Неотен
Неотения ( / n i ˈ ɒ t ən i / ), [1] [2] [3] [4] также называется ювенилизацией , [5] Это задержка или замедление физиологического или соматического развития организма, обычно животного. Неотения встречается у современных людей по сравнению с другими приматами. [6] В прогенезе или педогенезе ускоряется половое развитие . [7]
И неотения, и прогенез приводят к педоморфизму. [8] (имеющие типичную детскую форму) или педоморфоз [9] (изменение в сторону типичных для детей форм), тип гетерохронии . [10] Это сохранение у взрослых черт, ранее наблюдавшихся только у молодых. Такое сохранение важно в эволюционной биологии , приручении и эволюционной биологии развития . Некоторые авторы определяют педоморфизм как сохранение личиночных признаков, наблюдаемое у саламандр . [11] [12] [13]
История и этимология
[ редактировать ]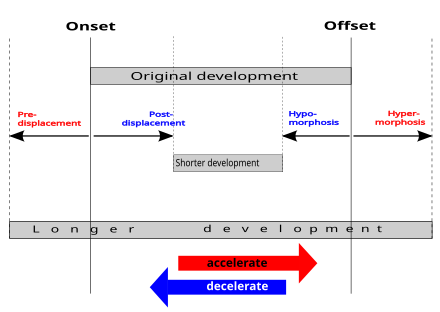
Юлиус Коллманн ввел термин «неотения» в 1885 году после того, как описал взросление аксолотля , оставаясь на водной стадии, напоминающей головастика , с жабрами, в отличие от других взрослых амфибий, таких как лягушки и жабы. [14] [15]
Слово неотения заимствовано из немецкого Neotenie , последнее построено Коллманом из греческого νέος ( неос , «молодой») и τείνειν ( teínein , «растягиваться, расширяться»). Прилагательное . бывает либо «неотеническим», либо «неотеническим» [16] В качестве противоположности слову «неотенический» разные авторитеты используют либо «геронтоморфный», либо «геронтоморфный». [17] [18] или « пераморфный ». [19] Богин указывает, что Коллманн имел в виду «сохранить молодость», но, очевидно, перепутал греческое teinein с латинским tenere , которое имело то значение, которое он хотел, «сохранить», так что новое слово будет означать «сохраняющий молодость». юности (во взрослую жизнь)». [15]
В 1926 году Луи Болк описал неотению как главный процесс гуманизации. [20] [15] В своей книге «Онтогенез и филогения» 1977 года [21] Стивен Джей Гулд отметил, что версия Болка представляет собой попытку оправдать «научный» расизм и сексизм, но признал, что Болк был прав в основной идее о том, что люди отличаются от других приматов тем, что становятся половозрелыми на инфантильной стадии развития тела. [15]
У людей
[ редактировать ]Неотения у людей — это замедление или задержка развития тела по сравнению с приматами , не являющимися людьми , что приводит к таким особенностям, как большая голова, плоское лицо и относительно короткие руки. Эти неотенические изменения могли быть вызваны половым отбором в эволюции человека . В свою очередь, они могли способствовать развитию человеческих способностей, таких как эмоциональное общение. Некоторые эволюционные теоретики предположили, что неотения была ключевой особенностью эволюции человека . [22] Дж.Б.С. Холдейн утверждает, что «основной тенденцией эволюции человека» является «большое продление детства и замедление взросления». [5] Делберт Д. Тиссен сказал, что «неотения становится более очевидной по мере того, как ранние приматы эволюционировали в более поздние формы» и что приматы «эволюционируют в сторону плоского лица». [23] Дуг Джонс утверждал, что тенденция эволюции человека к неотении могла быть вызвана половым отбором в эволюции человека по неотеническим чертам лица у женщин со стороны мужчин, при этом возникающая в результате неотения на мужских лицах была «побочным продуктом» полового отбора для неотенических женских лиц. [24]
У домашних животных
[ редактировать ]Неотения наблюдается у домашних животных, таких как собаки и мыши. [25] Это связано с тем, что доступно больше ресурсов, меньше конкуренции за эти ресурсы, а при снижении конкуренции животные тратят меньше энергии на получение этих ресурсов. Это позволяет им созревать и размножаться быстрее, чем их дикие собратья. [25] Среда, в которой выращиваются домашние животные, определяет, присутствует ли у этих животных неотения. При возникновении этих условий у вида может возникнуть эволюционная неотения, и вид становится половозрелым раньше своего «нормального развития». Другим объяснением неотении у домашних животных может быть отбор по определенным поведенческим характеристикам. Поведение связано с генетикой, что означает, что когда выбирается поведенческий признак, физический признак также может быть выбран из-за таких механизмов, как неравновесие по сцеплению . Часто поведение молодых особей выбирается для