Страшный материал
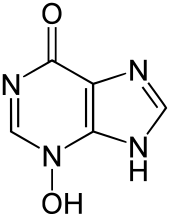
В 1938 году австрийский этолог Карл фон Фриш сделал свое первое сообщение о существовании химического сигнала тревоги, известного как Шрекстофф (вещество страха) у гольянов . Сигнал тревоги — это реакция человека, «отправителя», в ответ на опасность, которая предупреждает других животных — получателей об опасности. [1] Этот химический сигнал тревоги подается только тогда, когда отправитель подвергается механическому повреждению, например, когда его поймал хищник , и обнаруживается обонятельной системой . Когда этот сигнал достигает получателей, они воспринимают больший риск нападения хищников и проявляют реакцию против хищников. Поскольку популяции рыб, обладающих этим признаком, выживают более успешно, этот признак сохраняется посредством естественного отбора . Хотя эволюция этого сигнала когда-то была предметом больших дискуссий, недавние данные свидетельствуют о том, что Шрекстофф эволюционировал как защита от стрессовых факторов окружающей среды, таких как патогены , паразиты и УФ-B- излучение , и что позже он был использован хищниками и добычей в качестве химического сигнала. .
Фон
[ редактировать ]Системы химической сигнализации были идентифицированы у ряда различных таксонов, включая брюхоногих моллюсков , [2] иглокожие , [3] амфибии [4] и рыбы . Одним из наиболее хорошо изученных химических сигналов тревоги является шрекстофф, использование которого широко распространено в надотряде Ostariophys (например, гольяны , харацины , сомы и др.). Около 64% всех видов пресноводных рыб и 27% всех видов рыб во всем мире относятся к надотряду остариофизан, что подчеркивает широкое использование и важность этой системы химической сигнализации у рыб. [5]
Шоковое вещество у остариофизанов
[ редактировать ]Было показано, что производство шрекстоффа требует метаболических затрат и, следовательно, является частью условной стратегии, которую могут использовать только люди, имеющие доступ к достаточным ресурсам. [6] Одним из предполагаемых активных ингредиентов Шрекстоффа является гипоксантин-3N-оксид (H3NO), который может вырабатываться клубными клетками, которые в дальнейшем будут называться «клетками сигнального вещества». [7] Было обнаружено, что функциональная группа оксида азота является основным химическим триггером антихищнического поведения у приемников. [8] Шрекстофф представляет собой смесь, и фрагменты гликозаминогликана , хондроитинсульфата , способны вызывать реакцию страха. [9] Полисахарид-предшественник является компонентом слизи, и предполагается, что его фрагменты образуются во время травмы. Подобно шрекстофу, полученному из экстракта кожи, хондроитинсульфат активирует подмножество обонятельных сенсорных нейронов.
