Иглокожие
| Иглокожие | |
|---|---|
 | |
| Современные и вымершие иглокожие шести классов: Fromia indica ( Asteroidea ); Офиокома сколопендрина ( Ophiuroidea ); Stomopneustes variolaris ( Echinoidea ); Оксикомантус Беннетти ( Crinoidea ); Actinopyga echinites ( Holothuroidea ); Ктеноцистоидея . | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Subkingdom: | Эуметазоа |
| Клэйд : | ПараХоксозоа |
| Клэйд : | билатерия |
| Клэйд : | Нефрозоа |
| Супертип: | Дейтеростомия |
| Клэйд : | Амбулаторный |
| Тип: | Иглокожие Брюгьер, 1791 г. [бывший Кляйн , 1734 г.] |
| Типовой род | |
| Эхинус Линней, 1758 г. | |
| Подтипы и классы [2] | |
Homalozoa † Гилл и Кастер, 1960 г.
†=Вымершие | |
Иглокожие / ( / ɪ ˈ k aɪ n ə ˌ d ɜːr m , ˈ ɛ k ə - ) [3] любое вторичноротое животное типа Echinodermata ( , / ɪ ˌ k aɪ n oʊ ˈ d ɜːr m ə t ə / ), к которому относятся морские звезды , хрупкие звезды морские морские ежи , песчаные доллары и морские огурцы , а также сидячие лилии или «каменные лилии». [4] Хотя двусторонне симметричны личинки (пентамерная симметрия) и , взрослые иглокожие узнаваемы по своей обычно пятилучевой радиальной симметрии встречаются на морском дне на любой глубине океана от приливной зоны до абиссальной зоны . Этот тип насчитывает около 7600 живых видов , что делает его второй по величине группой вторичноротых после хордовых , а также крупнейшим типом , обитающим только в морских условиях . Первые дефинитивные иглокожие появились в начале кембрия .
Иглокожие важны как с экологической, так и с геологической точки зрения. есть несколько других группировок, столь обильных С экологической точки зрения, в биотической пустыне глубокого моря , а также в более мелких океанах . Большинство иглокожих способны размножаться бесполым путем и регенерировать ткани, органы и конечности; в некоторых случаях они могут подвергнуться полной регенерации из одной конечности. С геологической точки зрения ценность иглокожих заключается в их окостеневших дермальных эндоскелетах , которые вносят основной вклад во многие известняковые образования и могут дать ценную информацию о геологической среде. Они были наиболее часто используемыми видами в регенеративных исследованиях в 19 и 20 веках. Кроме того, некоторые ученые считают, что радиация иглокожих была ответственна за мезозойскую морскую революцию .
Таксономия и эволюция
[ редактировать ]Название иглокожие происходит от древнегреческого ἐχῖνος ( ekhînos ) «еж» и δέρμα ( dérma ) «кожа». [5] Иглокожие — билатерии , то есть их предки были зеркально-симметричными. Среди билатерий они принадлежат к отделу вторичноротых , а это означает, что бластопор , первое отверстие, образующееся во время развития эмбриона, становится анусом , а не ртом. [6] [7] Характерными чертами взрослых иглокожих являются наличие водной сосудистой системы с внешними трубчатыми ножками и известкового эндоскелета, состоящего из косточек, соединенных сеткой коллагеновых волокон. [8]
Филогения
[ редактировать ]Исторически таксономисты полагали, что Ophiuroidea были сестрой Asteroidea как клады Asterozoa или что они были сестрой (Holothuroidea + Echinoidea) как клады Cryptosyringida. [9] Однако анализ 219 генов всех классов иглокожих, проведенный в 2014 году, привел к убедительной поддержке Asterozoa. [10] Независимый анализ транскриптомов РНК 23 видов всех классов иглокожих, проведенный в 2015 году, дал одно и то же дерево. [9]
- Внешняя филогения
Контекст иглокожих внутри Bilateria: [11]
| билатерия |
| ||||||||||||||||||||||||

- Внутренняя филогения
| Эхино‑ | |
| дерматология |



Разнообразие
[ редактировать ]Существует около 7600 современных видов иглокожих, а также около 13 000 вымерших видов. [8] [12] Все иглокожие — морские животные , но их среда обитания варьируется от мелководных приливных зон до абиссальных глубин. Традиционно выделяются два основных подразделения: более знакомые подвижные Eleutherozoa , включающие Asteroidea ( морские звезды , около 1745 видов), Ophiuroidea ( хрупкие звезды , около 2300 видов), Echinoidea ( морские ежи и песчаные доллары , около 900 видов). и Holothuroidea ( морские огурцы , около 1430 видов); и Pelmatozoa , некоторые из которых сидячие , а другие подвижные. К ним относятся криноидеи ( перьевые звезды и морские лилии , насчитывающие около 580 видов) и вымершие бластоиды и паракриноиды . [13] [14]
- Подтип иглокожих
 Хрупкая звезда Ophionereis reticulata.
Хрупкая звезда Ophionereis reticulata. Stichopus Морской огурец chromnotus из Малайзии.
Stichopus Морской огурец chromnotus из Малайзии. Морские звезды разных цветов
Морские звезды разных цветов Морской еж Strongylocentrotus purpuratus.
Морской еж Strongylocentrotus purpuratus.





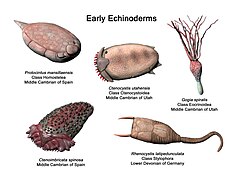
Ископаемая история
[ редактировать ]Самым старым кандидатом ископаемого иглокожего является Аркаруа из докембрия Австралии . Эти окаменелости имеют форму диска, с радиальными гребнями по краю и пятиконечной центральной впадиной, отмеченной радиальными линиями. Однако окаменелости не имеют стереомы или внутренней структуры, указывающей на наличие водной сосудистой системы, поэтому их невозможно окончательно идентифицировать. [15]
Первые общепризнанные иглокожие появляются в нижнем кембрии ; астерозои появились в ордовике , а криноидеи были доминирующей группой в палеозое . Иглокожие оставили после себя обширную летопись окаменелостей. [16] Предполагается, что предком всех иглокожих было простое, подвижное, двусторонне-симметричное животное, обладавшее ртом, кишкой и анальным отверстием. Этот предковый организм принял прикрепленный образ жизни с питанием взвесью и развил радиальную симметрию. Несмотря на это, личинки всех иглокожих двусторонне симметричны, и при метаморфозе у всех развивается радиальная симметрия. Как и их предок, морские звезды и криноидеи все еще прикрепляются к морскому дну, принимая взрослую форму. [17]
Первые иглокожие были неподвижны. [17] но превратились в животных, способных свободно передвигаться. Вскоре у них развились эндоскелетные пластинки со стереомной структурой и наружные ресничные бороздки для питания. [18] Палеозойские иглокожие имели шаровидную форму, прикреплены к субстрату и были ориентированы ротовой поверхностью вверх. У этих ранних иглокожих были амбулакральные бороздки, идущие вниз по бокам тела, окаймленные с обеих сторон брахиолами, как перышки современных криноидей. В конце концов, за исключением криноидей, все классы иглокожих изменили свою ориентацию и стали ртом вниз. До того, как это произошло, подии, вероятно, выполняли функцию питания, как и у современных криноидей. Локомоторная функция подиумов появилась позже, когда в результате переориентации рта подиумы впервые соприкоснулись с субстратом. [17]
- Дополнительная информация: Дибрахицистис.
 Ордовикские цистоидные эхиносфаэриты северо-востока Эстонии.
