Разделение биомассы
Распределение биомассы – это процесс, посредством которого растения распределяют свою энергию между листьями , стеблями , корнями и репродуктивными частями . Эти четыре основных компонента растения играют важную морфологическую роль: листья поглощают CO 2 и энергию солнца для создания соединений углерода , стебли растут выше конкурентов, чтобы достичь солнечного света, корни поглощают воду и минеральные питательные вещества из почвы, закрепляя растение, и репродуктивные части способствуют продолжению вида. Растения распределяют биомассу в ответ на ограничение или избыток ресурсов, таких как солнечный свет, углекислый газ, минеральные питательные вещества и вода, а рост регулируется постоянным балансом между распределением биомассы между частями растения. Равновесие между ростом корней и побегов возникает потому, что корням необходимы соединения углерода, образующиеся в результате фотосинтеза в побегах, а побегам необходим азот, поглощаемый корнями из почвы. [1] Распределение биомассы приближается к пределу роста; ограничение под землей будет концентрировать биомассу к корням, а ограничение над землей будет способствовать большему росту побегов. [2]
Растения фотосинтезируют , создавая углеродные соединения для роста и хранения энергии. Сахара, образующиеся в результате фотосинтеза, затем транспортируются по флоэме с помощью системы потока под давлением и используются для роста или сохраняются для дальнейшего использования. Разделение биомассы приводит к разделению этого сахара таким образом, чтобы максимизировать рост, обеспечить максимальную приспособленность и обеспечить успешное воспроизводство. Растительные гормоны играют большую роль в распределении биомассы, поскольку они влияют на дифференцировку и рост клеток и тканей, изменяя экспрессию генов и изменяя морфологию . [3] Реагируя на стимулы окружающей среды и соответствующим образом распределяя биомассу, растения могут лучше усваивать ресурсы из окружающей среды и максимизировать рост.
Абиотические факторы разделения
[ редактировать ]
Для растений важно иметь возможность сбалансировать поглощение и использование доступных ресурсов, и они корректируют свой рост, чтобы получить больше дефицитных, ограничивающих рост ресурсов: солнечного света, углекислого газа, минеральных питательных веществ и воды. [4] Равновесие распределения биомассы можно объяснить Законом минимума Либиха и смоделировать с помощью метафоры бочки Либиха , в которой ограничивающий ресурс роста растений подобен самой короткой планке на бочке, полной воды. [5] Бочка может удерживать воду только до уровня самой короткой планки, и, аналогично, растения могут расти только со скоростью, допускаемой ограниченным ресурсом. Чтобы продолжить рост, биомассу необходимо разделить, чтобы помочь изолировать эти ресурсы. [ нужна ссылка ]
Солнечный свет
[ редактировать ]Основным светочувствительным механизмом растений является фитохромная система с пигментами, расположенными по всему растению и позволяющими улавливать изменения красного и дальнего красного света. [2] Обнаружение качества света с помощью фитохромов помогает вызвать изменения в распределении биомассы. Показано, что растения, выращенные в условиях низкой освещенности, отдают больше биомассы побегам (в основном листьям). Измеряя площадь листьев растений при различных уровнях освещенности или солнечного света, было установлено, что более низкие уровни света приводят к увеличению общей поверхности листьев растений. [6] Если солнечный свет ограничен, растение увеличивает рост побегов и уменьшает энергию, передаваемую корням, поскольку более низкие темпы фотосинтеза уменьшают потребность в воде и минеральных питательных веществах. [2] Поскольку при слабом освещении происходит большее выделение побега, такая же корреляция наблюдается и для растений при ярком освещении; более высокая освещенность приводит к уменьшению общей площади нижней поверхности, поскольку для поглощения солнечного света требуется меньшая площадь поверхности. Когда у растения более чем достаточно фотосинтетической способности, оно вместо этого отдает приоритет росту корней, чтобы поглощать воду и питательные вещества. [ нужна ссылка ]
Углекислый газ
[ редактировать ]В ситуации, когда углерод является ограничивающим ресурсом, увеличение уровня CO 2 увеличивает скорость фотосинтеза. [7] Это также приведет к увеличению поглощения питательных веществ и использования воды, что приведет к большему росту корней. При низких концентрациях CO 2 растения создают более крупные и многочисленные листья, чтобы поглощать больше CO 2 . [7] Влияние концентрации углекислого газа в атмосфере на распределение биомассы важно для понимания воздействия на растения в условиях изменения климата. В растительных клетках соотношение углерода и азота увеличивается, когда концентрация CO 2 выше, что снижает уровень разложения. Уменьшение разложения, вызванное высокими концентрациями углекислого газа, приводит к снижению доступности азота для растений. [8]

Минеральные питательные вещества
[ редактировать ]В целом, доступность питательных веществ оказывает сильное влияние на разделение: растения, растущие в районах с бедными питательными веществами, распределяют большую часть своей биомассы в подземные структуры. [9] Например, доступность азота в почве является решающим фактором распределения биомассы. Например, в системах с низкой продуктивностью (низкий уровень азота в почве), таких как бореальные леса , деревья отдают большую часть своей биомассы корням. Но по мере увеличения продуктивности почвы биомасса в первую очередь распределяется по надземным структурам, таким как листья и стебли. [9] В целом длина корней уменьшается по мере увеличения концентрации питательных веществ. [4] Высокие концентрации минеральных питательных веществ, которые приводят к токсичности, также могут оказывать сильное влияние на рост и разделение. Например, было показано, что в токсичных концентрациях кадмия и свинца Fagus sylvatica развивает более компактную и менее разветвленную корневую систему, при этом образуя мало корневых волосков. [10] Эта измененная структура позволяет уменьшить площадь поверхности корней и уменьшить воздействие токсичных питательных веществ, а также сохранить биомассу для тех частей растения, рост которых был бы более полезным.
