла с мелкими полипами, Pocillopora damicornis и Acropora acunimata , были помещены в аквариум с коралловыми водорослями, и примерно в то время, когда молодые морские звезды достигли полного количества рук, они начали питаться кораллами. [9]
Молодь A. planci , достигшая стадии питания кораллами, затем в течение нескольких лет выращивалась в той же большой замкнутой системе с морской водой, которая использовалась для ранней молоди. Их переместили в резервуары большего размера и продолжали снабжать кораллами, чтобы еда не была ограничивающим фактором для скорости роста. Кривые роста размера в зависимости от возраста имели сигмоидальную форму , как это наблюдается у большинства морских беспозвоночных. [51] Начальный период относительно медленного роста произошел, когда морские звезды питались коралловыми водорослями. За этим последовала фаза быстрого роста, которая привела к половой зрелости в конце второго года жизни. На этом этапе морские звезды имели диаметр около 200 мм. Они продолжали быстро расти и составили около 300, а через 4 года имели тенденцию к снижению. Развитие гонад было выше в третий и последующие годы, чем в 2 года, и стал очевиден сезонный характер гаметогенеза и нереста, при этом температура воды была единственным заметным признаком в комнатном аквариуме. Большинство экземпляров A. planci погибло от «старости» в течение 5,0–7,5 лет, т. е. плохо питалось и мельчало.
Полевые наблюдения за жизненным циклом
[ редактировать ]Приведенные выше данные получены в результате лабораторных исследований A. planci , которые гораздо легче получить, чем эквивалентные данные, полученные в полевых условиях. Однако лабораторные наблюдения согласуются с ограниченными полевыми наблюдениями за жизненным циклом.
Как и в лабораторных исследованиях, где было обнаружено, что личинки A. planci выбирают кораллиновые водоросли для поселения, ранние молодые особи (<20 мм в диаметре) были обнаружены на сублиторальных коралловых водорослях ( Porolithon onkodes ) на наветренной передней части рифа Сува (Фиджи). [52] Молодь была обнаружена в различных местах обитания, где она была сильно скрыта - под коралловыми блоками и щебнем в зоне валунов обнаженной передней части рифа, на мертвых основаниях видов Acropora в более защищенных местах, в узких пространствах внутри гребня рифа и на предрифовом склоне до глубины 8 м.
Было обнаружено, что темпы роста на рифе Сува составляют 2,6, 16,7 и 5,3 мм/месяц увеличения диаметра до кормления кораллов, на ранней стадии кормления кораллов и на стадии взрослой особи соответственно. [52] Это согласуется с сигмоидальной зависимостью размера от возраста, наблюдаемой в лабораторных исследованиях, т.е. медленным начальным ростом, фазой очень быстрого роста, начинающейся при кормлении кораллов и спадающей после того, как морская звезда достигает половой зрелости. В рифах Филиппин самки и самцы созревали при длине 13 и 16 см соответственно. [44]
Пень [53] идентифицировал полосы на верхних поверхностных шипах A. planci и приписал их годовым полосам роста. Он не сообщил о темпах роста, основанных на этих определениях возраста, а также данных о маркировке и повторной поимке, но сообщил, что полосы роста выявили морские звезды в возрасте 12+ лет: намного старше, чем те, которые стали «старческими» и умерли в лаборатории.
В небольшом количестве полевых исследований уровень смертности молоди A. planci оказался очень высоким, например, 6,5% в день для месячных и 0,45% в день для 7-месячных детей. Большая часть смертности приходится на хищников, таких как мелкие крабы, которые обитают в субстрате и на нем вместе с молодью. [54] Однако возможно, что эти показатели могут не отражать смертность в диапазоне местообитаний, занимаемых мелкой молодью.
Экология
[ редактировать ]Экологическое воздействие на рифы
[ редактировать ]


A. planci — один из наиболее эффективных хищников склерактиновых кораллов (каменистых или твердых кораллов). Большинство организмов, питающихся кораллами, вызывают только потерю тканей или локальные повреждения, но взрослые особи A. planci могут уничтожать целые колонии кораллов. [55]
Общественная тревога по поводу новостей о высокой плотности A. planci на Большом Барьерном рифе нашла отражение во многих газетных сообщениях. [ нужна ссылка ] и такие публикации, как «Реквием по рифу» , в которых также предполагалось, что существовало сокрытие масштабов ущерба. [56] Возникла популярная идея, что кораллы, а вместе с ними и целые рифы разрушаются морскими звездами. Фактически, как описано выше, морские звезды охотятся на кораллы, переваривая поверхность живых тканей скелетов кораллов. Эти скелеты сохраняются вместе с массой коралловых водорослей, необходимых для целостности рифов. Первоначальное изменение (эффект первого порядка) заключается в утрате покрова живой коралловой ткани. [ нужна ссылка ]
A. planci является компонентом фауны большинства коралловых рифов, и воздействие популяций A. planci на коралловые рифы во многом зависит от плотности популяции. При низкой плотности (от 1 до, возможно, 30 особей на гектар) скорость, с которой морские звезды поедают кораллы, меньше скорости роста кораллов, т. е. площадь поверхности живых кораллов увеличивается. Однако морские звезды могут влиять на структуру кораллового сообщества. Поскольку морские звезды не питаются без разбора, они могут привести к тому, что распределение видов кораллов и размеров колоний будет отличаться от того, которое наблюдалось бы без них. Это становится очевидным при сравнении коралловых рифов, где A. planci не обнаружен, с более типичными рифами, где обитает A. planci . [40]
Некоторые экологи предполагают, что морские звезды играют важную и активную роль в поддержании биоразнообразия коралловых рифов , обеспечивая экологическую преемственность . До того, как перенаселение стало серьезной проблемой, терновый венец не позволял быстрорастущим кораллам вытеснить медленнорастущие разновидности кораллов. [57]
При высокой плотности (вспышки эпидемий, эпидемии), которую можно определить как ситуацию, когда морских звезд становится слишком много для обеспечения кораллов пищей, коралловый покров приходит в упадок. Морские звезды должны расширить свой рацион за счет предпочтительных видов, размера и формы колонии. Морские звезды часто объединяются во время кормления, даже при низкой плотности, но при высокой плотности очищенные коралловые участки становятся почти или полностью сплошными.Эффекты второго порядка существуют для этих больших площадей кораллов, на которых охотятся:
- Голые коралловые скелеты быстро заселяются нитчатыми водорослями.