Ордовикские цистоидные эхиносфаэриты северо-востока Эстонии. Ископаемые криноидей коронки
Ископаемые криноидей коронки




Онлайн-база данных модельных организмов
[ редактировать ]Echinobase — это база данных модельных организмов для ряда иглокожих. В настоящее время полностью поддерживаются 4 вида иглокожих (страницы генов, BLAST, треки JBrowse, загрузки генома), включая Strongylocentrotus purpuratus (фиолетовый морской еж), Lytechinus variegatus (зеленый морской еж), Patiria miniata (звезда летучей мыши) и Acanthaster planci (корона- шипованная морская звезда). Частично поддерживаемые виды (страницы генов отсутствуют) включают Lytechinus pictus (окрашенный морской еж), Asterias Rubens (сахарная звезда) и Anneissia japonica (криноидей перьевой звезды). [19] [20]
Анатомия и физиология
[ редактировать ]Иглокожие произошли от животных с двусторонней симметрией . Хотя взрослые иглокожие обладают пентарадиальной симметрией, их личинки представляют собой реснитчатые , свободно плавающие организмы с двусторонней симметрией. Позже, в ходе метаморфоза, левая сторона тела растет за счет правой, которая со временем рассасывается. Затем левая сторона растет по пентарадиально- симметричному принципу, при котором тело состоит из пяти частей вокруг центральной оси. [21] Внутри Asterozoa могут быть некоторые исключения из правил. У большинства морских звезд рода Leptasterias шесть рук, хотя могут встречаться и пятирукие особи. Brisingida также содержит несколько шестируких видов. Среди хрупких звезд существуют шестирукие виды, такие как Ophiothela danae , Ophiactis savigni и Ophionotus hexactis , а Ophiacantha vivipara часто имеет более шести. [22]
На каком-то этапе жизни иглокожие имеют вторичную радиальную симметрию в частях тела, что, скорее всего, является адаптацией к сидячему или малоподвижному существованию. [23] Многие криноидеи и некоторые морские звезды симметричны, кратные основным пяти; Морские звезды, такие как Labidiaster annulatus , имеют до пятидесяти конечностей, а морская лилия Comaster schlegelii - двести. [24]
Генетические исследования показали, что гены, управляющие развитием туловищной области, в процессе развития отсутствуют (единственным известным исключением является ген, активный на кончиках рук морских звезд), что указывает на то, что их тело состоит только из развитой головной области. [25] [26]
Кожа и скелет
[ редактировать ]Иглокожие имеют мезодермальный скелет в дерме, состоящий из пластинок на основе кальцита, известных как косточки . Если они твердые, они образуют тяжелый скелет, поэтому имеют пористую структуру, похожую на губку, известную как стереома. [27] [28] Слуховые косточки могут сливаться вместе, как у морских ежей, или могут сочленяться, образуя гибкие суставы, как у морских звезд, хрупких звезд и криноидей. Косточки могут иметь внешние выступы в виде шипов, гранул или бородавок и поддерживаются прочным эпидермисом . Скелетные элементы иногда располагаются особым образом, например, жевательный орган, называемый « фонарем Аристотеля » у морских ежей, опорные стебли криноидей и структурное «известковое кольцо» морских огурцов. [21]
Хотя отдельные косточки прочные и легко окаменевают, полные скелеты морских звезд, хрупких звезд и криноидей в летописи окаменелостей редки. С другой стороны, морские ежи часто хорошо сохраняются в меловых пластах или известняках. Во время окаменения полости в стереоме заполняются кальцитом, который сливается с окружающей породой. Разламывая такую породу, палеонтологи могут наблюдать характерные узоры спайности, а иногда даже сложную внутреннюю и внешнюю структуру панциря. [29]
Эпидермис содержит пигментные клетки, которые часто обеспечивают яркие цвета иглокожих, включая темно-красный, черно-белые полосы и насыщенный фиолетовый. [30] Эти клетки могут быть светочувствительными, из-за чего многие иглокожие полностью меняют внешний вид с наступлением ночи. Реакция может произойти быстро: морской еж Centrostephanus longispinus меняет цвет всего за пятьдесят минут под воздействием света. [31]
Одной из характеристик большинства иглокожих является особый вид ткани, известный как соединительная ткань . Этот материал на основе коллагена может изменять свои механические свойства под нервным контролем, а не с помощью мышц. Эта ткань позволяет морской звезде перейти от гибкого перемещения по морскому дну к тому, чтобы стать твердой, открывая двустворчатый моллюск или предотвращая свое извлечение из расщелины. Точно так же морские ежи могут фиксировать свои обычно подвижные шипы в вертикальном положении в качестве защитного механизма при нападении. [32] [33]
Водная сосудистая система
[ редактировать ]
Иглокожие обладают уникальной водной сосудистой системой, сетью заполненных жидкостью каналов, видоизмененных из целома (полости тела), которые участвуют в газообмене, питании, сенсорной рецепции и передвижении. Эта система варьируется у разных классов иглокожих, но обычно открывается наружу через ситовидный мадрепорит на аборальной (верхней) поверхности животного. Мадрепорит связан с тонким протоком, каменным каналом, который доходит до кольцевого канала, окружающего рот или пищевод . Кольцевой канал разветвляется на множество радиальных каналов, которые у астероидов тянутся вдоль рукавов, а у ежовых примыкают к раковине в амбулакральных участках. От радиальных каналов отходят короткие латеральные каналы, каждый из которых заканчивается ампулой. Часть ампулы может выступать через пору (или пару пор у морских ежей) наружу, образуя подиум или ножку трубки . Водная сосудистая система помогает распределять питательные вещества по телу животного; это наиболее заметно в ножках трубки, которые могут расширяться или сжиматься за счет перераспределения жидкости между ножкой и внутренней ампулой. [34] [35]
Несколько иная организация водной сосудистой системы у офиуроидов: мадрепорит может располагаться на поверхности рта, а у подиумов присоски отсутствуют. [36] У голотуроидов система редуцирована, часто с небольшим количеством трубчатых ножек, за исключением специализированных питающихся щупалец, и мадрепорит открывается в целом. У некоторых голотуроидов, таких как Apodida, отсутствуют трубчатые ножки и каналы вдоль тела; у других есть продольные каналы. [37] Устройство криноидей аналогично устройству астероидов, но у трубчатых ножек нет присосок, и они совершают возвратно-поступательные движения для передачи частиц пищи, захваченных руками, к центральному рту. На астероидах такое же движение используется для перемещения животного по земле. [38]
Другие органы
[ редактировать ]Иглокожие обладают простой пищеварительной системой, которая варьируется в зависимости от рациона животного. Морские звезды в основном плотоядны и имеют рот, пищевод, желудок, состоящий из двух частей, кишечник и прямую кишку, причем анус расположен в центре аборальной поверхности тела. За некоторыми исключениями, представители отряда Paxillosida не имеют заднего прохода. [39] [40] У многих видов морских звезд большой сердечный желудок может быть вывернут для переваривания пищи вне тела. Некоторые другие виды способны поедать цельные продукты питания, например, моллюски . [41] У хрупких звезд, питающихся по-разному, есть слепая кишка без кишечника и ануса; они выбрасывают пищевые отходы через рот. [42] Морские ежи — травоядные животные и используют свой специализированный ротовой аппарат, чтобы щипать, разрывать и пережевывать пищу, в основном водоросли . У них есть пищевод, большой желудок и прямая кишка с анусом на вершине яичка. [43] Морские огурцы в основном являются детритофагами , сортирующими осадок с помощью модифицированных трубчатых ножек вокруг рта — щечных щупалец. Песок и грязь сопровождают пищу через простой кишечник, имеющий длинный извитой кишечник и большую клоаку . [44] Морские лилии — это подвесные питатели , пассивно ловящие планктон , который попадает в их вытянутые руки. Комки пищи, содержащей слизь, передаются в рот, который соединен с анусом петлей, состоящей из короткого пищевода и более длинной кишки. [45]
Целомические полости иглокожих имеют сложную структуру. Помимо водной сосудистой системы, иглокожие имеют гемальный целом , перивисцеральный целом , гонадный целом и часто также окологемальный целом. [46] В процессе развития целом иглокожих делится на метацель, мезоцель и протоцель (также называемый соматоцелем, гидроцелем и аксоцелем соответственно). [47] Водная сосудистая система, гемальная система и перигемальная система образуют трубчатую целомическую систему. [48] Иглокожие необычны тем, что имеют как целомическую систему кровообращения (водную сосудистую систему), так и гемальную систему кровообращения, поскольку у большинства групп животных есть только одна из двух. [49]
