Боль у рыбы

| Часть серии о |
| Права животных |
|---|
Рыбы соответствуют нескольким критериям, которые указывают на то, что животные, кроме человека, испытывают боль. Эти удовлетворенные критерии включают подходящую нервную систему и сенсорные рецепторы, опиоидные рецепторы и снижение реакции на вредные раздражители при приеме анальгетиков и местных анестетиков , физиологические изменения в отношении вредных раздражителей, проявление защитных двигательных реакций, проявление обучения избеганию и поиск компромисса между избеганием вредных раздражителей. и другие мотивационные требования.
Чувствуют ли рыбы боль так же, как люди, или иначе, является спорным вопросом. Боль – это сложное психическое состояние , обладающее особым качеством восприятия, но также связанное со страданием , которое является эмоциональным состоянием . Из-за этой сложности наличие боли у животного или другого человека, если на то пошло, не может быть однозначно определено с использованием методов наблюдения, но вывод о том, что животные испытывают боль, часто делается на основе вероятного присутствия феноменального сознания , которое выводится. из сравнительной физиологии мозга, а также физических и поведенческих реакций. [1]
Если рыба чувствует боль, это имеет этические последствия и последствия для благополучия животных, включая последствия воздействия загрязняющих веществ, а также практики коммерческого и любительского рыболовства , аквакультуры , декоративной рыбы и генетически модифицированной рыбы , а также рыбы, используемой в научных исследованиях .
Фон
[ редактировать ]Возможность того, что рыбы и другие животные, кроме человека, испытывают боль, имеет долгую историю. Первоначально это было основано на теоретических и философских аргументах, но в последнее время оно перешло к научным исследованиям.
Философия
[ редактировать ]
Идея о том, что животные, отличные от человека, могут не чувствовать боли, восходит к французскому философу 17-го века Рене Декарту , который утверждал, что животные не испытывают боли и страданий, потому что у них нет сознания . [2] [3] [4] В 1789 году британский философ и социал-реформист Джереми Бентам обратился в своей книге «Введение в принципы морали и законодательства» к вопросу нашего обращения с животными следующими часто цитируемыми словами: «Вопрос не в том, могут ли они рассуждать? и не «Могут ли они говорить?», а «Могут ли они страдать?» [5] Чарльз Дарвин сказал, что «низшие животные, как и человек, явно чувствуют удовольствие и боль, счастье и несчастье». [6]
Питер Сингер , специалист по биоэтике и автор книги « Освобождение животных» , опубликованной в 1975 году, предположил, что сознание не обязательно является ключевым вопросом: то, что у животных мозг меньше или они «менее сознательны», чем люди, не означает, что они не способны чувствовать боль. Далее он утверждает, что мы не предполагаем, что новорожденные, люди, страдающие нейродегенеративными заболеваниями головного мозга, или люди с ограниченными возможностями обучения испытывают меньшую боль, чем мы. [7]
Бернард Роллин , главный автор двух федеральных законов США, регулирующих обезболивание животных, пишет, что в 1980-е годы исследователи оставались неуверенными в том, испытывают ли животные боль, а ветеринаров, обучавшихся в США до 1989 года, учили просто игнорировать боль животных. [8] В ходе общения с учеными и другими ветеринарами Роллина регулярно просили «доказать», что животные находятся в сознании, и предоставить «научно приемлемые» основания утверждать, что они чувствуют боль. [8]
В 1990-е годы дискуссии получили дальнейшее развитие о роли философии и науки в понимании когнитивных способностей и менталитета животных . [9] В последующие годы утверждалось, что существует сильная поддержка предположения о том, что некоторые животные (скорее всего, амниоты ) обладают по крайней мере простыми сознательными мыслями и чувствами. [10] и что точка зрения, что животные чувствуют боль иначе, чем люди, теперь является точкой зрения меньшинства. [2]
Научное исследование
[ редактировать ]Отсутствие неокортекса , по-видимому, не мешает организму испытывать аффективные состояния. Сходящиеся данные указывают на то, что животные, кроме человека, обладают нейроанатомическими, нейрохимическими и нейрофизиологическими субстратами состояний сознания, а также способностью демонстрировать намеренное поведение . Следовательно, масса доказательств указывает на то, что люди не уникальны в обладании нейрологическими субстратами , генерирующими сознание. Животные, кроме человека, включая всех млекопитающих и птиц, а также многие другие существа, включая осьминогов, также обладают этими неврологическими субстратами. [11]
В 20-м и 21-м веках было проведено множество научных исследований боли у животных, кроме человека.
Доктор Линн Снеддон вместе со своими коллегами Брейтуэйтом и Джентлом первыми открыли ноцицепторы (болевые рецепторы) у рыб. Она заявила, что рыбы демонстрируют связанные с болью изменения в физиологии и поведении, которые уменьшаются обезболивающими, и они демонстрируют более высокую мозговую активность при болезненной стимуляции. [12] Профессор Виктория Брейтуэйт в своей книге « Чувствуют ли рыбы боль?» , писал, что рыбы, подобно птицам и млекопитающим, обладают способностью к самосознанию и могут чувствовать боль. [13] Дональд Брум , профессор по защите животных Кембриджского университета, Англия, сказал, что большинство болевых систем млекопитающих также встречается у рыб, которые могут чувствовать страх и испытывать эмоции, которые контролируются в мозге рыбы в областях, анатомически отличающихся, но функционально очень похожих на те, которые есть у рыб. млекопитающие. [14]
Американская ветеринарно-медицинская ассоциация признает, что рыбы чувствуют боль, утверждая, что имеющиеся данные подтверждают позицию, согласно которой к рыбам следует относиться с теми же соображениями, что и к наземным позвоночным, в отношении облегчения боли. [15] Королевское общество по предотвращению жестокого обращения с животными в Великобритании в 1980 году заказало независимую группу экспертов. Они пришли к выводу, что разумно полагать, что все позвоночные способны в той или иной степени страдать. [16] RSPCA Австралии недавно добавило, что доказательства того, что рыбы способны испытывать боль и страдания, растут в течение нескольких лет. [17] Группа Европейского Союза по здоровью и благополучию животных (Европейское управление по безопасности пищевых продуктов) заявила, что совокупность доказательств указывает на то, что некоторые виды рыб могут испытывать боль. [18] В отчете Британского комитета по защите сельскохозяйственных животных за 2014 год « Мнение о благополучии выращиваемой рыбы» говорится, что научный консенсус заключается в том, что рыбы могут обнаруживать вредные раздражители и реагировать на них, а также испытывать боль. [19]
Млекопитающие
[ редактировать ]В 2001 году были опубликованы исследования, показавшие, что крысы, страдающие артритом, самостоятельно выбирают обезболивающие опиаты. [20] В 2014 году ветеринарный журнал Small Animal Practice опубликовал статью о распознавании боли, которая началась так: «Способность испытывать боль присуща всем млекопитающим…» [21] сообщалось а в 2015 году в научном журнале Pain , что несколько видов млекопитающих ( крыса , мышь , кролик , кошка и лошадь ) принимают такое выражение лица в ответ на вредный стимул, которое соответствует выражению боли у людей. [22]
Птицы
[ редактировать ]Одновременно с исследованиями на крысах, страдающих артритом, были опубликованы исследования, показавшие, что птицы с нарушениями походки самостоятельно выбирают диету, содержащую карпрофен , человеческий анальгетик . [23] В 2005 году было написано: «Птичья боль, вероятно, аналогична боли, которую испытывают большинство млекопитающих». [24] а в 2014 году «...признано, что птицы воспринимают вредные раздражители и реагируют на них, а также что птицы чувствуют боль» [25]
Рептилии и амфибии
[ редактировать ]Были опубликованы ветеринарные статьи, в которых говорится, что обе рептилии [26] [27] [28] и земноводные [29] [30] [31] испытывают боль аналогично людям, и что анальгетики эффективны у этих двух классов позвоночных.
Аргумент по аналогии
[ редактировать ]В 2012 году американский философ Гэри Варнер проанализировал исследовательскую литературу о боли у животных. Его выводы суммированы в следующей таблице. [32]
| Аргумент по аналогии [32] | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Свойство | |||||||||
| Рыба | Земноводные | Рептилии | Птицы | Млекопитающие | |||||
| Имеет ноцицепторы | |||||||||
| Имеет мозг | |||||||||
| Ноцицепторы и мозг связаны | ? [а] / | ? [б] / | ? / | ||||||
| Имеет эндогенные опиоиды | |||||||||
| Анальгетики влияют на реакцию | ? [с] | ? [д] | |||||||
| Реакция на повреждающие раздражители аналогична человеческой | |||||||||
Примечания
[ редактировать ]Рассуждая по аналогии, Варнер утверждает, что любое животное, проявляющее свойства, перечисленные в таблице, может испытывать боль. На этом основании он заключает, что все позвоночные, включая рыб, вероятно, испытывают боль, но беспозвоночные, за исключением головоногих , вероятно, не испытывают боли. [32] [37]
Ракообразные
Однако некоторые исследования показывают, что ракообразные действительно демонстрируют реакции, соответствующие признакам боли и страдания. [38]
Испытывать боль
[ редактировать ]Хотя существует множество определений боли , почти все они включают два ключевых компонента.
Во-первых, ноцицепция . необходима [39] Это способность обнаруживать вредные раздражители, которые вызывают рефлекторную реакцию, которая быстро перемещает все животное или пораженную часть его тела в сторону от источника раздражителя. Понятие ноцицепции не предполагает какого-либо неблагоприятного, субъективного «ощущения» — это рефлекторное действие. Примером у людей может служить быстрое отдергивание пальца, прикоснувшегося к чему-то горячему, – отдергивание происходит до того, как действительно возникает какое-либо ощущение боли.
Второй компонент – это само переживание «боли», или страдания – внутренняя, эмоциональная интерпретация ноцицептивного опыта. Опять же, у людей это происходит, когда отдернутый палец начинает болеть через несколько мгновений после отдергивания. Таким образом, боль — это личное эмоциональное переживание. Боль нельзя напрямую измерить у других животных, включая людей; можно измерить реакцию на предположительно болезненные стимулы, но не сам опыт. Чтобы решить эту проблему при оценке способности других видов испытывать боль, используется метод аргументации по аналогии. Это основано на том принципе, что если животное реагирует на раздражитель так же, как мы, оно, скорее всего, пережило аналогичный опыт.
ноцицепция
[ редактировать ]
Ноцицепция обычно включает передачу сигнала по цепочке нервных волокон от места болевого раздражителя на периферии к спинному и головному мозгу. Этот процесс вызывает рефлекторную дуговую реакцию, генерируемую в спинном мозге и не затрагивающую головной мозг, например, вздрагивание или отдергивание конечности. Ноцицепция в той или иной форме встречается у всех основных таксонов животных . [39] Ноцицепцию можно наблюдать с помощью современных методов визуализации; и физиологическую часто можно обнаружить и поведенческую реакцию на ноцицепцию. Однако ноцицептивные реакции у животных-жертв могут быть настолько тонкими, что обученные наблюдатели (люди) не могут их воспринимать, тогда как естественные хищники могут и впоследствии нацеливаются на раненых особей. [40]
Эмоциональная боль
[ редактировать ]Иногда проводят различие между «физической болью» и «эмоциональной» или « психологической болью ». Эмоциональная боль — это боль, испытываемая при отсутствии физической травмы, например, боль, которую испытывает человек после потери любимого человека или разрыва отношений. Утверждалось, что только приматы и люди могут чувствовать «эмоциональную боль», потому что они единственные животные, у которых есть неокортекс — часть коры головного мозга, которая считается «областью мышления». Однако исследования предоставили доказательства того, что обезьяны, собаки, кошки и птицы могут проявлять признаки эмоциональной боли и демонстрировать поведение, связанное с депрессией во время или после болезненного опыта , в частности, отсутствие мотивации, вялость, анорексию и невосприимчивость к другим животным. [7]
Физическая боль
[ редактировать ]Нервные импульсы ноцицептивной реакции могут передаваться в мозг, тем самым регистрируя местоположение, интенсивность, качество и неприятность стимула. Этот субъективный компонент боли включает сознательное осознание как ощущения, так и неприятности (отвратительный, негативный аффект ). Мозговые процессы, лежащие в основе сознательного осознания неприятностей (страданий), недостаточно изучены.