Вода
[ редактировать ]Растения, выращенные в засушливых условиях, часто имеют пониженное общее производство биомассы, но они также отдают большую часть своей биомассы корням и развивают более высокое соотношение корней и побегов. [4] Когда растения отдают большую часть своей биомассы корням, они могут улучшить поглощение воды, проникая глубже в грунтовые воды и расширяя корневую массу дальше в стороны. Увеличение количества корневых волосков также способствует увеличению абсорбции. При сильной почвенной засухе биомасса корней и побегов не увеличивается, поскольку устанавливается состояние покоя. [11] Доступность воды также влияет на площадь поверхности листьев, поскольку слишком большая площадь поверхности может привести к чрезмерной транспирации в условиях маловодия.
Биотические факторы разделения
[ редактировать ]Взаимодействия между организмами также могут изменить разделение из-за конкуренции за ресурсы, совместного использования ресурсов или уменьшения биомассы растений.
Соревнование
[ редактировать ]Межвидовая и внутривидовая конкуренция может вызвать уменьшение доступных ресурсов для отдельного растения и изменить способ распределения биомассы. Например, конкуренция между растениями вызывает уменьшение радиального роста ветвей и стебля при одновременном увеличении роста корней и высоты стебля. [12] Это дает обоснование важности фенотипической пластичности для поддержания приспособленности в конкурентной среде; чем лучше растение изменяет свою морфологию, тем более конкурентоспособным оно будет [ нужна ссылка ] .
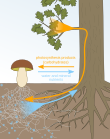
Мутуализм с микоризой
[ редактировать ]Отношения между растениями и микоризными грибами являются примером мутуализма , поскольку растения снабжают грибы углеводами, а микоризные грибы помогают растениям поглощать больше воды и питательных веществ. Поскольку микоризные грибы увеличивают поглощение растениями подземных ресурсов, растения, которые образуют мутуалистические отношения с грибами, стимулируют рост побегов и более высокое соотношение побегов и корней. [13]
Травоядные
[ редактировать ]Травоядность вызывает кратковременное уменьшение массы листьев и/или массы стебля, что изменяет стабильность и пропорции частей растения. Чтобы восстановить баланс между поглощением питательных веществ/воды и скоростью фотосинтеза, растения посылают больше энергии стеблям и листьям. Травоядность, вызывающая высокий уровень дефолиации (более 25% площади листьев), увеличивает рост побега, стремясь достичь того же соотношения побегов и корней, что и до дефолиации. [14] Таким образом, дефолиация также снижает рост корней и поглощение питательных веществ до тех пор, пока соотношение корней и побегов не восстановится до того, как было до дефолиации.
