Форонид
| Форониды Временной диапазон: | |
|---|---|
 | |
| Форонис сп. | |
 | |
| Форонопсис Хармери | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| (без рейтинга): | Спиралия |
| Супертип: | Лофотрохозоа |
| Клэйд : | Лофофората |
| Клэйд : | Брахиозоа |
| Тип: | Форониды Хачек , 1888 г. |
| Роды | |
| |
Форониды (научное название Phoronida , иногда называемые подковообразными червями ) — небольшой тип морских животных, которые фильтруют пищу с помощью лофофора («короны» из щупалец) и строят вертикальные хитиновые трубки для поддержки и защиты своих мягких тел. Они обитают в большинстве океанов и морей, включая Северный Ледовитый океан, но исключая Антарктический океан , а также между приливной зоной и на глубине около 400 метров. Большинство взрослых форонид имеют длину 2 см и ширину около 1,5 мм, хотя длина самых крупных из них составляет 50 см.
Название группы происходит от ее типового рода : Phoronis . [2] [3]
Обзор
[ редактировать ]Нижний конец тела представляет собой ампулу (припухлость, похожую на колбу), которая удерживает животное в трубке и позволяет ему очень быстро втягивать свое тело при угрозе. Когда лофофор вытянут в верхней части тела, реснички (волоски) по бокам щупалец притягивают частицы пищи ко рту, который находится внутри и немного сбоку от основания лофофора. Нежелательный материал можно исключить, закрыв крышку над ртом, или отторгнуть щупальцами, реснички которых могут переключаться в реверс. Затем пища перемещается вниз в желудок, который находится в ампуле. Твердые отходы перемещаются вверх по кишечнику и наружу через задний проход , который находится снаружи и немного ниже лофофора.
Кровеносный сосуд идет вверх по середине тела от желудка к круглому сосуду у основания лофофора, а оттуда вверх по каждому щупальцу проходит одиночный слепой сосуд. Пара кровеносных сосудов у стенки тела ведет вниз от лофофорного кольца к желудку, а также к слепым ветвям по всему телу. Сердца нет, но основные сосуды могут волнообразно сокращаться, обеспечивая движение крови. Форониды не вентилируют свои туловища насыщенной кислородом водой, а полагаются на дыхание через лофофор. Кровь содержит гемоглобин , что необычно для таких мелких животных и, по-видимому, является адаптацией к бескислородной и гипоксической среде. Кровь Phoronis Architecta несет в два раза больше кислорода , чем кровь человека того же веса. Две метанефридии фильтруют жидкость организма, возвращая все полезные продукты и выбрасывая оставшиеся растворимые отходы через пару пор рядом с анусом.
Один вид строит колонии путем почкования или деления на верхнюю и нижнюю части, а все форониды размножаются половым путем с весны до осени. Из яиц большинства видов образуются свободноплавающие личинки актинотрох, питающиеся планктоном. Актинотрох оседает на морское дно примерно через 20 дней, а затем за 30 минут претерпевает радикальные изменения: личиночные щупальца заменяются взрослым лофофором; анус перемещается снизу сразу за пределы лофофора; и это меняет кишку с вертикального на U-образный изгиб, при этом желудок оказывается в нижней части тела. Один вид образует «слизневидную» личинку, личинки некоторых видов не известны. Форониды живут около года.
Некоторые виды живут отдельно, в вертикальных трубках, погруженных в мягкий осадок , тогда как другие образуют запутанные массы, погребенные в камнях и ракушках или инкрустирующие их. Виды, способные проникать в такие материалы, как известняк и мертвые кораллы, делают это с помощью химических выделений. [4] В некоторых местообитаниях популяции форонид достигают десятков тысяч особей на квадратный метр. Личинки актинотрох широко распространены среди планктона и иногда составляют значительную долю биомассы зоопланктона. Хищники включают рыбу, брюхоногих моллюсков (улиток) и нематод (крошечных круглых червей). Один вид форонид неприятен для многих эпибентических хищников. различными паразитами Полости тела, пищеварительный тракт и щупальца форонид поражаются . Неизвестно, имеют ли форониды какое-либо значение для человека. Международный союз охраны природы (МСОП) не включил ни один вид форонид в список находящихся под угрозой исчезновения.
