Физиология рыб

Физиология рыб — это научное исследование того, как компоненты рыбы функционируют вместе в живой рыбе. [2] Ее можно противопоставить анатомии рыб , которая изучает форму или морфологию рыб. На практике анатомия и физиология рыб дополняют друг друга: первая касается строения рыбы, ее органов или составных частей и того, как они соединяются друг с другом, что можно наблюдать на анатомическом столе или под микроскопом, а вторая - изучение того, как эти компоненты функционируют вместе в живой рыбе. Для этого сначала нам необходимо узнать об их морфологии кишечника.
Дыхание [ править ]


Этот раздел нуждается в дополнительных цитатах для проверки . ( февраль 2024 г. ) |
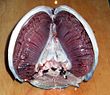
Большинство рыб обменивают газы с помощью жабр по обе стороны глотки ( горла). Жабры представляют собой ткани, состоящие из нитевидных структур, называемых нитями . Эти нити выполняют множество функций и «участвуют в переносе ионов и воды, а также в обмене кислорода, углекислого газа, кислоты и аммиака. [3] [4] Каждая нить содержит капиллярную сеть, обеспечивающую большую площадь поверхности для обмена кислорода и углекислого газа . Рыбы обменивают газы, втягивая через рот богатую кислородом воду и перекачивая ее через жабры. У некоторых рыб капиллярная кровь течет в направлении, противоположном воде, вызывая противоток . Жабры выталкивают бедную кислородом воду через отверстия по бокам глотки.
Рыбы из нескольких групп могут жить вне воды в течение длительного времени. Рыбы-земноводные , такие как илистый прыгун, могут жить и передвигаться по суше до нескольких дней или жить в стоячей или иным образом обедненной кислородом воде. Многие такие рыбы могут дышать воздухом с помощью различных механизмов. Кожа ангиллидных угрей может напрямую поглощать кислород. Ротовая полость электрического угря может дышать воздухом. Сомы семейств Loricariidae , Callichthyidae и Scoloplacidae поглощают воздух через пищеварительный тракт. [5] Двоякодышащие рыбы , за исключением австралийской двоякодышащей рыбы, и бичиры имеют парные легкие, похожие на легкие четвероногих , и должны всплывать на поверхность, чтобы заглатывать свежий воздух через рот и выводить отработанный воздух через жабры. Гар и боуфин имеют васкуляризированный плавательный пузырь, который функционирует одинаково. Вьюны , трахиры и многие сомы дышат, пропуская воздух через кишечник. Илистые прыгуны дышат, поглощая кислород через кожу (подобно лягушкам). У ряда рыб развились так называемые дополнительные органы дыхания , извлекающие кислород из воздуха. Лабиринтные рыбы (например, гурами и петушки ) имеют лабиринтный орган над жабрами , выполняющий эту функцию. Некоторые другие рыбы имеют структуры, напоминающие по форме и функциям лабиринтные органы, в первую очередь змееголовы , щукоголовые и семейство сомов Clariidae .
Воздух для дыхания в первую очередь полезен рыбам, обитающим в мелких водах с сезонными изменениями, где концентрация кислорода в воде может сезонно снижаться. Рыбы, зависящие исключительно от растворенного кислорода, такие как окунь и цихлиды , быстро задыхаются, в то время как дышащие воздухом выживают гораздо дольше, в некоторых случаях в воде, которая представляет собой немногим больше, чем влажный ил. В крайнем случае, некоторые рыбы, дышащие воздухом, способны неделями выживать во влажных норах без воды, переходя в состояние спячки (летней спячки), пока вода не вернется.
Рыб, дышащих воздухом, можно разделить на облигатно дышащих воздухом и факультативно дышащих воздухом. Облигатно дышащие воздухом, такие как африканские двоякодышащие рыбы , вынуждены периодически вдыхать воздух, иначе они задыхаются. Факультативно дышащие воздухом, такие как сом Hypostomus plecostomus , дышат воздухом только в случае необходимости, а в противном случае они могут полагаться на свои жабры для получения кислорода. Большинство рыб, дышащих воздухом, являются факультативными дышащими воздухом, которые избегают энергетических затрат на подъем на поверхность и затрат на приспособленность, связанных с воздействием поверхностных хищников. [5]

системы обмена потоками
Все базальные позвоночные дышат жабрами . Жабры расположены сразу за головой, гранича с задними краями ряда отверстий, идущих от пищевода наружу. Каждая жабра поддерживается хрящевой или костной жаберной дугой . [9] Жабры позвоночных обычно развиваются в стенках глотки вдоль ряда жаберных щелей, открывающихся наружу. Большинство видов используют противоточную систему обмена для усиления диффузии веществ в жабры и из них, при этом кровь и вода текут в противоположных направлениях друг к другу, что повышает эффективность поглощения кислорода из воды. [6] [7] [8] Свежая насыщенная кислородом вода, поступившая через рот, непрерывно «прокачивается» через жабры в одном направлении, тогда как кровь в пластинках течет в противоположном направлении, создавая противоток крови и воды, от которого зависит выживание рыбы. [8]
Жабры состоят из гребнеобразных нитей, жаберных пластинок , которые помогают увеличить площадь поверхности для обмена кислорода. [10] Когда рыба дышит, она через определенные промежутки времени втягивает в рот воду. Затем он сводит стороны горла вместе, выталкивая воду через жаберные отверстия так, что она проходит через жабры наружу. У костных рыб три пары дуг, у хрящевых — от пяти до семи пар, а у примитивных бесчелюстных — семь. У предка позвоночных, несомненно, было больше дуг, поскольку у некоторых из их хордовых родственников было более 50 пар жабр. [11]
У высших позвоночных жабры не развиваются, жаберные дуги формируются во время внутриутробного развития и закладывают основу таких важных структур, как челюсти , щитовидная железа , гортань , колумелла (соответствует стремени у млекопитающих ), а у млекопитающих молоточек и наковальня. . [11] Жаберные щели рыб могут быть эволюционными предками миндалин , вилочковой железы и евстахиевых труб , а также многих других структур, произошедших из эмбриональных жаберных мешков . [ нужна ссылка ]
Ученые исследовали, какая часть тела отвечает за поддержание дыхательного ритма. Они обнаружили, что нейроны , расположенные в стволе мозга рыб, отвечают за генезис дыхательного ритма. [12] Положение этих нейронов несколько отличается от центров респираторного генеза млекопитающих, но они расположены в одном и том же отделе мозга, что вызвало споры о гомологии дыхательных центров у водных и наземных видов. И при дыхании водных, и наземных животных точные механизмы, с помощью которых нейроны могут генерировать этот непроизвольный ритм, до сих пор полностью не изучены (см. Непроизвольный контроль дыхания ).
Еще одной важной особенностью дыхательного ритма является то, что он модулируется для адаптации к потреблению кислорода организмом. Как наблюдалось у млекопитающих, рыбы «дышат» быстрее и тяжелее, когда выполняют физические упражнения . Механизмы, с помощью которых происходят эти изменения, активно обсуждаются учеными на протяжении более 100 лет. [13] Авторов можно разделить на 2 школы:
- Те, кто считает, что большая часть дыхательных изменений заранее запрограммирована в мозге, а это означает, что нейроны центров локомоции мозга соединяются с дыхательными центрами в ожидании движений.
- Те, кто считает, что большая часть респираторных изменений является результатом обнаружения мышечных сокращений и что дыхание адаптируется вследствие мышечного сокращения и потребления кислорода. Это означало бы, что мозг обладает некими механизмами обнаружения, которые запускают дыхательную реакцию при мышечном сокращении.
Многие теперь согласны с тем, что оба механизма, вероятно, присутствуют и дополняют друг друга или работают вместе с механизмом, который может обнаруживать изменения насыщения крови кислородом и/или углекислым газом.