Гемальная и перигемальная системы происходят из исходного целома, образуя открытую и редуцированную систему кровообращения. Обычно он состоит из центрального кольца и пяти радиальных сосудов. нет Настоящего сердца , а в крови часто отсутствует дыхательный пигмент. Газообмен происходит через кожные жабры или папулы у морских звезд, генитальные сумки у хрупких звезд, околоротовые жабры у морских ежей и клоакальные деревья у голотурий. Обмен газов также происходит через ножки трубки. У иглокожих отсутствуют специализированные органы выделения (удаления отходов), поэтому азотистые отходы , главным образом в виде аммиака , диффундируют через дыхательные поверхности. [34]
Целомическая жидкость содержит целомоциты или иммунные клетки. Существует несколько типов иммунных клеток, которые различаются в зависимости от класса и вида. Все классы обладают типом фагоцитирующих амебоцитов, которые поглощают вторгающиеся частицы и инфицированные клетки, объединяются или сгущаются и могут участвовать в цитотоксичности . Эти клетки обычно большие и зернистые и считаются основной линией защиты от потенциальных патогенов. [50] В зависимости от класса иглокожие могут иметь сферальные клетки (для цитотоксичности, воспаления и антибактериальной активности), вибрирующие клетки (для движения целомической жидкости и свертывания крови) и кристаллические клетки (которые могут служить для осморегуляции у голотурий). [50] [51] Целомоциты секретируют антимикробные пептиды против бактерий и содержат набор лектинов и белков комплемента как часть врожденной иммунной системы , характеристика которой еще не изучена. [52]
Иглокожие имеют простую радиальную нервную систему , состоящую из модифицированной нервной сети взаимосвязанных нейронов без центрального мозга , хотя у некоторых из них имеются ганглии . Нервы расходятся от центральных колец вокруг рта в каждую руку или вдоль стенки тела; ветви этих нервов координируют движения организма и синхронизацию ножек трубки. У морских звезд есть сенсорные клетки в эпителии, простые глазные пятна и чувствительные к прикосновению ножки-трубочки, похожие на щупальца, на кончиках рук. У морских ежей нет особых органов чувств, но есть статоцисты , которые помогают в гравитационной ориентации, и у них тоже есть сенсорные клетки в эпидермисе, особенно в трубчатых ножках, шипах и педицелляриях . Хрупкие звезды, криноидеи и трепанги вообще не имеют органов чувств, но у некоторых роющих голотурий отряда Apodida имеется по одной статоцисте, примыкающей к каждому лучевому нерву, а у некоторых имеется глазное пятно у основания каждого щупальца. [53]
Гонады . по крайней мере периодически занимают большую часть полостей тела морских ежей [54] и морские огурцы, тогда как менее объемистые криноидеи, хрупкие звезды и морские звезды имеют по две гонады в каждом плече. Хотя считается, что у предков современных иглокожих было одно половое отверстие, у многих организмов есть несколько гонопоров , через которые могут высвобождаться яйцеклетки или сперматозоиды. [53]
Регенерация
[ редактировать ]
Многие иглокожие обладают огромной способностью к регенерации . Многие виды регулярно подвергаются автотомии и регенерации рук и внутренних органов . Морские огурцы часто выделяют части своих внутренних органов, если чувствуют угрозу, восстанавливая их в течение нескольких месяцев. Морские ежи постоянно заменяют утраченные в результате повреждений иглы, а морские звезды и морские лилии легко теряют и регенерируют свои конечности. В большинстве случаев из одной отрезанной руки не может вырасти новая морская звезда из-за отсутствия хотя бы части диска. [55] [56] [57] [58] Однако у некоторых видов одна рука может выжить и развиться в полноценную особь, а руки иногда намеренно отделяют с целью бесполого размножения . [56] [57] [58] В периоды потери пищеварительного тракта морские огурцы живут за счет накопленных питательных веществ и поглощают растворенные органические вещества непосредственно из воды. [59]
Регенерация утраченных частей включает как эпиморфоз , так и морфаллаксис . При эпиморфозе стволовые клетки — либо из резервного пула, либо полученные в результате дедифференцировки — образуют бластему и генерируют новые ткани. Морфаллактическая регенерация включает в себя перемещение и ремоделирование существующих тканей для замены утраченных частей. [60] прямая трансдифференцировка одного типа ткани в другой при замещении ткани. Наблюдается также [61]
Воспроизведение
[ редактировать ]Половое размножение
[ редактировать ]Иглокожие становятся половозрелыми примерно через два-три года, в зависимости от вида и условий окружающей среды. Почти все виды имеют раздельный мужской и женский пол , хотя некоторые из них гермафродиты . Яйцеклетки и сперматозоиды обычно выпускаются в открытую воду, где происходит оплодотворение. Выпуск сперматозоидов и яйцеклеток у некоторых видов синхронизирован, обычно в соответствии с лунным циклом. У других видов особи могут объединяться во время репродуктивного сезона, что увеличивает вероятность успешного оплодотворения. Внутреннее оплодотворение наблюдалось у трех видов морской звезды, трех хрупких звезд и глубоководного трепанга. Даже на бездонных глубинах , куда не проникает свет, иглокожие часто синхронизируют свою репродуктивную деятельность. [62]
Некоторые иглокожие высиживают яйца . Это особенно характерно для видов, живущих в холодной воде, где планктонные личинки не могут найти достаточно пищи. Эти оставшиеся яйца обычно немногочисленны и содержат крупные желтки для питания развивающихся эмбрионов. У морских звезд самка может носить икру в специальных мешочках, под мышками, под выгнутым телом или даже в сердечном желудке. [63] Многие хрупкие звезды — гермафродиты; они часто высиживают яйца, обычно в специальных камерах на поверхности рта, но иногда в яичнике или целоме. [64] У этих морских звезд и хрупких звезд развитие обычно идет непосредственно к взрослой форме, не проходя через двустороннюю личиночную стадию. [65] Несколько морских ежей и один вид песчаного доллара откладывают икру в полостях или возле ануса, удерживая их на месте своими шипами. [66] Некоторые морские огурцы используют свои щечные щупальца для переноса яиц на нижнюю или заднюю часть тела, где они и сохраняются. У очень небольшого числа видов яйца сохраняются в целоме, где они развиваются живородяще , а затем выходят наружу через разрывы стенки тела. [67] У некоторых криноидей эмбрионы развиваются в специальных сумках для размножения, где яйца хранятся до тех пор, пока их не найдет сперма, выпущенная самцом. [68]
Бесполое размножение
[ редактировать ]
Один из видов морской звезды , Ophidiaster granifer , размножается бесполым путем партеногенеза . [69] У некоторых других астерозойных взрослые особи размножаются бесполым путем, пока не созреют, а затем размножаются половым путем. У большинства этих видов бесполое размножение осуществляется путем поперечного деления с разделением диска на две части. И потерянная область диска, и отсутствующие руки отрастают заново, поэтому у человека могут быть руки разной длины. [58] [70] В период отрастания у них есть несколько крошечных и одно большое плечо, поэтому их часто называют «кометами». [57] [71]
Взрослые трепанги размножаются бесполым путем поперечным делением. Голотурия парвула часто использует этот метод, разделяясь на две части немного впереди средней точки. Каждая из двух половинок регенерирует недостающие органы в течение нескольких месяцев, но недостающие половые органы часто развиваются очень медленно. [72]
Личинки некоторых иглокожих способны к бесполому размножению. Давно известно, что это происходит у морских звезд и хрупких звезд, но совсем недавно оно наблюдалось у трепанга, песчаного доллара и морского ежа. [73] Это может происходить путем автотомизации частей, которые развиваются во вторичные личинки, путем почкования или поперечного деления . Автотомированные части или почки могут развиваться непосредственно в полностью сформированные личинки или могут пройти стадию гаструлы или даже бластулы . Новые личинки могут развиваться из преорального капюшона (бугоркообразного образования над ртом), боковой стенки тела, задне-боковых рук или их задних концов. [73] [74] [75]
Клонирование обходится личинке дорого как по ресурсам, так и по времени разработки. Личинки подвергаются этому процессу, когда еды много. [76] или температурные условия оптимальны. [75] Клонирование может происходить с целью использования тканей, которые обычно теряются во время метаморфоза. [77] Личинки некоторых песчанок клонируют себя, когда обнаруживают растворенную рыбью слизь, что указывает на присутствие хищников. [75] [77] Бесполое размножение дает множество личинок меньшего размера, которые лучше убегают от планктоноядных рыб, что означает, что этот механизм может быть адаптацией против хищников. [78]
Развитие личинок
[ редактировать ]