Было опубликовано несколько списков критериев для установления того, испытывают ли животные, кроме человека, боль, например: [41] [42] Некоторые критерии, которые могут указывать на способность других видов, включая рыб, чувствовать боль, включают: [42]
- Имеет подходящую нервную систему и сенсорные рецепторы.
- Имеет опиоидные рецепторы и снижает реакцию на вредные раздражители при приеме анальгетиков и местных анестетиков.
- Физиологические изменения в ответ на вредные раздражители
- Демонстрирует защитные двигательные реакции, которые могут включать ограниченное использование пораженного участка, например, хромоту, растирание, удержание или аутотомию.
- Показывает обучение избеганию
- Показывает компромисс между избеганием вредных стимулов и другими мотивационными требованиями.
- Высокие когнитивные способности и чувствительность
Адаптивное значение
[ редактировать ]Адаптивная ценность ноцицепции очевидна; организм, обнаруживший вредный раздражитель, немедленно отводит конечность, придаток или все тело от вредного раздражителя и тем самым избегает дальнейшего (потенциального) повреждения. Однако особенностью боли (по крайней мере, у млекопитающих) является то, что она может приводить к гипералгезии (повышенной чувствительности к вредным раздражителям) и аллодинии (повышенной чувствительности к невредным раздражителям). Когда возникает такая повышенная сенсибилизация, адаптивная ценность менее очевидна. Во-первых, боль, возникающая из-за повышенной сенсибилизации, может быть непропорциональна фактическому повреждению тканей. Во-вторых, повышенная сенсибилизация может также стать хронической, сохраняясь после заживления тканей. Это может означать, что беспокойство вызывает не фактическое повреждение тканей, вызывающее боль, а боль, вызванная повышенной сенсибилизацией. Это означает, что процесс сенсибилизации иногда называют дезадаптивным . Часто предполагают, что гипералгезия и аллодиния помогают организмам защитить себя во время заживления, но экспериментальные доказательства, подтверждающие это, отсутствуют. [43] [44]
В 2014 году адаптивная ценность сенсибилизации из-за травмы была проверена с использованием хищнических взаимодействий между длинноперым прибрежным кальмаром ( Doryteuthis pealeii ) и черноморским окунем ( Centropristis striata ), которые являются естественными хищниками этого кальмара. Если на раненых кальмаров напал окунь, они начали свое защитное поведение раньше (на что указывает большее расстояние оповещения и большее расстояние начала полета), чем неповрежденные кальмары. анестетик (1% этанол и MgCl 2 Если перед травмой вводится ), это предотвращает сенсибилизацию и блокирует поведенческий эффект. Авторы утверждают, что это исследование является первым экспериментальным доказательством, подтверждающим аргумент о том, что ноцицептивная сенсибилизация на самом деле является адаптивной реакцией на травмы. [40]
Был задан вопрос: «Если рыбы не могут чувствовать боль, то почему скаты имеют чисто защитные шипы на хвосте, которые доставляют яд? Предковыми хищниками скатов являются рыбы. И почему у многих рыб есть защитные плавники, некоторые из которых также содержат яд, вызывающий боль в люди?" [45]
Результаты исследования
[ редактировать ]Периферическая нервная система
[ редактировать ]Рецепторы
[ редактировать ]Примитивные рыбы, такие как миноги ( Petromyzon marinus ), имеют в коже свободные нервные окончания, реагирующие на тепло и механическое давление. Однако поведенческие реакции, связанные с ноцицепцией, не зарегистрированы, а также трудно определить, являются ли механорецепторы у миноги действительно ноцицептивно-специфичными или просто давильно-специфичными. [46]
Ноцицепторы у рыб были впервые идентифицированы в 2002 году. [47] [48] Целью исследования было определить, присутствуют ли ноцицепторы в тройничном нерве на голове форели, а также наблюдать физиологические и поведенческие последствия длительной вредной стимуляции. В губы радужной форели вводили уксусную кислоту, а другой группе вводили пчелиный яд. Эти вещества были выбраны потому, что протоны кислоты стимулируют ноцицептивные нервы млекопитающих и лягушек. [49] хотя яд оказывает воспалительное действие на млекопитающих [50] и оба, как известно, болезненны для людей. Рыбы демонстрировали аномальное поведение, такое как раскачивание из стороны в сторону и потирание губами стенок и дна аквариума. У них увеличилась частота дыхания, и они сократили количество плаваний. Кислотная группа тоже потерла губы о гравий. Растирание поврежденного участка для облегчения боли было продемонстрировано на людях и других млекопитающих. [51] На лице и голове радужной форели было расположено 58 рецепторов. Двадцать два из этих рецепторов можно отнести к ноцицепторам, поскольку они реагировали на механическое давление и тепло (более 40 °С). Восемнадцать также отреагировали на уксусную кислоту. Реакция рецепторов на механическую, повреждающую термическую и химическую стимуляцию четко характеризовала их как полимодальные ноцицепторы. Они имели сходные свойства с теми, что обнаружены у земноводных, птиц. [52] [53] и млекопитающие, включая человека. [54] Форели, которой вводили яд или кислоту, потребовалось примерно 3 часа, чтобы возобновить прием пищи, тогда как группе, получавшей физиологический раствор, и контрольной группе потребовалось примерно 1 час. Это может быть охранительное поведение, когда животные избегают использования больной конечности, предотвращая продолжающуюся боль и причинение вреда этой области. [52]
Радужная форель ( Oncorhynchus mykiss ) имеет полимодальные ноцицепторы на морде и морде, которые реагируют на механическое давление, температуры в вредном диапазоне (> 40 ° C) и 1% уксусную кислоту (химический раздражитель). В целом было обнаружено, что кожные рецепторы более чувствительны к механическим раздражителям, чем у млекопитающих и птиц, при этом некоторые из них реагируют на раздражители лишь при 0,001 г. У человека требуется не менее 0,6 г. Это может быть связано с тем, что рыбья кожа легче повреждается, что требует более низких порогов ноцицепторов. [47] [55] [56] [57] Дальнейшие исследования показали, что ноцицепторы более широко распространены на теле радужной форели, а также трески и карпа. Наиболее чувствительные участки тела — вокруг глаз, ноздрей, мясистые части хвоста, грудных и спинных плавников . [13] [58]
У радужной форели также есть роговицы ноцицепторы . Из 27 рецепторов, исследованных в одном исследовании, семь были полимодальными ноцицепторами и шесть — механотермическими ноцицепторами. Механические и температурные пороги были ниже, чем у кожных рецепторов, что указывает на более высокую чувствительность роговицы. [59]
Костные рыбы обладают ноцицепторами, которые по функциям аналогичны таковым у млекопитающих. [12]
Нервные волокна
[ редактировать ]Существует два типа нервных волокон, вызывающих боль у рыб. Нервные волокна группы C представляют собой тип чувствительных нервных волокон, которые не имеют миелиновой оболочки и имеют небольшой диаметр, что означает низкую скорость нервной проводимости . Страдания, которые люди связывают с ожогами, зубной болью или травмами, вызваны активностью С-волокон. Типичный кожный нерв человека содержит 83% нервных волокон группы С. [60] Волокна А-дельта — это еще один тип чувствительных нервных волокон, однако они миелинизированные и поэтому передают импульсы быстрее, чем немиелинизированные С-волокна. Волокна А-дельта передают сигналы холода, давления и некоторых болевых сигналов и связаны с острой болью, которая приводит к «отстранению» от вредных раздражителей.
Костные рыбы содержат волокна как группы C, так и A-дельта, составляющие 38,7% (в совокупности) волокон хвостового нерва карпа и 36% тройничного нерва радужной форели. Однако только 5% и 4% из них составляют волокна C у карпа и радужной форели соответственно. [60] [61]
У некоторых видов хрящевых рыб имеются А-дельта-волокна, однако С-волокна либо отсутствуют, либовстречаются в очень небольшом количестве. [60] [62] [63] Agnatha и миноги) в основном ( мисины содержат волокна группы C. [64]
Центральная нервная система
[ редактировать ]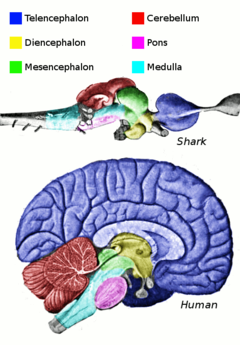
Центральная нервная система (ЦНС) рыб содержит спинной мозг , продолговатый мозг и головной мозг , разделенный на конечный мозг , промежуточный мозг , средний мозг и мозжечок .
У рыб, как и у других позвоночных, ноцицепция проходит от периферических нервов по спинномозговым нервам и передается через спинной мозг в таламус . Таламус связан с конечным мозгом множеством связей через паллиум серого вещества , которое, как было продемонстрировано, получает нервные реле для вредных и механических раздражителей. [64] [65]
Основными путями, которые передают информацию о боли от периферии в мозг, являются спиноталамический тракт (тело) и тройничный тракт (голова). Оба были изучены у бесчелюстных, костистых и пластиножаберных рыб (тройничное у обыкновенного карпа, спиноталамический тракт морской малиновки Prionotus carolinus ). [66]
Мозг
[ редактировать ]Если сенсорные реакции у рыб ограничены спинным и задним мозгом, их можно рассматривать как просто рефлекторные. Однако записи спинного мозга, мозжечка, тектума и конечного мозга как у форели, так и у золотой рыбки ( Carassius auratus ) показывают, что все они реагируют на вредные раздражители. Это указывает на ноцицептивный путь от периферии к высшим ЦНС рыб. [67]
Микроматричный анализ экспрессии генов показывает, что мозг активен на молекулярном уровне в переднем , среднем и заднем мозге обыкновенного карпа и радужной форели. Несколько генов, участвующих в ноцицепции млекопитающих, такие как нейротрофический фактор головного мозга (BDNF) и каннабиноидный рецептор CB1, регулируются в мозге рыб после ноцицептивного события. [68] [69]
Соматосенсорные вызванные потенциалы (СВП) — это слабые электрические реакции в ЦНС после стимуляции периферических сенсорных нервов. Это также указывает на существование пути от периферических ноцицепторов к высшим областям мозга. У золотой рыбки, радужной форели, атлантического лосося ( Salmo salar ) и атлантической трески ( Gadus morhua ) было продемонстрировано, что предположительно безвредная и вредная стимуляция вызывает СЭП в различных областях мозга, включая конечный мозг. [70] которые могут опосредовать координацию болевой информации. [71] Более того, множественные исследования функциональной магнитно-резонансной томографии (фМРТ) с участием нескольких видов рыб показали, что при предполагаемой боли в переднем мозге наблюдается глубокая активность, которая очень напоминает ту, что наблюдается у людей, и может быть принята как свидетельство пережитого. боли у млекопитающих. [72] [73]
Таким образом, у рыб, переживающих потенциально болезненное событие, активируются «высшие» области мозга на молекулярном, физиологическом и функциональном уровнях. Снеддон заявил: «Это придает большое значение предположению о том, что рыбы испытывают некоторую форму боли, а не ноцицептивные ощущения». [74]
Опиоидная система и эффекты анальгетиков
[ редактировать ]
Кистистые рыбы имеют функциональную опиоидную систему, которая включает наличие опиоидных рецепторов, аналогичных таковым у млекопитающих. [75] [76] Опиоидные рецепторы уже присутствовали у происхождения челюстных позвоночных 450 миллионов лет назад. [77] Все четыре основных опиоидных рецепторов типа ( дельта , каппа , мю и NOP ) консервативны у позвоночных, даже у примитивных бесчелюстных рыб (агнатстомы). [64]
Те же анальгетики и анестетики, которые применяются у людей и других млекопитающих, часто применяются для рыб в ветеринарной медицине. Эти химические вещества действуют на ноцицептивные пути, блокируя сигналы в мозг, где эмоциональные реакции на сигналы далее обрабатываются определенными частями мозга, обнаруженными у амниот (« высших позвоночных »). [78] [79]
Эффекты морфия
[ редактировать ]
Предварительное лечение морфином ( болеутоляющим средством для людей и других млекопитающих) оказывает дозозависимое антиноцицептивное действие. [80] и смягчает поведенческие реакции и реакцию вентиляции радужной форели на вредные раздражители.