Костистая рыба [ править ]

У костистых рыб жабры лежат в жаберной камере, покрытой костной крышечкой. Подавляющее большинство видов костистых рыб имеют пять пар жабр, хотя некоторые из них утратили их в ходе эволюции. Жаберная крышка может играть важную роль в регулировании давления воды внутри глотки, чтобы обеспечить надлежащую вентиляцию жабр, чтобы костлявым рыбам не приходилось полагаться на таранную вентиляцию (и, следовательно, на почти постоянное движение) для дыхания. Клапаны внутри рта предотвращают выход воды. [11]
Жаберные дуги костистых рыб обычно не имеют перегородки, поэтому из дуги выступают только жабры, поддерживаемые отдельными жаберными лучами. У некоторых видов сохраняются жаберные тычинки. Хотя у всех костных рыб, кроме самых примитивных, дыхалец отсутствует, связанная с ним псевдоветвь часто сохраняется, располагаясь у основания жаберной крышки. Однако часто он сильно редуцирован и состоит из небольшой массы клеток без какой-либо сохранившейся жаберной структуры. [11]

Морские костистые кости также используют жабры для выделения электролитов . Большая площадь поверхности жабр создает проблемы для рыб, которые стремятся регулировать осмолярность своих внутренних жидкостей. Морская вода менее разбавлена, чем эти внутренние жидкости, поэтому морские рыбы теряют большое количество воды осмотически через жабры. Чтобы восстановить воду, они пьют большое количество морской воды и выделяют соль . Однако пресная вода более разбавлена, чем внутренние жидкости рыб, поэтому пресноводные рыбы получают воду осмотически через жабры. [11]
У некоторых примитивных костных рыб и земноводных личинки . несут наружные жабры, ответвляющиеся от жаберных дуг [14] С возрастом они уменьшаются, их функцию берут на себя собственно жабры у рыб и легкие у большинства земноводных. У некоторых амфибий внешние личиночные жабры сохраняются и во взрослом возрасте, а сложная внутренняя жаберная система, наблюдаемая у рыб, по-видимому, безвозвратно утрачена очень рано в эволюции четвероногих . [15]
Хрящевая рыба [ править ]
Как и другие рыбы, акулы извлекают кислород из морской воды, когда она проходит через их жабры . В отличие от других рыб, жаберные щели акулы не прикрыты, а лежат в ряд за головой. расположена модифицированная щель, называемая дыхальцем Сразу за глазом , которая помогает акуле поглощать воду во время дыхания и играет важную роль у донных акул. акул дыхалец редуцированы или отсутствуют У активных пелагических . [16] Пока акула движется, вода проходит через рот и жабры в процессе, известном как «таранная вентиляция». В состоянии покоя большинство акул перекачивают воду через жабры, чтобы обеспечить постоянный приток насыщенной кислородом воды. Небольшое количество видов утратили способность перекачивать воду через жабры и вынуждены плавать без отдыха. Эти виды являются облигатными таранами-вентиляторами и, предположительно, задохнутся, если не смогут двигаться. Облигатная таранная вентиляция характерна и для некоторых пелагических видов костистых рыб. [17]
Процесс дыхания и кровообращения начинается, когда дезоксигенированная кровь попадает в двухкамерное сердце акулы . Здесь акула перекачивает кровь к жабрам через вентральную аорты артерию , где она разветвляется на афферентные плечевые артерии. Реоксигенация происходит в жабрах, и реоксигенированная кровь течет в выносящие плечевые артерии, которые объединяются, образуя дорсальную аорту . Кровь течет из спинной аорты по всему телу. Дезоксигенированная кровь из организма затем течет через задние кардинальные вены и попадает в задние кардинальные синусы . Оттуда кровь поступает в желудочек сердца и цикл повторяется. [18]
Акулы и скаты обычно имеют пять пар жаберных щелей, которые открываются прямо наружу тела, хотя у некоторых более примитивных акул их шесть или семь пар. Соседние щели разделены хрящевой жаберной дугой, из которой выступает длинная листовидная перегородка , частично поддерживаемая еще одним фрагментом хряща, называемым жаберным лучом . Отдельные пластинки жабр лежат по обе стороны от перегородки. Основание арки может также поддерживать жаберные гребни — небольшие выступающие элементы, которые помогают фильтровать пищу из воды. [11]
Меньшее отверстие, дыхальце , находится в задней части первой жаберной щели . У него есть небольшая ложная ветвь , которая по строению напоминает жабры, но получает только кровь, уже насыщенную кислородом настоящие жабры. [11] Считается, что дыхальце гомологично ушному отверстию у высших позвоночных. [19]
Большинство акул полагаются на таранную вентиляцию, нагнетая воду в рот и через жабры, быстро плывя вперед. У тихоходных или донных видов, особенно у скатов и скатов, дыхальце может быть увеличено, и рыба дышит, всасывая воду через это отверстие, а не через рот. [11]
Химеры отличаются от других хрящевых рыб тем, что лишились и дыхальца, и пятой жаберной щели. Остальные щели прикрыты жаберной крышкой , развившейся из перегородки жаберной дуги впереди первой жабры. [11]
Миноги и миксины [ править ]
У миног и миксин жаберных щелей как таковых нет. Вместо этого жабры заключены в сферические мешочки с круглым отверстием наружу. Как и жаберные щели высших рыб, каждый мешочек содержит по две жабры. В некоторых случаях отверстия могут сливаться вместе, образуя жаберную крышку. У миног семь пар сумок, а у миксин их может быть от шести до четырнадцати, в зависимости от вида. У миксины мешочки внутренне соединяются с глоткой. У взрослых миног под собственно глоткой развивается отдельная дыхательная трубка, отделяющая пищу и воду от дыхания путем закрытия клапана на ее переднем конце. [11]
Тираж [ править ]

Кровеносные системы всех позвоночных животных закрыты , так же , как и у человека. Тем не менее, системы рыб , земноводных , рептилий и птиц демонстрируют различные стадии эволюции системы кровообращения . У рыб система имеет только один контур: кровь перекачивается через капилляры жабр в капилляры тканей тела. Это известно как одноцикловая циркуляция. Таким образом, сердце рыбы представляет собой всего лишь один насос (состоящий из двух камер). У рыб замкнутая система кровообращения . Сердце перекачивает кровь по одному контуру по всему телу. У большинства рыб сердце состоит из четырех частей, включая две камеры, вход и выход. [20] Первая часть — это венозный синус , тонкостенный мешок, который собирает кровь из вен рыбы , прежде чем позволить ей течь во вторую часть, предсердие , которое представляет собой большую мышечную камеру. Предсердие служит односторонней предкамерой, направляет кровь в третью часть, желудочек . Желудочек — это еще одна толстостенная мышечная камера, которая перекачивает кровь сначала в четвертую часть, артериальную луковицу , большую трубку, а затем из сердца. Артериальная луковица соединяется с аортой , через которую кровь поступает к жабрам для насыщения кислородом.
У земноводных и большинства рептилий используется двойная система кровообращения , но сердце не всегда полностью разделено на два насоса. амфибий трехкамерное Сердце .
Пищеварение [ править ]
Челюсти позволяют рыбам поедать самую разнообразную пищу, включая растения и другие организмы. Рыбы заглатывают пищу через рот и расщепляют ее в пищеводе . В желудке пища переваривается, а у многих рыб перерабатывается в мешочках в форме пальцев, называемых пилорическими придатками , которые выделяют пищеварительные ферменты и поглощают питательные вещества. Такие органы, как печень и поджелудочная железа, добавляют ферменты и различные химические вещества по мере продвижения пищи по пищеварительному тракту. Кишечник завершает процесс пищеварения и всасывания питательных веществ.
У большинства позвоночных пищеварение представляет собой четырехэтапный процесс, в котором участвуют основные структуры пищеварительного тракта , начиная с приема пищи, помещения пищи в рот и заканчивая выведением непереваренного материала через задний проход. Изо рта пища попадает в желудок, где в виде комка она подвергается химическому расщеплению. Затем он перемещается в кишечник, где процесс расщепления пищи на простые молекулы продолжается, и результаты всасываются в виде питательных веществ в кровеносную и лимфатическую систему.
Хотя точная форма и размер желудка широко варьируются у разных позвоночных, относительное положение отверстий пищевода и двенадцатиперстной кишки остается относительно постоянным. В результате орган всегда несколько изгибается влево, а затем поворачивается назад, чтобы встретиться с пилорическим сфинктером. Однако у миног , миксин , химер , двоякодышащих рыб и некоторых костистых рыб желудка вообще нет, а пищевод открывается прямо в кишечник. Все эти животные потребляют диету, которая либо требует небольшого хранения пищи, либо не требует предварительного переваривания желудочными соками, либо и то, и другое. [11]
Тонкая кишка — это часть пищеварительного тракта, следующая за желудком, за которой следует толстая кишка , и именно здесь большая часть переваривания происходит и всасывания пищи. У рыб отделы тонкой кишки нечеткие, и кишки можно использовать термины «передняя» или «проксимальная вместо двенадцатиперстной кишка» . [21] Тонкий кишечник имеется у всех костистых рыб , хотя его форма и длина сильно различаются у разных видов. У костистых рыб он относительно короткий, обычно примерно в полтора раза длиннее тела рыбы. Обычно он имеет несколько пилорических придатков , небольших мешообразных структур по всей длине, которые помогают увеличить общую площадь поверхности органа для переваривания пищи. Илеоцекальный клапан у костистых рыб отсутствует, граница между тонкой кишкой и прямой кишкой обозначена только концом пищеварительного эпителия. [11]
У некостистых рыб, таких как акулы , осетровые и двоякодышащие рыбы , тонкого кишечника как такового нет . Вместо этого пищеварительная часть кишечника образует спиральную кишку , соединяющую желудок с прямой кишкой. При этом типе кишечника сама кишка относительно прямая, но имеет длинную складку, идущую по внутренней поверхности по спирали, иногда на десятки витков. Этот клапан значительно увеличивает как площадь поверхности, так и эффективную длину кишечника. Выстилка спиральной кишки аналогична слизистой оболочке тонкой кишки костистых рыб и четвероногих немлекопитающих. [11] У миног спиральный клапан чрезвычайно мал, возможно, потому, что их рацион требует незначительного переваривания. У миксины вообще нет спирального клапана, пищеварение происходит почти на всей длине кишечника, не подразделяющегося на разные отделы. [11]
Толстая кишка — последняя часть пищеварительной системы , обычно встречающаяся у позвоночных животных. Его функция — поглощать воду из остатков неперевариваемой пищи, а затем выводить ненужные отходы из организма. [22] У рыб нет настоящего толстого кишечника, а есть просто короткая прямая кишка, соединяющая конец пищеварительной части кишки с клоакой. У акул это включает ректальную железу , которая выделяет соль, помогая животному поддерживать осмотический баланс с морской водой. По строению железа несколько напоминает слепую кишку, но не является гомологичной структурой. [11]
Как и многие водные животные, большинство рыб выделяют азотистые отходы в виде аммиака . Часть отходов диффундирует через жабры. Отходы фильтруются почками . крови
Морские рыбы имеют тенденцию терять воду из-за осмоса . Их почки возвращают воду в организм. происходит обратное У пресноводных рыб : они имеют тенденцию получать воду осмотически. Их почки производят разбавленную мочу для выведения. У некоторых рыб есть специально адаптированные почки, которые различаются по функциям, что позволяет им перемещаться из пресной воды в соленую.