Производство шрекстоффа и реакция на него меняются в ходе онтогенеза . Например, молодые ручьевые колюшки ( Culaea inconstans ) с большей вероятностью попадают в ловушки для гольянов с наживкой из экстрактов кожи конкретного вида, чем взрослые особи. [10] Этот результат показывает, что молодые ручьевые колюшки не так легко связывают шрекстофф и потенциальное присутствие хищника, как взрослые особи. Остается неясным , усиливается ли эта связь с течением времени в результате обучения или физиологического развития. [11]
Помимо изменений в онтогенезе, степень образования шрекстофа варьируется в течение сезона размножения. Самцы толстоголовых гольянов ( Pimephales promelas ) прекращают производство шрекстофа во время сезона размножения, но в это время все еще демонстрируют антихищное поведение в ответ на шрекстофф. [12] В настоящее время производство Schreckstoff может быть остановлено, поскольку самцы толстоголовых гольянов часто получают механические повреждения во время строительства гнезд. Для самца было бы вредно производить шрекстофф во время строительства гнезда, поскольку это непреднамеренно отталкивало бы самок, тем самым снижая вероятность получения партнера. Прекратив производство шрекстофа во время сезона размножения, самцы обходят эту проблему. Прекращение производства сигнальных веществ клетками, по-видимому, контролируется андрогенами . [13]
Гипотезы эволюции шрекстоффа
[ редактировать ]Был предложен ряд различных гипотез эволюции шрекстофа. [14] Первая гипотеза заключается в том, что эволюция шрекстоффов была обусловлена родственным отбором . Подтверждение этой гипотезы может включать доказательства того, что особи живут группами близкородственных родственников и что выпуск химических сигналов тревоги увеличивает вероятность того, что родственные особи будут избегать хищников. Вторая гипотеза, привлечение хищников, предполагает, что выпуск шрекстоффа может привлечь дополнительных хищников, которые будут мешать хищничеству, увеличивая вероятность того, что добыча убежит и переживет нападение. Эта гипотеза предполагает, что шрекстофф будет привлекать хищников, и они будут мешать друг другу либо из-за конкуренции за пойманную добычу, либо из-за хищничества друг друга. Кроме того, предполагается, что, несмотря на то, что жертва уже получила механические повреждения, она может убежать и оправиться от нападения. Тестирование и проверка этих предположений поддержат гипотезу о привлекательности хищников. Третья гипотеза предполагает, что Шрекстофф выполняет иммунную функцию, обеспечивая защиту от патогены , паразиты и/или УФБ- излучение . Для подтверждения этой гипотезы необходимо наблюдать корреляцию между выработкой сигнальных веществ клетками и наличием патогенов и паразитов. Прямые доказательства того, что Шрекстофф подавляет рост водных патогенов и паразитов, дадут дополнительную поддержку гипотезе иммунитета. Другая гипотеза состоит в том, что шрекстофф представляет собой продукт распада слизи и клубневых клеток, вызванный травмой. Выбор реакции на сигнал тревоги осуществляется в первую очередь на уровне получателя.
Гипотеза родственного отбора
[ редактировать ]Одна из первых гипотез эволюции Шрекстоффа была основана на У. Д. Гамильтона . теории родственного отбора [15] Согласно теории родственного отбора, отправитель химического сигнала тревоги был бы готов понести расходы на отправку этого сигнала, если бы выгоды для родственных лиц были достаточно высокими. В ситуации, когда отправитель сигнала платит большие издержки (т. е. он выпускает сигнал химической тревоги, поскольку ему нанесен потенциально смертельный механический ущерб), выгоды для близкородственных родственников должны быть огромными. В рамках родственного отбора выбираются модели поведения, которые кажутся вредными для отправителя, поскольку они приносят пользу индивидуумам, которые, вероятно, имеют общие аллели по общему происхождению . отправителя Таким образом, частота аллелей в следующем поколении увеличивается за счет их присутствия в успешных родственниках.
Чтобы применить теорию родственного отбора к эволюции Шрекстоффа, необходимо выполнить ряд условий. Во-первых, должны существовать доказательства того, что отправитель передает шрекстофф в пользу получателей. Во-вторых, необходимо показать, что особи отряда Ostariophys общаются главным образом с членами семьи. Если любое из этих двух предположений нарушено, гипотеза родственного отбора не будет поддержана.
Существуют некоторые доказательства в поддержку первого предположения о том, что высвобождение шрекстоффа дает измеримые преимущества получателям этого химического сигнала. Лабораторный эксперимент [16] выявили, что толстоголовые гольяны, подвергшиеся воздействию шрекстофа, выживали на 39,5% дольше, чем контрольная группа, когда их помещали в аквариум с хищной северной щукой ( Esox lucius ). Это открытие предполагает, что шрекстофф повышает бдительность получателей, что приводит к сокращению времени реакции после обнаружения хищника.