По состоянию на 2010 год бесспорных окаменелостей тел форонид нет. [5] Имеются веские доказательства того, что форониды создали следы окаменелостей, обнаруженных в силурийском , девонском , пермском , юрском и меловом периодах , и, возможно, в ордовике и триасе . Форониды, брахиоподы и мшанки (эктопрокты) вместе называются лофофоратами , поскольку все они используют лофофоры для питания. Примерно с 1940-х по 1990-е годы генеалогические древа , основанные на эмбриологических и морфологических особенностях, помещали лофофораты среди или в качестве сестринской группы вторичноротых включает , супертипа, который хордовых и иглокожих . Хотя меньшинство придерживается этой точки зрения, большинство исследователей теперь считают форониды членами протостомов супертипа Lophotrochozoa . [6] Хотя аналитики, использующие молекулярную филогению, уверены, что члены Lophotrochozoa более тесно связаны друг с другом, чем те, кто не являются членами, отношения между членами по большей части неясны. Некоторые анализы рассматривают форониды и брахиоподы как сестринские группы , в то время как другие помещают форониды как подгруппу внутри брахиопод. [7]
Сравнение похожих типов
[ редактировать ]| Особенность | Форониды [8] | Брахиоподы [9] | Мшанки [10] | Энтопрокты [11] |
|---|---|---|---|---|
| Щупальца полые | Да | Да | Да | Нет |
| Защита и поддержка | трубка Прямая хитиновая | Корпус с двумя клапанами | Различные, включая хитин, минерализованные скелеты, растительные формы и массу студенистого материала. | Никто |
| Поток подачи | Сверху вниз | Через стороны корпуса, наружу через переднюю часть | Сверху вниз | Снизу вверх |
| Анус | Внешнее кольцо щупалец | Мантии . или нет, и твердые отходы выбрасываются изо рта [12] | Внешнее кольцо щупалец | Внутреннее кольцо щупалец |
| Колониальный | Один вид | Нет | Все, кроме одного рода | Большинство видов колониальные |
| Целом | Да | Да | Да | Нет |
Описание
[ редактировать ]Строение тела
[ редактировать ]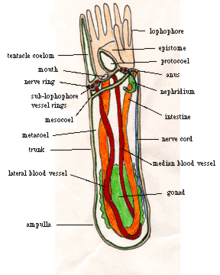
Большинство взрослых форонид имеют длину от 2 до 20 см и ширину около 1,5 мм. [8] хотя самые крупные имеют длину 50 см. [13] Их кожа не имеет кутикулы , но выделяет жесткие хитиновые трубки . [8] аналогичен материалу, используемому членистоногих экзоскелетах в , [14] а иногда усилены частицами осадка и другим мусором. [1] Трубки большинства видов прямые, но у Phoronis vancouverensis горизонтальные и запутанные. [15] Форониды могут перемещаться внутри своих трубок, но никогда не покидают их. [8] Нижний конец тела представляет собой ампулу (колбовидное вздутие в трубчатой структуре). [16] ), [8] который фиксирует животное в трубке и позволяет ему втянуть тело в случае угрозы, [13] уменьшение тела до 20 процентов от его максимальной длины. [1] Продольные мышцы очень быстро втягивают тело, а круговые медленно разгибают тело за счет сжатия внутренней жидкости. [13]
Для питания и дыхания каждый форонид имеет на верхнем конце лофофор — «корону» из щупалец, с помощью которых животное фильтрует пищу . У мелких видов «корона» представляет собой простой круг, у средних видов она изогнута в форме подковы с щупальцами на внешней и внутренней сторонах, а у самых крупных видов концы подковы закручиваются в сложные спирали. Эти более сложные формы увеличивают площадь, доступную для питания и дыхания . [8] Щупальца полые, удерживаются в вертикальном положении под давлением жидкости и могут перемещаться индивидуально с помощью мышц. [13]
Рот находится внутри основания короны щупалец, но сбоку. Кишка проходит ото рта к одной стороне желудка , в нижней части ампулы. Кишечник идет от желудка вверх по другой стороне тела и выходит у заднего прохода снаружи и немного ниже макушки щупалец. Кишечник и кишечник поддерживаются двумя брыжейками (перегородками, проходящими по всей длине тела), соединенными со стенкой тела, а еще одна брыжейка соединяет кишечник с кишечником. [8]
Тело разделено на целомы , [8] отсеки выстланы мезотелием . [17] Основная полость тела под коронкой щупалец называется метацелом , а щупальца и их основание разделяют мезоцелом . [8] Над ртом находится эпистома — полая крышка, закрывающая рот. [13] Полость в эпистоме иногда называют протоцелом, хотя другие авторы не согласны с тем, что это целом. [18] а Руперт, Фокс и Барнс считают, что он строится по другому процессу. [8]
Трубка состоит из трехслойного органического внутреннего цилиндра и агглютинированного внешнего слоя. [19]
Питание, кровообращение и выведение
[ редактировать ]Когда лофофор вытянут, реснички (маленькие волоски) по бокам щупалец вытягивают воду вниз между щупальцами и наружу у основания лофофора. Более короткие реснички на внутренних сторонах щупалец отбрасывают частицы пищи в бороздку по кругу под и внутри щупалец, а реснички в бороздке выталкивают частицы в рот. [13] Форониды направляют свои лофофоры в течение воды и быстро меняют ориентацию, чтобы максимально увеличить площадь захвата пищи при изменении течения. В их рацион входят водоросли , диатомовые водоросли , жгутиконосцы , перидинии , мелкие личинки беспозвоночных и детрит. [1] Нежелательный материал можно исключить, закрыв эпистому (веку над ртом) или отторгнуть щупальцами, реснички которых могут переключаться в реверс. Кишечник использует реснички и мышцы для перемещения пищи к желудку и выделяет ферменты, которые переваривают часть пищи, но желудок переваривает большую часть пищи. [13] Форониды также поглощают аминокислоты (строительные блоки белков). [20] ) через шкуру, в основном летом. [1] Твердые отходы перемещаются вверх по кишечнику и наружу через задний проход , который находится снаружи и немного ниже лофофора. [21]