У акул пищеварение может занять длительное время. Пища перемещается изо рта в J-образный желудок, где она сохраняется и происходит первоначальное переваривание. [23] Нежелательные предметы могут никогда не пройти через желудок, и вместо этого акула либо рвет, либо выворачивает желудок наизнанку и выбрасывает ненужные предметы изо рта. Одно из самых больших различий между пищеварительными системами акул и млекопитающих заключается в том, что у акул гораздо более короткий кишечник. Такая короткая длина достигается за счет спирального клапана с множеством витков на одном коротком участке вместо длинной трубчатой кишки. Клапан обеспечивает большую площадь поверхности, требуя, чтобы пища циркулировала внутри короткой кишки до полного переваривания, когда оставшиеся продукты жизнедеятельности попадают в клоаку . [23]
Эндокринная система [ править ]
Регуляция социального поведения [ править ]
Окситоцин — это группа нейропептидов , обнаруженных у большинства позвоночных. Одна из форм окситоцина действует как гормон , связанный с человеческая любовь. В 2012 году исследователи вводили цихлидам социального вида Neolamprologus pulcher либо эту форму изотоцина, либо контрольный физиологический раствор. Они обнаружили, что изотоцин повышает «чувствительность к социальной информации», что позволяет предположить, что «это ключевой регулятор социального поведения, который развивался и существовал с древних времен». [24] [25]
загрязнения Последствия
Рыба может биоаккумулировать загрязняющие вещества, которые сбрасываются в водные пути. Эстрогенные соединения, обнаруженные в пестицидах, противозачаточных средствах, пластике, растениях, грибах, бактериях и синтетических лекарствах, попадающих в реки, влияют на эндокринную систему местных видов. [26] В Боулдере, штат Колорадо, белая присоска , обнаруженная ниже по течению от городских очистных сооружений, демонстрирует нарушенное или аномальное половое развитие. Рыба подверглась воздействию более высокого уровня эстрогена, что привело к феминизации рыбы. [27] У самцов имеются женские репродуктивные органы, у обоих полов снижена плодовитость и выше вылупившаяся смертность. [28]
Пресноводные места обитания в Соединенных Штатах широко загрязнены распространенным пестицидом атразином . [29] Существуют разногласия по поводу степени вреда этого пестицида эндокринной системе пресноводных рыб и земноводных . Исследователи, не финансируемые промышленностью, постоянно сообщают о вредных последствиях, в то время как исследователи, финансируемые промышленностью, постоянно сообщают об отсутствии вредных последствий. [29] [30] [31]
В морской экосистеме хлорорганические загрязнители, такие как пестициды, гербициды (ДДТ) и хлордан , накапливаются в тканях рыб и нарушают работу их эндокринной системы. [32] Высокая частота бесплодия и высокий уровень хлорорганических соединений были обнаружены у капотоголовых акул вдоль побережья Мексиканского залива во Флориде. Эти соединения , нарушающие работу эндокринной системы , по структуре схожи с естественными гормонами рыб. Они могут модулировать гормональные взаимодействия у рыб путем: [33]
- связывание с клеточными рецепторами, вызывающее непредсказуемую и аномальную активность клеток
- блокирование рецепторных участков, ингибирование активности
- способствуя созданию дополнительных рецепторных участков, усиливая действие гормона или соединения
- взаимодействуя с естественными гормонами, изменяя их форму и воздействие
- влияя на синтез или метаболизм гормонов, вызывая неправильный баланс или количество гормонов
Осморегуляция [ править ]


Двумя основными типами осморегуляции являются осмоконформеры и осморегуляторы. Осмоконформеры активно или пассивно согласовывают осмолярность своего тела с окружающей средой. Большинство морских беспозвоночных являются осмоконформерами, хотя их ионный состав может отличаться от состава морской воды.
Осморегуляторы тела жестко регулируют осмолярность , которая всегда остается постоянной и чаще встречается в животном мире. Осморегуляторы активно контролируют концентрацию солей, несмотря на концентрацию солей в окружающей среде. Пример – пресноводная рыба. Жабры активно поглощают соль из окружающей среды с помощью клеток, богатых митохондриями. Вода диффундирует в рыбу, поэтому она выделяет очень гипотоническую (разбавленную) мочу, чтобы удалить всю лишнюю воду. У морской рыбы внутренняя осмотическая концентрация ниже, чем у окружающей морской воды, поэтому она имеет тенденцию терять воду и накапливать соль. Он активно выделяет соли из жабр . Большинство рыб являются стеногалинными , что означает, что они обитают либо в соленой, либо в пресной воде и не могут выжить в воде с концентрацией соли, отличной от той, к которой они приспособлены. Однако некоторые рыбы демонстрируют потрясающую способность к эффективной осморегуляции в широком диапазоне солености; рыбы с такой способностью известны как эвригалинные виды, например лосось. . Было замечено, что лосось обитает в двух совершенно разных средах — морской и пресной воде — и ему свойственно адаптироваться к обоим, внося поведенческие и физиологические изменения.
В отличие от костистых рыб, за исключением целаканта , [34] Кровь и другие ткани акул и Chondrichthyes обычно изотоничны морской среде из-за высокой концентрации мочевины и N-оксида триметиламина (ТМАО), что позволяет им находиться в осмотическом балансе с морской водой. Эта адаптация не позволяет большинству акул выжить в пресной воде, и поэтому они ограничены морской средой. Существует несколько исключений, например, акула-бык , которая разработала способ изменения функции почек для выделения большого количества мочевины. [35] Когда акула умирает, мочевина расщепляется бактериями до аммиака, в результате чего труп постепенно начинает сильно пахнуть аммиаком. [36] [37]
Акулы применили другой, эффективный механизм сохранения воды, а именно осморегуляцию. Они сохраняют мочевину в крови в относительно более высоких концентрациях. Мочевина повреждает живые ткани, поэтому, чтобы справиться с этой проблемой, некоторые рыбы сохраняют оксид триметиламина . Это обеспечивает лучшее решение проблемы токсичности мочевины. Акулы, имеющие немного более высокую концентрацию растворенных веществ (т.е. выше 1000 мОсм, что соответствует концентрации растворенных веществ в море), не пьют воду, как пресноводные рыбы.
Терморегуляция [ править ]
Гомеотермия и пойкилотермия определяют, насколько стабильна температура организма. Большинство эндотермных , гомойотермны организмов, как и млекопитающие . Однако животные с факультативной эндотермией часто являются пойкилотермными, то есть их температура может значительно варьироваться. Точно так же большинство рыб являются эктотермными животными , поскольку все их тепло поступает из окружающей воды. Однако большинство из них являются гомойотермами, поскольку их температура очень стабильна.
Большинство организмов имеют предпочтительный температурный диапазон, однако некоторые могут адаптироваться к более низким или более высоким температурам, чем те, к которым они обычно привыкли. Предпочтительной температурой организма обычно является температура, при которой физиологические процессы организма могут протекать с оптимальной скоростью. Когда рыбы адаптируются к другим температурам, эффективность их физиологических процессов может снизиться, но они продолжат функционировать. Это называется термонейтральной зоной , в которой организм может существовать неопределенно долго. [38]
Х. М. Вернон провел работу по определению температуры смерти и температуры паралича (температуры теплового оцепенения) различных животных. Он обнаружил, что виды одного и того же класса показали очень схожие значения температуры: у исследованных земноводных - 38,5 ° C, у рыб - 39 ° C, у рептилий - 45 ° C, а у различных моллюсков - 46 ° C.