Второе предположение о том, что особи отряда Ostariophys общаются с близкими членами семьи, по-видимому, не подтверждается эмпирическими данными. В стаях ( европейских гольянов Phoxinus phoxinus ) не было обнаружено различий в родстве внутри косяков и между ними. [17] указывает на то, что люди не общаются более тесно с родственниками, чем с неродственниками. Состав косяков не изучен у всех представителей отряда остариофизан, и еще могут быть обнаружены косяки, состоящие полностью из членов семейства. Тем не менее, открытие того, что производство шрекстоффов сохраняется у видов, у которых эта функция явно не связана с родственными выгодами, дает убедительные доказательства против родственного отбора как механизма эволюции шрекстоффов.
Также было обнаружено, что толстоголовые гольяны производят меньше клеток эпидермального тревожного вещества (и, следовательно, меньше шрекстоффа) в присутствии знакомых товарищей по мелководью. [18] Результаты этого исследования указывают на один из двух сценариев, ни один из которых не совместим с гипотезой о том, что Шрекстофф развился путем родственного отбора. Во-первых, если бы Шрекстофф эволюционировал путем родственного отбора, можно было бы ожидать, что в присутствии родственных клеток будет производиться больше клеток эпидермального сигнального вещества, чем неродственных. Это означает, что знакомые косяки толстоголовых гольянов должны быть близкородственными родственниками, а производство шрекстоффа должно быть увеличено, когда они находятся в косяках со знакомыми особями. Исследование не показало, что это так. [19] Во-вторых, эти результаты показывают, что особи либо вообще не общаются со своими сородичами, либо производство шрекстоффа варьируется в зависимости от того, насколько рыба знакома с особями, с которыми она стайивается. В заключение, данные не подтверждают гипотезу о том, что шрекстофф развился потому, что он усилил инклюзивную приспособленность отправителя за счет увеличения выживаемости родственников.
Гипотеза аттрактанта хищников
[ редактировать ]Гипотеза аттрактанта хищников предполагает, что основная цель шрекстоффа — привлечь в этот район дополнительных хищников. [20] Согласно этой гипотезе, с первоначальным хищником будут взаимодействовать дополнительные хищники, и эти взаимодействия предоставят отправителю возможность спастись. Для подтверждения этой гипотезы необходимо соблюдение ряда условий. Во-первых, шрекстофф должен привлекать хищников. Во-вторых, последующие хищники должны прервать событие хищничества, тем самым увеличивая вероятность того, что жертва сбежит. В-третьих, отправитель должен иметь возможность восстановиться после механического повреждения, полученного во время нападения хищников.
Исследование [21] обеспечивает поддержку первого условия, согласно которому выпуск шрекстофа должен привлекать хищников. Этот эксперимент показал, что шрекстофф, извлеченный из кожи толстоголовых гольянов, привлекал как северную щуку ( Esox lucius ), так и хищных жуков-ныряльщиков ( Colymbetescultilis ). Кроме того, естественное исследование показало, что хищная рыба в семь раз чаще попадает в приманку, наживленную губкой, пропитанной экстрактом кожи толстоголового гольяна, чем губкой, смоченной либо в воде, либо в экстракте кожи неостариофизановой каторжной цихлиды (которая, по-видимому, не производит шрекстоффа). ). [22]
В то время как в двух предыдущих исследованиях были приведены примеры систем, в которых Шрекстофф привлекает дополнительных хищников, была обнаружена система, для которой это было не так. [23] Пятнистый окунь ( Micropterus punctulatus ) подвергался воздействию экстрактов кожи (содержащих шрекстофф) и мышц (контроль, не содержащих шрекстоффа) пяти различных одновременно встречающихся видов добычи. Пятнистого окуня не привлекала никакая обработка Шрекстоффом. Этот результат указывает на то, что шрекстофф не всегда привлекает соответствующих хищников в этом районе. Северная щука является интродуцированным видом во многих регионах, поэтому вряд ли она могла эволюционировать совместно с толстоголовыми гольянами в ходе эволюции системы шрекстоффа. Эта система может быть более значимой с экологической точки зрения, и мало доказательств того, что шрекстофф эволюционировал как аттрактант хищников. В заключение, продолжаются дебаты о том, выполнено ли первое условие этой гипотезы.