Кровеносный сосуд [8] начинается от брюшины ( оболочки , рыхло окружающей желудок), [13] со слепыми капиллярами, снабжающими желудок. [8] Кровеносный сосуд ведет вверх по середине тела к круглому сосуду у основания лофофора, а оттуда вверх по каждому щупальцу идет одиночный слепой сосуд. Пара кровеносных сосудов у стенки тела ведет вниз от лофофорного кольца, и у большинства видов они объединены в один немного ниже лофофорного кольца. Нисходящий сосуд(ы) ведет обратно в брюшину, а также к слепым ветвям по всему телу. Сердца нет, но мышцы главных сосудов волнообразно сокращаются, обеспечивая движение крови. [13] В отличие от многих животных, живущих в трубках, форониды не вентилируют свой хобот насыщенной кислородом водой, а полагаются на дыхание лофофором, простирающимся над гипоксическими отложениями. В крови есть гемоциты, содержащие гемоглобин , что необычно для таких мелких животных и, по-видимому, является адаптацией к бескислородной и гипоксической среде. Кровь Phoronis Architecta несет столько же кислорода на см3. 3 как у большинства позвоночных ; объем крови в см 3 на грамм массы тела в два раза больше, чем у человека. [8]
Подоциты на стенках кровеносных сосудов осуществляют первую стадию фильтрации растворимых отходов в основную целомовую жидкость. Две метанефридии , каждая с воронкообразным приемным отверстием, вторично фильтруют жидкость, [8] возвращение любых полезных продуктов целому [22] и сброс оставшихся отходов через пару нефридиопоров рядом с анусом. [8]
Нервная система и движение
[ редактировать ]Между ртом и анусом находится нервный центр, а у основания лофофора — нервное кольцо. [1] Кольцо снабжает нервами щупальца и под кожей мышцы стенок тела. Phoronis ovalis имеет под кожей два нервных ствола, тогда как у других видов - один. [8] Туловище(я) имеет гигантские аксоны (нервы, передающие сигналы очень быстро), которые координируют втягивание тела при угрозе опасности. [13]
За исключением втягивания тела в трубку, форониды обладают ограниченным и медленным движением: частичный выход из трубки; сгибание тела при разгибании; и попадание еды в рот лофофором. [8]
Размножение и жизненный цикл
[ редактировать ]Только самый маленький вид подковообразных червей, Phoronis ovalis , естественным образом строит колонии путем почкования или деления на верхнюю и нижнюю части, которые затем вырастают в полноценные тела. В экспериментах другие виды успешно разделились, но только тогда, когда обе части имеют достаточное количество половых желез (репродуктивных [23] ) салфетка. [24] Все форониды размножаются половым путем с весны до осени. Некоторые виды являются гермафродитами (имеют как мужские, так и женские репродуктивные органы). [25] ), но перекрестно оплодотворять (оплодотворять яйца других членов [26] ), а другие раздельнополы (имеют раздельнополые [27] ). [1] Гаметы ) ( сперматозоиды и яйцеклетки [28] ) производятся в опухших половых железах вокруг желудка. [8] Гаметы плывут через метацелом к метанефридиям. [13] Сперматозоиды выходят через нефридиопоры, а часть захватывается лофофорами особей того же вида. Виды, откладывающие мелкие оплодотворенные яйца, выпускают их в воду в виде планктона. [1] в то время как виды с более крупными яйцами высиживают их либо в трубке тела, либо приклеивают к центру лофофора с помощью клея. [13] Высиженные яйца выпускаются для питания планктоном, когда они развиваются в личинки. [1]
Развитие яиц представляет собой смесь характеристик вторичноротых и протостомов . Ранние деления яйцеклетки голобластические (клетки делятся полностью) и радиальные (постепенно образуют стопку кружков). Процесс носит регулятивный характер (судьба каждой клетки зависит от взаимодействия с другими клетками, а не от жесткой программы в каждой клетке), и в экспериментах по делению ранних эмбрионов рождались полноценные личинки. Мезодерма образуется из мезенхимы, происходящей от архентерона .Целом формируется шизоцелией , а бластопор (вмятина на зародыше) становится ртом. [8]

Слизнеобразная личинка Phoronis ovalis , единственного известного вида с лецитотрофными (не питающимися) личинками, лишена щупалец и плавает около 4 дней, 3–4 дня ползает по морскому дну, а затем проникает в карбонатное дно. [29] [30] [31] О трех видах ничего не известно. У остальных видов развиваются свободноплавающие личинки актинотрох , питающиеся планктоном. Актинотрох представляет собой вертикальный цилиндр с анальным отверстием внизу и окаймлен ресничками. Вверху находится лопасть [1] или капюшон, под которым находятся: ганглий, соединенный с участком ресничек за пределами вершины капюшона; [8] пара протонефридиев (меньше и проще метанефридиев у взрослого); [1] рот; и кормящие щупальца, окружающие рот. [8] После плавания около 20 дней актинотрох оседает на морском дне и за 30 минут претерпевает катастрофическую метаморфозу (радикальное изменение): капюшон и личиночные щупальца рассасываются, а из метасомального мешка личинки формируется ювенильное тело.Взрослый лофофор создается вокруг рта, и за счет роста вентральной стороны, которая чрезвычайно длинна по сравнению со спинной стороной, в кишечнике образуется U-образный изгиб, так что анус оказывается сразу под лофофором и снаружи. [1] Наконец взрослый форонид строит трубку. [8] [32]
Форониды живут около года. [1]
Экология
[ редактировать ]