Тип растения
[ редактировать ]Поскольку разные растения имеют различное строение и формы роста, распределение биомассы у них также неодинаково. Например, вечнозеленые деревья имеют более высокую массовую долю листьев (LMF) по сравнению с лиственными деревьями. Кроме того, соотношение стебля и корня у травянистых растений варьируется в большей степени, чем у древесных, поскольку пластичность корней у травянистых растений выше. Кроме того, травянистые однодольные или злаки по сравнению с травянистыми двудольными имеют большее соотношение массы корня к общей массе (RMF). [15] Разницу между травянистыми однодольными и двудольными можно объяснить различиями в эффективности поглощения питательных веществ или необходимостью злаков хранить больше крахмала и питательных веществ для повторного роста после пожара или выпаса скота. На распределение биомассы также может влиять метод фиксации углерода растением: C3 или C4 . Для травянистых однодольных тип фотосинтеза не влияет на процентиль LMF или RMF, но у травянистых двудольных виды C4 имеют более низкие RMF. [15] Жизненный цикл растений также может обуславливать различные стратегии и соотношения листьев, корней и стеблей; того же размера По сравнению с эвдикотелидонными многолетними растениями , однолетние растения тратят больше энергии на рост листьев и стеблей, чем многолетние виды. [15]
Возраст
[ редактировать ]Схема распределения биомассы также может меняться по мере старения растений. В насаждениях Pinus strobus в возрасте 2, 15, 30 и 65 лет отношение корней к побегам составляло 0,32, 0,24, 0,16 и 0,22 соответственно, что свидетельствует об уменьшении массы корней по сравнению с массой побегов в первые несколько десятилетий роста. [16] Возраст также влияет на то, как деревья распределяют биомассу по различным компонентам ствола. Рост ветвей и площади поверхности листьев уменьшается с возрастом по мере увеличения перегородок ствола. [16]
Методы/Измерения
[ редактировать ]Многие исследования по разделению биомассы состоят из манипулятивных экспериментов , которые увеличивают или уменьшают уровни ресурсов, ограничивающих рост, и наблюдают за эффектами. Часто в этих исследованиях используются горшечные растения, выращенные в теплице, для измерения влияния плотности, избытка питательных веществ, слабого освещения и т. д. Другие исследования могут быть сосредоточены на наблюдении и анализе растений, встречающихся в природе, или использовать анализ данных предыдущих измерений. Методы и измерения для определения распределения биомассы могут быть довольно трудными из-за веса и объема более крупных растений. Кроме того, измерение размера и веса корней требует промывания и тщательного удаления почвы для получения точных измерений. [2] Биомасса растения часто измеряется с точки зрения массовой доли листьев (LMF), массовой доли стебля (SMF) и массовой доли корня (RMF), где сухая масса части растения равна общей сухой массе растения. [2] Латеральное, радиальное увеличение и увеличение высоты используются для количественной оценки скорости роста. [ нужна ссылка ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Фаррар, Дж. Ф.; Джонс, Д.Л. (2000). «Контроль поглощения углерода корнями» . Новый фитолог . 147 (1): 43–53. дои : 10.1046/j.1469-8137.2000.00688.x . ISSN 1469-8137 .
- ^ Перейти обратно: а б с д и Пуртер, Хендрик; Никлас, Карл Дж.; Райх, Питер Б.; Олексин, Яцек; Пут, Питер; Моммер, Лисье (2012). «Распределение биомассы по листьям, стеблям и корням: метаанализ межвидовых вариаций и контроль окружающей среды» . Новый фитолог . 193 (1): 30–50. дои : 10.1111/j.1469-8137.2011.03952.x . ISSN 1469-8137 . ПМИД 22085245 .
- ^ Мохаммед, Тунгос (6 августа 2018 г.). «Вещества, способствующие росту растений в растениеводстве: обзор» .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Перейти обратно: а б с Ву, Фучжун; Бао, Вейкай; Ли, Фанглан; Ву, Нин (01 мая 2008 г.). «Влияние стресса засухи и поступления азота на рост, распределение биомассы и эффективность использования воды сеянцев Sophora davidii». Экологическая и экспериментальная ботаника . 63 (1–3): 248–255. дои : 10.1016/j.envexpbot.2007.11.002 .
- ^ Плог, Р.Р. ван дер; Бём, В.; Киркхэм, МБ (1999). «О происхождении теории минерального питания растений и закона минимума» . Журнал Американского общества почвоведения . 63 (5): 1055–1062. Бибкод : 1999SSASJ..63.1055V . дои : 10.2136/sssaj1999.6351055x . ISSN 1435-0661 .
- ^ Ван, Дж. Джефф; Бауэрле, Уильям Л.; Маддер, Брайан Т. (1 мая 2006 г.). «Влияние световой акклиматизации на фотосинтез, рост и распределение биомассы у сеянцев американского каштана (Castanea dentata)» . Лесная экология и управление . 226 (1): 173–180. дои : 10.1016/j.foreco.2005.12.063 . ISSN 0378-1127 .