Когда уксусную кислоту вводят в губы радужной форели, они демонстрируют аномальное поведение, такое как раскачивание из стороны в сторону и потирание губами стенок и дна резервуаров, а скорость их вентиляции увеличивается. Инъекции морфина уменьшают как аномальное поведение, связанное с вредными стимулами, так и увеличение частоты дыхания. [81] Когда тот же вредный стимул применяется к рыбкам данио ( Danio rerio ), они реагируют снижением своей активности. Как и в случае с радужной форелью, морфин, введенный перед инъекцией кислоты, ослабляет снижение активности дозозависимым образом. [71]
Введение уксусной кислоты в губы радужной форели вызывает снижение у них естественной неофобии (боязни новизны); это устраняется введением морфия. [13]
У золотых рыбок, которым вводили морфин или физиологический раствор, а затем подвергали воздействию неприятных температур, рыбы, которым вводили физиологический раствор, действовали с защитным поведением, указывающим на тревогу, настороженность и страх, тогда как те, которым вводили морфин, этого не делали. [82]
Эффекты других анальгетиков
[ редактировать ]Нейромедиатор, вещество Р, а также анальгетические опиоидные энкефалины и β-эндорфин , которые действуют как эндогенные анальгетики у млекопитающих, присутствуют в рыбах. [83]
Различные анальгетики по-разному действуют на рыб. В исследовании эффективности трех типов анальгетиков бупренорфина (опиоида), карпрофена (нестероидного противовоспалительного препарата) и лидокаина (местного анестетика) в качестве показателей боли использовались частота вентиляции легких и время возобновления кормления. Бупренорфин оказывал ограниченное влияние на реакцию рыб, карпрофен смягчал последствия вредной стимуляции вовремя возобновления кормления, однако лидокаин снижал все поведенческие показатели. [84] Прием аспирина предотвращает изменение поведения, вызванное уксусной кислотой. [85]
Трамадол также повышает ноцицептивный порог у рыб, что является еще одним доказательством наличия у рыб антиноцицептивной опиоидной системы. [13] [86]
Эффекты налоксона
[ редактировать ]Налоксон является антагонистом мю-опиоидных рецепторов , который у млекопитающих сводит на нет анальгезирующее действие опиоидов. Как взрослые, так и пятидневные личинки рыбок данио демонстрируют поведенческие реакции, свидетельствующие о боли в ответ на инъекцию или разбавление уксусной кислоты. Антиноцицептивные свойства морфина или бупренорфина обращаются вспять, если взрослые, [71] или личинки, [87] одновременно лечатся налоксоном. И налоксон, и пролиллейцилглицинамид (еще один антагонист опиатов у млекопитающих) снижали обезболивающее действие морфина при электрошоке, получаемом золотыми рыбками, что указывает на то, что они могут действовать как антагонисты опиатов у рыб. [88] [89]
Физиологические изменения
[ редактировать ]Физиологические изменения рыб в ответ на вредные раздражители включают увеличение скорости вентиляции. [68] [80] [81] [90] и кортизола . уровень [86]
Защитные реакции
[ редактировать ]



Исследования показывают, что рыбы проявляют защитные поведенческие реакции на предположительно болезненные стимулы.
Когда уксусная кислота или пчелиный яд вводится в губы радужной форели, они демонстрируют аномальное раскачивание из стороны в сторону грудных плавников , трутся губами о стенки и дно резервуаров. [91] и увеличить скорость вентиляции. [90] Когда уксусную кислоту вводят в губы рыбок данио, они реагируют снижением своей активности. Величина этой поведенческой реакции зависит от концентрации уксусной кислоты. [71]
Поведенческие реакции на вредный раздражитель различаются у разных видов рыб. ), подвергшийся вредному стимулированию, Обыкновенный карп ( Cyprinus carpio демонстрирует аномальное раскачивание и трётся губами о стенки аквариума, но не меняет других поведений или скорости вентиляции. Напротив, рыбки данио ( Danio rerio ) сокращают частоту плавания и увеличивают скорость дыхания, но не демонстрируют аномального поведения. Радужная форель, как и данио, сокращает частоту плавания и увеличивает скорость вентиляции. [92] Нильская тилапия ( Oreochromis niloticus ) в ответ на подрезание хвостового плавника увеличивает свою плавательную активность и проводит больше времени в светлой зоне аквариума. [93]
Со времени этой первой работы Снеддон и ее коллеги показали, что радужная форель, обыкновенный карп и рыбка данио, испытывающие вредную стимуляцию, демонстрируют быстрые изменения в физиологии и поведении, которые сохраняются до 6 часов и, следовательно, не являются простыми рефлексами. [66]
Пятидневные личинки рыбок данио демонстрируют зависимое от концентрации увеличение двигательной активности в ответ на различные концентрации разбавленной уксусной кислоты. Это увеличение двигательной активности сопровождается увеличением мРНК ЦОГ-2 , демонстрируя, что ноцицептивные пути также активируются. [87]
Рыбы по-разному реагируют на разные вредные раздражители, даже если они внешне похожи. Это указывает на то, что реакция является гибкой, а не просто ноцицептивным рефлексом. Атлантическая треска, которой вводили в губу уксусную кислоту и капсаицин или прокалывали губу коммерческим рыболовным крючком, демонстрировала разные реакции на эти три типа вредной стимуляции. Треска, обработанная уксусной кислотой и капсаицином, чаще зависала у дна аквариума и реже использовала укрытия. Однако у пойманной на крючок трески наблюдалось лишь кратковременное покачивание головой. [90]
Обучение избеганию
[ редактировать ]Ранние эксперименты предоставили доказательства того, что рыбы учатся реагировать на предположительно вредные раздражители. Например, рыбы-жабы ( Batrachoididae ) хрюкают при ударе электрическим током, но после повторных ударов они хрюкают просто при виде электрода. [94] [95] Более поздние исследования показывают, что и золотая рыбка, и форель учатся избегать мест, где их подвергают ударам электрическим током. Колюшки получают некоторую защиту от хищных рыб благодаря своим шипам. Исследователи обнаружили, что щука и окунь сначала расхватали их, но затем отвергли. После нескольких опытов щука и окунь научились вообще избегать колюшек. Когда иглы колюшки были удалены, их защита исчезла. [96] Более того, это обучение избеганию является гибким и зависит от интенсивности стимула. [86] [97] [98] [99]
Компромиссы в мотивации
[ редактировать ]
Болезненный опыт может изменить мотивацию нормальных поведенческих реакций.
В исследовании 2007 года золотых рыбок приучали питаться в том месте аквариума, где впоследствии их подвергали удару электрическим током. Количество попыток кормления и время, проведенное в зоне кормления/шока, уменьшалось с увеличением интенсивности шока, а с увеличением лишения пищи количество и продолжительность попыток кормления увеличивались, равно как и реакции побега при входе в эту зону. Исследователи предположили, что у золотых рыбок есть компромисс между мотивацией к кормлению и мотивацией избегать острого вредного раздражителя. [98]
Радужная форель естественным образом избегает новизны (т. е. она неофобна ). Виктория Брейтуэйт описывает исследование, в котором яркий кубик Лего помещается в аквариум с радужной форелью. Форель, которой вводили в губу небольшое количество физиологического раствора, категорически избегала кубика Лего, однако форель, которому вводили уксусную кислоту, проводила значительно больше времени возле блока Лего. Когда исследование было повторено, но рыбам также давали морфин, реакция избегания вернулась у рыб, которым вводили уксусную кислоту, и ее нельзя было отличить от реакций рыб, которым вводили физиологический раствор. [13] [100]
Чтобы изучить возможность компромисса между реакцией на вредный стимул и хищничеством, исследователи представили радужной форели конкурирующий стимул — сигнал хищника. Рыбы, подвергшиеся токсичному стимулированию, перестают проявлять реакцию против хищников, что указывает на то, что боль становится их основной мотивацией. В том же исследовании изучался потенциальный компромисс между реакцией на вредный стимул и социальным статусом. Реакция форели, подвергшейся ядовитому воздействию, варьировалась в зависимости от того, насколько знакома рыба, с которой ее поместили. Исследователи предположили, что результаты мотивационных изменений и компромиссов свидетельствуют о центральной обработке боли, а не просто демонстрируют ноцицептивный рефлекс. [100] [101]
Оплата стоимости обезболивания
[ редактировать ]Рыбки данио, получившие доступ к бесплодной, ярко освещенной камере или обогащенной камере, предпочитают обогащенную зону. Когда этим рыбам вводят уксусную кислоту или физиологический раствор в качестве контроля, они все равно выбирают ту же обогащенную камеру. Однако, если анальгетик растворяется в бесплодной, менее предпочтительной камере, рыбки данио, которым введена ядовитая кислота, теряют свое предпочтение и тратят болееполовину своего времени проводят в ранее менее благоприятном и бесплодном помещении. Это предполагает компромисс в мотивации, и, более того, они готовы заплатить цену, чтобы войти в менее предпочтительную среду для доступа к обезболиванию. [41]
Когнитивные способности и чувствительность
[ редактировать ]Способность рыб к обучению, продемонстрированная в ряде исследований, указывает на сложные когнитивные процессы , которые более сложны, чем простое ассоциативное обучение . Примеры включают способность узнавать социальных товарищей, избегание (в течение нескольких месяцев или лет) мест, где они столкнулись с хищником или были пойманы на крючок, и формирование мысленных карт . [83]
Утверждалось, что, хотя высокие когнитивные способности могут указывать на большую вероятность испытать боль, они также дают этим животным большую способность справляться с ней, в результате чего у животных с более низкими когнитивными способностями возникают большие проблемы с преодолением боли. [102]
Критерии восприятия боли
[ редактировать ]Ученые также предположили, что в сочетании с аргументами по аналогии можно использовать критерии физиологии или поведенческих реакций для оценки возможности восприятия боли животными, отличными от человека. Ниже приводится таблица критериев, предложенных Sneddon et al. [41]
| Критерии | ||||
|---|---|---|---|---|
| Бесчелюстная рыба
| Хрящевая рыба
| Костистая рыба
| Лопастеперая рыба
| |
| Имеет ноцицепторы | ? | ? | ? | |
| Пути к центральной нервной системе | ? | ? | ? | |
| Центральная обработка в мозге | ? | ? | ? | |
| Рецепторы анальгезирующих препаратов | ? | ? | ? | |
| Физиологические реакции | ? | ? | ? | |
| Отстранение от вредных раздражителей | ? | ? | ? | |
| Поведенческие изменения по сравнению с нормой | ? | ? | ? | |
| Защитное поведение | ? | ? | ? | |
| Реакция снижается при приеме анальгетиков | ? | ? | ? | |
| Самостоятельное введение анальгезии | ? | ? | ? | |
| Реакции с высоким приоритетом над другими стимулами | ? | ? | ? | |
| Оплатить стоимость доступа к обезболиванию | ? | ? | ? | |
| Измененный поведенческий выбор/предпочтения | ? | ? | ? | |
| Облегчение обучения | ? | ? | ? | |
| Трение, хромота или охрана | ? | ? | ? | |
| Плата за избежание стимулирования | ? | ? | ? | |
| Компромиссы с другими требованиями | ? | ? | ? |



В таблице, ![]() указывает на положительные доказательства, а ? означает, что он не был протестирован или доказательств недостаточно.