Второе условие, которое необходимо выполнить для поддержки гипотезы о привлечении хищников, заключается в том, что дополнительные хищники должны время от времени нарушать события хищничества, увеличивая вероятность того, что добыча ускользнет. В системе северная щука/толстоголовый гольян дополнительная северная щука может помешать хищничеству одним из двух способов. [24] Во-первых, дополнительная северная щука того же размера мешает хищничеству, вступая в контакт с основным хищником (кусая его и т. д.). Во-вторых, дополнительная щука большего размера, привлеченная шрекстоффом, может охотиться на первоначального хищника.
Вероятность того, что толстоголовые гольяны сбегут после поимки северной щуки, значительно увеличивается, когда вторая щука вмешивается в процесс нападения хищников. [25] Популяция северной щуки имеет возрастную структуру с уклоном в сторону более молодых и мелких особей. Если молодая щука нападает на толстоголового гольяна и привлекает более старшего и более крупного сородича, то молодая щука может подвергаться риску каннибализма и будет склонна отпустить добычу, чтобы сосредоточиться на побеге. Что касается второго условия, дополнительные хищники, похоже, действительно мешают событиям хищничества, увеличивая вероятность того, что отправитель сбежит. Последнее условие, согласно которому особи должны успешно восстановиться после нападения хищников, по-видимому, выполнено. Подтверждением этого состояния служат наблюдения, что у многих мелких рыб в естественных популяциях наблюдаются шрамы, предположительно оставленные неудачными попытками хищников. [26]
Хотя доказательства того, что шрекстофф привлекает хищников, неоднозначны, исследования показывают, что несколько хищников будут мешать друг другу, а добыча может восстановиться после нападения хищников, когда ей удастся спастись. Степень, в которой хищников привлекает хищничество, зависит от скорости, с которой шрекстофф распространяется по водной среде, что, в свою очередь, зависит от параметров потока воды. Эта гипотеза указывает на то, что шрекстофф развился как способ увеличения вероятности выживания во время нападения хищников, а его роль в качестве сигнала хищника для сородичей развилась впоследствии. Гипотеза о привлечении хищников, подкрепленная большим количеством эмпирических исследований, чем гипотеза родственного отбора, оставалась популярной в течение довольно долгого времени.
Шрекстофф как возможная защита от патогенов, паразитов и ультрафиолетового излучения.
[ редактировать ]Последняя гипотеза утверждает, что Шрекстофф обладает иммунной функцией и может быть первой линией защиты от патогенов, паразитов и/или УФ-В-излучения. Доказательства этой гипотезы убедительны. Недавнее комплексное исследование [27] выявили, что воздействие паразитов и патогенов, проникающих через кожу остариофизанов, стимулирует выработку сигнальных клеток. Кроме того, повышенное воздействие УФ-излучения коррелировало с увеличением производства сигнальных клеток.
Роль шрекстофа в иммунном ответе была еще больше подтверждена обнаружением того, что экстракты кожи толстоголовых гольянов ингибируют рост Saprolegniaferax (водной плесени) в культуре. Напротив, экстракты кожи меченосцев ( Xiphophorus helleri ), которые, как полагают, не производят шрекстофф, увеличили рост S.ferax по сравнению с контролем. Кадмий , тяжелый металл и иммунодепрессант у позвоночных. [28] подавляет выработку сигнальных клеток при заражении рыб сапролегнией . [29] Кроме того, последующее исследование [30] лечили толстоголовых гольянов кортизолом, известным иммунодепрессантом, который значительно снижал накопление сигнальных клеток в сочетании с активностью лейкоцитов. Результаты этих обширных исследований убедительно свидетельствуют о том, что основной функцией Шрекстоффа является обеспечение иммунитета против ряда экологических угроз, направленных на эпидермис рыб.