Чтобы справляться с низкими температурами, некоторые рыбы развили способность сохранять работоспособность даже при температуре воды ниже нуля; некоторые используют природный антифриз или белки-антифризы, чтобы противостоять образованию кристаллов льда в тканях.
Большинство акул «хладнокровны» или, точнее, пойкилотермны , что означает, что их внутренняя температура тела соответствует температуре окружающей среды. Члены семейства Lamnidae (такие как короткоперая акула-мако и большая белая акула ) гомойотермны и поддерживают более высокую температуру тела, чем окружающая вода. У этих акул полоска аэробных красных мышц, расположенная недалеко от центра тела, генерирует тепло, которое тело сохраняет с помощью противоточного механизма обмена с помощью системы кровеносных сосудов , называемой rete mirabile («чудесная сеть»). Обыкновенная акула-молотилка имеет аналогичный механизм поддержания повышенной температуры тела, который, как полагают, развился независимо. [ не удалось пройти проверку ] . [39]
Тунец может поддерживать температуру определенных частей своего тела выше температуры окружающей морской воды. Например, синий тунец поддерживает внутреннюю температуру тела 25–33 ° C (77–91 ° F), а в воде холодную до 6 ° C (43 ° F). Однако, в отличие от типичных эндотермических существ, таких как млекопитающие и птицы, тунец не поддерживает температуру в относительно узком диапазоне. [40] Тунец достигает эндотермии , сохраняя тепло, выделяемое в результате нормального метаболизма . Rete mirabile («чудесная сеть»), переплетение вен и артерий на периферии тела, передает тепло от венозной крови к артериальной через противоточную систему обмена, тем самым смягчая эффекты поверхностного охлаждения. Это позволяет тунцу повышать температуру высокоаэробных тканей скелетных мышц, глаз и мозга. [40] [41] который поддерживает более высокую скорость плавания и снижает расход энергии, а также позволяет им выживать в более прохладных водах в более широком диапазоне океанских сред, чем у других рыб. [ нужна ссылка ] Однако у всех тунцов сердце работает при температуре окружающей среды , так как получает охлажденную кровь, а коронарное кровообращение происходит непосредственно от жабр . [41]
- Гомеотермия : Хотя большинство рыб исключительно экзотермны , есть исключения. Некоторые виды рыб поддерживают повышенную температуру тела. Эндотермные костистые рыбы (костистые рыбы) относятся к подотряду Scombroidei и включают марлиновых , тунцов, включая «примитивный» вид скумбрии , Gasterochisma melampus . Все акулы семейства Lamnidae — короткоплавники мако, длинноплавники мако, белая, беловая акула и лососевая акула — эндотермичны, и данные свидетельствуют о том, что этот признак существует у семейства Alopiidae ( акул-молотилок ). Степень эндотермии варьируется от марлиновых , которые согревают только глаза и мозг, до голубого тунца и морских акул , у которых температура тела превышает температуру окружающей воды более чем на 20 ° C (36 ° F). [42] См. также гигантотермия . Считается, что эндотермия, хотя и требует больших метаболических затрат, дает такие преимущества, как увеличение мышечной силы, более высокие скорости обработки данных в центральной нервной системе и более высокие скорости пищеварения .
У некоторых рыб rete mirabile позволяет повысить температуру мышц в тех областях, где находится сеть вен и артерий. Рыба способна терморегулировать определенные участки своего тела. Кроме того, это повышение температуры приводит к повышению базальной метаболической температуры. Рыба теперь способна расщеплять АТФ с большей скоростью и, в конечном итоге, может плавать быстрее.
Глаз рыбы -меч может генерировать тепло , чтобы лучше обнаруживать добычу на глубине до 600 метров (2000 футов). [43]
Мышечная система [ править ]


Рыбы плавают за счет сокращения продольных красных мышц и косо ориентированных белых мышц. Красная мышца является аэробной и нуждается в кислороде, который поставляет миоглобин . Белая мышца анаэробна и не нуждается в кислороде. Красные мышцы используются для продолжительной активности, например, при движении на медленной скорости во время океанских миграций. Белые мышцы используются для всплесков активности, таких как прыжки или резкие скачки скорости для ловли добычи. [44]
, мышцы В основном рыбы имеют белые мышцы, но у некоторых рыб, таких как скомброиды и лососевые варьируются от розового до темно-красного цвета. Красные миотомальные мышцы получают свой цвет от миоглобина , молекулы, связывающей кислород, которую тунец экспрессирует в количествах, гораздо больших, чем у большинства других рыб. Богатая кислородом кровь дополнительно обеспечивает доставку энергии к мышцам. [40]
Большинство рыб двигаются, поочередно сокращая парные группы мышц по обе стороны позвоночника. Эти сокращения образуют S-образные кривые, которые движутся вниз по телу. Когда каждая кривая достигает спинного плавника , к воде прикладывается обратная сила, которая вместе с плавниками перемещает рыбу вперед. функционируют Плавники рыбы как закрылки самолета. Плавники также увеличивают площадь поверхности хвоста, увеличивая скорость. Обтекаемое тело рыбы уменьшает трение о воду.
Типичной характеристикой многих животных, которые используют волнообразное передвижение, является то, что у них есть сегментированные мышцы или блоки миомеров , идущие от головы к хвостам, которые разделены соединительной тканью, называемой миосептами. Кроме того, некоторые сегментированные группы мышц, например латеральная гипаксиальная мускулатура у саламандры, ориентированы под углом к продольному направлению. Для этих косо ориентированных волокон деформация в продольном направлении больше, чем деформация в направлении мышечных волокон, что приводит к архитектурному передаточному числу, превышающему 1. Более высокий начальный угол ориентации и большее дорсовентральное выпячивание приводит к более быстрому сокращению мышц, но приводит к меньшее количество производимой силы. [45] Предполагается, что животные используют механизм переменной передачи, который позволяет саморегулировать силу и скорость для удовлетворения механических требований сокращения. [46] Когда перистая мышца подвергается воздействию небольшой силы, сопротивление изменениям ширины мышцы заставляет ее вращаться, что, следовательно, приводит к более высокому передаточному числу архитектурной передачи (AGR) (высокая скорость). [46] Однако под действием высокой силы перпендикулярная компонента силы волокон преодолевает сопротивление изменениям ширины, и мышца сжимается, создавая более низкую AGR (способную поддерживать более высокую выходную силу). [46]
Большинство рыб во время плавания сгибаются в виде простого однородного пучка за счет сокращений продольных красных мышечных волокон и косо ориентированных белых мышечных волокон внутри сегментированной осевой мускулатуры. волокон Напряжение (εf), испытываемое продольными красными мышечными волокнами, эквивалентно продольному растяжению (εx). Более глубокие белые мышечные волокна рыб имеют разнообразное расположение. Эти волокна организованы в конусообразные структуры и прикрепляются к листам соединительной ткани, известным как миосепты; каждое волокно имеет характерную дорсовентральную (α) и медиолатеральную (φ) траекторию. Теория сегментированной архитектуры предсказывает, что εx > εf. Это явление приводит к архитектурному передаточному числу, определяемому как продольная деформация, деленная на деформацию волокна (εx / εf), превышающую единицу, и усиление продольной скорости; кроме того, это возникающее усиление скорости может быть усилено за счет изменения архитектурной зубчатой передачи за счет изменений мезолатеральной и дорсовентральной формы - закономерность, наблюдаемая в перистых мышцах. сокращения. Передаточное отношение красного к белому (красный εf / белый εf) отражает совокупный эффект растяжения продольных красных мышечных волокон и косых белых мышечных волокон. [45] [47]
Плавучесть [ править ]


Тело рыбы плотнее воды, поэтому рыба должна компенсировать разницу, иначе она утонет. У многих костных рыб есть внутренний орган, называемый плавательным пузырем или газовым пузырем, который регулирует их плавучесть путем манипулирования газами. Таким образом, рыба может оставаться на текущей глубине воды, подниматься или опускаться, не тратя энергию на плавание. Мочевой пузырь имеется только у костистых рыб. У более примитивных групп, таких как некоторые гольяны , бичиры и двоякодышащие рыбы , мочевой пузырь открыт пищеводу и выполняет функцию легкого . Он часто отсутствует у быстроплавающих рыб, таких как семейства тунцов и скумбрии. Состояние мочевого пузыря, открытого в пищевод, называется физостомой , закрытым состоянием физоклиста . В последнем случае содержание газа в мочевом пузыре контролируется через сеть мирабилис — сеть кровеносных сосудов, осуществляющих газообмен между мочевым пузырем и кровью. [48]