Развитие начинается с двусторонне-симметричного эмбриона, сначала развивается целобластула. Гаструляция отмечает открытие «второго рта», в результате которого иглокожие помещаются внутри вторичноротых, а мезодерма, в которой будет находиться скелет, мигрирует внутрь. Вторичная полость тела, целом, образуется в результате разделения трех полостей тела. Личинки часто планктонны , но у некоторых видов яйца сохраняются внутри самки, а у некоторых самка высиживает личинки. [79] [80]
Личинки проходят несколько стадий, которые имеют особые названия, полученные от таксономических названий взрослых особей или от их внешнего вида. Например, у морского ежа есть личинка «эхиноплютеус», а у хрупкой звезды — личинка «офиоплутеус». У морской звезды имеется личинка « бипиннария », которая развивается в многорукую личинку « брахиолярию ». Личинка трепанга — это «аурикулярия», а личинка криноидея — «желточник». Все эти личинки двусторонне симметричны и имеют полоски ресничек, с помощью которых они плавают; у некоторых, обычно известных как личинки «плютеуса», есть руки. Когда они полностью развились, они оседают на морском дне, где претерпевают метаморфозы, а личиночные конечности и кишечник дегенерируют. Левая сторона личинки развивается в ротовую поверхность молоди, а правая становится аборальной поверхностью. На этом этапе развивается пентарадиальная симметрия. [79] [81]
личинки , Предковым личиночным типом иглокожих считается личинка, питающаяся планктоном, живущая и питающаяся в толще воды, но у современных иглокожих около 68% видов развиваются с использованием питающейся желтком . [13] Наличие желточного мешка означает, что производится меньшее количество яиц, у личинок более короткий период развития и меньший потенциал распространения, но больше шансов на выживание. [13]
Распространение и среда обитания
[ редактировать ]Иглокожие распространены по всему миру практически на всех глубинах, широтах и средах океана. Взрослые особи в основном ведут донный образ жизни , обитая на морском дне, тогда как личинки часто ведут пелагический образ жизни и живут в виде планктона в открытом океане. Однако некоторые взрослые особи голотуроидов, такие как Pelagothuria, ведут пелагический образ жизни. [82] Некоторые криноидеи являются псевдопланктонными и прикрепляются к плавающим бревнам и мусору, хотя наиболее широко такое поведение наблюдалось в палеозое, до того как конкуренция со стороны таких организмов, как ракушки, ограничила масштабы такого поведения. [83]
Образ жизни
[ редактировать ]Передвижение
[ редактировать ]
Иглокожие в основном используют свои трубчатые ножки для передвижения, хотя некоторые морские ежи также используют свои шипы. Ножки трубки обычно имеют кончик в форме присоски, в которой за счет сокращения мышц может создаваться вакуум. Это в сочетании с некоторой липкостью из-за выделения слизи обеспечивает адгезию. Ножки трубки сокращаются и расслабляются волнами, которые движутся по прилипшей поверхности, и животное медленно движется вперед. [84]
Хрупкие звезды — самые ловкие из иглокожих. Любая из рук может образовывать ось симметрии, направленную вперед или назад. Затем животное движется скоординировано, приводя его в движение четырьмя другими руками. Во время передвижения метательные руки могут совершать змееподобные или гребные движения. [85] Морские звезды передвигаются с помощью своих трубчатых ног, сохраняя руки почти неподвижными, в том числе у таких видов, как Pycnopodia , у которых руки гибкие. Поверхность рта покрыта тысячами трубчатых ножек, которые движутся вразнобой друг с другом, но не в метахронном ритме ; Однако в некотором смысле ножки трубки скоординированы, поскольку животное устойчиво скользит вперед. [86] У некоторых роющих морских звезд на трубчатых ножках вместо присосок имеются точки, и они способны «скользить» по морскому дну с большей скоростью. [87]
Морские ежи используют свои трубчатые ножки для передвижения так же, как морские звезды. Некоторые также используют свои шарнирные шипы, чтобы толкать себя вперед или отрывать поверхность рта от субстрата. Если морской еж перевернут, он может вытянуть свои трубчатые ножки в одной амбулакральной области достаточно далеко, чтобы они оказались в пределах досягаемости от субстрата, а затем последовательно прикреплять лапы из прилегающей области, пока не выпрямится. Некоторые виды проникают в скалу, обычно шлифовая поверхность ротовым аппаратом. [88]

Морские огурцы, как правило, медлительные животные. Многие из них могут передвигаться по поверхности морского дна или рыться в песке или иле, используя перистальтические движения; у некоторых на нижней поверхности есть короткие трубчатые ножки, с помощью которых они могут ползти, как морские звезды. Некоторые виды передвигаются с помощью щечных щупалец, а другие умудряются плавать с помощью перистальтических движений или ритмичных изгибов. Многие живут в трещинах, дуплах и норах и почти не передвигаются. Некоторые глубоководные виды являются пелагическими и могут плавать в воде с перепончатыми сосочками, образующими паруса или плавники. [89]
Большинство криноидей подвижны, но морские лилии сидячие и прикрепляются к твердым субстратам стеблями. Движение у большинства морских лилий ограничивается сгибанием (их стебли могут сгибаться), а также перекатыванием и развертыванием рук; некоторые виды могут перемещаться по морскому дну ползком. Морские перья не прикреплены и обычно живут в расщелинах, под кораллами или внутри губок, причем единственной видимой частью являются их рукава. Некоторые морские перья вылетают ночью и садятся на близлежащие возвышенности, чтобы лучше использовать течения, несущие пищу. Многие виды могут «ходить» по морскому дну, поднимая тело с помощью рук, или плавать, используя руки. Однако большинство видов морских перьев в основном ведут оседлый образ жизни и редко удаляются далеко от выбранного места укрытия. [90]
Кормление
[ редактировать ]Способы питания у разных таксонов иглокожих сильно различаются. Морские лилии и некоторые хрупкие звезды, как правило, являются пассивными фильтраторами. [91] [92] улавливание взвешенных частиц из проходящей воды. Большинство морских ежей являются травоядными; [93] морские огурцы — питатели отложений; [94] и большинство морских звезд — активные охотники. [95]
Морские лилии улавливают частицы пищи с помощью ножек трубки на своих распростертых перышках, перемещают их в амбулакральные бороздки, обволакивают слизью и доставляют в рот с помощью ресничек, выстилающих бороздки. [91] Точные диетические потребности криноидей мало изучены, но в лаборатории их можно кормить диатомовыми водорослями. [96]
Звезды-корзины — подвесные кормушки, поднимающие свои разветвленные конечности для сбора зоопланктона , в то время как другие хрупкие звезды используют несколько способов питания. Некоторые из них представляют собой подвесные кормушки, закрепляющие частицы пищи с помощью нитей слизи, шипов или трубчатых ножек на поднятых руках. Другие — падальщики и питатели детрита. Другие также являются прожорливыми хищниками и способны заарканить свою водную добычу, внезапно окружив ее гибкими руками. Затем конечности сгибаются под диском, чтобы передать пищу в челюсти и рот. [92]
Многие морские ежи питаются водорослями, часто соскребая тонкий слой водорослей, покрывающий поверхность камней, своим специальным ротовым аппаратом, известным как фонарь Аристотеля. Другие виды пожирают более мелкие организмы, которых они могут поймать своими трубчатыми ножками. Они также могут питаться мертвой рыбой и другими животными. [93] Песчаные доллары могут осуществлять взвешенное питание и питаться фитопланктоном , детритом, кусочками водорослей и бактериальным слоем, окружающим песчинки. [97]
Морские огурцы часто являются мобильными кормушками для отложений или суспензий, используя свои щечные подиумы для активного захвата пищи, а затем помещая частицы по отдельности в свои щечные полости. Другие заглатывают большое количество осадка, поглощают органические вещества и пропускают неперевариваемые минеральные частицы через кишечник. Таким образом они разрушают и перерабатывают большие объемы субстрата, часто оставляя на морском дне характерные гряды отложений. Некоторые морские огурцы инфаунально живут в норах передним концом вниз и анусом на поверхности, заглатывая осадок и пропуская его через кишечник. Другие норы живут передним концом вверх и ждут, пока детрит упадет во входы в норы, или сгребают мусор с близлежащей поверхности своими щечными подиями. [94]
Почти все морские звезды питаются детритом или хищниками, хотя некоторые из них питаются взвесью. Мелкая рыба, приземлившаяся на верхнюю поверхность, может быть поймана педицилиями, а мертвые животные могут быть убраны, но основной добычей являются живые беспозвоночные, в основном двустворчатые моллюски. Чтобы питаться одним из них, морская звезда перемещается по нему, прикрепляет трубчатые ножки и оказывает давление на клапаны, выгибая спину. Когда образуется небольшая щель между створками, морская звезда вставляет в добычу часть желудка, выделяет пищеварительные ферменты и медленно разжижает мягкие части тела. Когда приводящая мышца двустворчатого моллюска расслабляется, в него вставляется больше желудка, и когда пищеварение завершено, желудок возвращается в свое обычное положение у морской звезды с теперь уже разжиженной пищей двустворчатых моллюсков внутри него. Другие морские звезды выворачивают желудок, чтобы питаться губками, морскими анемонами, кораллами, детритом и пленками водорослей. [95]