- Большие заросли оленевидных кораллов вида Acropora могут обрушиться и превратиться в руины, что уменьшит топографическую сложность рифа.
- Иногда поверхности, на которых охотятся, дополнительно заселяются макроводорослями, мягкими кораллами и губками. Они имеют тенденцию занимать поверхность рифов на длительное время в качестве альтернативы сообществам твердых кораллов; однажды установившись, они ограничивают пополнение личинками твердых кораллов.
С эстетической точки зрения во всех вышеперечисленных случаях поверхность рифа не так привлекательна, как поверхность живого коралла, но и никак не мертва.
Эффект третьего порядка может возникнуть в результате инвазии нитчатых водорослей. Животные, которые прямо или косвенно зависят от твердых кораллов, например, в качестве убежища и пищи, должны проиграть, а травоядные и менее специализированные кормушки выиграют. Вероятно, это будет наиболее заметно в ихтиофауне, и долгосрочные исследования сообществ коралловых рифовых рыб подтверждают это ожидание. [58] [59]
Популяционные вспышки
[ редактировать ]
Было доказано, что большие популяции морских звезд с терновым венцом (иногда эмоционально называемые чумой) встречались в 21 месте коралловых рифов в период с 1960-х по 1980-е годы. [60] Эти места варьировались от Красного моря через тропический Индо-Тихоокеанский регион до Французской Полинезии. По крайней мере две подтвержденные повторные вспышки произошли в 10 из этих мест.
Значения плотности морских звезд от 140 до 1000/га считаются в различных отчетах популяциями вспышки, тогда как плотность морских звезд менее 100/га считается низкой; [34] однако при плотности ниже 100/га питание A. planci может превышать рост кораллов с чистой потерей кораллов.
Результаты исследований многих мест рифов по всему ареалу морских звезд показали, что большая численность видов Acanthaster . можно классифицировать как:
- Первичные вспышки, при которых резкий рост численности населения как минимум на две величины не может быть объяснен наличием предыдущей вспышки.
- Вторичные вспышки вполне вероятно могут быть связаны с предыдущими вспышками за счет воспроизводства предыдущей когорты морских звезд. Они могут выглядеть как новобранцы, чтобы оттеснить существующую популяцию от вспышки.
- Хронические ситуации, когда постоянная популяция от умеренной до высокой плотности существует на рифе, где кораллов мало, из-за постоянного питания морскими звездами. [61]
Большой Барьерный риф (ББР) — самая выдающаяся система коралловых рифов в мире из-за его большой длины, количества отдельных рифов и видового разнообразия. Когда впервые были замечены высокие плотности акантастера , вызывавшие высокую смертность кораллов, в 1960–65 годах у острова Грин , недалеко от Кэрнса, это вызвало серьезную тревогу. Впоследствии популяции с высокой плотностью были обнаружены на ряде рифов к югу от острова Грин, в центральном регионе ББР. [62] [63] [64] Некоторые популярные издания предположили, что всему рифу грозит гибель. [65] [66] и они повлияли и отразили некоторую общественную тревогу по поводу состояния и будущего ГББР. [ нужна ссылка ]
В ряде исследований демографические вспышки были смоделированы на территории GBR как средство понимания этого явления. [67] [68]
Правительства Австралии и Квинсленда финансировали исследования и создали консультативные комитеты в период большой обеспокоенности по поводу природы вспышек морских звезд в ГББР. Считалось, что они не смирились с беспрецедентным характером и масштабами этой проблемы. [69] Многих учёных критиковали за то, что они не смогли дать однозначных, но необоснованных ответов. Другие были более категоричны в своих ответах. [70] Ученых критиковали за сдержанность и разногласия относительно природы и причин вспышек в ГББР, которые иногда называют «войнами морских звезд». [71] [70]
Причины популяционных вспышек
[ редактировать ]Серьезные дискуссии и некоторые твердые взгляды указывают на причины этого явления. Некоторые гипотезы были сосредоточены на изменениях в выживаемости молоди и взрослых морских звезд - «гипотеза удаления хищников»:
- Чрезмерный вылов тритонов , хищников морских звезд. [72]
- Перелов хищников морских звезд [73]
- Сокращение популяций хищников из-за разрушения среды обитания [30]
- Более теплая температура моря способствует развитию личинок [74]
- Антропогенное воздействие, такое как поступление аллохтонных питательных веществ [75]
Многие сообщения о охоте рыбы на Акантастера представляют собой единичные наблюдения или предполагают хищничество, исходя из характера рыбы. Например, горбатый губан может охотиться на морскую звезду среди своего обычного рациона. [76] Было замечено, что отдельные рыбы-фугу и триггеры кормят морских звезд с терновым венцом в Красном море, но нет никаких доказательств того, что они являются существенным фактором контроля над популяцией. [77] Однако исследование, основанное на содержимом желудков крупных хищных рыб, которые являются потенциальными хищниками морских звезд, не обнаружило никаких признаков присутствия морских звезд в кишках рыбы. Эти хищные рыбы были выловлены в коммерческих целях на коралловых рифах Оманского залива и исследованы на местных рыбных рынках. [78]
Одна из проблем с концепцией хищников крупных молодых и взрослых морских звезд, вызывающих полную смертность, заключается в том, что морские звезды обладают хорошими регенеративными способностями и не могут оставаться на месте, пока их едят. Кроме того, чтобы умереть, их нужно съесть полностью или почти полностью; У 17–60% морских звезд в различных популяциях отсутствовали или регенерировали руки. [34] Очевидно, что морские звезды подвергаются различным уровням сублетального нападения хищников. Когда повреждение включает в себя большую часть диска вместе с рукавами, количество регенерирующих рукавов на диске может быть меньше количества потерянных. [61]
Другая гипотеза - это «гипотеза агрегации», согласно которой большие скопления A. planci выглядят как очевидные вспышки, поскольку они поглотили все прилегающие кораллы. По-видимому, это означает, что, очевидно, существует плотная популяционная вспышка, когда произошла более диффузная популяционная вспышка, которая была достаточно плотной, чтобы массово охотиться на больших площадях твердых кораллов.