указывает на положительные доказательства, а ? означает, что он не был протестирован или доказательств недостаточно.
Социальные последствия
[ редактировать ]
Учитывая, что некоторые интерпретировали существующую научную информацию как предположение о том, что рыбы могут чувствовать боль, [103] было предложено принципы предосторожности к коммерческому рыболовству, что, вероятно, будет иметь множественные последствия. применять [103]
И ученые, и защитники животных выразили обеспокоенность по поводу возможных страданий (боли и страха), причиняемых рыбам при ловле рыбы . [104] [105] [106]
Другие социальные последствия того, что рыба испытывает боль, включают острое и хроническое воздействие загрязняющих веществ, коммерческое и спортивное рыболовство (например, травмы во время траления, мечение/обрезание плавников во время оценки запасов, повреждение тканей, физическое истощение и серьезный дефицит кислорода во время отлова, боль и стресс во время убоя). , использование живой наживки), аквакультура (например, мечение/обрезание плавников, высокая плотность посадки, приводящая к повышенной агрессии, лишение пищи для лечения болезней или перед сбором урожая, удаление из воды для рутинного содержания, боль во время убоя), декоративная рыба (например, отлов сублетальное отравление, постоянное неблагоприятное физическое состояние из-за селекционного разведения), научные исследования (например, генетическая модификация) могут иметь пагубные последствия для благосостояния, преднамеренно навязанные неблагоприятные физические, физиологические и поведенческие состояния, электроловля, мечение, обрезка плавников или иное маркирование рыба, процедуры обращения с ней, которые могут привести к травмам. [46] [107]
Броуман и др. [108] предполагают, что если нормативно-правовая база продолжит действовать по нынешней траектории (добавив больше таксонов водных животных к уже регулируемым), деятельность в некоторых секторах может быть строго ограничена или даже запрещена. Они также утверждают, что распространение правовой защиты на водных животных является выбором общества, но они подчеркивают, что этот выбор не следует приписывать сильной поддержке со стороны ряда исследований, которые еще не существуют и, возможно, никогда не будут существовать, а также последствиям принятия этого решения. необходимо тщательно взвесить.
Законодательство
[ редактировать ]В Великобритании законодательство, защищающее животных во время научных исследований, «Закон о животных (научных процедурах) 1986 года», защищает рыб с момента, когда они становятся способными к самостоятельному питанию. [109] Законодательство, защищающее животных в большинстве других случаев в Великобритании, - это «Закон о защите животных 2006 года», в котором говорится, что в Законе «животное» означает позвоночное животное, кроме человека». [110] явно включая рыбу.
В США законом, защищающим животных во время научных исследований, является «Закон о защите животных». [111] Это исключает защиту «хладнокровных» животных, в том числе рыб. [112]
В норвежском Законе о правах животных 1974 года говорится, что он касается млекопитающих, птиц, лягушек, саламандр, рептилий, рыб и ракообразных. [113]
В статье Говарда Броумана и его коллег, опубликованной в 2018 году, представлен обзор того, что означают различные точки зрения на боль и благополучие рыб в контексте аквакультуры, коммерческого рыболовства, любительского рыболовства и исследований. [108]
Споры
[ редактировать ]Нервная система
[ редактировать ]Рецепторы и нервные волокна
[ редактировать ]Утверждалось, что рыбы не могут чувствовать боль, поскольку у них нет достаточной плотности соответствующих нервных волокон. Типичный кожный нерв человека содержит 83% нервных волокон группы С. [114] однако те же нервы у людей с врожденной нечувствительностью к боли содержат только 24–28% волокон С-типа. [114] Основываясь на этом, Джеймс Роуз из Университета Вайоминга утверждал, что отсутствие волокон С-типа у хрящевых акул и скатов указывает на то, что передача сигналов, ведущая к восприятию боли, скорее всего, невозможна, а низкие цифры у костистых рыб (например, 5% для карпа и форели) указывают на то, что для этих рыб это также маловероятно. [114] Волокна А-дельта-типа, которые, как полагают, вызывают реакцию избегания, часто встречаются у костистых рыб, хотя у акул и скатов они не обнаружены. [114] Роуз заключает, что рыбы хорошо выжили в эволюционном смысле без полного спектра ноцицепции, типичной для человека или других млекопитающих. [114] Профессор Кулум Браун из Университета Маккуори в Сиднее утверждает, что отсутствие доказательств использовалось как доказательство отсутствия; Это фундаментальное неправильное толкование научного метода, которое было принято за предположение, что акулы и скаты не могут чувствовать боли. Он утверждает, что тот факт, что ноцицепция возникает у бесчелюстных рыб, [115] а также у костистых рыб, [116] предполагает, что наиболее экономное объяснение состоит в том, что акулы действительно обладают такими способностями, но нам еще предстоит понять, что рецепторы или волокна, которые мы идентифицировали, действуют по-новому. Он указывает, что альтернативное объяснение состоит в том, что пластиножаберные утратили способность ноцицепции, и нужно было бы придумать очень убедительный аргумент в пользу адаптивного значения такой потери для одного таксона во всем животном мире. [117] Профессор Брум из Кембриджского университета утверждает, что чувство боли дает активным сложным позвоночным селективное преимущество посредством обучения и реагирования, позволяя им выживать в окружающей среде. Системы боли и страха филогенетически чрезвычайно древние, поэтому маловероятно, что они внезапно появились у млекопитающих или людей. [118]
Мозг
[ редактировать ]В 2002 году Роуз опубликовал обзоры, в которых утверждалось, что рыбы не могут чувствовать боль, потому что у них нет неокортекса в мозгу. [119] [120] Этот аргумент также исключает восприятие боли у большинства млекопитающих, а также у всех птиц и рептилий. [52] [72] Однако в 2003 году исследовательская группа под руководством Линн Снеддон пришла к выводу, что мозг радужной форели активирует нейроны так же, как человеческий мозг, когда испытывает боль. [121] [122] Роуз раскритиковал исследование, заявив, что оно ошибочно, главным образом потому, что оно не предоставило доказательств того, что рыбы обладают «сознательным сознанием, особенно таким осознанием, которое по смыслу похоже на наше». [123]
Роуз, а в последнее время и Брайан Ки. [124] [125] из Университета Квинсленда, утверждают, что, поскольку мозг рыбы сильно отличается от мозга человека, рыбы, вероятно, не обладают сознанием, как люди, и хотя рыбы могут реагировать так же, как люди реагируют на боль, реакции в случае с рыбой есть и другие причины. Исследования, показавшие, что рыбы могут чувствовать боль, путают ноцицепцию с ощущением боли, говорит Роуз. «Боль зависит от осознания. Ключевым вопросом является различие между ноцицепцией и болью. Человек, находящийся под наркозом в операционной, по-прежнему будет физически реагировать на внешний раздражитель, но он или она не будет чувствовать боли». [126] По мнению Роуза и Ки, литература, посвященная вопросу сознания рыб, склонна к антропоморфизму, и необходимо проявлять осторожность, чтобы избежать ошибочного приписывания рыбам человеческих способностей. [127] Однако ни одно другое животное не может напрямую сообщать о том, что оно чувствует и думает, а Роуз и Ки не опубликовали экспериментальные исследования, показывающие, что рыбы не чувствуют боли. [128] Снеддон предполагает, что вполне возможно, что виды с другим эволюционным путем могли развить разные нервные системы для выполнения одних и тех же функций (то есть конвергентная эволюция ), как показали исследования мозга птиц. [129] Ки соглашается, что феноменальное сознание, вероятно, встречается у млекопитающих и птиц, но не у рыб. [124] Специалист по поведению животных Темпл Грандин утверждает, что рыбы все еще могут обладать сознанием без неокортекса, потому что «разные виды могут использовать разные структуры и системы мозга для выполнения одних и тех же функций». [122] Снеддон предполагает, что предположение о внезапном возникновении функции без примитивной формы противоречит законам эволюции. [130]
Другие исследователи также полагают, что сознание животных не требует неокортекса, но может возникнуть из гомологичных подкорковых сетей мозга. [11] Было высказано предположение, что цепи ствола мозга могут генерировать боль. Сюда входят исследования детей с анэнцефалией , которые, несмотря на отсутствие значительной части коры головного мозга, выражают эмоции. Существуют также данные исследований активации, показывающие чувства, опосредованные стволом мозга, у нормальных людей и реакции отказа плода на вредную стимуляцию, но до развития коры головного мозга. [131]
В статьях, опубликованных в 2017 и 2018 годах, Майкл Вудрафф [132] [133] обобщено значительное количество научных статей, которые, вопреки выводам Роуза и Ки, убедительно подтверждают гипотезу о том, что нейроанатомическая организация мантии рыб и ее связи с субпаллиальными структурами, особенно с прегломерулярным ядром и тектумом, сложны достаточно, чтобы быть аналогом схемы коры головного мозга и таламуса, которая, как предполагается, лежит в основе разума млекопитающих. К этим анатомическим наблюдениям он добавил нейрофизиологические и поведенческие данные, которые также подтверждают гипотезу о том, что паллий является важной частью иерархической сети, предложенной Фейнбергом и Маллаттом и лежащей в основе сознания рыб. [134]
Защитные реакции
[ редактировать ]Работа Снеддона охарактеризовала поведенческие реакции радужной форели, карпа и рыбки данио. [66] Однако, когда Ньюби и Стивенс повторили эти эксперименты без анестезии, раскачивания и трения не наблюдалось, что позволяет предположить, что некоторые из предполагаемых болевых реакций, наблюдаемых Снеддоном и его коллегами, вероятно, были связаны с выходом рыбы из анестезии. Но Ньюби и Стивенс, пытаясь повторить исследование, проведенное лабораторией Снеддона, использовали протокол, отличный от уже опубликованного. Отсутствие аномального поведения при трении и возобновление кормления в эксперименте Ньюби и Стивенса можно объяснить тем, что они вводили такую высокую концентрацию кислоты. Если в центральную нервную систему не поступает ноцицептивная информация, то никаких поведенческих изменений не произойдет. Снеддон утверждает, что это демонстрирует важность следования экспериментальному плану опубликованных исследований для получения сопоставимых результатов. [135] [136] [137]
Некоторые исследователи спорят по поводу определения боли, используемого в поведенческих исследованиях, поскольку записанные наблюдения были противоречивыми, неподтвержденными и неповторяемыми другими исследователями. [60] В 2012 году Роуз утверждала, что рыбы возобновляют «нормальное питание и активность сразу или вскоре после операции». [60] Но Стоскопф предположил, что рыбы могут тонко реагировать на хронические раздражители. К ним относятся изменения цвета, изменения позы и различное использование толщи воды, и эти более тонкие проявления поведения могут быть упущены, в то время как Вагнер и Стивенс заявили, что необходимы дальнейшие исследования, изучающие большее количество моделей поведения. [138] [139]
Нордгрин сказал, что обнаруженные ими поведенческие различия в ответ на некомфортные температуры показали, что рыбы чувствуют как рефлекторную, так и когнитивную боль. [140] «Эксперимент показывает, что рыбы не только реагируют на болевые раздражители рефлексами, но и меняют свое поведение после события», — сказал Нордгрин. «Вместе с тем, что мы знаем из экспериментов, проведенных другими группами, это указывает на то, что рыбы сознательно воспринимают тестовую ситуацию как болезненную и переключаются на поведение, свидетельствующее о том, что они пережили аверсивный опыт». [140] В 2012 году Роуз и другие проанализировали это и дальнейшие исследования, которые пришли к выводу, что у рыб наблюдалась боль. Они пришли к выводу, что результаты такого исследования обусловлены плохим дизайном и неправильной интерпретацией, и что исследователи не смогли отличить бессознательное обнаружение повреждающих стимулов (ноцицепцию) от сознательной боли. [60]
В 2018 году Снеддон, Дональд Брум , Кулум Браун и другие опубликовали статью, в которой выяснилось, что, несмотря на эмпирические доказательства, скептики по-прежнему отрицают у рыб что-либо, кроме рефлекторных реакций, и заявляют, что они неспособны к сложным когнитивным способностям. Недавние исследования [141] [142] по обучению показали, что рыбы-чистильщики, а также попугаи лучше справляются с сложными учебными задачами, чем шимпанзе, орангутанги или капуцины, в которых им приходится учиться отличать надежные источники пищи от ненадежных. Золотые рыбки учатся избегать мест, где их ударили током. Даже если в этом месте ранее была доставлена пища и у рыб есть сильная мотивация проводить там время, они избегают ее в течение трех дней, после чего обменивают свой голод на риск получить еще один шок. Это показывает сложное принятие решений, выходящее за рамки простых рефлексов. [128]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Эбботт Ф.В., Франклин К.Б., Уэстбрук Ф.Р. (январь 1995 г.). «Формалиновый тест: оценка свойств первой и второй фаз болевой реакции у крыс». Боль . 60 (1): 91–102. дои : 10.1016/0304-3959(94)00095-В . ПМИД 7715946 . S2CID 35448280 .