У некоторых рыб сетчатка мирабиле наполняет плавательный пузырь кислородом. противоточная система обмена Между венозными и артериальными капиллярами используется . Понижая уровень pH в венозных капиллярах, кислород отделяется от гемоглобина крови . Это вызывает увеличение концентрации кислорода в венозной крови, позволяя кислороду диффундировать через капиллярную мембрану в артериальные капилляры, где кислород все еще связывается с гемоглобином. Цикл диффузии продолжается до тех пор, пока концентрация кислорода в артериальных капиллярах не станет пересыщенной (больше, чем концентрация кислорода в плавательном пузыре). В этот момент свободный кислород из артериальных капилляров диффундирует в плавательный пузырь через газовую железу. [49]
В отличие от костистых рыб, у акул нет плавательного пузыря, наполненного газом, для обеспечения плавучести. Вместо этого акулы полагаются на большую печень, наполненную маслом, содержащим сквален , и хрящи, плотность которых составляет примерно половину нормальной плотности костей. [50] Их печень составляет до 30% от общей массы тела. [35] Эффективность печени ограничена, поэтому акулы используют динамическую подъемную силу , чтобы поддерживать глубину, когда они не плавают. Песчаные тигровые акулы хранят воздух в желудке, используя его как плавательный пузырь. Большинству акул необходимо постоянно плавать, чтобы дышать, и они не могут долго спать, не утонув (если вообще не погружаясь). Однако некоторые виды, такие как акула-нянька , способны перекачивать воду через жабры, что позволяет им отдыхать на дне океана. [51]
Сенсорные системы [ править ]
Большинство рыб обладают высокоразвитыми органами чувств. Почти все дневные рыбы обладают цветовым зрением, по крайней мере, таким же хорошим, как у человека (см. зрение у рыб ). У многих рыб также есть хеморецепторы, отвечающие за необычайные ощущения вкуса и обоняния. Хотя у них есть уши, многие рыбы могут не очень хорошо слышать. У большинства рыб есть чувствительные рецепторы, образующие систему боковой линии , которая улавливает слабые течения и вибрации, а также улавливает движение близлежащих рыб и добычи. [52] Акулы могут чувствовать частоты в диапазоне от 25 до 50 Гц через свою боковую линию. [53]
Рыбы ориентируются по ориентирам и могут использовать мысленные карты, основанные на нескольких ориентирах или символах. Поведение рыб в лабиринтах показывает, что они обладают пространственной памятью и зрительной дискриминацией. [54]
Видение [ править ]
Зрение — важная сенсорная система для большинства видов рыб. Глаза рыб похожи на глаза наземных позвоночных, таких как птицы и млекопитающие, но имеют более сферическую линзу . Их сетчатка обычно имеет как палочки, так и колбочки (для скотопического и фотопического зрения ), и большинство видов обладают цветовым зрением . Некоторые рыбы могут видеть ультрафиолет , а некоторые — поляризованный свет . Среди бесчелюстных рыб , у миноги хорошо развиты глаза а у миксины лишь примитивные глазные пятна . [55] Зрение рыб показывает адаптацию к визуальной среде: например, глаза глубоководных рыб приспособлены к темноте.
Слушание [ править ]
Слух – важная сенсорная система для большинства видов рыб. Порог слуха и способность локализовать источники звука снижаются под водой, где скорость звука выше, чем на воздухе. Подводный слух зависит от костной проводимости , а локализация звука, по-видимому, зависит от различий в амплитуде, определяемых костной проводимостью. [56] Однако водные животные, такие как рыбы, имеют более специализированный слуховой аппарат, который эффективно работает под водой. [57]
Рыбы могут чувствовать звук через свои боковые линии и отолиты (уши). Некоторые рыбы, например некоторые виды карпа и сельди , слышат посредством своего плавательного пузыря, который функционирует как слуховой аппарат. [58]
Слух хорошо развит у карпа , у которого имеется веберов орган — три специализированных позвоночных отростка, передающих колебания плавательного пузыря на внутреннее ухо.
Хотя проверить слух акул сложно, они могут обладать острым слухом и, возможно, могут услышать добычу на расстоянии многих миль. [59] Небольшое отверстие на каждой стороне головы (не дыхалец ) ведет прямо во внутреннее ухо через тонкий канал. имеет Боковая линия аналогичное расположение и открыта для окружающей среды через ряд отверстий, называемых порами боковой линии . Это напоминание об общем происхождении этих двух органов, воспринимающих вибрацию и звук, которые сгруппированы в акустико-латеральную систему. У костных рыб и четвероногих наружное отверстие во внутреннее ухо утеряно.
Хеморецепция [ править ]

Акулы обладают острым обонянием , расположенным в коротком протоке (который не слит, в отличие от костистых рыб) между передним и задним носовыми отверстиями, при этом некоторые виды способны обнаружить всего лишь одну миллионную часть крови в морской воде. [60]
Акулы обладают способностью определять направление определенного запаха на основе времени обнаружения запаха в каждой ноздре. [61] Это похоже на метод, который используют млекопитающие для определения направления звука.
Их больше привлекают химические вещества, содержащиеся в кишечнике многих видов, и в результате они часто задерживаются вблизи канализационных стоков или в них. Некоторые виды, такие как акулы-няньки , имеют внешние усики , которые значительно повышают их способность чувствовать добычу.
Магнитоцепция [ править ]
Электрорецепция [ править ]

У некоторых рыб, таких как сом и акулы, есть органы, которые улавливают слабые электрические токи порядка милливольт. [62] Другие рыбы, такие как южноамериканские электрические рыбы Gymnotiformes , могут производить слабые электрические токи, которые они используют для навигации и социального общения. У акул ампулы Лоренцини электрорецепторными органами являются . Их число исчисляется сотнями и тысячами. Акулы используют ампулы Лоренцини для обнаружения электромагнитных полей , которые производят все живые существа. [63] Это помогает акулам (особенно акуле-молоту ) находить добычу. Акула обладает наибольшей электрической чувствительностью среди всех животных. Акулы находят добычу, спрятанную в песке, обнаруживая электрические поля создаваемые ими . Океанские течения , движущиеся в магнитном поле Земли, также генерируют электрические поля, которые акулы могут использовать для ориентации и, возможно, навигации. [64]
- Ампулы Лоренцини позволяют акулам чувствовать электрические разряды.
- Электрические рыбы способны создавать электрические поля с помощью модифицированных мышц своего тела.
Боль [ править ]
Эксперименты, проведенные Уильямом Таволгой, доказали, что у рыб есть боли реакции и страха. Например, в экспериментах Таволги рыба-жаба хрюкала при ударе током, а со временем стала хрюкать при одном лишь виде электрода. [65]
В 2003 году шотландские ученые из Эдинбургского университета и Института Рослина пришли к выводу, что радужная форель демонстрирует поведение, часто связанное с болью у других животных. Пчелиный яд и уксусная кислота, введенные в губы, заставляли рыб раскачиваться и тереться губами о стенки и дно аквариума, что, по мнению исследователей, было попыткой облегчить боль, подобно тому, что делали бы млекопитающие. [66] [67] [68] Нейроны сработали по схеме, напоминающей структуру нейронов человека. [68]
Профессор Джеймс Д. Роуз из Университета Вайоминга заявил, что исследование было ошибочным, поскольку оно не предоставило доказательств того, что рыбы обладают «сознательным сознанием, особенно тем видом осознания, которое по смыслу похоже на наше». [69] Роуз утверждает, что, поскольку мозг рыбы настолько отличается от мозга человека, рыбы, вероятно, не обладают таким же сознанием, как люди, поэтому реакции, подобные человеческим реакциям на боль, имеют другие причины. Годом ранее Роуз опубликовала исследование, в котором утверждается, что рыбы не могут чувствовать боль, потому что в их мозгу отсутствует неокортекс . [70] Однако специалист по поведению животных Темпл Грандин утверждает, что рыбы все равно могут обладать сознанием без неокортекса, потому что «разные виды могут использовать разные структуры и системы мозга для выполнения одних и тех же функций». [68]
Защитники защиты животных выражают обеспокоенность по поводу возможных страданий рыб, вызванных ловлей рыбы. Некоторые страны, например Германия, запретили определенные виды рыболовства, а британское RSPCA теперь официально преследует лиц, жестоко обращающихся с рыбой. [71]
Репродуктивные процессы [ править ]
Развитие оогоний у костистых рыб варьируется в зависимости от группы, а определение динамики оогенеза позволяет понять процессы созревания и оплодотворения. Изменения в ядре , ооплазме и окружающих слоях характеризуют процесс созревания ооцита. [72]
Постовуляторные фолликулы представляют собой структуры, образующиеся после выхода ооцита; они не выполняют эндокринной функции, имеют широкий неровный просвет и быстро реабсорбируются в процессе, включающем апоптоз фолликулярных клеток. Дегенеративный процесс, называемый фолликулярной атрезией, приводит к реабсорбции непророжденных вителлогенных ооцитов. Этот процесс также может происходить, но реже, в ооцитах на других стадиях развития. [72]
Некоторые рыбы являются гермафродитами , имеющими семенники и яичники либо на разных стадиях жизненного цикла, либо, как в деревнях , имеют их одновременно.