Если шрекстофф развился как защита от патогенов, паразитов и УФ-В-излучения, то выпуск шрекстоффа в окружающую среду впоследствии позволил как хищникам, так и жертвам использовать эту систему. Хищники в некоторых системах могут использовать шрекстофф как сигнал к легкой трапезе, либо прерывая событие хищничества, чтобы украсть добычу для себя, либо охотясь на первоначального хищника. Соседние сородичи затем используют шрекстофф как химический сигнал, предупреждая их о присутствии хищника в этом районе.
Экологические соображения
[ редактировать ]Наиболее убедительные исследования на сегодняшний день показывают, что клетки тревожного вещества служат реакцией иммунной системы, а впоследствии развиваются экологические последствия использования этого вещества в качестве химического сигнала тревоги. Это открытие порождает ряд интересных исследовательских вопросов. Во-первых, как упоминалось ранее, самцы многих видов остариофизан прекращают выработку клеток сигнального вещества в период размножения, по-видимому, для того, чтобы самки не были случайно отброшены из гнезда, когда самцы подвергаются механическим повреждениям во время строительства гнезда. В свете иммунной гипотезы клетки тревожного вещества, возможно, вместо этого производятся меньше во время сезона размножения, поскольку повышенный уровень тестостерона может снизить иммунные реакции. [31] Кроме того, это открытие указывает на то, что самцы подвергаются большему риску УФ-излучения, а также заражения паразитами и патогенами в период размножения.
Роль шрекстоффа как иммунного ответа имеет дополнительные последствия в эпоху растущих изменений окружающей среды. [32] Факторы экологического стресса, включая УФ-В-излучение, загрязнение окружающей среды и паразитов, увеличиваются в окружающей среде и, вероятно, будут продолжать увеличиваться с течением времени. Воздействие UVB-излучения увеличивается из-за уменьшения содержания стратосферного озона . [33] заболевания становятся все более важными как в местном, так и в глобальном масштабе, [34] загрязняющие вещества, включая тяжелые металлы , попадают в экосистемы . [35] Если концентрация кадмия, тяжелого металла, влияющего на способность рыб производить шрекстофф в ответ на стрессовые факторы окружающей среды, увеличится, иммунный ответ многих остариофизановых рыб будет нарушен. [36]
Ссылки
[ редактировать ]- ^ (Смит 1992).
- ^ (Стенцлер и Атема 1997; Атема и Стенцлер 1977)
- ^ (Снайдер и Снайдер, 1970)
- ^ (Хьюс и Блаустейн 1985; Хьюс 1998; Луттершмидт и др. 1994)
- ^ (Нельсон 1994).
- ^ (Визенден и Смит, 1998).
- ^ (Аргентини 1976; Пфайффер 1982).
- ^ Браун и др. (2000)
- ^ Матхуру и др. (2012)
- ^ (Чиверс и Смит, 1994).
- ^ (Чиверс и Смит, 1994).
- ^ (Смит 1973).
- ^ (Смит 1973).
- ^ (см. обзор Smith 1992).
- ^ (Гамильтон 1963, 1964a, 1964b).
- ^ Матис и Смит (1992)
- ^ Нэйш и др. (1993)
- ^ (Визенден и Смит, 1998).
- ^ Визенден и Смит (1998)
- ^ (Смит 1992).
- ^ Матис и др. (1995)
- ^ (Висенден и Тиль, 2002).
- ^ Кэшнер (2004)
- ^ (Матис и др. 1995).
- ^ Чиверс и др. (1996)
- ^ (Смит и Лемли 1986; Реймхем 1988).
- ^ Чиверс и др. (2007)
- ^ (Санчес-Дардон и др. 1999)
- ^ Чиверс и др. (2007)
- ^ Хальбгевакс и др. (2009)
- ^ (Фолстад и Картер 1992).