Форониды обитают во всех океанах и морях, включая Арктику. [33] и исключая Антарктический океан , [1] и появляются между приливной зоной и примерно на 400 метров ниже. Некоторые из них встречаются отдельно, в вертикальных трубках, погруженных в мягкие отложения, такие как песок, грязь или мелкий гравий. Другие образуют запутанные массы из множества особей, погребенных в камнях и ракушках или инкрустирующих их. В некоторых местообитаниях популяции форонид достигают десятков тысяч особей на квадратный метр. Личинки актинотрохов известны среди планктона . [1] и иногда составляют значительную долю биомассы зоопланктона. [34]
Phoronis australis просверливает стенку трубки анемоны Ceriantheomorphe цериантидной brasiliensis и использует ее как основу для построения собственной трубки. В одном цериантиде может находиться до 100 форонид. В этом неравном отношении анемон не испытывает ни существенной пользы, ни вреда, тогда как форонид получает пользу от: основания для своей трубки; пища (оба животных являются фильтраторами); и защита, поскольку цериантид прячется в свою трубку, когда угрожает опасность, и это предупреждает форонид о необходимости втянуться в свою собственную трубку. [35]
Хотя хищники форонид малоизвестны, к ним относятся рыбы, брюхоногие моллюски (улитки) и нематоды (мельчайшие круглые черви). [1] Phoronopsis viridis , плотность которого достигает 26 500 особей на квадратный метр на приливных отмелях в Калифорнии (США), неприятен для многих эпибентических хищников, включая рыб и крабов. Невкусность наиболее сильна в верхней части, включая лофофор, который подвергается воздействию хищников при питании форонид. Когда лофофоры были удалены в эксперименте, форониды были более вкусными, но этот эффект уменьшался в течение 12 дней по мере регенерации лофофоров. Эта широко эффективная защита, которая кажется необычной среди беспозвоночных, населяющих мягкие отложения, может быть важна для того, чтобы позволить Phoronopsis viridis достичь высокой плотности. [36] Некоторые паразиты заселяют форониды: прогенетические метацеркарии и цисты трематод форонид в целомических полостях ; неопознанные грегарины в пищеварительном тракте форонид; и анцистрокомидный реснитчатый паразит Heterocineta в щупальцах. [1]
Неизвестно, имеют ли форониды какое-либо значение для человека. Международный союз охраны природы (МСОП) не включил ни один вид форонид в список находящихся под угрозой исчезновения. [1]
Эволюционная история
[ редактировать ]Ископаемая запись
[ редактировать ]
По состоянию на 2016 год бесспорных окаменелостей тел форонид нет. [5] нижнего кембрия Исследуя окаменелости Чэнцзян , в 1997 году Чен и Чжоу интерпретировали Iotuba chengjiangensis как форонид, поскольку у него были щупальца и U-образный кишечник. [38] а в 2004 году Чен интерпретировал Eophoronis как форонид. [39] Однако в 2006 году Конвей Моррис рассматривал Iotuba и Eophoronis как синонимы одного и того же рода, который, по его мнению, был похож на приапулид Louisella . [40] В 2009 году Бальтазар и Баттерфилд нашли в западной Канаде два экземпляра нового ископаемого Lingulosacculus nuda , датированного примерно 505 миллионами лет назад, у которого были две раковины, подобные раковинам брахиопод , но не минерализованные . По мнению авторов, U-образная кишка выходила за пределы замочной линии и за пределы меньшей раковины. Это препятствовало бы прикреплению мышц для закрытия и открытия раковин, а 50% длины животного за пределами шарнирной линии потребовались бы продольные мышцы, а также кутикула для защиты. Следовательно, они предполагают, что Lingulosacculus мог быть членом группы стволов форонид внутри язычкообразных брахиопод. [41] Другая альтернатива заключается в том, что Eccentrotheca [42] [43] лежит где-то в линии стебля форонид.
Имеются веские доказательства того, что виды , которые были найдены в Phoronis создали следы окаменелостей ихногена Talpina девонском , юрском и меловом периодах . [44] Животное Тальпина бурило известковые водоросли , кораллы , раковины) ежовых рыб раковины , раковины моллюсков и ростры белемнитов ( . [45] Гедереллиды или гедереллоиды представляют собой окаменелые трубки, обычно изогнутые и шириной от 0,1 до 1,8 мм, встречающиеся от силура до перми и, возможно, в ордовике и триасе . Их ветвящиеся колонии могли быть созданы форонидами. [37]
Генеалогическое древо
[ редактировать ]Форониды, брахиоподы и мшанки (эктопрокты) вместе называются лофофоратами , поскольку все они питаются лофофорами. [6] Примерно с 1940-х по 1990-е годы генеалогические древа, на эмбриологических и морфологических особенностях, помещали лофофораты среди или в качестве сестринской группы вторичноротых основанные . [7] супертип, включающий хордовых и иглокожих . На раннем этапе развития эмбрионов вторичноротые формируют анус раньше рта, а протостомы сначала формируют рот. [46]
Нильсен (2002) считает, что форониды и брахиоподы связаны с крыложаберными вторичноротыми , которые также фильтруют пищу с помощью щупалец, поскольку токоведущие клетки лофофоров всех трех имеют по одной ресничке на клетку, в то время как лофофоры мшанок , которых он считает как протостомы, имеют несколько ресничек на клетку. [47] Хельмкампф, Бруххаус и Хаусдорф (2008) резюмируют эмбриологический и морфологический анализ нескольких авторов, которые сомневаются или не согласны с тем, что форониды и брахиоподы являются вторичноротыми: [6]
- В то время как вторичноротые имеют три целомические полости, лофофораты, такие как форониды и брахиоподы, имеют только две. [18]