- ^ Перейти обратно: а б Карбоне Л. (2004). Чего хотят животные: опыт и пропаганда политики благополучия лабораторных животных . Издательство Оксфордского университета. п. 149. ИСБН 978-0-19-516196-0 .
- ^ Раднер Д., Раднер М. (1989). Сознание животных . Буффало: Книги Прометея. ISBN 978-0-87975-459-4 .
- ^ Харрисон П. (1992). «Декарт о животных». Философский ежеквартальный журнал . 42 (167): 219–227. дои : 10.2307/2220217 . JSTOR 2220217 .
- ^ Бентам Дж (1879). Введение в принципы морали и законодательства . Кларендон Пресс.
- ^ Дарвин, Чарльз (1871). Происхождение человека в отношении пола .
- ^ Перейти обратно: а б Снеддон Л.У. «Могут ли животные чувствовать боль?» . Добро пожаловать Траст. Архивировано из оригинала 13 апреля 2012 года . Проверено 24 сентября 2015 г.
- ^ Перейти обратно: а б Роллин Б. (1989). Неуслышанный крик: сознание животных, боль животных и наука . Издательство Оксфордского университета. стр. 117–118. ISBN 978-0-19-286104-7 . цитируется в Carbone 2004, с. 150
- ^ Аллен С. (январь 1998 г.). «Оценка познания животных: этологические и философские перспективы». Журнал зоотехники . 76 (1): 42–7. дои : 10.2527/1998.76142x . ПМИД 9464883 .
- ^ Гриффин Д.Р., Спек ГБ (январь 2004 г.). «Новые доказательства сознания животных». Познание животных . 7 (1): 5–18. дои : 10.1007/s10071-003-0203-x . ПМИД 14658059 . S2CID 8650837 .
- ^ Перейти обратно: а б Низкий P (7 июля 2012 г.). Панксепп Дж., Рейсс Д., Эдельман Д., Ван Суиндерен Б., Лоу П., Кох С. (ред.). «Кембриджская декларация о сознании» (PDF) . Кембриджский университет.
- ^ Перейти обратно: а б Снеддон, Линн У. (1 апреля 2015 г.). «Боль у водных животных» . Журнал экспериментальной биологии . 218 (7): 967–976. дои : 10.1242/jeb.088823 . ISSN 0022-0949 . ПМИД 25833131 . S2CID 130495 .
- ^ Перейти обратно: а б с д и Брейтуэйт V (2010). Рыбы чувствуют боль? . Издательство Оксфордского университета. ISBN 978-0-19-161396-8 .
- ^ Брум, Дональд (1 января 2016 г.). «Рыбий мозг и поведение указывают на способность чувствовать боль» . Чувство животных . 1 (3). дои : 10.51291/2377-7478.1031 . ISSN 2377-7478 .
- ^ «Руководство AVMA по эвтаназии животных: издание 2013 г.» (PDF) . Американская ветеринарная медицинская ассоциация . 2013. с. 12 . Проверено 4 октября 2021 г.
- ^ Председатель, лорд Медуэй (1980). Отчет RSPCA о комиссии по расследованию стрельбы и рыбной ловли (1976–1979) . Опубликовано Группой по расследованию стрельбы и рыбной ловли. Архивировано из оригинала 3 марта 2021 года.
- ^ «Чувствуют ли рыбы боль? – База знаний RSPCA» . Проверено 16 октября 2021 г.
- ^ «Общий подход к благополучию рыб и концепции чувствительности рыб» . Журнал EFSA . 7 (2): 954. 2009. doi : 10.2903/j.efsa.2009.954 . ISSN 1831-4732 .
- ^ Комитет по защите сельскохозяйственных животных (2014 г.). «Отчет Комитета по защите сельскохозяйственных животных за 2014 год, Мнение о благополучии выращиваемой рыбы» (PDF) : 30 . Проверено 19 октября 2021 г.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Колпаерт, Тарайр Дж.П., Аллиага М., Брюинз Слот Л.А., Аттал Н., Кук В. (март 2001 г.). «Самовведение опиатов как показатель хронической ноцицептивной боли у крыс с артритом». Боль . 91 (1–2): 33–45. дои : 10.1016/s0304-3959(00)00413-9 . ПМИД 11240076 . S2CID 24858615 .
- ^ Мэтьюз К., Кронен П.В., Ласеллес Д., Нолан А., Робертсон С., Стегалл П.В. и др. (июнь 2014 г.). «Руководство по распознаванию, оценке и лечению боли: члены Глобального совета по боли WSAVA и соавторы этого документа». Журнал практики мелких животных . 55 (6): Е10-68. дои : 10.1111/jsap.12200 . ПМИД 24841489 .
- ^ Чемберс, Коннектикут, Могил Дж.С. (май 2015 г.). «Онтогенез и филогения лицевого выражения боли» . Боль . 156 (5): 798–799. doi : 10.1097/j.pain.0000000000000133 . ПМИД 25887392 . S2CID 2060896 .
- ^ Дэнбери Т.К., Уикс Калифорния, Чемберс Дж.П., Уотерман-Пирсон А.Э., Кестин С.С. (март 2000 г.). «Самовыбор обезболивающего препарата карпрофен хромыми цыплятами-бройлерами». Ветеринарный журнал . 146 (11): 307–11. дои : 10.1136/vr.146.11.307 . ПМИД 10766114 . S2CID 35062797 .
- ^ Машин К.Л. (2005). «Птичья анальгезия». Семинары по медицине птиц и экзотических домашних животных . 14 (4): 236–242. дои : 10.1053/j.saep.2005.09.004 .
- ^ Пол-Мерфи Дж., Хокинс М.Г. (2014). «Глава 26 – Особенности птиц: распознавание боли у домашних птиц». В Gaynor JS, Muir III WW (ред.). Справочник по ветеринарному лечению боли . Elsevier Науки о здоровье. ISBN 978-0-323-08935-7 .
- ^ Мосли, Калифорния (2005). «Анестезия и анальгезия у рептилий». Семинары по медицине птиц и экзотических домашних животных . 14 (4): 243–262. дои : 10.1053/j.saep.2005.09.005 .
- ^ Мосли С. (январь 2011 г.). «Боль и ноцицепция у рептилий». Ветеринарные клиники Северной Америки. Практика с экзотическими животными . 14 (1): 45–60. дои : 10.1016/j.cvex.2010.09.009 . ПМИД 21074702 .
- ^ Сладкий К.К., Манс С (2012). «Клиническая аналгезия у рептилий». Журнал медицины экзотических домашних животных . 21 (2): 158–167. дои : 10.1053/j.jepm.2012.02.012 .
- ^ Мачин К.Л. (март 1999 г.). «Земноводная боль и анальгезия». Журнал медицины зоопарков и дикой природы . 30 (1): 2–10. JSTOR 20095815 . ПМИД 10367638 .
- ^ Мачин К.Л. (январь 2001 г.). «Обезболивание для рыб, амфибий и рептилий». Ветеринарные клиники Северной Америки. Практика с экзотическими животными . 4 (1): 19–33. дои : 10.1016/S1094-9194(17)30048-8 . ПМИД 11217460 .
- ^ Стивенс CW (январь 2011 г.). «Аналгезия у амфибий: доклинические исследования и клиническое применение» . Ветеринарные клиники Северной Америки. Практика с экзотическими животными . 14 (1): 33–44. дои : 10.1016/j.cvex.2010.09.007 . ПМК 3056481 . ПМИД 21074701 .
- ^ Перейти обратно: а б с Варнер Дж.Э. (2012). «Глава 5: Какие животные разумны?». Личность, этика и познание животных: положение животных в двухуровневом утилитаризме Хэйра . Издательство Оксфордского университета. doi : 10.1093/acprof:oso/9780199758784.001.0001 . ISBN 978-0-19-975878-4 . Таблица в статье составлена на основе таблицы 5.2, стр. 113.
- ^ Генетт С.А., Жиру MC, Вашон П. (2013). «Восприятие боли и анестезия у исследовательских лягушек» . Экспериментальные животные . 62 (2): 87–92. дои : 10.1538/expanim.62.87 . ПМИД 23615302 .
- ^ Мосли С. (2006). «Боль, ноцицепция и анальгезия у рептилий: когда змея кричит: «ой!» " (PDF) . Североамериканская ветеринарная конференция . 20 : 1652–1653.