- ^ Перейти обратно: а б Чу, Селия К.; Коулман, Джеймс С.; Муни, Гарольд А. (1 апреля 1992 г.). «Контроль распределения биомассы между корнями и побегами: обогащение атмосферы CO2, а также приобретение и распределение углерода и азота в диком редисе». Экология . 89 (4): 580–587. Бибкод : 1992Oecol..89..580C . дои : 10.1007/BF00317167 . ISSN 1432-1939 . ПМИД 28311891 . S2CID 24092595 .
- ^ Бернакки, CJ; Коулман, Дж.С.; Баззаз, ФА; МакКоннохей, KDM (2000). «Распределение биомассы однолетних видов на старых полях, выращиваемых в условиях повышенного содержания CO2: нет доказательств оптимального разделения». Биология глобальных изменений . 6 (7): 855–863. Бибкод : 2000GCBio...6..855B . дои : 10.1046/j.1365-2486.2000.00370.x . ISSN 1365-2486 . S2CID 85074078 .
- ^ Перейти обратно: а б Камбуи, Калифорния; Свеннерштам Х; Граффман Л; Нордин А; Ганетег У; и др. (2011). «Схемы распределения растительной биомассы зависят от источника азота» . ПЛОС ОДИН . 6 (4): e19211. Бибкод : 2011PLoSO...619211C . дои : 10.1371/journal.pone.0019211 . ПМК 3081341 . ПМИД 21544211 .
- ^ Брекл, Южный-Западный; Кале, Х. (1 июля 1992 г.). «Влияние токсичных тяжелых металлов (Cd, Pb) на рост и минеральное питание бука (Fagus sylvatica L.)». Растительность . 101 (1): 43–53. дои : 10.1007/BF00031914 . ISSN 1573-5052 . S2CID 25502214 .
- ^ Сюй, Чжэньчжу; Чжоу, Гуаншэн; Симидзу, Хидеюки (01 июня 2010 г.). «Реакция растений на засуху и полив» . Сигнализация и поведение растений . 5 (6): 649–654. дои : 10.4161/psb.5.6.11398 . ISSN 1559-2324 . ПМК 3001553 . ПМИД 20404516 .
- ^ Ян, Сяочжоу; Чжан, Вэнь-хуэй; Хэ, Цю-юэ (01.01.2019). «Влияние внутривидовой конкуренции на рост, архитектуру и распределение биомассы Quercus Liaotungensis» . Журнал взаимодействия растений . 14 (1): 284–294. дои : 10.1080/17429145.2019.1629656 . ISSN 1742-9145 .
- ^ Вересоглу, Ставрос Д.; Менексес, Джордж; Риллиг, Маттиас К. (01 апреля 2012 г.). «Влияют ли арбускулярные микоризные грибы на аллометрическое распределение биомассы растения-хозяина на побеги и корни? Метаанализ исследований с 1990 по 2010 год». Микориза . 22 (3): 227–235. дои : 10.1007/s00572-011-0398-7 . ISSN 1432-1890 . ПМИД 21710352 . S2CID 17929759 .
- ^ Михаляк, Чарльз А.; Линкольн, Дэвид Э. (1 марта 1989 г.). «Разделение растительной биомассы и химическая защита: реакция на дефолиацию и ограничение нитратов». Экология . 80 (1): 122–126. Бибкод : 1989Oecol..80..122M . дои : 10.1007/BF00789940 . ISSN 1432-1939 . ПМИД 23494354 . S2CID 6606602 .
- ^ Перейти обратно: а б с Пуртер, Хендрик; Ягодзинский, Анджей М.; Руис-Пейнадо, Рикардо; Куя, Сим; Ло, Юньцзянь; Олексин, Яцек; Усольцев Владимир А.; Бакли, Томас Н.; Райх, Питер Б.; Сак, Лоурен (ноябрь 2015 г.). «Как распределение биомассы меняется в зависимости от размера и различается между видами? Анализ 1200 видов растений с пяти континентов» . Новый фитолог . 208 (3): 736–749. дои : 10.1111/nph.13571 . ISSN 0028-646X . ПМК 5034769 . ПМИД 26197869 .
- ^ Перейти обратно: а б Пайхл, Матиас; Араин, М. Альтаф (15 декабря 2007 г.). «Аллометрия и разделение надземной и подземной биомассы деревьев в возрастной последовательности белых сосновых лесов» . Лесная экология и управление . 253 (1): 68–80. дои : 10.1016/j.foreco.2007.07.003 . ISSN 0378-1127 .