- Крыложаберные могут быть подгруппой энтеропнеустов («желудевых червей»). Это говорит о том, что предковый вторичноротый больше похож на подвижный червеобразный энтеропнеуст, чем на сидячую колониальную крыложаберность. Тот факт, что и лофофораты, и крыложабры используют щупальца для питания, вероятно, не является синапоморфией лофофоратов и вторичноротых, а развился независимо как конвергентная адаптация к сидячему образу жизни. [6] [48] [49]
- Мезодерма не образуется путем энтероцелии у форонид и мшанок, но образуется у вторичноротых, при этом существуют разногласия относительно того, образуют ли брахиоподы мезодерму путем энтероцелии. [6]
Начиная с 1988 года анализ, основанный на молекулярной филогении , который сравнивает биохимические особенности, такие как сходство в ДНК , поместил форониды и брахиоподы в число Lophotrochozoa , супертипа протостомов, который включает моллюсков , кольчатых червей и плоских червей , но исключает другой основной супертип протостомов Ecdysozoa. , к числу членов которого относятся членистоногие . [6] [7] Коэн писал: «Если этот вывод верен, он подрывает практически все морфологические реконструкции филогении, сделанные за последнее столетие или даже больше». [54]
Хотя анализ молекулярной филогении дает уверенность в том, что члены Lophotrochozoa более тесно связаны друг с другом, чем те, кто не являются членами, отношения между членами по большей части неясны. [7] [55] Lophotrochozoa Lophophorata обычно делятся на: (животные , имеющие лофофоры), включая Phoronida и Brachiopoda; Trochozoa (животные, многие из которых имеют личинки трохофоров ), включая моллюсков, кольчатых червей , эхиуранов , сипункуланов и немертин ; и некоторые другие типы (такие как Platyhelminthes , Gastrotricha , Gnathostomulida , Micrognathozoa и Rotifera ). [7]
Молекулярная филогения указывает на то, что Phoronida тесно связаны с Brachiopoda, но Bryozoa (Ectoprocta) не имеют близкого родства с этой группой, несмотря на то, что используют аналогичный лофофор для питания и дыхания. [55] [56] [57] Это означает, что традиционное определение «Lophophorata» не является монофилетическим . В последнее время термин «Lophophorata» стал применяться только к Phoronida и Brachiopoda, и Халанич (2004) считает, что это изменение вызовет путаницу. [7] Некоторые анализы рассматривают Phoronida и Brachiopoda как сестринские группы, в то время как другие помещают Phoronida как подгруппу внутри Brachiopoda. [7] подразумевая, что Brachiopoda парафилетичны . [58] Анализ Коэна и Вейдмана (2005) приходит к выводу, что форониды представляют собой подгруппу нечленораздельных брахиопод (тех, у которых шарнир между двумя створками не имеет зубов и гнезд). [9] ) и сестринская группа других нечленораздельных подгрупп. Авторы также предполагают, что предки моллюсков и клады брахиопод+форонид разошлись между 900 и 560 млн лет назад, наиболее вероятно около 685 млн лет назад. [57]
Таксономия
[ редактировать ]| Взрослые виды [1] | Виды личинок [1] |
|---|---|
| Форонис овалис | (ползучая личинка) |
| Форонис гиппокрепия | Актинотроха гиппокрепия [30] |
| Форонис иджимай , также называемый Форонис Ванкуверенсис | Актинотроха Ванкуверская |
| Форонис южный | (неизвестный) |
| Форонис муллери | Актинотроха жаберная |
| Форонис псаммофила | Актинотроха сабатьери |
| Форонис бледный | Актинотроха бледная |
| Форонопсис альбомакулата | (неизвестный) |
| Форонопсис Хармери | Актинотроха Хармери |
| Форонопсис калифорнийский | (неизвестный) |
В типе два рода , без названий классов и отрядов . Зоологи дали личинкам, обычно называемым актинотрохами , отдельное от взрослых название рода. [1]
В 1999 году Темерева и Малахов описали Phoronis svetlanae . [59] В 2000 году Темерева описала новый вид — Phoronopsis malahovi . [60] в то время как Эмиг считает его синонимом Phoronopsisharmeri . [1] Сантагата считает, что Phoronis Architecta — это вид, отличный от Phoronis psammophila и Phoronis muelleri , и что «видовое разнообразие [форонид] в настоящее время недооценивается». [61] В 2009 году Темерева описала, что может быть личинками Phoronopsis albomaculata и Phoronopsiscalifornica . Она написала, что, хотя существует 12 бесспорных видов взрослых форонид, идентифицировано 25 морфологических типов личинок. [34]
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х Эмиг, Кристиан К. (2003). «Тип: Форониды» (PDF) . В Бернхарде Гржимеке; Девра Г. Клейман; Майкл Хатчинс (ред.). Энциклопедия жизни животных Гржимека . Том. 2: Перворотые (2-е изд.). Томсон Гейл. стр. 491–495. ISBN 978-0-7876-5362-0 . Архивировано из оригинала (PDF) 27 марта 2010 г. Проверено 1 марта 2011 г.
- ^ «Форонида» . Словарь Merriam-Webster.com . «Новая латынь, от Форонис + -ида ».
- ^ «Форонис» . Словарь Merriam-Webster.com . «Новая латынь, вероятно, от латинского Phoronis ( Ио , мифическая жрица Аргоса , любимая Зевсом)».
- ^ Коралловые рифы Персидского залива: адаптация к экстремальным климатическим условиям
- ^ Jump up to: а б Тейлор, Пол Д.; Олев Винн ; Марк А. Уилсон (2010). «Эволюция биоминерализации в лофофоратах » . Специальные статьи по палеонтологии . 84 : 317–333.
- ^ Jump up to: а б с д и ж Хельмкампф, Мартин; Ирис Бруххаус; Бернхард Хаусдорф (август 2008 г.). «Филогеномный анализ лофофоратов (брахиопод, форонид и мшанок) подтверждает концепцию Lophotrochozoa» . Труды Королевского общества Б. 275 (1645): 1927–1933. дои : 10.1098/rspb.2008.0372 . ПМК 2593926 . ПМИД 18495619 .