- ^ (Чиверс и др. 2007).
- ^ (Блаустейн и др. 1997)
- ^ Кизекер и др. 2004 г.
- ^ (Дженсон и Бро-Расмуссен, 1992).
- ^ (Чиверс и др. 2007).
Источники
[ редактировать ]- Аргентини, М. 1976. Выделение возбудителя страха из кожи гольяна Phoxinus phoxinus (L). Университет Цюриха.
- Атема Дж. и Д. Стенцлер. 1977. Тревожное вещество морской грязевой улитки Nassarius obsoletus : биологическая характеристика и возможная эволюция. Журнал химической экологии 3:173-187.
- Блаустейн, А.Р., Дж. М. Кизекер, Д. П. Чиверс и Р. Г. Энтони. 1997. Атмосферное УФ-В-излучение вызывает уродства у эмбрионов амфибий. Труды Национальной академии наук США 94:13735-13737.
- Браун, Дж.Э., Дж.С.Дж. Адриан, Э. Смит, Х. Лит и С. Бреннан. 2000. Феромоны тревоги Ostariophysan: лабораторные и полевые испытания функционального значения оксидов азота. Журнал химической экологии 26:139-154.
- Чиверс, Д. П., Дж. Э. Браун и Р. Дж. Ф. Смит. 1996. Эволюция химических сигналов тревоги: привлечение хищников приносит пользу отправителям сигналов тревоги. Американский натуралист 148:649-659.
- Чиверс, Д.П. и Р.Дж.Ф. Смит. 1994. Внутри- и межвидовое избегание участков, отмеченных экстрактом кожи ручьевой колюшки ( Culea inconstans ) в естественной среде обитания. Журнал химической экологии 20:1517-1523.
- Чиверс, Д. П., Б. Д. Визенден, К. Дж. Хиндман, Т. А. Михалак, Р. К. Куш, С. Г. Каминский, К. Л. Лак, MCO Ferrari, * Р. Дж. Поллок, К. Ф. Хальбгевакс, М. С. Поллок, С. Алемади, К. Т. Джеймс, Р. К. Савалоя, К. П. Гоатер, А. Корвин, Р.С. Мирза, Дж.М. Кизекер, Г.Е. Браун, Дж.С.Дж. Адриан, П.Х. Кроун, А.Р. Блаустейн и А. Матис. 2007. Клетки эпидермального «сигнального вещества» рыб, поддерживающие не тревожные функции: возможная защита от патогенов, паразитов и УФ-излучения. Труды Королевского общества B 274: 2611-2619.
- Фесслер, СММ и М.Дж. Кайзер. 2008. Филогенетически опосредованные реакции борьбы с хищниками у двустворчатых моллюсков. Серия достижений морской экологии 363:217-225.
- Фолстад И. и Картер А.Дж. 1992. Паразиты, яркие самцы и нарушения иммунокомпетентности. Американский натуралист 139: 603-622.
- Фриш, К.в. 1938. О психологии стаи рыб. Естественные науки 26:601-606.
- Хальбгевакс, К.Ф., Р.К. Куш, Т.А. Марчант и Д.П. Чиверс. 2009. Эпидермальные клубные клетки и врожденная иммунная система гольянов. Труды Королевского общества B 98: 891-897.
- Гамильтон, WG 1963. Эволюция альтруистического поведения. Американский натуралист 97:354-356.
- Гамильтон, WG 1964a. Генетическая эволюция социального поведения. I. Журнал теоретической биологии 7:1-16.
- Гамильтон, WG 1964b. Генетическая эволюция социального поведения. II. Журнал теоретической биологии 7.
- Хьюс, Д.К. 1988. Реакция тревоги у личинок западной жабы Bufo boreas: выброс личинок химических веществ естественным хищником и его влияние на эффективность отлова хищников. Поведение животных 36:125-133.