Более 97% всех известных рыб яйцекладущие . [73] то есть яйца развиваются вне тела матери. Примеры яйцекладущих рыб включают лосося , золотую рыбку , цихлид , тунца и угрей . У большинства этих видов оплодотворение происходит вне тела матери, при этом самцы и самки сбрасывают свои гаметы в окружающую воду. Тем не менее, некоторые яйцекладущие рыбы практикуют внутреннее оплодотворение, при этом самцы используют своего рода интромиттирующий орган для доставки спермы в половое отверстие самки, в первую очередь яйцекладущие акулы, такие как рогатая акула , и яйцекладущие скаты, такие как скаты . В этих случаях самец оснащен парой модифицированных брюшных плавников, известных как класперы .
Морская рыба может производить большое количество икры, которая часто выбрасывается в толщу открытой воды. Яйца имеют средний диаметр 1 миллиметр (0,039 дюйма). Яйца обычно окружены внезародышевыми оболочками, но вокруг этих оболочек не образуется твердая или мягкая оболочка. У некоторых рыб толстая кожистая шерсть, особенно если им приходится выдерживать физическую силу или высыхание. Яйца такого типа также могут быть очень маленькими и хрупкими.
 Яйцо миноги
Яйцо миноги Яйцо кошачьей акулы ( кошелек русалки )
Яйцо кошачьей акулы ( кошелек русалки ) Яйцо бычьей акулы
Яйцо бычьей акулы Яйцо химеры
Яйцо химеры

Только что вылупившуюся молодь яйцекладущих рыб называют личинками . Они обычно плохо сформированы, несут большой желточный мешок (для питания) и по внешнему виду сильно отличаются от молодых и взрослых особей. Личиночный период у яйцекладущих рыб относительно короткий (обычно всего несколько недель), личинки быстро растут и меняют внешний вид и структуру (процесс, называемый метаморфозом ), превращаясь в молодь. Во время этого перехода личинки должны переключиться с желточного мешка на питание добычей зоопланктона - процесс, который обычно зависит от недостаточной плотности зоопланктона, приводящей к голоданию многих личинок.
У яйцеживородящих рыб икра развивается внутри тела матери после внутреннего оплодотворения, но получает мало или вообще не получает питания непосредственно от матери, вместо этого оно зависит от желтка . Каждый эмбрион развивается в своей яйцеклетке. Известные примеры яйцеживородящих рыб включают гуппи , акулы-ангелы и целаканты .
Некоторые виды рыб живородящие . У таких видов мать сохраняет яйца и питает эмбрионы. Обычно живородящие рыбы имеют структуру, аналогичную плаценте млекопитающих , соединяющую кровоснабжение матери с кровоснабжением эмбриона. Примерами живородящих рыб являются окуни для серфинга , расщепленные плавники и лимонная акула . У некоторых живородящих рыб наблюдается оофагия , при которой развивающиеся эмбрионы поедают другие яйца, произведенные матерью. Это наблюдалось в первую очередь среди акул, таких как короткоплавник мако и porbeagle , но известно и для некоторых костистых рыб, таких как полуклюв Nomorhamphus ebrardtii . [74] Внутриутробный каннибализм — еще более необычный способ живорождения, при котором самые крупные эмбрионы поедают более слабых и меньших братьев и сестер. Такое поведение также чаще всего встречается среди акул, таких как серая акула-нянька , но также сообщалось и о Nomorhamphus ebrardtii . [74]
У многих видов рыб плавники были модифицированы, чтобы обеспечить внутреннее оплодотворение .
Аквариумисты обычно называют яйцеживородящих и живородящих рыб живородящими .
- Многие виды рыб являются гермафродитами . Синхронные гермафродиты обладают и яичниками , и семенниками одновременно . У сиквенсальных гермафродитов присутствуют оба типа ткани в гонадах , причем преобладает один тип, тогда как рыбы принадлежат к соответствующему полу.
Социальное поведение [ править ]
Социальное поведение рыб, называемое «обмелением», предполагает совместное плавание группы рыб. Такое поведение является защитным механизмом в том смысле, что в больших количествах существует безопасность и снижается вероятность быть съеденным хищниками. Обмеление также увеличивает успех спаривания и поиска пищи. С другой стороны, стайное поведение — это поведение внутри стаи, при котором можно увидеть, как рыба синхронно выполняет различные маневры. [75] Параллельное плавание — это форма «социального копирования», когда рыбы в стае копируют направление и скорость соседних рыб. [76]
Эксперименты, проведенные доктором Стивеном по изучению стайного поведения рыб, пришли к выводу, что в течение дня рыбы имеют более высокую склонность оставаться вместе в результате баланса между уходом отдельных рыб и поиском собственного направления и взаимным притяжением между рыбами одного и того же происхождения. разновидность. Выяснилось, что ночью рыбы плавали заметно быстрее, однако часто поодиночке и без координации. Часто можно было видеть группы из двух или трех человек, но они рассеивались через пару секунд. [77]
Теоретически, время, в течение которого рыба остается вместе в косяке, должно отражать ее стоимость пребывания, а не ухода. [78] Прошлый лабораторный эксперимент, проведенный на карповых, показал, что временной бюджет социального поведения внутри стаи варьируется пропорционально количеству присутствующих рыб. Это происходит из-за соотношения затрат и выгод, которое меняется соответственно с размером группы, измеряемым риском нападения хищников по сравнению с потреблением пищи. [79] Когда соотношение затрат и выгод благоприятствует стайному поведению, тогда решение остаться в группе или присоединиться к ней является благоприятным. В зависимости от этого соотношения рыба соответственно примет решение уйти или остаться. Таким образом, считается, что поведение стайки определяется постоянным потоком решений, принимаемых отдельной рыбой. [75]
См. также [ править ]
Ссылки [ править ]
- ^ Шваб, Иван Р. (2002). «Больше, чем просто крутые оттенки» . Британский журнал офтальмологии . 86 (10): 1075. дои : 10.1136/bjo.86.10.1075 . ПМЦ 1771297 . ПМИД 12349839 .
- ^ Проссер, К. Лэдд (1991). Сравнительная физиология животных, экологическая и метаболическая физиология животных (4-е изд.). Хобокен, Нью-Джерси: Вили-Лисс. стр. 1–12. ISBN 978-0-471-85767-9 .
- ^ Хоар WS и Рэндалл DJ (1984) Физиология рыб: жабры: Часть A - Анатомия, газообмен и регуляция кислотно-щелочного баланса Academic Press. ISBN 978-0-08-058531-4 .
- ^ Хоар В.С. и Рэндалл DJ (1984) Физиология рыб: жабры: Часть B - Перенос ионов и воды Academic Press. ISBN 978-0-08-058532-1 .
- ↑ Перейти обратно: Перейти обратно: а б Армбрустер, Джонатан В. (1998). «Модификации пищеварительного тракта для удержания воздуха у лорикариевых и сколоплацидных сомов» (PDF) . Копейя . 1998 (3): 663–675. дои : 10.2307/1447796 . JSTOR 1447796 . Проверено 25 июня 2009 г.
- ↑ Перейти обратно: Перейти обратно: а б Кэмпбелл, Нил А. (1990). Биология (Второе изд.). Редвуд-Сити, Калифорния: Benjamin/Cummings Publishing Company, Inc., стр. 836–838 . ISBN 0-8053-1800-3 .
- ↑ Перейти обратно: Перейти обратно: а б Хьюз GM (1972). «Морфометрия жабр рыб». Физиология дыхания . 14 (1–2): 1–25. дои : 10.1016/0034-5687(72)90014-x . ПМИД 5042155 .
- ↑ Перейти обратно: Перейти обратно: а б с Сторер, Трейси И.; Узергер, Р.Л.; Стеббинс, Роберт С.; Нибаккен, Джеймс В. (1997). Общая зоология (шестое изд.). Нью-Йорк: МакГроу-Хилл. стр. 668–670 . ISBN 0-07-061780-5 .
- ^ Скотт, Томас (1996). Краткая энциклопедия биологии . Вальтер де Грюйтер. п. 542 . ISBN 978-3-11-010661-9 .
- ^ Эндрюс, Крис; Адриан Экселл; Невилл Кэррингтон (2003). Руководство по здоровью рыб . Книги Светлячка.
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот Ромер, Альфред Шервуд ; Парсонс, Томас С. (1977). Тело позвоночного . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 316–327. ISBN 978-0-03-910284-5 .
- ^ Рассел, Дэвид Ф. (1986). «Генерация дыхательного паттерна у взрослых миног (Lampetra fluviatilis): интернейроны и сброс импульсов» . Журнал сравнительной физиологии А. 158 (1): 91–102. дои : 10.1007/BF00614523 . ПМИД 3723432 . S2CID 19436421 .
- ^ Уолдроп, Тони Г.; Ивамото, Гэри А. (2006). «Точка: Контрапункт: супраспинальные локомоторные центры вносят/не вносят существенного вклада в гиперпноэ при динамических упражнениях» . Журнал прикладной физиологии . 100 (3): 1077–1083. doi : 10.1152/japplphysicalol.01528.2005 .