- ^ Jump up to: а б с д и ж г Галаныч, К.М. (15 декабря 2004 г.). «Новый взгляд на филогению животных» (PDF) . Ежегодный обзор экологии, эволюции и систематики . 35 : 229–256. doi : 10.1146/annurev.ecolsys.35.112202.130124 . Проверено 9 марта 2011 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х Руперт Э.Э., Фокс Р.С. и Барнс Р.Д. (2004). «Лофофората: Форонида». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 817–821 . ISBN 978-0-03-025982-1 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Руперт Э.Э., Фокс Р.С. и Барнс Р.Д. (2004). «Брахиопода». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 821–829 . ISBN 978-0-03-025982-1 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Руперт, Э.Э.; Фокс, РС; Барнс, Р.Д. (2004). «Мшанка». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 829–845 . ISBN 978-0-03-025982-1 .
- ^ Руперт Э.Э., Фокс Р.С. и Барнс Р.Д. (2004). «Камптозоа и циклиофора». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 808–812 . ISBN 978-0-03-025982-1 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Доэрти, Пи Джей (2001). «Лофофораты». В Андерсоне, Д.Т. (ред.). Зоология беспозвоночных (2-е изд.). Издательство Оксфордского университета. стр. 356–363. ISBN 978-0-19-551368-4 .
- ^ Jump up to: а б с д и ж г час я дж к л м Доэрти, Пи Джей (1998). «Лофофораты - Phoronida, Brachiopoda и Ectoprocta». В Д.Т. Андерсоне (ред.). Зоология беспозвоночных (1-е изд.). Издательство Оксфордского университета, Австралия. стр. 346–349. ISBN 978-0-19-553941-7 .
- ^ Руперт Э.Э., Фокс Р.С. и Барнс Р.Д. (2004). «Членистоногие». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 518 . ISBN 978-0-03-025982-1 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хинтон, Сэм (1987). Морская жизнь южной Калифорнии: знакомство с животным миром калифорнийских пляжей к югу от Санта-Барбары . Издательство Калифорнийского университета. ISBN 978-0-520-05924-5 . Проверено 28 августа 2011 г.
- ^ Моррис, Кристофер Г. (1992). «Ампула». Словарь академической прессы по науке и технике . Профессиональное издательство Персидского залива. п. 102. ИСБН 978-0-12-200400-1 .
- ^ Руперт Э.Э., Фокс Р.С. и Барнс Р.Д. (2004). «Компартментализация». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 205–206 . ISBN 978-0-03-025982-1 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Бартоломеус, Т. (февраль 2001 г.). «Ультраструктура и формирование слизистой оболочки полости тела Phoronis muelleri (Phoronida, Lophophorata)». Зооморфология . 120 (3): 135–148. дои : 10.1007/s004350000030 . S2CID 23516505 .
- ^ Темерева, Елена; Щербакова Татьяна; Цетлин, Александр (2020). «Первые данные о строении трубок, образованных форонидами». Зоология . 143 : 125849. doi : 10.1016/j.zool.2020.125849 . ПМИД 33099239 . S2CID 224887177 .
- ^ «Структуры жизни» . Национальный институт общих медицинских наук. Архивировано из оригинала 7 июня 2014 г. Проверено 3 марта 2011 г.
- ^ Маргулис, Линн ; Карлин В. Шварц (1998). «Форонида» . Пять королевств: иллюстрированное руководство по типам жизни на Земле (3-е изд.). Эльзевир. стр. 340–341 . ISBN 978-0-7167-3027-9 . Проверено 27 марта 2011 г.
- ^ Руперт Э.Э., Фокс Р.С. и Барнс Р.Д. (2004). «Введение в билатерию». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр. 212–213 . ISBN 978-0-03-025982-1 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ «Гонады – Определение» . Бесплатный словарь Мерриам-Вебстера . Мерриам-Вебстер, Инкорпорейтед . Проверено 3 марта 2011 г.
- ^ Ринкевич, Барух; Валерия Матранга (2009). «Стволовые клетки в бесполом размножении морских беспозвоночных» . Стволовые клетки в морских организмах . Спрингер. п. 123. ИСБН 978-90-481-2766-5 . Проверено 29 марта 2011 г.
- ^ «Гермафродит – Определение» . Бесплатный словарь Мерриам-Вебстера . Мерриам-Вебстер, Инкорпорейтед . Проверено 3 марта 2011 г.
- ^ «Взаимное оплодотворение – Определение» . Бесплатный словарь Мерриам-Вебстера . Мерриам-Вебстер, Инкорпорейтед . Проверено 10 марта 2011 г.
- ^ «Двудомные – определение» . Бесплатный словарь Мерриам-Вебстера . Мерриам-Вебстер, Инкорпорейтед . Проверено 3 марта 2011 г.
- ^ «Гамета – Определение» . Бесплатный словарь Мерриам-Вебстера . Мерриам-Вебстер, Инкорпорейтед . Проверено 3 марта 2011 г.
- ^ Бейли-Брок, Джули Х.; Кристиан К. Эмиг (2000). «Гавайские форониды (Lophophorata) и их распространение в Тихоокеанском регионе» (PDF) . Тихоокеанская наука . 54 (2): 119–126 . Проверено 11 марта 2011 г.
- ^ Jump up to: а б Эмиг, CC (1982). Дж. Х. С. Блакстер (ред.). Достижения морской биологии . Академическая пресса. стр. 22–23. ISBN 978-0-12-026119-2 . Проверено 12 марта 2011 г.
- ^ Темерева Елена Н.; Цитрин, Евгений Б. (2013). «Развитие, организация и ремоделирование мышц форонид от эмбриона до метаморфоза (Lophotrochozoa: Phoronida)» . Биология развития BMC . 13:14 . дои : 10.1186/1471-213X-13-14 . ПМЦ 3658900 . ПМИД 23617418 .
- ^ Темерева Елена Н.; Малахов, Владимир Владимирович (2015). «Метаморфическое ремоделирование морфологии и полости тела у Phoronopsisharmeri (Lophotrochozoa, Phoronida): эволюция плана тела и жизненного цикла форонид» . Эволюционная биология BMC . 15 (1): 229. Бибкод : 2015BMCEE..15..229T . дои : 10.1186/s12862-015-0504-0 . ПМЦ 4618516 . ПМИД 26489660 .
- ^ Темерева Е.Н.; Малахов В.В.; Яковис Э.Л.; Фокин М.В. (сентябрь – октябрь 2000 г.). « Phoronis ovalis (Phoronida, Lophophorata) в Белом море: первая находка форонид в Арктическом бассейне». Доклады биологических наук . 374 : 523–525. ПМИД 11103334 .
- ^ Jump up to: а б Темерева Е.Н. (2009). «Новые данные о распространении, морфологии и систематике личинок форонид (Lophophorata: Phoronida)» (PDF) . Зоология беспозвоночных . 6 (1): 47–48. дои : 10.15298/invertzool.06.1.05 . Проверено 11 марта 2011 г.
- ^ Стампар, Серхио; Кристиан К. Эмиг; Андре К. Морандини; Гильерме Коджа; Ана Паула Бальбони; Фабио Ланг да Силвейра (2010). «Есть ли какая-либо опасность у симбиотического вида, связанного с видом, находящимся под угрозой исчезновения? Случай червя-форонида, растущего на трубке Ceriantheomorphe » (PDF) . Да. Биол. Мар . 51 : 205–211. Архивировано из оригинала (PDF) 12 ноября 2010 г. Проверено 2 марта 2011 г.
- ^ Ларсон, Эми А.; Джон Дж. Стахович (2008). «Химическая защита форонид, обитающих в мягких осадках, от местных эпибентических хищников» . Серия «Прогресс в области морской экологии» . 374 : 101–111. дои : 10.3354/meps07767 . ISSN 0171-8630 .
- ^ Jump up to: а б Тейлор, PD; Уилсон, Массачусетс (2008). «Морфология и сходство гедереллоидных мшанок» ( PDF) . В Хагемане, С.Дж.; Ки, ММ-младший; Уинстон, Дж. Э. (ред.). Исследования мшанок 2007: Материалы 14-й Международной конференции по мшанке . Музей естественной истории Вирджинии. стр. 301–309. Архивировано из оригинала (PDF) 26 марта 2010 г. Проверено 26 марта 2011 г.
- ^ Чен, JY; Г. Чжоу. «Биология фауны Чэнцзян». Бюллетень Национального музея естественных наук, Тайбэй . 10 :11–105. - цитируется Эмигом (март 2010 г.) и Тейлором, Винном и Уилсоном (2010 г.)
- ^ Чен, JY (2004). Рассвет животного мира (на китайском языке). Нанкин : Jiangsu Science and Technology Press. п. 366. - цитируется Тейлор, Винн и Уилсон (2010).
- ^ Конвей Моррис, С. (29 июня 2006 г.). «Дилемма Дарвина: реалии кембрийского «взрыва» » . Философские труды Королевского общества B: Биологические науки . 361 (1470): 1069–1083. дои : 10.1098/rstb.2006.1846 . ПМЦ 1578734 . ПМИД 16754615 .
- ^ Бальтазар, Уве; Николас Дж. Баттерфилд (2009). «Раннекембрийские брахиоподы с «мягким панцирем» как возможные форониды стеблевой группы» (PDF) . Acta Palaeontologica Polonica . 54 (2): 307–314. дои : 10.4202/app.2008.0042 . S2CID 54072910 . Проверено 8 марта 2011 г.
- ^ Сковстед, CB; Брок, Джорджия; Патерсон-младший; Холмер, Ле; Бадд, GE (2008). «Склеритом Eccentrotheca из нижнего кембрия Южной Австралии: сходство с лофофоратами и значение для филогении томмотиид». Геология . 36 (2): 171–174. Бибкод : 2008Geo....36..171S . дои : 10.1130/g24385a.1 .
- ^ Сковстед, CB; Брок; Топпер, ТП; Патерсон-младший; Холмер, Л.Е. (2011). «Построение склеритома, биофации, биостратиграфия и систематика томмотиид Eccentrotheca helenia sp. nov. из раннего кембрия Южной Австралии». Палеонтология . 54 (2): 253–286. Бибкод : 2011Palgy..54..253S . дои : 10.1111/j.1475-4983.2010.01031.x .
- ^ Бромли, Р.Г. (2004). «Стратиграфия морской биоэрозии» . В Д. Макилрое (ред.). Применение ихнологии к палеоэкологическому и стратиграфическому анализу . Геологическое общество. п. 461. ИСБН 978-1-86239-154-3 . Проверено 12 марта 2011 г.
- ^ Эмиг, Кристиан К. (март 2010 г.). «Ископаемые Форониды и их предполагаемые ихнотаксоны» (PDF) . Геологические книжки . дои : 10.4267/2042/35531 . Проверено 4 марта 2011 г.
- ^ «Введение в дейтеростомию» . Музей палеонтологии Калифорнийского университета . Проверено 8 марта 2010 г.
- ^ Нильсен, К. (июль 2002 г.). «Филогенетическое положение Entoprocta, Ectoprocta, Phoronida и Brachiopoda» . Интегративная и сравнительная биология . 42 (3): 685–691. дои : 10.1093/icb/42.3.685 . ПМИД 21708765 . S2CID 15883626 .
- ^ Кэмерон, CB; Гэри-младший; Свалла, Би Джей (25 апреля 2000 г.). «Эволюция строения тела хордовых: новые данные из филогенетического анализа типов вторичноротых» . Труды Национальной академии наук . 97 (9): 4469–4474. дои : 10.1073/pnas.97.9.4469 . ПМК 18258 . ПМИД 10781046 .
- ^ Халанич, Кеннет М. (февраль 1996 г.). «Конвергенция в пищевых аппаратах лофофоратов и крыложаберных полухордовых, выявленная с помощью 18S рДНК: интерпретация». Биологический бюллетень . 190 (1): 1–5. дои : 10.2307/1542669 . JSTOR 1542669 . ПМИД 29244547 . S2CID 44165536 .
- ^ Гирибет, Гонсало; Эджкомб, Грегори (3 марта 2020 г.). Древо жизни беспозвоночных . Издательство Принстонского университета.
- ^ Марлетаз, Фердинанд; Пейненбург, Катя ТСА; Гото, Тайчиро; Сато, Нориюки; Рохсар, Дэниел С. (2019). «Новая спиральная филогения относит загадочных стреловидных червей к гнатиферам» . Современная биология . 29 (2): 312–318.e3. Бибкод : 2019CBio...29E.312M . дои : 10.1016/j.cub.2018.11.042 . ПМИД 30639106 .
- ^ Лаумер, Кристофер Э.; Беккуш, Николя; Кербл, Александра; Гетц, Фрейя; Невес, Рикардо К.; Соренсен, Мартин В.; Кристенсен, Рейнхардт М.; Хейнол, Андреас; Данн, Кейси В.; Гирибет, Гонсало; Ворсаа, Катрин (2015). «Спиральная филогения определяет эволюцию микроскопических линий» . Современная биология . 25 (15): 2000–2006. Бибкод : 2015CBio...25.2000L . дои : 10.1016/j.cub.2015.06.068 . ПМИД 26212884 .
- ^ Телфорд, MJ; Бурла, С.Дж.; Эконому, А.; Папийон, Д.; Рота-Стабелли, О. (27 апреля 2008 г.). «Эволюция Ecdysozoa» . Философские труды Королевского общества Б. 363 (1496): 1529–1537. дои : 10.1098/rstb.2007.2243 . ПМК 2614232 . ПМИД 18192181 .
- ^ Коэн, Бернард Л. (февраль 2000 г.). «Монофилия брахиопод и форонид: согласование молекулярных данных с классификацией Линнея (подтип Phoroniformea nov.)» . Труды Королевского общества Б. 267 (1440): 225–231. дои : 10.1098/рспб.2000.0991 . ПМК 1690528 . ПМИД 10714876 .
- ^ Jump up to: а б Гирибет, Гонсало (апрель 2008 г.). «Сборка лофотрохозойного (=спирального) древа жизни» . Труды Королевского общества Б. 363 (1496): 1513–1522. дои : 10.1098/rstb.2007.2241 . ПМК 2614230 . ПМИД 18192183 .
- ^ Гэри, Джеймс Р. (июль 2002 г.). «Менее известные таксоны протостомов: введение и дань уважения Роберту П. Хиггинсу» . Интегративная и сравнительная биология . 42 (3): 611–618. дои : 10.1093/icb/42.3.611 . ПМИД 21708757 . S2CID 2117991 .
- ^ Jump up to: а б Коэн, Бернард Л.; Агата Вейдман (1 декабря 2005 г.). «Молекулярные доказательства того, что форониды являются подтаксоном брахиопод (Brachiopoda: Phoronata) и что генетическая дивергенция типов многоклеточных животных началась задолго до раннего кембрия» (PDF) . Организмы, разнообразие и эволюция . 5 (4): 253–273. Бибкод : 2005ODivE...5..253C . дои : 10.1016/j.ode.2004.12.002 . ISSN 1439-6092 . Проверено 15 марта 2011 г.
- ^ де Роза, Рено (ноябрь – декабрь 2001 г.). «Молекулярные данные указывают на сходство с протостомами брахиопод» . Систематическая биология . 50 (6): 848–859. дои : 10.1080/106351501753462830 . ПМИД 12116636 .
- ^ Темерева Е.Н.; В.В. Малахов (1999). «Новый вид форонид, обитающий в скалах, Phoronis svetlanae (Lophophorata, Phoronida) из Японского моря» . Зоологический журнал . 78 (5): 626–630. ISSN 0044-5134 . Проверено 31 марта 2011 г.
- ^ Темерева Е.Н. (2000). «Новый вид форонид Phoronopsis malakhovi (Lophophorata, Phoronida) из Южно-Китайского моря». Зоологический журнал . 79 (9): 1088–1093.
- ^ Сантагата, Скотт (2009). «Форида Мексиканского залива» . У Дэррила Л. Фелдера; Сильвия А. Эрл (ред.). Мексиканский залив Происхождение, воды и биота: биоразнообразие . Издательство Техасского университета A&M. стр. 1133–1134. ISBN 978-1-60344-094-3 . Проверено 29 марта 2011 г.
Внешние ссылки
[ редактировать ]- ФОРОНИДА
- Мировая база данных Форонид
- Phoronidae - Путеводитель по морскому зоопланктону юго-восточной Австралии , Тасманский институт аквакультуры и рыболовства
- Веб-сайт биоэрозии в колледже Вустера
| Базы данных органов управления : Национальные |
|---|