Защита от хищников
[ редактировать ]
Несмотря на низкую пищевую ценность и обилие неперевариваемого кальцита, иглокожие являются пищей многих организмов, в том числе костистых рыб , акул , гаг , чаек , крабов , брюхоногих моллюсков , других иглокожих, каланов , песцов и человека. Более крупные морские звезды охотятся на более мелких; большое количество яиц и личинок, которые они производят, составляют часть зоопланктона , потребляемого многими морскими существами. С другой стороны, криноидеи относительно свободны от хищников. [98]
Защита от хищников включает наличие шипов, токсинов (врожденных или доставленных через ножки трубки), а также выделение липких опутывающих нитей морскими огурцами. Хотя большинство шипов иглокожих тупые, у морских звезд с терновым венцом они длинные и острые и могут вызвать болезненные колотые раны, поскольку покрывающий их эпителий содержит токсин. [99] Из-за захватывающей соединительной ткани, которая может быстро переходить из вялого состояния в жесткое, иглокожих очень трудно вытеснить из расщелин. У некоторых морских огурцов есть скопление кювьеровых трубочек , которые могут выбрасываться из ануса в виде длинных липких нитей, чтобы запутать и навсегда вывести из строя нападавшего. Морские огурцы иногда защищаются, разрывая стенки тела и освобождая кишечник и внутренние органы. [100] Морские звезды и хрупкие звезды при нападении могут подвергнуться автотомии , в результате чего оторвется рука; это может отвлечь хищника на время, достаточное для того, чтобы животное могло убежать. Некоторые виды морских звезд умеют уплывать от опасности. [101]
Экология
[ редактировать ]
Иглокожие — многочисленные беспозвоночные, взрослые особи которых играют важную роль в донных экосистемах , а личинки — основной компонент планктона. Среди экологических ролей взрослых особей - выпас морских ежей, переработка отложений сердечных ежей, а также взвешивание и кормление отложениями криноидей и морских огурцов. [13] [82] Некоторые морские ежи могут вгрызаться в твердую породу, дестабилизируя ее поверхность и выбрасывая питательные вещества в океан. Коралловые рифы также пробуриваются таким способом, но скорость нарастания карбонатного материала часто превышает скорость эрозии, вызываемой морским ежом. [102] Иглокожие улавливают около 0,1 гигатонны углекислого газа в год в виде карбоната кальция , что делает их важными участниками глобального углеродного цикла . [103]
У иглокожих иногда наблюдаются большие колебания численности популяции, которые могут трансформировать экосистемы. Например, в 1983 году массовая гибель тропического морского ежа Diadema antillarum в Карибском море привела к изменению системы рифов с преобладанием кораллов на систему с преобладанием водорослей. [104] Морские ежи являются одними из основных травоядных животных на рифах, и обычно существует прекрасный баланс между ежами, водорослями и другими водорослями, на которых они пасутся. Уменьшение численности хищников (выдр, омаров и рыб) может привести к увеличению численности ежей, что приведет к чрезмерному выпасу лесов из водорослей , в результате чего оголенные водорослями «бесплодные ежи» станут « бесплодными ежами ». [105] На Большом Барьерном рифе необъяснимое увеличение численности морских звезд с терновым венцом ( Acanthaster planci ), которые питаются живой тканью кораллов, значительно увеличило смертность кораллов и сократило биоразнообразие коралловых рифов . [106]
Использование людьми
[ редактировать ]Как еда и лекарство
[ редактировать ]

В 2019 году заготовлено 129 052 тонны иглокожих. Большую часть из них составили морские огурцы (59 262 тонны) и морские ежи (66 341 тонна). [107] Они используются в основном в пищу, но также и в традиционной китайской медицине . [108] В некоторых странах Юго-Восточной Азии трепанги считаются деликатесом; как таковые, они находятся в непосредственной опасности чрезмерного сбора урожая. [109] Популярные виды включают ананасовую катушку Thelenota ananas ( сусухан ) и красный морской огурец Holothuria edulis . Эти и другие виды в просторечии известны как bêche de mer или трепанг в Китае и Индонезии . Морские огурцы варят двадцать минут, а затем сушат естественным путем, а затем на огне, что придает им дымный привкус. В Китае их используют как основу для студенистых супов и рагу. [110] В пищу употребляются как мужские, так и женские гонады морских ежей, особенно в Японии и Франции . Вкус описывается как мягкий и тающий, напоминающий смесь морепродуктов и фруктов. [111] [112] Были предприняты эксперименты по разведению морских ежей, чтобы попытаться компенсировать чрезмерную эксплуатацию . [113]
В исследованиях
[ редактировать ]Из-за быстрого роста личинок морские ежи широко используются в исследованиях, особенно в качестве модельных организмов в биологии развития и экотоксикологии. [114] [115] [116] [117] Strongylocentrotus purpuratus и Arbacia punctulata . С этой целью в эмбриологических исследованиях используются [118] Большой размер и прозрачность яйцеклеток позволяют наблюдать за сперматозоидами в процессе оплодотворения яйцеклеток . [114] Потенциал регенерации рук хрупких звезд изучается в связи с пониманием и лечением нейродегенеративных заболеваний у людей. [119] Геномные данные, относящиеся к модельным организмам иглокожих, собраны в Echinobase . [19] [20]
Другое использование
[ редактировать ]Известковые раковины или панцири иглокожих используются в качестве источника извести фермерами в районах, где известняк недоступен, а некоторые из них используются при производстве рыбной муки . [120] Ежегодно для этих целей используется четыре тысячи тонн животных. Эта торговля часто осуществляется совместно с фермерами, выращивающими моллюсков , для которых морские звезды представляют серьезную угрозу, поедая их выращенные запасы. Другие виды использования морской звезды, которую они добывают, включают производство корма для животных, компостирование и подготовку сушеных образцов для декоративно-прикладного искусства. [119]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Смит, А., Замора, С. и Альваро, Дж. Самые старые фауны иглокожих из Гондваны показывают, что разнообразие плана тела иглокожих было быстрым. Nat Commun 4, 1385 (2013). https://doi.org/10.1038/ncomms2391 .
- ^ Штер, Сабина (2014). «Иглокожие» . ВОРМС . Всемирный регистр морских видов . Проверено 23 февраля 2014 г.
- ^ «иглокожие» . Dictionary.com Полный (онлайн). nd
- ^ Холл, Даниэль (3 февраля 2022 г.). «Иглокожие | Смитсоновский океан» . Ocean.si.edu . Проверено 31 марта 2023 г.
- ^ «иглокожие» . Интернет-словарь этимологии .
- ^ Дорит, Уокер и Барнс 1991 , стр. 777–779.
- ^ Фокс, Ричард. « Астериас форбеси » . Анатомия беспозвоночных онлайн . Ландерский университет . Проверено 19 мая 2012 г.
- ^ Jump up to: Перейти обратно: а б Рэй, Грегори А. (1999). «Иглокожие: животные с колючей кожей: морские ежи, морские звезды и их союзники» . Древо Жизни . Архивировано из оригинала 23 мая 2015 года . Проверено 19 октября 2012 г.
- ^ Jump up to: Перейти обратно: а б Эскрива, Гектор; Райх, Адриан; Данн, Кейси; Акасака, Кодзи; Вессель, Гэри (2015). «Филогеномный анализ иглокожих подтверждает наличие сестринских групп астерозоа и эхинозоа» . ПЛОС ОДИН . 10 (3): e0119627. Бибкод : 2015PLoSO..1019627R . дои : 10.1371/journal.pone.0119627 . ПМЦ 4368666 . ПМИД 25794146 .
- ^ Телфорд, MJ; Лоу, CJ; Кэмерон, CB; Ортега-Мартинес, О.; Аронович, Дж.; Оливери, П.; Копли, Р.Р. (2014). «Филогеномный анализ отношений классов иглокожих поддерживает Asterozoa» . Труды Королевского общества B: Биологические науки . 281 (1786): 20140479. doi : 10.1098/rspb.2014.0479 . ПМК 4046411 . ПМИД 24850925 .
- ^ Эджкомб, Грегори Д.; Гирибет, Гонсало; Данн, Кейси В.; Хейнол, Андреас; Кристенсен, Рейнхардт М.; Невес, Рикардо К.; Роуз, Грег В.; Ворсаае, Катрин; Соренсен, Мартин В. (июнь 2011 г.). «Отношения многоклеточных животных более высокого уровня: недавний прогресс и оставшиеся вопросы» . Организмы, разнообразие и эволюция . 11 (2): 151–172. дои : 10.1007/s13127-011-0044-4 . S2CID 32169826 .
- ^ «Сеть разнообразия животных - Иглокожие» . Зоологический музей Мичиганского университета . Проверено 26 августа 2012 г.
- ^ Jump up to: Перейти обратно: а б с д Утик, Свен; Шаффельке, Бритта; Бирн, Мария (1 января 2009 г.). «Тип бума-спада? Экологические и эволюционные последствия изменений плотности у иглокожих» . Экологические монографии . 79 (1): 3–24. Бибкод : 2009ЭкоМ...79....3У . дои : 10.1890/07-2136.1 .