- ^ Сарский, Хенрик (1957). «Происхождение личинки и метаморфозы у амфибий». Американский натуралист . 91 (860). Институт Эссекса: 287. doi : 10.1086/281990 . JSTOR 2458911 . S2CID 85231736 .
- ^ Клак, Дж. А. (2002): Завоевывать позиции: происхождение и эволюция четвероногих. Издательство Индианского университета , Блумингтон, Индиана. 369 стр.
- ^ Гилбертсон, Лэнс (1999). Руководство зоологической лаборатории . Нью-Йорк: McGraw-Hill Companies, Inc. ISBN 978-0-07-237716-3 .
- ^ Уильям Дж. Беннетта (1996). «Глубокое дыхание» . Архивировано из оригинала 14 августа 2007 года . Проверено 28 августа 2007 г.
- ^ «АКУЛЫ И СКВАТЫ, SeaWorld/Busch Gardens ЖИВОТНЫЕ, СИСТЕМА КРОВООБРАЩЕНИЯ» . Буш Энтертейнмент Корпорейшн. Архивировано из оригинала 24 апреля 2009 года . Проверено 3 сентября 2009 г.
- ^ Лорин М. (1998): Важность глобальной экономности и исторической предвзятости в понимании эволюции четвероногих. Часть I – систематика, эволюция среднего уха и подвешивание челюстей. Annales des Sciences Naturelles, Zoologie, Париж , 13e Série 19: стр. 1–42.
- ^ Сетаро, Джон Ф. (1999). Кровеносная система . Microsoft Энкарта 99.
- ^ Гийом, Жан; Издательство Праксис; Садасивам Кошик; Пьер Берго; Роберт Метайллер (2001). Питание и кормление рыб и ракообразных . Спрингер. п. 31. ISBN 978-1-85233-241-9 . Проверено 9 января 2009 г.
- ^ Национальный институт рака (2 февраля 2011 г.). «Словарь онкологических терминов NCI — толстая кишка» . Проверено 16 сентября 2012 г.
- ↑ Перейти обратно: Перейти обратно: а б Мартин, Р. Эйдан. «Нет мужества, нет славы» . Центр исследований акул ReefQuest . Проверено 22 августа 2009 г.
- ^ Реддон, Арканзас; О'Коннор, CM; Марш-Ролло, ЮВ; Бальшин, С (2012). «Влияние изотоцина на социальные реакции рыб, размножающихся совместно». Поведение животных . 84 (4): 753–760. дои : 10.1016/j.anbehav.2012.07.021 . S2CID 13037173 .
- ^ Рыбья версия окситоцина стимулирует их социальное поведение Science 2.0 , 10 октября 2012 г.
- ^ Пинто, Патрисия; Эстевао, Мария; Пауэр, Дебора (август 2014 г.). «Влияние эстрогенов и соединений, разрушающих эстроген, на минерализованные ткани рыб» . Морские наркотики . 12 (8): 4474–94. дои : 10.3390/md12084474 . ПМЦ 4145326 . ПМИД 25196834 .
- ^ Уотерман, Джим. «Исследование влияния сточных вод на половое развитие рыб в Боулдер-Крик» . Информационная сеть по устойчивому развитию района Боулдер .
- ^ Арукве, Августин (август 2001 г.). «Клеточные и молекулярные реакции на эндокринные модуляторы и влияние на воспроизводство рыб». Бюллетень о загрязнении морской среды . 42 (8): 643–655. дои : 10.1016/S0025-326X(01)00062-5 . ПМИД 11525282 .
- ↑ Перейти обратно: Перейти обратно: а б Ривз, К. (2015). «О лягушках и риторике: Атразиновые войны» . Техническая коммуникация Ежеквартально . 24 (4): 328–348. дои : 10.1080/10572252.2015.1079333 . S2CID 53626955 .
- ^ Сутер, Гленн; Кормье, Сьюзен (2015). «Проблема предвзятых данных и потенциальные решения для оценок здоровья и окружающей среды». Оценка человеческого и экологического риска . 21 (7): 1–17. дои : 10.1080/10807039.2014.974499 . S2CID 84723794 .
- ^ Рор, младший (2018) «Атразин и амфибии: история прибылей, противоречий и анимуса». В: Д. А. Делла Сала и М. И. Гольдштейн (ред.) Энциклопедия антропоцена , том 5, страницы 141–148. Оксфорд: Эльзевир.
- ^ Фаулер, Сара; Кавана, Рэйчел. «Акулы, скаты и химеры: статус хондрихтиевых рыб» (PDF) . Всемирный союз охраны природы . Проверено 3 мая 2017 г.
- ^ Арукве, Августин (2001). «Клеточные и молекулярные реакции на эндокринные модуляторы и влияние на воспроизводство рыб». Бюллетень о загрязнении морской среды . 42 (8): 643–655. дои : 10.1016/s0025-326x(01)00062-5 . ПМИД 11525282 .
- ^ Химия жидкостей организма латимерии, Latimeria chalumnae.
- ↑ Перейти обратно: Перейти обратно: а б Компаньо, Леонард; Дандо, Марк; Фаулер, Сара (2005). Акулы мира . Полевые гиды Коллинза. ISBN 978-0-00-713610-0 . OCLC 183136093 .
- ^ Джон А. Мьюзик (2005). «Методы управления промыслом пластиножаберных: 14. Использование акул» . ФАО: Департамент рыболовства и аквакультуры . Проверено 16 марта 2008 г.
- ^ Томас Баттен. «МАКО АКУЛА Isurus oxyrinchus» . Грант Делавэрского моря, Университет штата Делавэр. Архивировано из оригинала 11 марта 2008 года . Проверено 16 марта 2008 г.
- ^ Уилмер, Пэт; Стоун, Грэм; Джонстон, Ян (2009). Экологическая физиология животных . Уайли. стр. 198–200 . ISBN 978-1-4051-0724-2 .
- ^ Мартин, Р. Эйдан (апрель 1992 г.). «Огонь во чреве зверя» . Центр исследований акул ReefQuest . Проверено 21 августа 2009 г.
- ↑ Перейти обратно: Перейти обратно: а б с Сепульведа, Калифорния; Диксон, Калифорния; Бернал, Д.; Грэм, Дж. Б. (1 июля 2008 г.). «Повышенная температура красных миотомальных мышц у большинства базальных видов тунца, Allothunnus Fallai » (PDF) . Журнал биологии рыб . 73 (1): 241–249. дои : 10.1111/j.1095-8649.2008.01931.x . Архивировано из оригинала (PDF) 7 февраля 2013 года . Проверено 2 ноября 2012 г.
- ↑ Перейти обратно: Перейти обратно: а б Ландейра-Фернандес, AM; Морриссетт, Дж. М.; Бланк, Дж. М.; Блок, бакалавр наук (16 октября 2003 г.). «Температурная зависимость Са2+-АТФазы (SERCA2) в желудочках тунца и скумбрии». AJP: Регуляторная, интегративная и сравнительная физиология . 286 (2): 398Р–404. CiteSeerX 10.1.1.384.817 . дои : 10.1152/ajpregu.00392.2003 . ПМИД 14604842 .
- ^ Блок, бакалавр; Финнерти, младший (1993). «Эндотермия у рыб: филогенетический анализ ограничений, предрасположенностей и давления отбора» (PDF) . Экологическая биология рыб . 40 (3): 283–302. дои : 10.1007/BF00002518 . S2CID 28644501 . [ постоянная мертвая ссылка ]
- ^ Дэвид Флешлер (15 октября 2012 г.) Sun-Sentinel Южной Флориды ,
- ^ Капур 2004 г.
- ↑ Перейти обратно: Перейти обратно: а б Брейнерд, Эл.; Азизи, Э. (2005). «Угол мышечных волокон, выпуклость сегментов и архитектурное передаточное число в сегментированной мускулатуре» . Журнал экспериментальной биологии . 208 (17): 3249–3261. дои : 10.1242/jeb.01770 . ПМИД 16109887 .
- ↑ Перейти обратно: Перейти обратно: а б с Азизи, Э.; Брейнерд, Эл.; Робертс, Ти Джей (2008). «Переменная передача в перистых мышцах» . ПНАС . 105 (5): 1745–1750. Бибкод : 2008PNAS..105.1745A . дои : 10.1073/pnas.0709212105 . ПМК 2234215 . ПМИД 18230734 .
- ^ Брейнерд, Эл.; Азизи, Э. (2007). «Архитектурное передаточное число и однородность напряжения мышечных волокон в сегментированной мускулатуре». Журнал экспериментальной зоологии . 307 (А): 145–155. дои : 10.1002/jez.a.358 . ПМИД 17397068 .
- ^ Кардонг, К. (2008). Позвоночные животные: Сравнительная анатомия, функции, эволюция (5-е изд.). Бостон: МакГроу-Хилл. ISBN 978-0-07-304058-5 .