Самки морской звезды тернового венца очень плодовиты. По яйцеклеткам в яичниках 200-, 300-. и самки диаметром 400 мм потенциально откладывают около 4, 30 и 50 миллионов яиц соответственно. [79] (см. также Гаметы и эмбрионы). Лукас применил другой подход, сосредоточив внимание на выживании личинок, возникающих из яиц. [80] Обоснованием такого подхода было то, что небольшие изменения в выживаемости личинок и стадиях развития приведут к очень большим изменениям во взрослой популяции, если принять во внимание две гипотетические ситуации.
Около 20 миллионов икринок от нерестящихся самок, имеющих выживаемость около 0,00001% на протяжении всего развития, заменят двух взрослых морских звезд в популяции с низкой плотностью, где пополняются личинки. Если, однако, выживаемость увеличится до 0,1% (одна из тысячи) на протяжении всего развития после одного нереста из 20 миллионов икринок, это приведет к появлению 20 000 взрослых морских звезд, в которых пополнятся личинки. Поскольку личинки представляют собой наиболее распространенную стадию развития, изменения в выживаемости, вероятно, имеют наибольшее значение на этой стадии развития.
Температура и соленость мало влияют на выживаемость личинок тернового венца. [49] Однако численность и виды конкретного компонента фитопланктона ( одноклеточные жгутиконосцы ), которым питаются личинки, оказывают глубокое влияние на выживаемость и скорость роста. Особенно важно обилие клеток фитопланктона. [81] Численность фитопланктона как автотрофов сильно зависит от концентрации неорганических питательных веществ, таких как азотистые соединения.
Биркеланд наблюдал корреляцию между обилием тернового венца на рифах, прилегающих к суше. Они произошли на материковых островах, в отличие от коралловых атоллов, примерно через три года после сильных дождей, последовавших за периодом засухи. [82] Он предположил, что стоки таких сильных дождей могут стимулировать цветение фитопланктона достаточного размера, чтобы производить достаточное количество пищи для личинок A. planci за счет поступления питательных веществ.
Сочетание наблюдений Биркеланда с влиянием неорганических питательных веществ на выживаемость личинок морских звезд в экспериментальных исследованиях подтвердило механизм вспышек морских звезд:
увеличение поверхностного стока → увеличение питательных веществ более плотный фитопланктон↑ → лучшая выживаемость личинок → увеличение популяций морских звезд
В дальнейшем эти связи были подтверждены, но исследования Олсона (1987), Кауфмана (2002) и Бирна (2016) показывают, что земной сток практически не влияет на выживаемость личинок. [83] [84] [85] [86] [87] Противоречивые данные, описывающие незначительную роль наземных сельскохозяйственных стоков, были названы «неудобным исследованием». [83]
Кроме того, эффект притока проявляется в том, что там, где большие популяции морских звезд производят большое количество личинок, вероятно интенсивное пополнение на рифах ниже по течению, куда личинки переносятся и затем оседают. [ нужна ссылка ]
Контроль численности населения
[ редактировать ]
Численность населения тернового венца растет с 1970-х годов. [88] Однако исторические данные о моделях и численности населения найти трудно, поскольку технология подводного плавания , необходимая для проведения переписи населения, была разработана только в предыдущие несколько десятилетий.
Чтобы предотвратить перенаселение тернового венца, вызывающее широкомасштабное разрушение среды обитания коралловых рифов, люди приняли различные меры контроля. Удаление вручную прошло успешно, [44] но относительно трудоемки. Введение бисульфата натрия морской звезде является наиболее эффективной мерой на практике. Бисульфат натрия смертелен для тернового венца, но не вредит окружающим рифам и океаническим экосистемам. [89] Чтобы контролировать районы с высоким уровнем заражения, команды дайверов добились уровня смертности до 120 в час на одного дайвера. [89] Было показано, что практика их расчленения приводит к 12 убийствам в час на одного дайвера, и дайверу, выполнявшему этот тест, вводили шипы трижды. В результате расчленения не рекомендуются именно по этой причине, а не из-за слухов о том, что они могут регенерировать.
Еще более трудоемкий, но менее рискованный для дайвера путь — закопать их под камнями или обломками. Этот маршрут подходит только для районов с низкой зараженностью и при наличии материалов для выполнения процедуры без повреждения кораллов.
Исследование, проведенное в 2015 году Университетом Джеймса Кука, показало, что обычный бытовой уксус также эффективен, поскольку кислотность приводит к разложению морских звезд в течение нескольких дней. Уксус также безвреден для окружающей среды и не ограничен правилами, касающимися продуктов животного происхождения, таких как желчь. [90] В 2019 году дайверы использовали 10%-ный раствор уксуса, чтобы сократить популяцию морских звезд на островах Раджа-Ампат . [91]
Новым успешным методом контроля популяции является введение тиосульфатно-цитратно-желчных солей-сахарозного агара (TCBS). Требуется всего одна инъекция, приводящая к смерти морской звезды в течение 24 часов от заразной болезни, характеризующейся «обесцвечиванием и некрозом кожи, изъязвлениями, потерей тургора тела, скоплением бесцветной слизи на многих шипах, особенно на их кончиках, и потерей шипов. Волдыри. на спинном покрове прорвало поверхность кожи и образовало большие открытые язвы, обнажившие внутренние органы». [92]
Был разработан автономный робот для уничтожения морских звезд под названием COTSBot , и по состоянию на сентябрь 2015 года он был близок к испытаниям на GBR. [93] COTSbot, оснащенный системой машинного зрения на основе нейронной сети , предназначен для поиска морских звезд с терновым венцом и введения им смертельных инъекций желчных солей . После того, как большая часть морских звезд уничтожена в определенном районе, водолазы-люди могут проникнуть и удалить выживших. полевые испытания робота начались в Мортон-Бей в Брисбене По словам исследователя Квинслендского технологического университета Мэтью Данбабина, с целью усовершенствования его навигационной системы. В заливе Мортон нет морских звезд с терновым венцом, но когда навигация будет усовершенствована, на рифе будет использоваться робот. [94] [95] [96]
В Индонезии проводятся исследования по использованию измельченных остатков COTS в качестве обогащающей добавки к корму для белокногих креветок . [97]
Ссылки
[ редактировать ]- ^ «Морская звезда Терновый венец» . Фонд Большого Барьерного рифа . Архивировано из оригинала 7 июля 2023 г. Проверено 06 сентября 2022 г.