- Хьюс, Д.К. и А.Р. Блаустейн. 1985. Исследование реакции тревоги у головастиков Bufo boreas и Rana cascadae . Поведенческая и нейронная биология 43:47-57.
- Дженсен А. и Ф. Бро-Расмуссен. 1992. Экологический кадмий в Европе. Обзоры загрязнения окружающей среды и токсикологии 125:101-108.
- Кизекер, Дж. М., Л. К. Белден, К. Ши и М. Дж. Руббо. 2004. Сокращение численности земноводных и возникающие болезни. Американский учёный 92:138-147.
- Луттершмидт, Висконсин, Г.А. Марвин и В.Х. Хатчисон. 1994. Реакция тревоги многодонтидной саламандры ( Desmognathus ochraphaeus ): конспецифичный и гетероспецифичный «шрекстофф». Журнал химической экологии 20: 2751-2757.
- Матис А. и Р.Дж.Ф. Смит. 1993. Химические сигналы тревоги увеличивают время выживания толстоголовых гольянов ( Pimephales promelas ) во время встреч с северной щукой ( Esox lucius ). Поведенческая экология 4:260-265.
- Матис А. и Р.Дж.Ф. Смит. 1994. Внутривидовые и межотрядные реакции ручьевой колюшки на химические сигналы тревоги. Экология 74:2395-2404.
- Матхуру А., Кибат К., Чеонг В.Ф., Шуй Г., Венк М.Р., Фридрих Р.В. и Джесутасан С. 2012. Фрагменты хондроитина представляют собой запахи, вызывающие у рыб страх . Текущая биология 22: 538-544.
- Нэйш, К.-А., Г. Р. Карвалью и Т. Дж. Питчер. 1993. Генетическая структура и микрораспределение косяков Phoxinus phoxinus , европейского гольяна. Журнал биологии рыб 43:75-89.
- Нельсон, Дж. С. 1994. Рыбы мира, 4-е издание. Wiley-Interscience, Нью-Йорк, штат Нью-Йорк.
- Пфайффер, В. 1982. Химические сигналы в общении. Страницы 307–325 в издательстве TJ In Hara. Хеморецепция у рыб. Эльзевир, Амстердам.
- Санчес-Дардон Дж., И. Вочча, А. Хонтела, С. Чильмончик, М. Дунье, Х. Бурманс, Б. Блейкли и М. Фурнье. 1999. Иммуномодуляция тяжелыми металлами, проверенная индивидуально или в смесях на радужной форели ( Oncorhynchus mykiss ), подвергшейся воздействию in vivo. Экологическая токсикология и химия 18: 1492-1497.
- Смит, RJF 1973. Тестостерон устраняет тревожные вещества у самцов толстоголовых гольянов. Канадский журнал зоологии 54:875-876.
- Смит, RJF 1992. Сигналы тревоги у рыб. Обзоры по биологии рыб и рыболовству 2:33-63.
- Смит, RJF и А.Д. Лемли. 1986. Выживание толстоголовых гольянов после травмирования хищниками и его возможная роль в эволюции сигналов тревоги. Экологическая биология рыб 15:147-149.
- Снайдер, НФР и Х.А. Снайдер. 1970. Тревожная реакция Diadema antillarum . Наука 168:276-278.
- Стенцлер Д. и Дж. Атема. 1977. Реакция тревоги морской грязевой улитки Nassarius obsoletus : Специфичность и приоритет поведения. Журнал химической экологии 3:159-171.
- Визенден, Б.Д. и Р.Дж.Ф. Смит. 1998. Переоценка влияния знакомства с мелководными товарищами на пролиферацию клеток тревожного вещества у остариофизановых рыб. Журнал биологии рыб 53:841-846.
- Визенден, Б.Д. и Т.А. Тиль. 2002. Полевая проверка привлечения хищников к сигнальному веществу гольяна. Журнал химической экологии 28:433-438.