того, чтобы легче приручить вид; агрессивность у некоторых видов приходит с взрослением, когда возникает необходимость конкурировать за ресурсы. Если нет необходимости в конкуренции, то нет и необходимости в агрессии. Выбор поведенческих характеристик несовершеннолетних может привести к неотении физических характеристик, потому что, например, при снижении потребности в таком поведении, как агрессия, нет необходимости в развитии черт, которые могли бы помочь в этой области. Признаки, которые могут стать неотенизированными из-за снижения агрессии, могут включать более короткую морду и меньший общий размер у одомашненных особей. Некоторые общие неотенозные физические черты у домашних животных (в основном собак, свиней, хорьков, кошек и даже лисиц) включают висячие уши, изменения репродуктивного цикла, вьющиеся хвосты, пегая окраска, меньший или укороченный позвонок, большие глаза, округлый лоб, большие уши и укороченная морда. [26] [27]

Когда роль собак расширилась от просто рабочих собак до компаньонов , люди начали селекционное разведение собак по морфологической неотении, и это селекционное разведение по «неотении или педоморфизму» «укрепило связь между человеком и собакой». [28] Люди разводили собак, чтобы у взрослых у них было больше «юношеских физических качеств», таких как короткая морда и широко посаженные глаза, которые ассоциируются со щенками, потому что люди обычно считают эти черты более привлекательными. Некоторые породы собак с короткой мордой и широкой головой, такие как комондор , сенбернар и мареммская овчарка, морфологически более неотеничны, чем другие породы собак. [29] Кавалер-кинг-чарльз-спаниели являются примером отбора на неотению, поскольку у них большие глаза, висячие уши и компактные лапы, что придает им морфологию, аналогичную взрослому щенку. [28]
В 2004 году исследование, в котором использовалось 310 черепов волков и более 700 черепов собак, представляющих 100 пород, пришло к выводу, что эволюция черепов собак, как правило, не может быть описана гетерохронными процессами, такими как неотения, хотя некоторые педоморфные породы собак имеют черепа, напоминающие черепа молодых особей. волки. [30] К 2011 году выводы того же исследователя сводились к следующему: «Собаки не являются педоморфными волками». [31]
У других видов
[ редактировать ]
Неотения наблюдалась у многих других видов. Важно отметить разницу между частичной и полной неотенией при рассмотрении других видов, чтобы различать ювенильные признаки, выгодные в краткосрочной перспективе, и черты, полезные на протяжении всей жизни организма; это может дать представление о причине неотении у вида. Частичная неотения - это сохранение личиночной формы после обычного возраста созревания с возможным половым развитием (прогенезом) и возможным созреванием во взрослую форму; это наблюдается у лягушки Lithobates clamitans . Полная неотения наблюдается у Ambystoma mexicanum и некоторых популяций Ambystoma tigrinum , которые остаются в личиночной форме на протяжении всей жизни. [32] [33] Lithobates clamitans частично неотенозный; это задерживает созревание зимой, поскольку доступно меньше ресурсов; в личиночной форме ему легче находить ресурсы. Это охватывает обе основные причины неотении; энергия, необходимая для выживания зимой вновь сформировавшейся взрослой особи, слишком велика, поэтому организм проявляет неотенические характеристики до тех пор, пока он не сможет лучше выжить во взрослом состоянии. По той же причине Ambystoma tigrinum сохраняет неотению; однако сохранение является постоянным из-за отсутствия доступных ресурсов на протяжении всего срока его действия. Это еще один пример экологической причины неотении. Некоторые виды птиц, такие как манакины Chiroxiphia Linearis и Chiroxiphia caudata , демонстрируют частичную неотению. Самцы обоих видов сохраняют юное оперение и во взрослом возрасте, теряя его, когда становятся полностью зрелыми. [34] У некоторых видов птиц сохранение молоди оперения связано со временем линьки у каждого вида. Чтобы гарантировать отсутствие совпадения между временем линьки и спаривания, у птиц может проявляться частичная неотения в оперении; самцы не приобретают яркого взрослого оперения до того, как самки будут готовы к спариванию. Неотения присутствует, потому что самцам нет необходимости рано линять, а попытка спариваться с неполовозрелыми самками была бы энергонеэффективной.