- ^ Jump up to: а б Биркеланд и Лукас (1990) , стр. 97–98.
- ^ Jump up to: а б Пити, Р.; Гарм А. и Холл М.Р. (2016). «Терновый венец обладает истинным видением, формирующим образ» . Границы в зоологии . 13 (1). 41. дои : 10.1186/s12983-016-0174-9 . ПМК 5013567 . ПМИД 27605999 .
- ^ Карпентер, Р.К. (1997) Беспозвоночные хищники и травоядные». Архивировано 1 мая 2012 г. в Wayback Machine. В: К. Биркеланд, Жизнь и смерть коралловых рифов , Спрингер. ISBN 978-0-412-03541-8 .
- ^ Касо, MJ (1974). «Внешняя морфология Acanthaster planci (Linnaeus)». Журнал Морской биологической ассоциации Индии . 16 (1): 83–93.
- ^ "Морская звезда Тернового венца (Acanthaster Planci)". Морская звезда «Терновый венец»: видео, фотографии и факты. Веб. 5 ноября 2014 г. < http://www.arkive.org/crown-of-thorns-starfish/acanthaster-planci/. Архивировано 6 ноября 2014 г. в Wayback Machine >.
- ^ Лоуренс, Джон М. (2013). Морские звезды: биология и экология астероидов . Джу Пресс. ISBN 978-1-4214-0787-6 . [ нужна страница ]
- ^ Лоуренс, Джон М. (2013). Морские звезды: биология и экология астероидов . Джу Пресс. ISBN 978-1-4214-0787-6 . [ нужна страница ]
- ^ Jump up to: а б Лукас, Дж. С.; Джонс, ММ (1976). «Гибридная морская звезда с терновым венцом ( Acanthaster planci x A. Brevispinus ), выращенная до зрелости в лаборатории». Природа . 263 (5576): 409–12. Бибкод : 1976Natur.263..409L . дои : 10.1038/263409a0 . ПМИД 972678 . S2CID 4218030 .
- ^ Блейк, диджей (1979). «Родство и происхождение морской звезды с терновым венцом Акантастера Жерве ». Дж. Нэт. Хист . 13 (3): 303–314. Бибкод : 1979JNatH..13..303B . дои : 10.1080/00222937900770241 .
- ^ Биркеланд и Лукас (1990) , стр. 13–19.
- ^ Лукас, Дж. С.; Нэш, У.Дж.; Нисида, М (1985). «Аспекты эволюции Acanthaster planci (L) (Echinodermata, Asteroidea)» Материалы Пятого Международного конгресса коралловых рифов, Таити, 27 мая - 1 июня 1985 г., Vol. 5 .
- ^ Бензи, JAH (1999). «Основные генетические различия между популяциями морских звезд с терновым венцом (Acanthaster planci) в Индийском и Тихом океанах». Эволюция . 53 (6): 1782–1795. дои : 10.2307/2640440 . JSTOR 2640440 . ПМИД 28565442 .
- ^ Фоглер, К.; Бензи, Дж.; Лессиос, Х.; Барбер, П.; Ворхайде, Г. (2008). «Угроза коралловым рифам увеличивается? Четыре вида морских звезд с терновым венцом» . Письма по биологии . 4 (6): 696–9. дои : 10.1098/rsbl.2008.0454 . ПМК 2614177 . ПМИД 18832058 .
- ^ «Оказывается, морских звезд, убивающих кораллы, существует четыре вида, а не один» . Yahoo! Новости . АФП. 30 сентября 2008 г. Архивировано из оригинала 4 октября 2008 г. Проверено 14 октября 2008 г.
- ^ Хадсон Р.Р.; Койн Дж. (2002). «Математические следствия концепции генеалогического вида» . Эволюция . 56 (8): 1557–1565. дои : 10.1111/j.0014-3820.2002.tb01467.x . ПМИД 12353748 . S2CID 8012779 .
- ^ Рубинов, Д. (2006). «Полезность штрих-кодов митохондриальной ДНК в сохранении видов». Консервировать. Биол . 20 (4): 1026–1033. Бибкод : 2006ConBi..20.1026R . дои : 10.1111/j.1523-1739.2006.00372.x . ПМИД 16922219 . S2CID 25740078 .
- ^ Бэбкок Р., Манди С., Кизинг Дж., Оливер Дж. (1992). «Предсказуемые и непредсказуемые события нереста: данные о поведении in situ беспозвоночных коралловых рифов, нерестящихся на свободном месте». Размножение и развитие беспозвоночных . 22 (1–3): 213–228. Бибкод : 1992InvRD..22..213B . дои : 10.1080/07924259.1992.9672274 .
- ^ Биркеланд и Лукас (1990) , стр. 131–132.
- ^ Барнетт, Д; и др. (1968). «Определение содержания стероидных сапонинов в тканях морских звезд и изучение их биосинтеза». Сравнительная биохимия и физиология Б . 90 (1): 141–145. дои : 10.1016/0305-0491(88)90050-8 .