- ^ Арноне, Мария Ина; Бирн, Мария; Мартинес, Педро (2015). «Иглокожие». Эволюционная биология развития беспозвоночных 6 . Вена: Шпрингер Вена. стр. 1–58. дои : 10.1007/978-3-7091-1856-6_1 . ISBN 978-3-7091-1855-9 .
- ^ Смит, Дэйв (28 сентября 2005 г.). «Вендские животные: Аркаруа » . Проверено 14 марта 2013 г.
- ^ Ваггонер, Бен (16 января 1995 г.). «Иглокожие: летопись окаменелостей» . Знакомство с иглокожими . Музей палеонтологии: Калифорнийский университет в Беркли . Проверено 14 марта 2013 г.
- ^ Jump up to: Перейти обратно: а б с Дорит, Уокер и Барнс 1991 , стр. 792–793.
- ^ UCMP Беркли. «Иглокожие: Морфология» . Музей палеонтологии Калифорнийского университета . Проверено 21 марта 2011 г.
- ^ Jump up to: Перейти обратно: а б Аршинов, Брэдли I; Кэри, Грегори А; Карими, Камран; Фоли, Сирша; Агалаков Сергей; Дельгадо, Франциско; Лотай, Ванит С; Ку, Кэролайн Дж; Пеллс, Трой Дж; Битман, Томас Р.; Ким, Юджин; Кэмерон, Р. Эндрю; Визе, Питер Д; Телмер, Шерил А; Кроче, Дженифер С; Эттенсон, Чарльз А; Хинман, Вероника Ф (7 января 2022 г.). «Эхинобаза: использование существующей базы данных модельных организмов для создания базы знаний, поддерживающей исследования в области геномики и биологии иглокожих» . Исследования нуклеиновых кислот . 50 (Д1): Д970–Д979. дои : 10.1093/nar/gkab1005 . ПМЦ 8728261 . ПМИД 34791383 .
- ^ Jump up to: Перейти обратно: а б Телмер, Шерил; Карими, Камран; Чесс, Мэйси; Агалаков Сергей; Аршинов, Брэдли; Лотай, Ванит; Ван, Дун Чжо; Чу, Стэнли; Пеллс, Трой; Визе, Питер; Хинман, Вероника; Эттенсон, Чарльз. «Echinobase: ресурс для поддержки исследовательского сообщества иглокожих» . Генетика . дои : 10.1093/генетика/iyae002 . ПМЦ 11075573 .
- ^ Jump up to: Перейти обратно: а б Руперт, Fox & Barnes 2004 , с. 873.
- ^ Бирн, Мари; О'Хара, Тим; ЦСИРО (2017). Австралийские иглокожие: биология, экология и эволюция . Клейтон Саут, Виктория. стр. 322–324. ISBN 978-1-4863-0763-0 . OCLC 960042778 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Холло, Габор; Новак, Михай (2012). «Гипотеза маневренности, объясняющая сохранение двусторонней симметрии в эволюции животных» . Биология Директ . 7 (1): 22. дои : 10.1186/1745-6150-7-22 . ПМЦ 3438024 . ПМИД 22789130 .
- ^ Мессинг, Чарльз. «Корона и чашечка» . Страницы криноидей Чарльза Мессинга . Архивировано из оригинала 29 октября 2013 года . Проверено 29 июля 2012 г.
- ^ Формери, Л.; Пелузо, П.; Конле, И.; Малник, Дж.; Томпсон-младший; Питель, М.; Улингер, КР; Рохсар, Д.С.; Ранг, ДР; Лоу, CJ (2023). «Молекулярные доказательства переднезаднего рисунка у взрослых иглокожих» . Природа . 623 (7987): 555–561. Бибкод : 2023Natur.623..555F . дои : 10.1038/s41586-023-06669-2 . ISSN 1476-4687 . ПМИД 37914929 .
- ↑ Новое генетическое исследование показывает, что морские звезды — это всего лишь кучка голов, слитых вместе.
- ^ Беренс, Питер; Бауэрляйн, Эдмунд (2007). Справочник по биоминерализации: биомиметическая и биоинспирированная химия» . Вайли-ВЧ. п. 393. ИСБН 978-3-527-31805-6 .
- ^ Бруска, Мур и Шустер, 2016 , стр. 979–980.
- ^ Дэвис, А. Морли (1925). Введение в палеонтологию . Томас Мерби. стр. 240–241.
- ^ Перилло, Маргарита; Улен, Натали; Фостер, Стефани; Сперрелл, Максвелл; Калестани, Кристина; Вессель, Гэри (19 августа 2020 г.). «Регуляция динамических состояний пигментных клеток при разрешении одной клетки» . электронная жизнь . 9 . дои : 10.7554/elife.60388 . ПМЦ 7455242 . ПМИД 32812865 .
- ^ Вебер, В.; Дамбах, М. (апрель 1974 г.). «Светочувствительность изолированных пигментных клеток морского ежа Centrostephanus longispinus ». Исследования клеток и тканей . 148 (3): 437–440. дои : 10.1007/BF00224270 . ПМИД 4831958 . S2CID 1669387 .
- ^ Мотокава, Тацуо (май 1984 г.). «Захват соединительной ткани у иглокожих». Биологические обзоры . 59 (2): 255–270. дои : 10.1111/j.1469-185X.1984.tb00409.x . S2CID 86391676 .
- ^ Бруска, Мур и Шустер 2016 , с. 980.
- ^ Jump up to: Перейти обратно: а б Дорит, Уокер и Барнс 1991 , стр. 780–791.
- ^ Бруска, Мур и Шустер, 2016 , стр. 980–982.
- ^ Дорит, Уокер и Барнс 1991 , стр. 784–785.
- ^ Бруска, Мур и Шустер 2016 , с. 982.
- ^ Дорит, Уокер и Барнс 1991 , стр. 790–793.
- ^ Жангу, Мишель (1982). «11. Пищеварительная система» . В Жангу, Мишель; Лоуренс, Джон М. (ред.). Питание иглокожих . Роттердам: А.А. Балкема. п. 244. ИСБН 978-90-6191-080-0 .
- ^ Ригби, П. Робин; Икен, Катрин; Сираяма, Ёсихиса (2007). Отбор проб биоразнообразия в прибрежных сообществах: протоколы NaGISA для местообитаний морских водорослей и макроводорослей . НУС Пресс. п. 44. ИСБН 978-9971693688 .
- ^ Руперт, Fox & Barnes 2004 , стр. 885.
- ^ Руперт, Fox & Barnes 2004 , стр. 891.
- ^ Рупперт, Fox & Barnes 2004 , стр. 902–904.
- ^ Руперт, Fox & Barnes 2004 , стр. 912.
- ^ Руперт, Fox & Barnes 2004 , стр. 920.
- ^ Мур, Джанет; Оверхилл, Райт (2006). Введение в беспозвоночных (2-е изд.). Издательство Кембриджского университета. п. 245. ИСБН 978-0521674065 .
- ^ Хикман, К. младший; Робертс, Л.; Ларсон, А. (2003). Разнообразие животных . Издательство Гранит Хилл. п. 271. ИСБН 978-0071195492 . ОСЛК 1148032077 .
- ^ «Макробентос Северного моря - Иглокожие > Введение» . etibioinformatics.nl .
- ^ Нильсен, Клаус (2012). Эволюция животных: взаимоотношения живых типов (3-е изд.). Издательство Оксфордского университета. п. 78. ИСБН 978-0199606023 .
- ^ Jump up to: Перейти обратно: а б Рамирес-Гомес, Франциско (27 сентября 2010 г.). «Иглокожий иммунитет» . Журнал выживания беспозвоночных .
- ^ Смит, Кортни (январь 2010 г.). «Иглокожий иммунитет» . Иммунитет беспозвоночных . Достижения экспериментальной медицины и биологии. Том. 708. С. 260–301. дои : 10.1007/978-1-4419-8059-5_14 . ISBN 978-1-4419-8058-8 . ПМИД 21528703 .
- ^ Смит, Л. Кортни; Гош, Джули; Бакли, Кэтрин М.; и др. (2010). «Иглокожий иммунитет». Иммунитет беспозвоночных . Достижения экспериментальной медицины и биологии. Том. 708. Бостон, Массачусетс: Спрингер. стр. 260–301. дои : 10.1007/978-1-4419-8059-5_14 . ISBN 978-1-4419-8058-8 . ПМИД 21528703 .
- ^ Jump up to: Перейти обратно: а б Руперт, Fox & Barnes 2004 , стр. 872–929.