Неотения обычно наблюдается у нелетающих насекомых, например, у самок отряда Strepsiptera . Нелетание насекомых несколько раз развивалось отдельно; Факторами, которые, возможно, способствовали отдельной эволюции нелетаемости, являются большая высота, географическая изоляция (острова) и низкие температуры. [35] В таких условиях окружающей среды расселение было бы невыгодно; В более холодном климате тепло теряется через крылья быстрее. Самки . некоторых групп насекомых становятся половозрелыми без метаморфоза, а у некоторых не развиваются крылья Отсутствие полета у некоторых самок насекомых связано с более высокой плодовитостью . [35] Тля — пример насекомых, у которых в зависимости от окружающей среды могут никогда не появиться крылья. Если на растении-хозяине ресурсов достаточно, нет необходимости отращивать крылья и рассеиваться. Если ресурсы истощаются, у их потомства могут развиться крылья, чтобы распространиться по другим растениям-хозяевам. [36]
Две среды, благоприятствующие неотении, - это большие высоты и прохладные температуры, поскольку неотенические особи обладают большей приспособленностью, чем особи, которые превращаются во взрослую форму. Энергия, необходимая для метаморфоза, снижает индивидуальную приспособленность, и неотенозные особи могут легче использовать доступные ресурсы. [37] Эта тенденция видна при сравнении видов саламандр на более низких и больших высотах; в прохладной высокогорной среде неотенозные особи выживают лучше и более плодовиты, чем те, которые метаморфизируются во взрослую форму. [37] Насекомые в более прохладных условиях, как правило, проявляют неотению в полете, поскольку крылья имеют большую площадь поверхности и быстро теряют тепло; Насекомым невыгодно превращаться во взрослых особей. [35]
Многие виды саламандр и земноводных в целом демонстрируют экологическую неотению. Аксолотль и олм — многожаберные виды саламандр, которые сохраняют свою молодую водную форму на протяжении всей взрослой жизни, что является примером полной неотении. Жабры - обычная особенность молодых особей амфибий, которые сохраняются после созревания; примерами являются тигровая саламандра и грубокожий тритон, оба из которых сохраняют жабры и во взрослом возрасте. [32]
Бонобо имеют много общих физических характеристик с людьми, включая неотенозные черепа. [38] Форма их черепа не меняется во взрослом возрасте (только увеличивается в размерах) из-за полового диморфизма и эволюционного изменения сроков развития. [38] Молодые особи стали половозрелыми до того, как их тела полностью развились во взрослом возрасте, и из-за селективного преимущества неотеническая структура черепа сохранилась. [ нужна ссылка ]
В некоторых группах, таких как семейства насекомых Gerridae , Delphacidae и Carabidae , затраты энергии приводят к неотении; многие виды этих семейств имеют маленькие неотенозные крылья или вообще не имеют их . [36] Некоторые виды сверчков сбрасывают крылья в зрелом возрасте; [39] В роде Ozopemon самцы (считающиеся первым примером неотении у жуков ) значительно меньше самок из-за инбридинга . [40] У термита Kalotermes flavicollis у линяющих самок наблюдается неотения. [41]