- ^ Коул, Эндрю; Пратчетт, Морган; Джонс, Джеффри (2008). «Разнообразие и функциональное значение коралловых рыб тропических коралловых рифов». Рыба и рыболовство . 9 (3): 286–307. дои : 10.1111/j.1467-2979.2008.00290.x .
- ^ Печеник, Ю. (2015). Биология беспозвоночных (7-е изд.). Нью-Йорк: Образование Макгроу-Хилл. п. 505.
- ^ Белк, Д. (1975). «Наблюдение за колонизацией водорослей на Acropora aspera , уничтоженном Acanthaster planci ». Гидробиология . 46 (1): 29–32. дои : 10.1007/bf00038724 . S2CID 25260922 .
- ^ Пьер Мадл (1998). «Acanthaster planci (Обзор морской звезды тернового венца)» . Проверено 28 августа 2006 г.
- ^ Кизинг Дж.К., Лукас Дж.С. (1992). «Полевые измерения скорости питания и передвижения морской звезды с терновым венцом Acanthaster planci (L.)». Журнал экспериментальной морской биологии и экологии . 156 : 89–104. дои : 10.1016/0022-0981(92)90018-6 .
- ^ Мюллер Б; АР Бос; Г Граф; Г. С. Гуманао (2011). «Скорость передвижения и характер движения четырех распространенных морских звезд Индо-Тихоокеанского региона (Echinodermata; Asteroidea) в зависимости от размера» . Водная биология . 12 (2): 157–164. дои : 10.3354/ab00326 .
- ^ Биркеланд и Лукас (1990) , стр. 149.
- ^ Де Вантье; Райхельт и Брэдбери (1986). «Защищает ли Spirobranchus giganteus хозяина Porites от хищников Acanthaster planci : давление хищников как механизм коэволюции?» . Серия «Прогресс в области морской экологии» . 32 : 307–310. Бибкод : 1986MEPS...32..307D . дои : 10.3354/meps032307 .
- ^ Пратчетт (2001). «Влияние коралловых симбионтов на пищевые предпочтения морской звезды с терновым венцом Acanthaster planci в западной части Тихого океана» . Серия «Прогресс в области морской экологии» . 214 : 111–119. Бибкод : 2001MEPS..214..111P . дои : 10.3354/meps214111 .
- ^ Jump up to: а б Чансанг, Х (1987). «Заражение Acanthaster planci в Андаманском море». Второй международный симпозиум по морской биологии Индо-Тихоокеанского региона Западное общество естествоиспытателей, Университет Гуама, 23-28 июня 1986 года . OCLC 17256640 .
- ^ Бэбкок; Манди и Уайтхед (1994). «Модели диффузии спермы и подтверждение in situ оплодотворения на больших расстояниях на свободно нерестящемся астероиде Acanthaster planci » . Биологический бюллетень . 186 (1): 17–28. дои : 10.2307/1542033 . JSTOR 1542033 . ПМИД 29283304 .
- ^ Пляж, DH; Хэнскомб, Нью-Джерси; Ормонд, РФГ (1975). «Нерест феромона морской звезды с терновым венцом». Природа . 254 (5496): 135–136. Бибкод : 1975Natur.254..135B . дои : 10.1038/254135a0 . ПМИД 1117997 . S2CID 4278163 .
- ^ Свитман, HPA (1995). «Полевое исследование хищничества рыб на молодых морских звездах с терновым венцом». Коралловые рифы . 14 (1): 47–53. Бибкод : 1995КорРе..14...47С . дои : 10.1007/bf00304071 . S2CID 6434509 .
- ^ Jump up to: а б с Моран, П.Дж. (1986). « Феномен Акантастера ». Океанография и морская биология . 24 : 379–480. ИНИСТ 8358915 НАИД 10027122155 . Переиздано как: Моран, Питер Джон (1988). Феномен Акантастера . Австралийский институт морских наук. дои : 10.5962/bhl.title.60630 . ISBN 978-0-642-13250-5 .
- ^ Ормонд, Руперт ФГ; Кэмпбелл, Эндрю К. (1974). «Формирование и распад скоплений Acanthaster planci в Красном море». Материалы Второго международного симпозиума по коралловым рифам . Том. 1. Комитет Большого Барьерного рифа. стр. 595–620. ISBN 978-0-909377-00-7 . ПроКвест 17931170 .
- ^ Хищники COTS
- ^ Пауэлл, Дж. (1979). «Звезды для королей». Морские рубежи . 25 (5): 282–285.
- ^ Jump up to: а б Глинн П.В. (1982). «Регулирование численности акантастера креветкой и червем» . В Гомес ЭД, Биркеланд К.Э., Бадемайер Р.В., Йоханнес Р.Э., Марш-младший Дж.А., Цуда Р.Т. (ред.). Материалы Четвертого Международного симпозиума по коралловым рифам, 1981. Том. 2 . Международный симпозиум по коралловым рифам, Манила (Филиппины), 18–22 мая 1981 г., стр. 607–612. ПроКвест 13786745 .
- ^ Jump up to: а б Глинн, Питер В. (1 июля 1984 г.). «Амфиномидный червь-хищник морской звезды Терновый венец и общее хищничество на астероидах в коралловых рифах восточной и западной части Тихого океана» . Бюллетень морской науки . 35 (1): 54–71.
- ^ Jump up to: а б Birkeland & Lucas (1990) , необходимы стр.
- ^ Бос, Арканзас; Гуманао, Г.С.; Салац, ФН (2008). «Недавно обнаруженный хищник морской звезды с терновым венцом». Коралловые рифы . 27 (3): 581. Бибкод : 2008CorRe..27..581B . дои : 10.1007/s00338-008-0364-9 . S2CID 34920961 .
- ^ Бос АР; Б. Мюллер; Г. С. Гуманао (2011). «Биология питания и симбиотические отношения кораллиморфных гоплитов Paracorynactis (Anthozoa: Hexacorallia)» . Зоологический бюллетень Raffles . 59 (2): 245–250.
- ^ Биркеланд и Лукас (1990) , стр. 63.