- ^ Кобл DJ, Тейлор Д.К., Мук Д.М. (май 2011 г.). «Обезболивающее действие мелоксикама, сульфата морфина, флуниксина меглюмина и гидрохлорида ксилазина у африканских когтистых лягушек (Xenopus laevis)» . Журнал Американской ассоциации наук о лабораторных животных . 50 (3): 355–60. ПМК 3103286 . ПМИД 21640031 .
- ^ Бейкер Б.Б., Сладкий К.К., Джонсон С.М. (январь 2011 г.). «Оценка обезболивающего эффекта перорального и подкожного введения трамадола у красноухих черепах» . Журнал Американской ветеринарной медицинской ассоциации . 238 (2): 220–7. дои : 10.2460/javma.238.2.220 . ПМК 3158493 . ПМИД 21235376 .
- ^ Эндрюс К. (2014). «Раздел 3.6.2: Боль от рыбы» . Животный разум: введение в философию познания животных . Рутледж. ISBN 978-1-317-67675-1 .
- ^ «Какой самый гуманный способ убивать ракообразных для потребления человеком?» . База знаний RSPCA . Королевское общество по предотвращению жестокого обращения с животными (RSPCA), Австралия . Проверено 20 сентября 2020 г.
- ^ Перейти обратно: а б Снеддон Л.У. (октябрь 2004 г.). «Эволюция ноцицепции у позвоночных: сравнительный анализ низших позвоночных» . Исследования мозга. Обзоры исследований мозга . 46 (2): 123–30. дои : 10.1016/j.brainresrev.2004.07.007 . ПМИД 15464201 . S2CID 16056461 .
- ^ Перейти обратно: а б Крук Р.Дж., Диксон К., Хэнлон Р.Т., Уолтерс Э.Т. (май 2014 г.). «Ноцицептивная сенсибилизация снижает риск нападения хищников» . Современная биология . 24 (10): 1121–5. Бибкод : 2014CBio...24.1121C . дои : 10.1016/j.cub.2014.03.043 . ПМИД 24814149 .
- ^ Перейти обратно: а б с Снеддон Л.У., Элвуд Р.В., Адамо С.А., Лич MC (2014). «Определение и оценка боли животных» . Поведение животных . 97 : 201–212. дои : 10.1016/j.anbehav.2014.09.007 . S2CID 53194458 .
- ^ Перейти обратно: а б Элвуд Р.В., Барр С., Паттерсон Л. (2009). «Боль и стресс у ракообразных?». Прикладная наука о поведении животных . 118 (3): 128–136. дои : 10.1016/j.applanim.2009.02.018 .
- ^ Прайс ТиДжей, Дюссор Дж. (май 2014 г.). «Эволюция: преимущество «неадаптивной» болевой пластичности» . Современная биология . 24 (10): Р384-6. Бибкод : 2014CBio...24.R384P . дои : 10.1016/j.cub.2014.04.011 . ПМК 4295114 . ПМИД 24845663 .
- ^ «Дезадаптивная боль» . Оксфордский справочник . Проверено 16 мая 2016 г.
- ^ Сафина С (2016). «Рыбья боль: Больная тема» . Чувство животных . 1 (3): 41. дои : 10.51291/2377-7478.1076 .
- ^ Перейти обратно: а б Хантингфорд Ф.А., Адамс С., Брейтуэйт В.А., Кадри С., Поттинджер Т.Г., Сандё П., Тернбулл Дж.Ф. (2006). «Обзорный документ: Текущие проблемы благополучия рыб» (PDF) . Журнал биологии рыб . 68 (2): 332–372. дои : 10.1111/j.0022-1112.2006.001046.x .
- ^ Перейти обратно: а б Снеддон Л.У., Брейтуэйт, Вирджиния, Джентл М.Дж. (июнь 2003 г.). «Есть ли у рыб ноцицепторы? Доказательства эволюции сенсорной системы позвоночных» . Слушания. Биологические науки . 270 (1520): 1115–21. дои : 10.1098/rspb.2003.2349 . ПМК 1691351 . ПМИД 12816648 .
- ^ Снеддон Л. (12 апреля 2021 г.). «Существует множество доказательств того, что рыбы чувствуют боль | Письмо» . Хранитель . Проверено 28 августа 2021 г.
- ^ Хамамото Д.Т., Форки М.В., Дэвис В.Л., Каяндер К.К., Симоне Д.А. (апрель 2000 г.). «Роль pH и осмолярности в возникновении реакции вытирания, вызванной уксусной кислотой, в модели ноцицепции у лягушек». Исследования мозга . 862 (1–2): 217–29. дои : 10.1016/S0006-8993(00)02138-7 . ПМИД 10799688 . S2CID 7290178 .
- ^ Ларивьер В.Р., Мелзак Р. (август 1996 г.). «Тест на пчелиный яд: новый тонизирующий болевой тест» . Боль . 66 (2–3): 271–7. дои : 10.1016/0304-3959(96)03075-8 . ПМИД 8880850 . S2CID 34628083 .
- ^ Роверони РК, Парада Калифорния, Сесилия М, Вейга ФА, Тамбели СН (ноябрь 2001 г.). «Разработка поведенческой модели боли в ВНЧС у крыс: формалиновый тест ВНЧС». Боль . 94 (2): 185–191. дои : 10.1016/S0304-3959(01)00357-8 . ПМИД 11690732 . S2CID 15199427 .
- ^ Перейти обратно: а б с Нежный MJ (ноябрь 1992 г.). «Боль у птиц» . Благополучие животных . 1 (4): 235–47. дои : 10.1017/S0962728600015189 . S2CID 255884717 .
- ^ Стивенс CW (1992). «Альтернативы использованию млекопитающих для исследования боли». Науки о жизни . 50 (13): 901–12. дои : 10.1016/0024-3205(92)90167-Н . ПМИД 1548975 .
- ^ Хандверкер Х.О., Антон Ф., Ри П.В. (1 февраля 1987 г.). «Схемы разрядки афферентных кожных нервных волокон из хвоста крысы во время длительной вредной механической стимуляции». Экспериментальное исследование мозга . 65 (3): 493–504. дои : 10.1007/BF00235972 . ПМИД 3556477 . S2CID 22840458 .
- ^ Снеддон LU (май 2003 г.). «Тройничная соматосенсорная иннервация головы костистых рыб с особым упором на ноцицепцию» . Исследования мозга . 972 (1–2): 44–52. дои : 10.1016/s0006-8993(03)02483-1 . ПМИД 12711077 . S2CID 14616224 .
- ^ Эшли П.Дж., Снеддон Л.У., Маккрохан С.Р. (август 2007 г.). «Ноцицепция у рыб: стимульно-реакционные свойства рецепторов на голове форели Oncorhynchus mykiss» . Исследования мозга . 1166 : 47–54. дои : 10.1016/j.brainres.2007.07.011 . ПМИД 17673186 . S2CID 15837167 .
- ^ Меттам Дж.Дж., Маккрохан С.Р., Снеддон Л.У. (февраль 2012 г.). «Характеристика хемосенсорных рецепторов тройничного нерва радужной форели Oncorhynchus mykiss: реакция на химические раздражители и углекислый газ» . Журнал экспериментальной биологии . 215 (Часть 4): 685–93. дои : 10.1242/jeb.060350 . ПМИД 22279076 .
- ^ Червова Л.С., Лапшин Д.Н. (2004). «Болевая чувствительность рыб и аналгезия, вызываемая опиоидными и неопиоидными агентами» (PDF) . В материалах четвертой международной конференции Ирана и России .
- ^ Эшли П.Дж., Снеддон Л.У., Маккрохан С.Р. (декабрь 2006 г.). «Свойства рецепторов роговицы костистых рыб» . Письма по неврологии . 410 (3): 165–8. дои : 10.1016/j.neulet.2006.08.047 . ПМИД 17101221 . S2CID 14375428 .
- ^ Перейти обратно: а б с д и ж Роуз Дж.Д., Арлингхаус Р., Кук С.Дж., Дигглс Б.К., Савинок В., Стивенс Э.Д., Винн К.Д. (2012). «Могут ли рыбы действительно чувствовать боль?» (PDF) . Рыба и рыболовство . 15 (1): 97–133. дои : 10.1111/faf.12010 .
- ^ Снеддон LU (февраль 2002 г.). «Анатомический и электрофизиологический анализ тройничного нерва костистой рыбы Oncorhynchus mykiss» . Письма по неврологии . 319 (3): 167–71. дои : 10.1016/S0304-3940(01)02584-8 . ПМИД 11834319 . S2CID 14807046 .
- ^ Сноу П.Дж., Плендерлейт М.Б., Райт Л.Л. (август 1993 г.). «Количественное исследование популяций первичных сенсорных нейронов трех видов пластиножаберных рыб». Журнал сравнительной неврологии . 334 (1): 97–103. дои : 10.1002/cne.903340108 . ПМИД 8408762 . S2CID 32762031 .
- ^ Брейтуэйт, В.А., Булкотт П. (май 2007 г.). «Восприятие боли, отвращение и страх у рыб» . Болезни водных организмов . 75 (2): 131–8. дои : 10.3354/dao075131 . ПМИД 17578252 .
- ^ Перейти обратно: а б с Вебер Э.С. (январь 2011 г.). «Рыбная анальгезия: боль, стресс, неприятие страха или ноцицепция?». Ветеринарные клиники Северной Америки. Практика с экзотическими животными . 14 (1): 21–32. дои : 10.1016/j.cvex.2010.09.002 . ПМИД 21074700 .
- ^ Нордгрин Дж., Хорсберг Т.Э., Ранхейм Б., Чен А.С. (декабрь 2007 г.). «Соматосенсорные вызванные потенциалы в конечном мозге атлантического лосося (Salmo salar) после гальванической стимуляции хвоста». Журнал сравнительной физиологии А. 193 (12): 1235–42. дои : 10.1007/s00359-007-0283-1 . ПМИД 17987296 . S2CID 19654379 .
- ^ Перейти обратно: а б с Снеддон Л.У. (2009). «Восприятие боли у рыб: показатели и конечные точки» . Журнал ИЛАР . 50 (4): 338–42. дои : 10.1093/ilar.50.4.338 . ПМИД 19949250 .
- ^ Данлоп Р., Лэминг П. (сентябрь 2005 г.). «Механорецепторные и ноцицептивные реакции в центральной нервной системе золотой рыбки (Carassius auratus) и форели (Oncorhynchus mykiss)» . Журнал боли . 6 (9): 561–8. дои : 10.1016/j.jpain.2005.02.010 . ПМИД 16139775 .
- ^ Перейти обратно: а б Рейли С.К., Куинн Дж.П., Коссинс А.Р., Снеддон Л.У. (май 2008 г.). «Новые гены-кандидаты идентифицированы в мозге во время ноцицепции у карпа (Cyprinus carpio) и радужной форели (Oncorhynchus mykiss)» . Письма по неврологии . 437 (2): 135–8. дои : 10.1016/j.neulet.2008.03.075 . ПМИД 18440145 . S2CID 18763423 .
- ^ Познер Л.П. (2009). «Введение: боль и страдания рыб: обзор доказательств» . Журнал ИЛАР . 50 (4): 327–8. дои : 10.1093/ilar.50.4.327 . ПМИД 19949248 .