У других видов, таких как северо-западная саламандра ( Ambystoma gracile ), условия окружающей среды – в данном случае большая высота – вызывают неотению. [42] Неотения также встречается у нескольких видов ракообразных семейства Ischnomesidae , обитающих в глубоких океанских водах. [43]
Субклеточная неотения
[ редактировать ]Неотения обычно используется для описания развития животных; однако неотения также наблюдается в клеточных органеллах . Было высказано предположение, что субклеточная неотения может объяснить, почему сперматозоиды имеют атипичные центриоли . В одной из двух центриолей сперматозоидов плодовой мухи сохраняется «ювенильная» структура центриоли, которую можно охарактеризовать как центриолярную «неотению». Эта неотеническая атипичная центриоль известна как проксимальная центриольная . Типичные центриоли формируются в результате пошагового процесса, в ходе которого формируется колесо, затем развивается в процентриоль и далее созревает в центриоль. Неотеническая центриоль плодовой мухи напоминает раннюю процентриоль. [44]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «неотения» . Dictionary.com Полный (онлайн). нд . Проверено 21 апреля 2019 г.
- ^ «неотения» . Словарь английского языка американского наследия (5-е изд.). ХарперКоллинз . Проверено 21 апреля 2019 г.
- ^ «неотения» . Лексико американский английский словарь . Издательство Оксфордского университета . Архивировано из оригинала 22 марта 2020 г.
- ^ «неотения» . Словарь Merriam-Webster.com . Проверено 21 апреля 2019 г.
- ^ Перейти обратно: а б Монтегю, А. (1989). Растущая молодость. Бергин и Гарви: Коннектикут.
- ^ Чой, Чарльз К. (1 июля 2009 г.). «Более инфантильность могла привести к большему размеру мозга» . Научный американец .
- ^ Волкенштейн, М.В. 1994. Физические подходы к биологической эволюции . Springer-Verlag: Берлин, [1] .
- ^ «Педоморфный» . 21 января 2022 г.
- ^ «Морфоза» . 6 июня 2022 г.
- ^ Ридли, Марк (1985). Эволюция . Блэквелл.
- ^ Уайтман, Х.Х. (1994). «Эволюция факультативного педоморфоза». Ежеквартальный обзор биологии . 69 (2): 205–221. дои : 10.1086/418540 . S2CID 83500486 .
- ^ Шелл, Справочник SC по трематодам Северной Америки к северу от Мексики , 1985, стр. 22
- ^ Гинецинская, Т. А. Трематоды, их жизненные циклы, биология и эволюция . Ленинград, СССР: Наука, 1968. Перевод 1988 г., [2] .
- ^ Коллманн, Дж. (1885). «Зимовка личинок европейских лягушек и тритонов и трансформация мексиканского аксолотля » . Труды Базельского общества естественных наук (на немецком языке). 7 :387-398. Из стр. 397–398: «Тогда возникает вопрос, является ли латентная стадия яиц, которая, с одной стороны, встречается в таких чрезвычайно удивительных формах у рыб, птиц и млекопитающих, а с другой стороны, латентная стадия у беспозвоночных¹) не является вариантом того же свойства организмов, которое я назвал неотенией и которое может проявиться на любой стадии развития». (Тогда возникает вопрос, не является ли, с одной стороны, латентная стадия яиц — которая встречается в столь удивительных формах у рыб, птиц и млекопитающих — [и] с другой стороны латентная стадия у беспозвоночных ¹) вариантом этого то же свойство организмов, которое я назвал «неотенией» и которое может проявиться на любой стадии развития.)