- ^ Jump up to: а б с д Бос, Артур Р.; Гуманао, Гирли С.; Мюллер, Бенджамин; Саседа-Кардоса, Марджо М.Э. (2013). «Борьба со вспышками морской звезды тернового венца (Acanthaster planci L.): успех удаления зависит от топографии рифа и сроков цикла воспроизводства». Управление океаном и прибрежной зоной . 71 : 116–22. Бибкод : 2013OCM....71..116B . дои : 10.1016/j.ocecoaman.2012.09.011 .
- ^ Бэбкок, Колорадо; Манди, Китай; Энгельгардт, У; Лассиг, Б. (1993). «Сезонные изменения фертильности и плодовитости Acanthaster planci». Управление морского парка Большого Барьерного рифа, Таунсвилл, Квинсленд (Австралия) .
- ^ Скотт, CM; Мехротра, Р.; Уржелл, П. (2014). «Наблюдение за нерестом Acanthaster planci в Сиамском заливе». Морское биоразнообразие . 45 (4): 621–2. дои : 10.1007/s12526-014-0300-x . S2CID 33626561 .
- ^ Герра, Ванесса; Хейнс, Гвилим; Бирн, Мария; Ясуда, Нина; Адачи, Сота; Накамура, Масако; Накачи, Шу; Харт, Майкл В. (14 декабря 2019 г.). «Неспецифическая экспрессия генов оплодотворения у тернового венца Acanthaster cf. Solaris: неожиданное свидетельство гермафродитизма у хищника коралловых рифов» . Молекулярная экология . 29 (2): 363–379. дои : 10.1111/mec.15332 . ISSN 0962-1083 . ПМИД 31837059 .
- ^ Хендерсон Дж. А., Лукас Дж. С. (1971). «Развитие личинок и метаморфоз Acanthaster planci (Asteroidea)». Природа . 232 (5313): 655–657. Бибкод : 1971Natur.232..655H . дои : 10.1038/232655a0 . ПМИД 16063148 . S2CID 4156996 .
- ^ Jump up to: а б Лукас, Джон (1973). «Репродуктивная и личиночная биология и Acanthaster planci (L.) в водах Большого Барьерного рифа» (PDF) . Микронесика . 9 (2): 197–203.
- ^ Jump up to: а б Ямагучи, М. (1973). «Ранние истории жизни астероидов коралловых рифов, с особым упором на Acanthaster planci (L.)». В Джонс, А.О. и Эндин, Р. (ред.). Биология и геология коралловых рифов Том. 2 . Нью-Йорк: Академическая пресса. п. 369.
- ^ Лукас Дж.С. (1984). «Рост, созревание и влияние диеты у Acanthaster planc i (L.) (Asteroidea) и гибридов, выращенных в лаборатории». Журнал экспериментальной морской биологии и экологии . 79 (2): 129–147. дои : 10.1016/0022-0981(84)90214-4 .
- ^ Jump up to: а б Занн, Леон; Броди, Джон; Берриман, Кристина; Написано Миликой (сентябрь 1987 г.). «Набор, экология, рост и поведение молодых особей Acanthaster Planci (L.) (Echinodermata: Asteroidea)» . Бюллетень морской науки . 41 (2): 561–575.
- ^ Стамп, Р. (1993). Характеристики истории жизни популяций Acanthaster planci (L.), потенциальные ключи к разгадке причин вспышек . Управление морского парка Большого Барьерного рифа, Таунсвилл, Квинсленд, Австралия. [ нужна страница ]
- ^ Кизинг Дж. К., Хэлфорд А. Р. (1992). «Полевые измерения выживаемости молоди Acanthaster planci: методы и предварительные результаты» . Серия «Прогресс в области морской экологии» . 85 (1–2): 107–114. Бибкод : 1992MEPS...85..107K . дои : 10.3354/meps085107 .
- ^ Пратчетт, Морган; Утик, Свен (2017). Биология, экология и управление морскими звездами с терновым венцом . Базель, Швейцария: Книги MDPI. дои : 10.3390/books978-3-03842-603-5 . ISBN 978-3-03842-603-5 .
- ^ Джеймс, Питер (1976). Реквием по рифу: история официального искажения информации о морской звезде с терновым венцом . Фонд Пресс. ISBN 978-0-9597383-0-8 . [ нужна страница ]
- ^ Ларри Зетлин (2004). Хищники Большого Барьерного рифа (ТВ-специальный выпуск). Квинсленд, Австралия: Gulliver Media Australia.
- ^ Уилсон, Шон К; Грэм, Николас Эй Джей; Пратчетт, Морган С; Джонс, Джеффри П.; Полунин, Николай В.К. (2006). «Множественные нарушения и глобальная деградация коралловых рифов: находятся ли рифовые рыбы в опасности или устойчивы?». Биология глобальных изменений . 12 (11): 2220–2234. Бибкод : 2006GCBio..12.2220W . дои : 10.1111/j.1365-2486.2006.01252.x . S2CID 59933337 .
- ^ Уилсон, СК; Долман, AM; Чил, Эй Джей; Эмсли, М. Дж. Пратчетт; и др. (2009). «Поддержание разнообразия рыб на нарушенных коралловых рифах» . Коралловые рифы . 28 (1): 3–14. Бибкод : 2009CorRe..28....3W . дои : 10.1007/s00338-008-0431-2 .
- ^ Биркеланд и Лукас (1990) , стр. 36–40.
- ^ Jump up to: а б Биркеланд и Лукас (1990) , стр. 38.
- ^ Эндин, Р. (1974). Материалы Второго международного симпозиума по коралловым рифам. Том. 1 . Комитет Большого Барьерного рифа, Брисбен (Австралия). ISBN 978-0-909377-00-7 . [ нужна страница ]
- ^ Эндин, Р; Стаблум, В. (1973). «Исследование некоторых аспектов заражения морской звездой тернового венца (Acanthaster planci) рифов Австралии и Большого Барьерного рифа». Бюллетень исследований атолла . 167 : 1–62. дои : 10.5479/si.00775630.167.1 .