- ^ Людвигсен С., Стенклев Н.К., Джонсен Х.К., Лаукли Е., Матре Д., Аас-Хансен О. (февраль 2014 г.). «Вызванные потенциалы у атлантической трески после предположительно безобидной и предположительно вредной электростимуляции: минимально инвазивный подход» . Физиология и биохимия рыб . 40 (1): 173–81. Бибкод : 2014FPBio..40..173L . дои : 10.1007/s10695-013-9834-2 . ПМК 3901938 . ПМИД 23896862 .
- ^ Перейти обратно: а б с д Рибейро М.М., Пинто А., Пинто М., Херас М., Мартинс И., Коррейя А. и др. (июль 2011 г.). «Подавление ноцицептивных реакций после системного введения амидированного киоторфина» . Британский журнал фармакологии . 163 (5): 964–73. дои : 10.1111/j.1476-5381.2011.01290.x . ПМК 3130928 . ПМИД 21366550 .
- ^ Перейти обратно: а б Браун С. (январь 2015 г.). «Рыбный интеллект, чувствительность и этика» . Познание животных . 18 (1): 1–17. дои : 10.1007/s10071-014-0761-0 . ПМИД 24942105 . S2CID 207050888 .
- ^ Снеддон Л.У., Лич MC (2016). «Антропоморфное отрицание рыбьей боли» . Чувство животных . 1 (3): 28. дои : 10.51291/2377-7478.1048 .
- ^ Снеддон Л.У. (2011). «Свидетельства о восприятии боли у рыбы и последствия употребления рыбы» . Журнал исследований сознания . 18 (9): 209–229.
- ^ Буатти М.К., Пастернак Г.В. (август 1981 г.). «Множественные опиатные рецепторы: филогенетические различия». Исследования мозга . 218 (1–2): 400–5. дои : 10.1016/0006-8993(81)91319-6 . ПМИД 6268247 . S2CID 6870252 .
- ^ Маррон Фдез де Веласко Э., Лоу П.Ю., Родригес Р.Е. (сентябрь 2009 г.). «Мю-опиоидный рецептор рыбок данио проявляет функциональные характеристики, такие же, как у мю-опиоидных рецепторов млекопитающих». Рыбка данио . 6 (3): 259–68. дои : 10.1089/zeb.2009.0594 . ПМИД 19761379 .
- ^ Дреборг С., Сундстрем Г., Ларссон Т.А., Лархаммар Д. (октябрь 2008 г.). «Эволюция опиоидных рецепторов позвоночных» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (40): 15487–92. Бибкод : 2008PNAS..10515487D . дои : 10.1073/pnas.0805590105 . ПМК 2563095 . ПМИД 18832151 .
- ^ Виньуэла-Фернандес I, Джонс Э., Уэльсский EM, Флитвуд-Уокер С.М. (сентябрь 2007 г.). «Механизмы боли и их значение для лечения боли у сельскохозяйственных животных и домашних животных». Ветеринарный журнал . 174 (2): 227–39. дои : 10.1016/j.tvjl.2007.02.002 . ПМИД 17553712 .
- ^ Снеддон Л.У. (2012). «Клиническая анестезия и анальгезия у рыб» . Журнал медицины экзотических домашних животных . 21 : 32–43. дои : 10.1053/j.jepm.2011.11.009 . S2CID 78473 .
- ^ Перейти обратно: а б Джонс С.Г., Камунде С., Лемке К., Стивенс Э.Д. (декабрь 2012 г.). «Зависимость антиноцицептивного эффекта морфина от дозы у рыбы, радужной форели». Журнал ветеринарной фармакологии и терапии . 35 (6): 563–70. дои : 10.1111/j.1365-2885.2011.01363.x . ПМИД 22229842 .
- ^ Перейти обратно: а б Снеддон Л.У. (2003). «Доказательства боли у рыб: использование морфия в качестве анальгетика» . Прикладная наука о поведении животных . 83 (2): 153–162. дои : 10.1016/s0168-1591(03)00113-8 .
- ^ Нордгрин Дж., Джозеф П., Гарнер Дж.П., Янчак А.М., Ранхейм Б., Мьюир В.М., Хорсберг Т.Э. (2009). «Термоноцицепция у рыб: влияние двух разных доз морфия на температурный порог и послетестовое поведение у золотых рыбок (Carassius auratus)». Прикладная наука о поведении животных . 119 (1–2): 101–107. дои : 10.1016/j.applanim.2009.03.015 .
- ^ Перейти обратно: а б Веник ДМ (май 2007 г.). «Когнитивные способности и разум: каких водных животных следует защищать?» . Болезни водных организмов . 75 (2): 99–108. дои : 10.3354/dao075099 . ПМИД 17578249 .
- ^ Меттам Дж.Дж., Оултон Л.Дж., Маккрохан С.Р., Снеддон Л.У. (2011). «Эффективность трех типов обезболивающих препаратов в уменьшении боли у радужной форели Oncorhynchus mykiss» . Прикладная наука о поведении животных . 133 (3): 265–274. дои : 10.1016/j.applanim.2011.06.009 .
- ^ Лопес-Луна Дж., Аль-Джубури К., Аль-Нуайми В., Снеддон Л.У. (апрель 2017 г.). «Снижение активности из-за вредной химической стимуляции улучшается при погружении рыбок данио в обезболивающие препараты» . Журнал экспериментальной биологии . 220 (Часть 8): 1451–1458. дои : 10.1242/jeb.146969 . ПМИД 28424313 . S2CID 24635067 .
- ^ Перейти обратно: а б с Волкерс КП, Барбоза Джуниор А, Менескаль-де-Оливейра Л, Хоффманн А (2013). «Вызванная стрессом антиноцицепция у рыб устраняется налоксоном» . ПЛОС ОДИН . 8 (7): е71175. Бибкод : 2013PLoSO...871175W . дои : 10.1371/journal.pone.0071175 . ПМК 3728202 . ПМИД 23936261 .
- ^ Перейти обратно: а б Стинберген П.Дж., Бардин Н. (2014). «Антиноцицептивные эффекты бупренорфина на личинок рыбок данио: альтернатива моделям на грызунах для изучения боли и ноцицепции?». Прикладная наука о поведении животных . 152 : 92–99. дои : 10.1016/j.applanim.2013.12.001 .
- ^ Эренсинг Р.Х., Мичелл Г.Ф., Кастин А.Дж. (октябрь 1982 г.). «Аналогичный антагонизм морфиновой анальгезии со стороны MIF-1 и налоксона у Carassius auratus». Фармакология, биохимия и поведение . 17 (4): 757–61. дои : 10.1016/0091-3057(82)90358-6 . ПМИД 6129644 . S2CID 31113845 .
- ^ Червова Л.С., Лапшин Д.Н. (2000). «Опиоидная модуляция болевого порога у рыб». Доклады биологических наук . 375 (1): 590–1. дои : 10.1023/а:1026681519613 . ПМИД 11211504 . S2CID 1180288 .
- ^ Перейти обратно: а б с Экрот-младший, Аас-Хансен О, Снеддон Л.У., Бичао Х., Довинг К.Б. (2014). «Физиологические и поведенческие реакции атлантической трески (Gadus morhua) на вредные раздражители» . ПЛОС ОДИН . 9 (6): е100150. Бибкод : 2014PLoSO...9j0150E . дои : 10.1371/journal.pone.0100150 . ПМК 4061104 . ПМИД 24936652 .
- ^ Грандин Т. (2015). «Глава 2 – Важность измерений для улучшения благосостояния скота, птицы и рыбы». В Грандин Т. (ред.). Улучшение благополучия животных: практический подход . ISBN 978-1-78064-468-4 .
- ^ Рейли С.К., Куинн Дж.П., Коссинс А.Р., Снеддон Л.У. (2008). «Поведенческий анализ ноцицептивных явлений у рыб: сравнение трех видов демонстрирует специфические реакции» . Прикладная наука о поведении животных . 114 (1): 248–259. дои : 10.1016/j.applanim.2008.01.016 .
- ^ Рокес Х.А., Аббинк В., Гердс Ф., ван де Вис Х., Флик Г. (ноябрь 2010 г.). «Отрезание хвостового плавника, болезненная процедура: исследования нильской тилапии и карпа». Физиология и поведение . 101 (4): 533–40. дои : 10.1016/j.physbeh.2010.08.001 . ПМИД 20705079 . S2CID 25859917 .
- ^ Дунайер Дж. (июль – август 1991 г.). «Рыба: чувствительность за пределами хватки похитителя» . Программа животных : 12–18 – через All-Creatures.org.
- ^ «Познание животных» . 22 апреля 2015 года . Проверено 15 сентября 2015 г.
- ^ Хугланд Р., Моррис Д., Тинберген Н. (1956). «Иглы колюшек (Gasterosteus и Pygosteus) как средство защиты от хищников (Perca и Esox)» . Поведение . 10 (3/4): 205–236. дои : 10.1163/156853956X00156 . ISSN 0005-7959 . JSTOR 4532857 .
- ^ Данлоп Р., Миллсопп С., Лэминг П. (2006). «Обучение избеганию у золотых рыбок (Carassius auratus) и форели (Oncorhynchus mykiss) и последствия для восприятия боли». Прикладная наука о поведении животных . 97 (2): 255–271. дои : 10.1016/j.applanim.2005.06.018 .
- ^ Перейти обратно: а б Миллсопп С., Лэминг П. (2007). «Компромисс между кормлением и предотвращением шока у золотых рыбок (Carassius auratus)». Прикладная наука о поведении животных . 113 (1–3): 247–254. дои : 10.1016/j.applanim.2007.11.004 .
- ^ Зерболио DJ, Royalty JL (1 сентября 1983 г.). «Соответствие и условное различение странностей у золотой рыбки как реакция избегания: доказательства концептуального обучения избеганию» . Обучение и поведение животных . 11 (3): 341–348. дои : 10.3758/BF03199786 . ISSN 1532-5830 .
- ^ Перейти обратно: а б Снеддон Л.У., Брейтуэйт, Вирджиния, Джентл М.Дж. (октябрь 2003 г.). «Новый объектный тест: исследование ноцицепции и страха у радужной форели» . Журнал боли . 4 (8): 431–40. дои : 10.1067/S1526-5900(03)00717-X . ПМИД 14622663 .
- ^ Эшли П.Дж., Рингроуз С., Эдвардс К.Л., Уоллингтон Е., Э., Маккрохан С.Р., Снеддон Л.У. (2009). «Влияние вредной стимуляции на реакцию борьбы с хищниками и статус доминирования у радужной форели» . Поведение животных . 77 (2): 403–410. дои : 10.1016/j.anbehav.2008.10.015 . S2CID 19225428 .
- ^ Веник ДМ (2001). «Эволюция боли» (PDF) . Фламандский ветеринарный журнал . 70 (1): 17–21. дои : 10.21825/vdt.89895 . S2CID 38767856 . Архивировано из оригинала (PDF) 30 декабря 2022 года . Проверено 25 сентября 2015 г.
- ^ Перейти обратно: а б Браун С. (2016). «Рыбья боль: неудобная правда» . Чувство животных . 1 (3): 32. дои : 10.51291/2377-7478.1069 .
- ^ Кук С.Дж., Снеддон Л.У. (2007). «Перспективы защиты животных при любительском рыболовстве». Прикладная наука о поведении животных . 104 (3–4): 176–198. CiteSeerX 10.1.1.630.459 . дои : 10.1016/j.applanim.2006.09.002 . S2CID 49233569 .