- ^ Перейти обратно: а б с д Богин, Барри (1999). Закономерности человеческого роста . Издательство Кембриджского университета. стр. 157–169. ISBN 978-0-521-56438-0 .
- ^ Неотения , Бесплатный словарь . 2011. По состоянию на 30 апреля 2011 г.
- ^ Хенке, В. (2007). Справочник по палеоантропологии, Том 1. Springer Books, Нью-Йорк.
- ^ Хетерингтон, Р. (2010). Связь с климатом: изменение климата и современная эволюция человека. Издательство Кембриджского университета.
- ^ Холл, Б.К., Халлгримссон, Б. Монро, WS (2008). Эволюция Стрикбергера: интеграция генов, организмов и популяций. Издательство Jones and Bartlett: Канада.
- ^ Болк, Луи (1926). Проблема воплощения: лекция, прочитанная 15 апреля 1926 г. в XXV. Заседание Анатомического общества Фрайбурга [ Проблема гуманизации: Лекция, прочитанная 15 апреля 1926 года на 25-м конгрессе Анатомического общества во Фрайберге ] (на немецком языке). Йена, Германия: Густав Фишер.
- ^ Гулд, Стивен Джей (1977). Онтогенез и филогения . Кембридж, Массачусетс: Белкнап (издательство Гарвардского университета). ISBN 978-0-674-63940-9 .
- ^ Ши, Брайан Т. (1989). «Гетерохрония в эволюции человека: пересмотр аргументов в пользу неотении». Американский журнал физической антропологии . 32 (С10): 69–101. дои : 10.1002/ajpa.1330320505 .
- ^ Тиссен, Д.Д. (1997). Горько-сладкая судьба: бурная эволюция человеческого поведения. Издательство Транзакций, Нью-Джерси
- ^ Джонс, Д.; и др. (1995). «Половой отбор, физическая привлекательность и неотения лица: межкультурные доказательства и последствия [а также комментарии и ответы]» . Современная антропология . 36 (5): 723–748. дои : 10.1086/204427 . S2CID 52840802 .
- ^ Перейти обратно: а б Прайс, Э. (1999). «Поведенческое развитие животных, подвергающихся приручению». Прикладная наука о поведении животных . 65 (3): 245–271. дои : 10.1016/S0168-1591(99)00087-8 .
- ^ Бертоне, Дж. (2006). Гериатрическая медицина и хирургия лошадей. Сондерс, Мичиган.
- ^ Правда, Л.Н. (1999). «Раннее приручение псовых: эксперимент с лисами на ферме». Американский учёный . 87 (2): 160–169. Бибкод : 1999AmSci..87.....T . дои : 10.1511/1999.2.160 .
- ^ Перейти обратно: а б МакГриви, П.Д. и Николас, ФРВ (1999). Некоторые практические решения проблем благосостояния в собаководстве. В сфере защиты животных. 8: 329–341.
- ^ Бек, AM и Катчер, AH (1996). Между домашними животными и людьми: важность дружеских отношений. Вест-Лафайет, Индиана: Издательство Университета Пердью. ISBN 1-55753-077-7
- ^ Дрейк, Эбби Грейс, «Эволюция и развитие морфологии черепа псовых: исследование морфологической интеграции и гетерохронии» (1 января 2004 г.). Докторские диссертации доступны в Proquest. Бумага ААИ3136721. связь
- ^ Дрейк, Эбби Грейс (2011). «Развенчание собачьих догм: исследование гетерохронии у собак с использованием 3D-геометрического морфометрического анализа формы черепа». Эволюция и развитие . 13 (2): 204–213. дои : 10.1111/j.1525-142X.2011.00470.x . ПМИД 21410876 . S2CID 20893501 .
- ^ Перейти обратно: а б Свингл, В. (1922). «Опыты по метаморфозу неотенозных амфибий» . Журнал экспериментальной зоологии . 36 (4): 397–421. дои : 10.1002/jez.1400360402 .