- ^ Кенчингтон, Р. (1978). «Кризис тернового венца в Австралии: ретроспективный анализ». Охрана окружающей среды . 5 (1): 11–20. Бибкод : 1978EnvCo...5...11K . дои : 10.1017/s0376892900005191 . S2CID 86566114 .
- ^ Джеймс, Питер (1976). Реквием по рифу: история официального искажения информации о морской звезде с терновым венцом . Фонд Пресс. ISBN 978-0-9597383-0-8 . [ нужна страница ]
- ^ Браун, Тео; Уилли, Кейт (1972). Терновый венец: гибель Большого Барьерного рифа? . Сидней: Ангус и Робертсон. ISBN 978-0-207-12390-0 . [ нужна страница ]
- ^ Райхельт, Р.Э.; Брэдбери, Р.Х.; Моран, П. Дж. (1990). «Морская звезда с терновым венцом, Acanthaster planci, на Большом Барьерном рифе» . Математическое и компьютерное моделирование . 13 (3): 45–60. дои : 10.1016/0895-7177(90)90008-б .
- ^ Скандол, Дж. П. (декабрь 1999 г.). «CotSim - интерактивная модель метапопуляции Acanthaster planci для центральной части Большого Барьерного рифа». Морские модели онлайн . 1–4 (1–4): 39–81. дои : 10.1016/S0079-6611(99)00003-8 .
- ^ Раймонд, Роберт (1986). Войны морских звезд: Коралловая смерть и терновый венец . Мельбурн: Макмиллан. ISBN 978-0-333-43015-6 .
- ^ Jump up to: а б Эндин, Р; Кэмерон, AM (1985). «Экокатастрофа на Большом Барьерном рифе». Материалы пятого международного конгресса по коралловым рифам, Таити, 27 мая - 1 июня 1985 г., Vol. 5: Разные документы (А) . Музей Антенны-EPHE, Муреа (Французская Полинезия). стр. 309–314.
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Starfish Wars (название - пьеса по «Звездным войнам »), 1986, Мельбурн, Роберт Рэймонд.
- ^ Висенте, Нардо (декабрь 1999 г.). «Природные сокровища под тяжелым давлением» . Океанорама (на французском языке). № 30. с. 7–12.
- ^ Брэдбери, Р.Х. (1991). «ПОНЯТИЕ АКАНТАСТРА ». Ценозы . 6 (3): 121–126. JSTOR 43461274 .
- ^ Утик, С.; Логан, М.; Лидди, М.; Фрэнсис, Д.; Харди, Н.; Ламаре, М. (2015). «Изменение климата как неожиданный сопутствующий фактор, способствующий вспышкам вспышек морских звезд, поедающих кораллы (Acanthaster planci)» . Научные отчеты . 5 : 8402. Бибкод : 2015NatSR...5E8402U . дои : 10.1038/srep08402 . ПМЦ 4325318 . ПМИД 25672480 .
- ^ Броди, Дж.; Фабрициус, К.; Де'Ат, Г.; Окадзи, К. (2005). «Является ли увеличение поступления питательных веществ причиной увеличения количества вспышек морских звезд с терновым венцом? Оценка доказательств». Бюллетень о загрязнении морской среды . 51 (1–4): 266–78. Бибкод : 2005MarPB..51..266B . дои : 10.1016/j.marpolbul.2004.10.035 . ПМИД 15757727 .
- ^ Рэндалл, Джон Э.; Руководитель Стивен М.; Сандерс, Адриан П.Л. (1978). «Пищевые привычки гигантского горбатого губана Cheilinus undulatus (Labridae)». Экологическая биология рыб . 3 (2): 235–8. Бибкод : 1978EnvBF...3..235R . дои : 10.1007/BF00691948 . S2CID 10744732 .
- ^ Ормонд, РФГ; Кэмпбелл, AC (1974). «Формирование и распад скоплений Acanthaster planci в Красном море». Материалы Второго международного симпозиума по коралловым рифам. Том. 1 .
- ^ Мендонса, Ванда Мариям; Аль-Джабри3, Мусаллам Мубарак; Аль Аджми, Ибрагим; Аль Мухаррами, Мохамед; Ареими, Мохамед Аль; Аль Агбари, Хусейн Али (2010). «Постоянные и расширяющиеся вспышки популяций кораллоядных морских звезд Acanthaster planci в северо-западной части Индийского океана: действительно ли они являются следствием неустойчивого удаления хищников-морских звезд в результате чрезмерного вылова рыбы на коралловых рифах или ответом на изменение окружающей среды?» . Зоологические исследования . 49 (1): 108–123.
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Чайник, Британская Колумбия; Лукас, Дж. С. (1 сентября 1987 г.). «Биометрические взаимосвязи между показателями органов, плодовитостью, потреблением кислорода и размером тела у Acanthaster Planci (L.) (Echinodermata; Asteroidea)» . Бюллетень морской науки . 41 (2): 541–551.
- ^ Хендерсон, Дж.А.; Лукас (1971). «Развитие личинок и метаморфоз Acanthaster planci (Asteroidea)». Природа . 232 (5313): 655–657. Бибкод : 1971Natur.232..655H . дои : 10.1038/232655a0 . ПМИД 16063148 . S2CID 4156996 .
- ^ Лукас, Дж (1982). «Количественные исследования питания и питания во время развития личинок астероида кораллового рифа 'Acanthaster planci' (L.)». Журнал экспериментальной морской биологии и экологии . 65 (2): 173–194. дои : 10.1016/0022-0981(82)90043-0 .
- ^ Биркеланд, К. (1982). «Земной сток как причина вспышек Acanthaster planci (Echinodermata: Asteroidea)». Морская биология . 69 (2): 175–185. Бибкод : 1982MarBi..69..175B . дои : 10.1007/bf00396897 . S2CID 85130293 .
- ^ Jump up to: а б Вулф, Кеннеди; Граба-Ландри, Алексия; Дворжанин, Саймон А.; Бирн, Мария (2017). «Суперзвезды: оценка порогов питательных веществ для повышения успеха личинок Acanthaster planci , обзор доказательств». Бюллетень о загрязнении морской среды . 116 (1–2): 307–314. Бибкод : 2017MarPB.116..307W . дои : 10.1016/j.marpolbul.2016.12.079 . ПМИД 28094041 .