- ^ Кардонг, К. (2008). Позвоночные животные: Сравнительная анатомия, функции, эволюция (5-е изд.). Бостон: МакГроу-Хилл.
- ^ Мартин, Р. Эйдан. «Как важно быть хрящевым» . Центр исследований акул ReefQuest . Проверено 29 августа 2009 г.
- ^ «Спят ли акулы» . Flmnh.ufl.edu . Проверено 23 сентября 2010 г.
- ^ Орр, Джеймс (1999). Рыба . Microsoft Encarta 99. ISBN 978-0-8114-2346-5 .
- ^ Поппер, А.Н.; К. Платт (1993). «Внутреннее ухо и боковая линия». Физиология рыб (1-е изд.). ЦРК Пресс.
- ^ Журнал студентов наук о жизни. «Подходящая методология лабиринта для изучения обучения рыб» (PDF) . Архивировано из оригинала (PDF) 6 июля 2011 года . Проверено 28 мая 2009 г.
- ^ Н. А. Кэмпбелл и Дж. Б. Рис (2005). Биология , седьмое издание. Бенджамин Каммингс, Сан-Франциско, Калифорния.
- ^ Шупак А. Шарони З. Янир Й. Кейнан Й. Алфи Й. Халперн П. (январь 2005 г.). «Подводный слух и локализация звука с воздушным интерфейсом и без него» . Отология и невротология . 26 (1): 127–130. дои : 10.1097/00129492-200501000-00023 . ПМИД 15699733 . S2CID 26944504 .
- ^ Грэм, Майкл (1941). «Слух рыб» . Природа . 147 (3738): 779. Бибкод : 1941Natur.147..779G . дои : 10.1038/147779b0 . S2CID 4132336 .
- ^ Уильямс, К. (1941). «Слух у рыб» . Природа . 147 (3731): 543. Бибкод : 1941Natur.147..543W . дои : 10.1038/147543b0 . S2CID 4095706 .
- ^ Мартин, Р. Эйдан. «Обнаружение слуха и вибрации» . Проверено 1 июня 2008 г.
- ^ Мартин, Р. Эйдан. «Запах и вкус» . Центр исследований акул ReefQuest . Проверено 21 августа 2009 г.
- ^ Функция двусторонних различий во времени прибытия запаха в обонятельной ориентации акул. Архивировано 8 марта 2012 г. в Wayback Machine , Джейн М. Гардинер, Джелле Атема, Current Biology - 13 июля 2010 г. (том 20, выпуск 13, стр. 1187-1191). )
- ^ Альберт, Дж. С. и WGR Крэмптон. 2005. Электрорецепция и электрогенез. стр. 431–472 в «Физиологии рыб», 3-е издание. Д. Х. Эванс и Дж. Б. Клэйборн (ред.). ЦРК Пресс.
- ^ Калмин А.Дж. (1982). «Обнаружение электрических и магнитных полей у пластиножаберных рыб». Наука . 218 (4575): 916–8. Бибкод : 1982Sci...218..916K . дои : 10.1126/science.7134985 . ПМИД 7134985 .
- ^ Мейер К.Г., Холланд К.Н., Папастаматиу Ю.П. (2005). «Акулы могут обнаружить изменения в геомагнитном поле» . Журнал Королевского общества, Интерфейс . 2 (2): 129–30. дои : 10.1098/rsif.2004.0021 . ПМК 1578252 . ПМИД 16849172 .
- ^ Дунайер, Джоан, «Рыба: чувствительность за пределами хватки похитителя», Программа животных, июль/август 1991 г., стр. 12–18.
- ↑ Вантресса Браун, «Рыбы чувствуют боль, говорят британские исследователи», Agence France-Presse, 1 мая 2003 г.
- ^ Кирби, Алекс (30 апреля 2003 г.). «Рыбы чувствуют боль, говорят ученые» . Новости Би-би-си . Проверено 4 января 2010 г.
- ↑ Перейти обратно: Перейти обратно: а б с Грандин, Темпл ; Джонсон, Кэтрин (2005). Животные в переводе . Нью-Йорк, Нью-Йорк: Скрибнер. стр. 183–184 . ISBN 978-0-7432-4769-6 .
- ^ «Роуз, JD 2003. Критика статьи: «Есть ли у рыб ноцицепторы: доказательства эволюции сенсорной системы позвоночных» » (PDF) . Архивировано из оригинала (PDF) 8 июня 2011 года . Проверено 21 мая 2011 г.
- ^ Джеймс Д. Роуз, Чувствуют ли рыбы боль? Архивировано 20 января 2013 года в Wayback Machine , 2002. Проверено 27 сентября 2007 года.
- ↑ Лик, Дж. «Рыболовы столкнутся с проверкой RSPCA», The Sunday Times – Великобритания, 14 марта 2004 г.
- ↑ Перейти обратно: Перейти обратно: а б Гимарайнш-Крус, Родриго Х., Родриго Х.; Сантос, Хосе Э. дос; Сантос, Гилмар Б. (июль – сентябрь 2005 г.). «Структура гонад и гаметогенез Loricaria lentiginosa Isbrücker (Pisces, Teleostei, Siluriformes)» . Преподобный Брас. Зоол . 22 (3): 556–564. дои : 10.1590/S0101-81752005000300005 . ISSN 0101-8175 .
- ^ Питер Скотт: Живородящие рыбы , с. 13. Тетра Пресс, 1997. ISBN 1-56465-193-2
- ↑ Перейти обратно: Перейти обратно: а б Мейснер, А.; Бернс, Дж. (декабрь 1997 г.). «Живорождение полуклювов родов Dermogenys и Nomorhamphus (Teleostei: Hemiramphidae)». Журнал морфологии . 234 (3): 295–317. doi : 10.1002/(SICI)1097-4687(199712)234:3<295::AID-JMOR7>3.0.CO;2-8 . ПМИД 29852651 . S2CID 46922423 .
- ↑ Перейти обратно: Перейти обратно: а б Питчер, Ти Джей (1998). Обмеление и стайное обучение рыб . Гарланд, Нью-Йорк, США: Гринберг, Г. и Харарвей. ММ, стр. 748–760.
- ^ Питчер, Ти Джей (1983). «Эвристические определения поведения рыбы при стайке». Поведение животных . 31 (2): 611–613. дои : 10.1016/S0003-3472(83)80087-6 . S2CID 53195091 .
- ^ Стивен, DM (1959). «Исследования стайного поведения рыб: I. Реакция двух видов на изменения освещенности и на обонятельные раздражители». Журнал экспериментальной биологии . 36 (2): 261–280. дои : 10.1242/jeb.36.2.261 .
- ^ Партридж, Б.Л. (1982). «Жёсткие определения школьного поведения недостаточны». Поведение животных . 30 : 298–299. дои : 10.1016/S0003-3472(82)80270-4 . S2CID 53170935 .
- ^ Питчер, Ти Джей (1982). «Доказательства позиционных предпочтений стайной скумбрии». Поведение животных . 30 (3): 932–934. дои : 10.1016/S0003-3472(82)80170-X . S2CID 53186411 .
Дальнейшее чтение [ править ]
- Бернье Н.Дж., Ван Дер Краак Г., Фаррелл А.П. и Браунер С.Дж. (2009) Физиология рыб: Академическое издательство рыбной нейроэндокринологии. ISBN 978-0-08-087798-3 .
- Эдди Ф.Б. и Хэнди Р.Д. (2012) Экологическая и экологическая физиология рыб Издательство Оксфордского университета. ISBN 978-0-19-954095-2 .
- Эванс Д.Х., Дж.Б. Клэйборн и С. Карри (редакторы) (2013) Физиология рыб, 4-е издание, CRC Press. ISBN 978-1-4398-8030-2 .
- Гроселл М., Фаррелл А.П. и Браунер С.Дж. (2010) Физиология рыб: многофункциональный кишечник рыбы Academic Press. ISBN 978-0-08-096136-1 .
- Хара Т.Дж. и Зелински Б. (2006) Физиология рыб: Сенсорные системы, неврология, Академическое издательство. ISBN 978-0-08-046961-4 .
- Капур Б.Г. и Ханна Б. (2004) «Справочник по ихтиологии», страницы 137–140, Springer. ISBN 978-3-540-42854-1 .
- Маккензи DJ, Фаррелл А.П. и Браунер С.Дж. (2007) Физиология рыб: Академическое издательство примитивных рыб. ISBN 978-0-08-054952-1 .
- Сломан К.А., Уилсон Р.В. и Балшайн С. (2006) Поведение и физиология рыбного залива, профессиональное издание. ISBN 978-0-12-350448-7 .
- Вуд CM, Фаррелл AP и Браунер CJ (2011) Физиология рыб: гомеостаз и токсикология неэссенциальных металлов Academic Press. ISBN 978-0-12-378634-0 .
Внешние ссылки [ править ]
физиологии Типы | ||
|---|---|---|
| Животные |  | |
| Растения | ||
| Клетки | ||
| Связанные темы | ||