- ^ Лик Дж. (14 марта 2004 г.). «Рыболовы столкнутся с проверкой RSPCA» . Санди Таймс . Архивировано из оригинала 23 сентября 2015 года . Проверено 15 сентября 2015 г.
- ^ Дигглс Б.К. (2016). «Рыбная боль: изменит ли это текущую лучшую практику в реальном мире?» . Чувство животных . 1 (3): 35. дои : 10.51291/2377-7478.1068 .
- ^ Снеддон Л.У. (2006). «Этика и благополучие: восприятие боли рыбами» . Бык. Евро. доц. Рыба. Патол . 26 (1): 6–10.
- ^ Перейти обратно: а б Броуман Х.И., Кук С.Дж., Коукс И.Г., Дербишир С.В., Касумян А., Ки Б. и др. (2018). «Благосостояние водных животных: где что находится, куда они идут и что это значит для исследований, аквакультуры, любительского рыболовства и коммерческого рыболовства» . Журнал морских наук ICES . 76 (1): 82–92. doi : 10.1093/icesjms/fsy067 . ISSN 1054-3139 .
- ^ «Закон о животных (научные процедуры) 1986 года» (PDF) . Министерство внутренних дел (Великобритания) . Проверено 23 сентября 2015 г.
- ^ «Закон о защите животных 2006 г.» . Правительство Великобритании. 2006 год . Проверено 25 сентября 2015 г.
- ^ «Раздел 7 USC – Сельское хозяйство» .
- ^ «Животные в исследованиях» . неавс. Архивировано из оригинала 18 сентября 2015 года . Проверено 25 сентября 2015 г.
- ^ Хенриксен С., Ваагланд Х., Сундт-Хансен Л., Мэй Р., Фьелльхайм А. (2003). «Последствия восприятия боли у рыб при вылове и выпуске, аквакультуре и коммерческом рыболовстве» (PDF) .
- ^ Перейти обратно: а б с д и Роуз Дж.Д., Арлингхаус Р., Кук С.Дж., Дигглс Б.К., Савинок В., Стивенс Э.Д., Винн К.Д. (2012). «Могут ли рыбы действительно чувствовать боль?» (PDF) . Рыба и рыболовство . 15 (1): 97–133. дои : 10.1111/faf.12010 .
- ^ Мэтьюз, Гэри; Викельгрен, Уоррен О. (1978). «Тройничные сенсорные нейроны морской миноги» . Журнал сравнительной физиологии А. 123 (4): 329–333. дои : 10.1007/bf00656966 . ISSN 0340-7594 . S2CID 1631932 .
- ^ Снеддон LU (апрель 2015 г.). «Боль у водных животных» . Журнал экспериментальной биологии . 218 (Часть 7): 967–76. дои : 10.1242/jeb.088823 . ПМИД 25833131 .
- ^ Браун, Кулум (1 января 2017 г.). «Оценка риска и филогенетический подход» . Чувство животных . 2 (16). дои : 10.51291/2377-7478.1219 . ISSN 2377-7478 .
- ^ Брум, Дональд (1 января 2016 г.). «Рыбий мозг и поведение указывают на способность чувствовать боль» . Чувство животных . 1 (3). дои : 10.51291/2377-7478.1031 . ISSN 2377-7478 .
- ^ Роуз Джей Ди (2002). «Нейроповеденческая природа рыб и вопрос осознания и боли» (PDF) . Обзоры в журнале Fisheries Science . 10 (1): 1–38. Бибкод : 2002RvFS...10....1R . CiteSeerX 10.1.1.598.8119 . дои : 10.1080/20026491051668 . S2CID 16220451 . Архивировано из оригинала (PDF) 10 октября 2012 года.
- ^ Роуз Джей Ди (2002). «Рыбы чувствуют боль?» . Архивировано из оригинала 20 января 2013 года . Проверено 27 сентября 2007 г.
- ^ «Рыбы чувствуют боль, говорят ученые» . Новости Би-би-си . 30 апреля 2003 года . Проверено 20 мая 2010 г.
- ^ Перейти обратно: а б Грандин Т., Джонсон С. (2005). Животные в переводе . Нью-Йорк: Скрибнер. стр. 183–184 . ISBN 978-0-7432-4769-6 .
- ^ Роуз Джей Ди (2003). Эриксон Х.Э. (ред.). «Критика статьи: «Есть ли у рыб ноцицепторы: доказательства эволюции сенсорной системы позвоночных» » (PDF) . Информационные ресурсы по благополучию рыб 1970–2003 гг., Информационные ресурсы по благополучию животных . Белтсвилл, Мэриленд: Министерство сельского хозяйства США. стр. 49–51. Архивировано из оригинала (PDF) 19 ноября 2008 года.
- ^ Перейти обратно: а б Ключ Б (2015). «Рыбы не чувствуют боли и ее значения для понимания феноменального сознания» . Биология и философия . 30 (2): 149–165. дои : 10.1007/s10539-014-9469-4 . ПМЦ 4356734 . ПМИД 25798021 .
- ^ Ключ Б (2016). «Почему рыбы не чувствуют боли» . Чувство животных . 1 (3): 1. дои : 10.51291/2377-7478.1011 .
- ^ «Рыбам не хватает мозга, чтобы чувствовать боль», — утверждает новейшая школа мысли . Телеграф . 10 февраля 2003 г.
- ^ Роуз Джей Ди (май 2007 г.). «Антропоморфизм и «психическое благополучие» рыб» . Болезни водных организмов . 75 (2): 139–54. дои : 10.3354/dao075139 . ПМИД 17578253 .
- ^ Перейти обратно: а б Снеддон, Линн; Лопес-Луна, Хавьер; Вулфенден, Дэвид; Лич, Мэтью; Валентим, Ана; Стинберген, Питер; Бардин, Набила; Карри, Аманда; Брум, Дональд; Браун, Кулум (1 января 2018 г.). «Отрицание разума рыб: мутение воды» . Чувство животных . 3 (21). дои : 10.51291/2377-7478.1317 . ISSN 2377-7478 . S2CID 55070090 .
 Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Текст был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Снеддон LU (28 августа 2012 г.). «Восприятие боли у рыб: почему критики не могут принять научные доказательства боли у рыб» (PDF) . Архивировано из оригинала (PDF) 23 сентября 2015 года.
- ^ Бекофф М., Шерман П.В. (апрель 2004 г.). «Размышления о животных» . Тенденции в экологии и эволюции . 19 (4): 176–80. дои : 10.1016/j.tree.2003.12.010 . ПМИД 16701251 . S2CID 17877550 .
- ^ Дербишир SW (2016). «Рыбам не хватает мозгов и психологии, чтобы чувствовать боль» . Чувство животных . 1 (3): 18. дои : 10.51291/2377-7478.1047 .
- ^ Вудрафф М. (2017). «Сознание костистых рыб: есть что-то вроде ощущения себя рыбой» . Чувство животных . 2 (13): 1. дои : 10.51291/2377-7478.1198 . S2CID 56306955 .
- ^ Вудрафф М (2018). «Чувствительность рыб: больше о доказательствах». Чувство животных . 3 (13): 16.
- ^ Фейнберг Т.Е., Маллат Дж.М. (2016). Древние истоки сознания: как мозг создавал опыт . Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-03433-3 .
- ^ Ньюби, Северная Каролина, Стивенс Э.Д. (2008). «Влияние «болевого» теста с уксусной кислотой на кормление, плавание и дыхательные реакции радужной форели (Oncorhynchus mykiss)». Прикладная наука о поведении животных . 114 (1): 260–269. дои : 10.1016/j.applanim.2007.12.006 .
- ^ Снеддон Л.У. (2009). «Влияние «болевого» теста с уксусной кислотой на кормление, плавание и дыхательные реакции радужной форели (Oncorhynchus mykiss): критика Ньюби и Стивенса (2008)» . Прикладная наука о поведении животных . 116 (1): 96–97. дои : 10.1016/j.applanim.2008.07.006 .
- ^ Ньюби, Северная Каролина, Стивенс Э.Д. (2009). «Влияние «болевого» теста с уксусной кислотой на кормление, плавание и дыхательные реакции радужной форели (Oncorhynchus mykiss): критика Ньюби и Стивенса (2008) - ответ». Прикладная наука о поведении животных . 116 (1): 97–99. дои : 10.1016/j.applanim.2008.07.009 .
- ^ Стоскопф, МК (1 февраля 1994 г.). «Боль и анальгезия у птиц, рептилий, земноводных и рыб» . Исследовательская офтальмология и визуальные науки . 35 (2): 775–780. ISSN 1552-5783 . ПМИД 8113029 .
- ^ Вагнер, Гленн Н.; Стивенс, Э. Дон (1 мая 2000 г.). «Влияние различных хирургических техник: шовный материал и расположение места разреза на поведение радужной форели (Oncorhynchus mykiss)» . Морское и пресноводное поведение и физиология . 33 (2): 103–114. Бибкод : 2000MFBP...33..103W . дои : 10.1080/10236240009387084 . ISSN 1023-6244 . S2CID 83911821 .
- ^ Перейти обратно: а б «Рыбы на самом деле могут чувствовать боль и реагировать на нее так же, как люди» . Университет Пердью. 29 апреля 2009 г. Архивировано из оригинала 5 декабря 2022 г. . Проверено 13 сентября 2009 г.
- ^ Сальвичек, Люси Х.; Прето, Лоран; Демарта, Ланила; Проктор, Дарби; Эсслер, Дженнифер; Пинто, Ана И.; Висмер, Шэрон; Стоинский, Тара; Броснан, Сара Ф.; Бшари, Редуан (21 ноября 2012 г.). «Взрослый губан-чистильщик превосходит обезьян-капуцинов, шимпанзе и орангутангов в сложной задаче по поиску пищи, полученной в результате сотрудничества чистильщик-клиент рифовой рыбы» . ПЛОС ОДИН . 7 (11): e49068. Бибкод : 2012PLoSO...749068S . дои : 10.1371/journal.pone.0049068 . ISSN 1932-6203 . ПМК 3504063 . ПМИД 23185293 .
- ^ Пепперберг, Ирен М.; Хартсфилд, Ли Энн (2014). «Могут ли серые попугаи (Psittacus erithacus) преуспеть в выполнении «сложной» задачи по поиску пищи, которую не смогли выполнить приматы (Pan troglodytes, Pongo abelii, Sapajus apella), но которую удалось решить рыбе-губану (Labroides dimidiatus)?» . Журнал сравнительной психологии . 128 (3): 298–306. дои : 10.1037/a0036205 . ISSN 1939-2087 . ПМИД 24798239 .
Внешние ссылки
[ редактировать ]- «Это официально: рыбы чувствуют боль» . Смитсоновский журнал . 8 января 2018 года . Проверено 31 октября 2021 г.
- «Ошибаемся ли мы, полагая, что рыбы не чувствуют боли?» . Хранитель . 30 октября 2018 года . Проверено 31 октября 2021 г.
- «Ученые говорят, что рыбы чувствуют боль. Это может привести к серьезным изменениям в рыбной отрасли» . Вашингтон Пост . 8 января 2018 г.
- «Наука показывает, что рыбы чувствуют боль» . Марк Бекофф . Хаффингтон Пост . 29 декабря 2015 г.
- «Чувствуют ли рыбы боль? Дебаты продолжаются» . Марк Лалланилья . ЖиваяНаука . 2 июля 2013 г.