- ^ Броди, Джон; Фабрициус, Катарина; Де'Ат, Гленн; Окадзи, Кен (2005). «Является ли увеличение поступления питательных веществ причиной увеличения количества вспышек морских звезд с терновым венцом? Оценка доказательств». Бюллетень о загрязнении морской среды . 51 (1–4): 266–78. Бибкод : 2005MarPB..51..266B . дои : 10.1016/j.marpolbul.2004.10.035 . ПМИД 15757727 .
- ^ Фабрициус, К.Э.; Окадзи, К.; Де'Ат, Г. (2010). «Три линии доказательств, связывающих вспышки морской звезды с терновым венцом Acanthaster planci с выделением личинок ограничения в пище». Коралловые рифы . 29 (3): 593–605. Бибкод : 2010CorRe..29..593F . дои : 10.1007/s00338-010-0628-z . S2CID 1469117 .
- ^ Кауфман, МГ; Гудфренд, В; Колер-Гэрриган, А; Уокер, Эд; Клюг, MJ (2002). «Влияние растворимых питательных веществ на микробные сообщества и продукцию комаров в местах обитания Ochlerotatus triseriatus» . Водная микробная экология . 29 : 73–88. дои : 10.3354/ame029073 .
- ^ Олсон, Ричард Рэндольф (1987). «Культивирование in situ как проверка гипотезы голодания личинок морской звезды Acanthaster planci » . Лимнология и океанография . 32 (4): 895–904. Бибкод : 1987LimOc..32..895O . дои : 10.4319/lo.1987.32.4.0895 .
- ↑ Учёные: Чтобы спасти Большой Барьерный риф, убивайте морские звезды . Новости CNN . 2 октября 2012 г.
- ^ Jump up to: а б «Управление морской звездой тернового венца» . Управление морского парка Большого Барьерного рифа. Апрель 1995 года. Архивировано из оригинала 12 октября 2008 года . Проверено 15 декабря 2008 г.
- ^ Колдервуд, Кэтлин; Роу, Изобель (23 сентября 2015 г.). «Бытовой уксус способствует борьбе с угрозой появления морских звезд в виде тернового венца на Большом Барьерном рифе» . Новости АВС . Проверено 25 марта 2016 г.
- ^ Келли, Джон (23 июня 2019 г.). «Аллангисты из округа Колумбия выкапывают свои старые трубки и ласты, чтобы сразиться с пожирателем кораллов» . Вашингтон Пост . Проверено 20 декабря 2023 г.
- ^ Ривера-Посада, JA; Пратчетт, М.; Кано-Гомес, А.; Аранго-Гомез, доктор юридических наук; Оуэнс, Л. (6 декабря 2011 г.). «Инъекция Acanthaster planci с тиосульфатно-цитратно-желчно-сахарозным агаром (TCBS). I. Индукция заболевания» . Болезни водных организмов . 97 (2): 85–95. дои : 10.3354/dao02401 . ПМИД 22303625 .
- ^ Маклиш, Кэти (31 августа 2015 г.). «Новый робот призван уничтожить морскую звезду с терновым венцом, угрожающую рифам» . Новости АВС . Архивировано из оригинала 8 ноября 2020 года . Проверено 28 сентября 2021 г.
- ^ Эспинер, Том , убивающий морских звезд, близок к испытаниям на Большом Барьерном рифе BBC News . 4 сентября 2015 г.
- ^ Григгс, Робот Мэри Бет, предназначенный для уничтожения морских звезд, будет выпущен на австралийские рифы Научно-популярная статья . . 4 сентября 2015 г.
- ^ «Водный робот ищет и уничтожает морские звезды, уничтожающие рифы» . 07.09.2015 . Проверено 25 марта 2016 г.
- ^ Мухаммад Сафир; Новалина Сердиати; Касим Мансюр; Фадлы Ю. Танту; Билли Сабилла (2022). «Оценка муки морской звезды с шипами короны (Acanthaster planci) как кандидата в качестве кормового ингредиента для креветок ваннамеи (Litopeneus vannamei)» . Медиа Акуатика . 7 (4). Университет Халу Олео: 170–8. ISSN 2503-4324 .
Дальнейшее чтение
[ редактировать ]- Биркеланд, К.; Лукас, Дж. С. (1990). Acanthaster planci: Основная проблема управления коралловыми рифами . Бока-Ратон, Флорида: CRC Press. ISBN 978-0-8493-6599-7 .
Внешние ссылки
[ редактировать ] в этой статье Использование внешних ссылок может не соответствовать политике и рекомендациям Википедии . ( Март 2016 г. ) |
- Обзор морской звезды с терновым венцом, наблюдаемой на Большом Барьерном рифе.
- Текущее состояние морской звезды с терновым венцом на Большом Барьерном рифе Австралии. Архивировано 20 ноября 2008 г. в Wayback Machine.
- История вспышек морских звезд с терновым венцом на Большом Барьерном рифе Австралии с 1986 года (Анимация)
- Последняя информация о морской звезде с терновым венцом на Большом Барьерном рифе Австралии.
- Управление морской звездой с терновым венцом
- Моделирование вспышки тернового венца
- Микродокументы. Архивировано 27 июля 2011 г. в Wayback Machine : Терновый венец. Архивировано 7 ноября 2012 г. в Wayback Machine.
- Экспедиция Acanthaster planci 1969 года на Восточные Каролинские острова.
- Инфографика о морской звезде с терновым венцом на Большом Барьерном рифе.
- АДАПТАЦИЯ: Коралловые рифы Исследователь климата Вануату Ализе Каррер исследует, как люди справляются с морскими звездами с терновым венцом
- Фотографии морской звезды Тернового венца из коллекции Sealife.
- Геном A. planci на эхинобазе
| Базы данных органов управления : Национальные |
|---|