- ^ Джеймс, Филип; Сиикавуопио, Стен; Йоханссон, Гунхильд Сельехауг (2018). «Руководство по репродуктивному циклу морских ежей и стадированию образцов гонад морского ежа» (PDF) . Нофима. Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 21 июня 2022 г.
ГИ [% массы тела, которую составляют гонады] у ежей в дикой природе может сильно различаться и может составлять от менее 1% до 20%, в то время как у культивируемых морских ежей значения ГИ могут достигать 35%.
- ^ Эдмондсон, Швейцария (1935). «Аутотомия и регенерация гавайских морских звезд» (PDF) . Периодические статьи Музея епископа . 11 (8): 3–20. Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Jump up to: Перейти обратно: а б МакАлари, Флоренс А. (1993). Структура популяции и воспроизводство делящихся морских звезд Linckia columbiae Grey на острове Санта-Каталина, Калифорния . 3-й симпозиум Калифорнийских островов. Служба национальных парков . Проверено 15 апреля 2012 г.
- ^ Jump up to: Перейти обратно: а б с См. последний абзац обзора выше. Анализ. Хочкисс, Фредерик ХК (1 июня 2000 г.). «О количестве лучей в морской звезде» . Американский зоолог . 40 (3): 340–354. дои : 10.1093/icb/40.3.340 .
- ^ Jump up to: Перейти обратно: а б с Фишер, WK (1 марта 1925 г.). «Бесполое размножение морских звезд Scleasterias » (PDF) . Биологический бюллетень . 48 (3): 171–175. дои : 10.2307/1536659 . JSTOR 1536659 . Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 15 июля 2011 г.
- ^ Добсон, МЫ; С.Е. Станчик; Л.А. Клементс; Р. М. Шоумен (1 февраля 1991 г.). «Транслокация питательных веществ во время ранней регенерации диска у Brittlestar Microphiopholis gracillima (Stimpson) (Echinodermata: Ophiuroidea)» . Биол Булл . 180 (1): 167–184. дои : 10.2307/1542439 . JSTOR 1542439 . ПМИД 29303627 . Проверено 14 июля 2011 г.
- ^ Кандия-Карневали, доктор медицины; Торндайк, MC; Матранга, В. (2009). «Регенерация иглокожих: обещание понять потенциал стволовых клеток». Ринкевич, Б.; Матранга, В. (ред.). Стволовые клетки в морских организмах . Дордрехт: Спрингер . стр. 165–186. дои : 10.1007/978-90-481-2767-2_7 . ISBN 978-90-481-2766-5 .
- ^ Машанов Владимир С.; Игорь Ю. Долматов; Томас Хайнцеллер (1 декабря 2005 г.). «Трансдифференциация при регенерации кишечника голотурий» . Биологический вестник . 209 (3): 184–193. дои : 10.2307/3593108 . JSTOR 3593108 . ПМИД 16382166 . S2CID 27847466 .
- ^ Янг, Крейг М.; Экельбаргер, Кевин Дж. (1994). Размножение, биология личинок и пополнение глубоководного бентоса . Издательство Колумбийского университета. стр. 179–194. ISBN 0231080042 .
- ^ Рупперт, Fox & Barnes 2004 , стр. 887–888.
- ^ Руперт, Fox & Barnes 2004 , стр. 895.
- ^ Руперт, Fox & Barnes 2004 , стр. 888.
- ^ Руперт, Fox & Barnes 2004 , стр. 908.
- ^ Руперт, Fox & Barnes 2004 , стр. 916.
- ^ Руперт, Fox & Barnes 2004 , стр. 922.
- ^ Ямагучи, М.; Дж. С. Лукас (1984). «Естественный партеногенез, развитие личинок и молоди и географическое распространение астероида коралловых рифов Ophidiaster granifer ». Морская биология . 83 (1): 33–42. Бибкод : 1984МарБи..83...33Г . дои : 10.1007/BF00393083 . S2CID 84475593 .
- ^ Макговерн, Тамара М. (5 апреля 2002 г.). «Способы полового и бесполого размножения хрупкой звезды Ophiactis savigni на островах Флорида-Кис» (PDF) . Серия «Прогресс в области морской экологии» . 230 : 119–126. Бибкод : 2002MEPS..230..119M . дои : 10.3354/meps230119 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Монкс, Сара П. (1 апреля 1904 г.). «Изменчивость и автотомия Фатарии ». Труды Академии естественных наук Филадельфии . 56 (2): 596–600. JSTOR 4063000 .
- ^ Килле, Фрэнк Р. (1942). «Регенерация репродуктивной системы после бинарного деления у трепанга Holothuria parvula (Selenka)» (PDF) . Биологический бюллетень . 83 (1): 55–66. дои : 10.2307/1538013 . JSTOR 1538013 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Jump up to: Перейти обратно: а б Ивз, Александра А.; Палмер, А. Ричард (11 сентября 2003 г.). «Размножение: широко распространенное клонирование личинок иглокожих». Природа . 425 (6954): 146. Бибкод : 2003Natur.425..146E . дои : 10.1038/425146a . hdl : 10402/era.17195 . ПМИД 12968170 . S2CID 4430104 .
- ^ Джекл, Уильям Б. (1 февраля 1994 г.). «Множественные способы бесполого размножения тропических и субтропических личинок морских звезд: необычная адаптация к расселению и выживанию генов» (PDF) . Биологический бюллетень . 186 (1): 62–71. дои : 10.2307/1542036 . JSTOR 1542036 . ПМИД 29283296 . Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 13 июля 2011 г.
- ^ Jump up to: Перейти обратно: а б с Вон, Дон (октябрь 2009 г.). «Клонирование личинок песчаного доллара Dendraster excentricus, вызванное хищниками : может ли мать иметь значение?» . Биологический бюллетень . 217 (2): 103–114. дои : 10.1086/BBLv217n2p103 . ПМИД 19875816 . S2CID 26030855 .
- ^ Макдональд, Кэтрин А.; Дон Вон (1 августа 2010 г.). «Резкое изменение пищевой среды вызывает клонирование Plutei Dendraster excentricus » . Биологический бюллетень . 219 (1): 38–49. дои : 10.1086/BBLv219n1p38 . ПМИД 20813988 . S2CID 24422314 . Проверено 16 июля 2011 г.
- ^ Jump up to: Перейти обратно: а б Вон, Дон; Ричард Р. Стратманн (14 марта 2008 г.). «Хищники вызывают клонирование личинок иглокожих» . Наука . 319 (5869): 1503. Бибкод : 2008Sci...319.1503V . дои : 10.1126/science.1151995 . JSTOR 40284699 . ПМИД 18339931 . S2CID 206509992 . Проверено 16 июля 2011 г.
- ^ Вон, Дон (март 2010 г.). «Зачем бежать и прятаться, если можно разделить? Доказательства клонирования личинок и уменьшения их размера как адаптивной индуцируемой защиты». Морская биология . 157 (6): 1301–1312. Бибкод : 2010МарБи.157.1301В . дои : 10.1007/s00227-010-1410-z . S2CID 84336247 .
- ^ Jump up to: Перейти обратно: а б Дорит, Уокер и Барнс 1991 , с. 778.
- ^ Бруска, Мур и Шустер 2016 , стр. 997–998.
- ^ ван Эгмонд, Вим (1 июля 2000 г.). «Галерея личинок иглокожих» . Микроскопия Великобритании . Проверено 2 февраля 2013 г.
- ^ Jump up to: Перейти обратно: а б Бруска, Мур и Шустер, 2016 , с. 968.
- ^ Сяофэн, Ван; Хагдорн, Ганс; Чуаньшан, Ван (сентябрь 2006 г.). «Псевдопланктонный образ жизни триасового криноида Traumatocrinus из Юго-Западного Китая». Летайя . 39 (3): 187–193. Бибкод : 2006Letha..39..187X . дои : 10.1080/00241160600715321 .
- ^ Смит, Дж. Э. (1937). «Структура и функции трубчатых ножек у некоторых иглокожих» (PDF) . Журнал Морской биологической ассоциации Соединенного Королевства . 22 (1): 345–357. Бибкод : 1937JMBUK..22..345S . дои : 10.1017/S0025315400012042 . S2CID 55933156 . Архивировано из оригинала (PDF) 15 ноября 2013 года.
- ^ Эстли, ХК (2012). «Передвижение, когда вы рядом: количественный анализ передвижения хрупкой звезды с тупыми шипами Ophiocoma echinata» . Журнал экспериментальной биологии . 215 (11): 1923–1929. дои : 10.1242/jeb.068460 . ПМИД 22573771 .