- ^ «Амбистома тигринум» . Сеть амфибий.
- ^ Фостер, М. (1987). «Задержка созревания, неотения и различия социальных систем у двух манакинов рода Chiroxyphia » . Эволюция . 41 (3): 547–558. дои : 10.2307/2409256 . JSTOR 2409256 . ПМИД 28563802 .
- ^ Перейти обратно: а б с Барбоза, П.; и др. (1989). «Особенности жизни лесных нелетающих чешуекрылых». Американский натуралист из Мидленда . 122 (2): 262–274. дои : 10.2307/2425912 . JSTOR 2425912 .
- ^ Перейти обратно: а б Харрисон, Р. (1980). «Полиморфизмы распространения у насекомых». Ежегодный обзор экологии и систематики . 11 : 95–118. doi : 10.1146/annurev.es.11.110180.000523 . JSTOR 2096904 .
- ^ Перейти обратно: а б Снайдер, Р. (1956). «Сравнительные особенности истории жизни Ambystoma gracile (Baird) из популяций на низких и больших высотах». Копейя . 1956 (1): 41–50. дои : 10.2307/1439242 . JSTOR 1439242 .
- ^ Перейти обратно: а б Ши, BT (1983). «Педоморфоз и неотения у карликового шимпанзе». Наука . 222 (4623): 521–522. Бибкод : 1983Sci...222..521S . дои : 10.1126/science.6623093 . JSTOR 1691380 . ПМИД 6623093 .
- ^ Харрисон, Р. (1980). «Полиморфизмы рассеяния у насекомых». Ежегодный обзор экологии и систематики . 11 : 95–118. doi : 10.1146/annurev.es.11.110180.000523 . JSTOR 2096904 .
- ^ Джордал, Британская Колумбия; Бивер, РА; Нормарк, BB; Фаррелл, Б.Д. (2002). «Чрезвычайное соотношение полов и эволюция мужской неотении у спаривающихся сибсов жуков Ozopemon » . Биологический журнал Линнеевского общества . 75 (3): 353–360. дои : 10.1046/j.1095-8312.2002.00025.x .
- ^ Солтани-Мазуни, Н.; Бордеро, К. (1987). «Изменения кутикулы, яичников и коллетериальных желез во время псевдогейтной и неотенической линьки у Kalotermes flavicollis (FABR.) (Isoptera: Kalotermitidae)». Международный журнал морфологии и эмбриологии насекомых . 16 (3–4): 221–225. дои : 10.1016/0020-7322(87)90022-5 .
- ^ Иглсон, Г.; Маккеун, Б. (1978). «Изменения активности щитовидной железы Ambystoma gracile (Baird) на различных личиночных, трансформирующих и постметаморфических фазах». Канадский журнал зоологии . 56 (6): 1377–1381. дои : 10.1139/z78-190 .
- ^ Брокеленд, В.; Брандт, А. (2004). «Два новых вида Ischnomesidae (Crustacea: Isopoda) из Южного океана, демонстрирующие неотению». Глубоководные исследования. Часть II . 51 (14–16): 1769–1785. Бибкод : 2004DSRII..51.1769B . дои : 10.1016/j.dsr2.2004.06.034 .
- ^ Авидор-Рейсс, Томер; Тернер, Катерина (2019), Клок, Малгожата (ред.), «Эволюция структуры центриолей: гетерохрония, неотения и гиперморфоз» , Аппарат Гольджи и центриоль: функции, взаимодействия и роль в заболеваниях , результаты и проблемы дифференцировки клеток , том. 67, Чам: Springer International Publishing, стр. 3–15, номер номера : 10.1007/978-3-030-23173-6_1 , ISBN. 978-3-030-23173-6 , PMC 7576685 , PMID 31435789 , получено 12 октября 2023 г.
Дальнейшее чтение
[ редактировать ]- Бергсторм, Карл Т. и Дугаткин, Ли Алан (2012). Эволюция , WW Нортон ISBN 039391349X