- ^ Бруска, Мур и Шустер, 2016 , стр. 982–983.
- ^ «Песчаная звезда — Luidia foliolata » . Морские звезды северо-запада Тихого океана. Архивировано из оригинала 9 сентября 2012 года . Проверено 26 сентября 2012 г.
- ^ Рупперт, Fox & Barnes 2004 , стр. 899–900.
- ^ Рупперт, Fox & Barnes 2004 , стр. 911–912.
- ^ Мессинг, Чарльз. «Механизм питания криноидей» . Страницы криноидей Чарльза Мессинга . Архивировано из оригинала 14 января 2013 года . Проверено 26 июля 2012 года .
- ^ Jump up to: Перейти обратно: а б Барнс, Роберт Д. (1982). Зоология беспозвоночных . Холт-Сондерс Интернэшнл. стр. 997–1007. ISBN 0-03-056747-5 .
- ^ Jump up to: Перейти обратно: а б Руперт, Fox & Barnes 2004 , с. 893.
- ^ Jump up to: Перейти обратно: а б Кэрфут, Том. «Узнайте о морских ежах: кормление, питание и рост» . Одиссея улитки . Архивировано из оригинала 22 марта 2013 года . Проверено 23 февраля 2013 г.
- ^ Jump up to: Перейти обратно: а б Руперт, Fox & Barnes 2004 , с. 914.
- ^ Jump up to: Перейти обратно: а б Руперт, Fox & Barnes 2004 , стр. 884–885.
- ^ Кэрфут, Том. «Узнайте о перьевых звездах: питание и рост» . Одиссея улитки . Архивировано из оригинала 12 января 2013 года . Проверено 23 февраля 2013 г.
- ^ Кэрфут, Том. «Узнайте о песчаных долларах: кормление и рост» . Одиссея улитки . Архивировано из оригинала 19 октября 2012 года . Проверено 23 февраля 2013 г.
- ^ Миллер, Джон Э. «Иглокожие: роль в природе» . Британская энциклопедия . Проверено 23 июня 2022 г.
- ^ Дорит, Уокер и Барнс 1991 , с. 779.
- ^ Дорит, Уокер и Барнс 1991 , стр. 789–790.
- ^ Младенов Филипп Владимирович; Игдура, Сулейман; Асотра, Сатиш; Берк, Роберт Д. (1 апреля 1989 г.). «Очистка и частичная характеристика фактора, способствующего автотомии, из морской звезды Pycnopodia Helianthoides » . Биологический вестник . 176 (2): 169–175. дои : 10.2307/1541585 . JSTOR 1541585 .
- ^ Эррера-Эскаланте, Т.; Лопес-Перес, РА; Лейте-Моралес, GE (2005). «Биоэрозия, вызванная морским ежом Diadema mexicanum (Echinodermata: Echinoidea) в заливах Уатулько, Западная Мексика» Журнал тропической биологии . 53 (3): 263–273. ПМИД 17469255 .
- ^ Лебрато, Марио; Иглесиас-Родригес, Дебора; Фили, Ричард А.; и др. (2010). «Глобальный вклад иглокожих в морской углеродный цикл: баланс CaCO3 и донные отсеки» . Экологические монографии . 80 (3): 441–467. Бибкод : 2010ЭкоМ...80..441Л . дои : 10.1890/09-0553.1 . hdl : 10261/79231 .
- ^ Осборн, Патрик Л. (2000). Тропическая экосистема и экологические концепции . Издательство Кембриджского университета. п. 464. ИСБН 0-521-64523-9 .
- ^ Лоуренс, Дж. М. (1975). «О взаимоотношениях морских растений и морских ежей» (PDF) . Океанографический морской биологический ежегодный обзор . 13 : 213–286. Архивировано из оригинала (PDF) 5 февраля 2011 года.
- ^ Биркеланд, Чарльз; Лукас, Джон С. (1990). «Воздействие A. planci на сообщества коралловых рифов». Acanthaster planci : Основная проблема управления коралловыми рифами . Флорида: CRC Press. стр. 148–159. ISBN 0849365996 .
- ^ «B-76 Морские ежи и другие иглокожие. Добыча вылова по видам, районам промысла и странам или районам» (PDF) . ФАО. стр. 463–465. Архивировано (PDF) из оригинала 9 октября 2022 года . Проверено 23 июня 2022 г.
- ^ Пангесути, Ратих; Арифин, Зайнал (2018). «Лекарственные и полезные для здоровья эффекты функциональных трепангов» . Журнал традиционной и дополнительной медицины . 8 (3): 341–351. дои : 10.1016/j.jtcme.2017.06.007 . ПМК 6035309 . ПМИД 29992105 .
- ^ «Морские огурцы находятся под угрозой со стороны азиатской торговли» . Нью-Йорк Таймс . Апрель 2009 года . Проверено 1 апреля 2009 г.
- ^ Wikisource: Британская энциклопедия 1911 года.
- ^ Лоуренс, Джон М. (2001). «Съедобные морские ежи». В Джоне М. Лоуренсе (ред.). Съедобные морские ежи: биология и экология . Развитие науки в области аквакультуры и рыболовства. Том. 37. стр. 1–4. дои : 10.1016/S0167-9309(01)80002-8 . ISBN 978-0444503909 .
- ^ Дэвидсон, Алан (2014) Оксфордский справочник по еде . Издательство Оксфордского университета , 3-е издание. п. 730.
- ^ Сартори Д., Скудери А.; Сансоне Г.; Гайон А. (февраль 2015 г.). «Эхинокультура: выращивание Paracentrotus lividus в системе замкнутой аквакультуры — эксперименты с искусственными диетами для поддержания полового созревания». Международная Аквакультура . 23 (1): 111–125. Бибкод : 2015AqInt..23..111S . дои : 10.1007/s10499-014-9802-6 . S2CID 18856183 .
- ^ Jump up to: Перейти обратно: а б «Взгляд морского ежа» . Станция визуализации микроскопа . Эксплораториум. Архивировано из оригинала 5 февраля 2018 года . Проверено 23 июня 2022 г.
- ^ Сартори, Д.; Гайон, А. (2015). «Токсичность полиненасыщенных альдегидов диатомовых водорослей для индо-тихоокеанского биоиндикатора Echinometra mathaei». Лекарственная и химическая токсикология . 39 (2): 124–8. дои : 10.3109/01480545.2015.1041602 . ПМИД 25945412 . S2CID 207437380 .
- ^ Гайон, А.; Скудери, А.; Пеллегрини, Д.; Сартори, Д. (2013). «Воздействие мышьяка влияет на развитие эмбриона морского ежа Paracentrotus lividus (Ламарк, 1816)». Бюллетень загрязнения окружающей среды и токсикологии . 39 (2): 124–8. дои : 10.3109/01480545.2015.1041602 . ПМИД 25945412 . S2CID 207437380 .
- ^ Харт, М.В. (январь – февраль 2002 г.). «Эволюция истории жизни и сравнительная биология развития иглокожих». Эволюция и развитие . 4 (1): 62–71. дои : 10.1046/j.1525-142x.2002.01052.x . ПМИД 11868659 . S2CID 39175905 .
- ^ Лонго, Ф.Дж.; Андерсон, Э. (июнь 1969 г.). «Дифференциация сперматозоидов морских ежей Arbacia punctulata и Strongylocentrotus purpuratus ». Журнал исследований ультраструктуры . 27 (5): 486–509. дои : 10.1016/S0022-5320(69)80046-8 . ПМИД 5816822 .
- ^ Jump up to: Перейти обратно: а б Баркхаус, К.; Найлз, М.; Дэвидсон, Луизиана (2007). «Обзор литературы по методам борьбы с морскими звездами при выращивании донных и наддонных моллюсков» (PDF) . Отчет канадской промышленности по рыболовству и водным наукам . 279 (7): 14–15. Архивировано из оригинала (PDF) 22 мая 2013 года.
- ^ «Морские звезды» . Дикий Сингапур . Проверено 4 февраля 2013 г.
Цитируемые тексты
[ редактировать ]- Бруска, Ричард К.; Мур, Венди; Шустер, Стивен М. (2016). Беспозвоночные (3-е изд.). Сандерленд, Массачусетс. ISBN 978-1-60535-375-3 . OCLC 928750550 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - Дорит, РЛ; Уокер, ВФ; Барнс, Р.Д. (1991). Зоология, Международное издание . Издательство Колледжа Сондерса. ISBN 978-0-03-030504-7 .
- Руперт, Эдвард Э.; Фокс, Ричард, С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных (7-е изд.). Cengage Обучение. ISBN 81-315-0104-3 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка )