Морской еж
| Морской еж Временной диапазон:
| |
|---|---|

| |
| Tripneustes ventricosus и Echinometra viridis. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Иглокожие |
| Подтип: | Эхинозоа |
| Сорт: | Эхинодея Леске , 1778 г. |
| Подклассы | |
| |
Морские ежи или ежи ( / ˈ ɜːr tʃ ɪ n z / ), также известные как морские ежи, обычно представляют собой колючие шаровидные животные , иглокожие из класса Echinoidea. Около 950 видов живут на морском дне, населяя все океаны и зоны глубин от приливной зоны до 5000 метров (16000 футов; 2700 саженей). [ 1 ] Их раковины (твердые панцири) круглые и колючие, обычно от 3 до 10 см (от 1 до 4 дюймов) в поперечнике. Морские ежи передвигаются медленно, ползая на трубчатых ногах , а иногда и толкая себя шипами. Они питаются в основном водорослями , но также поедают медленно движущихся или сидячих животных. Их хищниками являются каланы , морские звезды , волчьи угри и спинороги .
Как и все иглокожие, взрослые морские ежи обладают пятикратной симметрией, а их личинки плутеуса обладают двусторонней (зеркальной) симметрией ; Последнее указывает на их принадлежность к Bilateria наряду с хордовыми , членистоногими , кольчатыми червями и моллюсками . Морские ежи встречаются в каждом океане и в любом климате, от тропиков до полярных регионов , и населяют морские донные (морские) места обитания, от скалистых берегов до глубин хадальной зоны . Летопись окаменелостей ежовых рыб датируется ордовикским периодом, около 450 миллионов лет назад. Ближайшими иглокожими родственниками морского ежа являются морские огурцы (Holothuroidea), которые, как и они, относятся к вторичноротым , кладе, включающей хордовых . ( Песчаные доллары представляют собой отдельный отряд морских ежей класса Echinoidea.)
Животные изучались с 19 века как модельные организмы в биологии развития , поскольку за их эмбрионами было легко наблюдать. Это продолжилось с изучением их геномов из-за их необычной пятикратной симметрии и родства с хордовыми. Такие виды, как грифельный еж, популярны в аквариумах, где они полезны для борьбы с водорослями. Ископаемые ежи использовались в качестве защитных амулетов .
Разнообразие
[ редактировать ]Морские ежи относятся к типу Echinodermata , в который также входят морские звезды, морские огурцы , песчаные доллары , хрупкие звезды и криноидеи . Как и другие иглокожие, они обладают пятикратной симметрией (так называемая пентамерия ) и передвигаются с помощью сотен крошечных, прозрачных, клейких « ножек-трубочек ». Симметрия не очевидна у живого животного, но легко видна в высушенном тесте . [ 2 ]
В частности, термин «морской еж» относится к «правильным ежам», которые являются симметричными и шаровидными, и включает в себя несколько различных таксономических групп с двумя подклассами: Euechinoidea («современные» морские ежи, в том числе неправильные) и Cidaroidea , или « «грифельно-карандашные ежи», имеющие очень толстые, тупые шипы, на которых растут водоросли и губки. «Неправильные» морские ежи представляют собой инфра-класс внутри Euechinoidea, называемый Irregularia , и включают Atelostomata и Neognathostomata . К неправильным ежам относятся сплюснутые песчанки , морские ежи и сердечные ежи . [ 3 ]
Вместе с голотуриями ( Holothuroidea ) они составляют подтип Echinozoa , для которого характерна шаровидная форма без рукавов и выступающих лучей. Морские огурцы и морские ежи неправильной формы вторично приобрели разнообразные формы. Хотя многие морские огурцы имеют разветвленные щупальца , окружающие их ротовые отверстия, они произошли от видоизмененных трубчатых ножек и не гомологичны рукам криноидей, морских звезд и хрупких звезд. [ 2 ]
-

-
 Песчаный доллар , морской еж неправильной формы ( Irregularia ).
Песчаный доллар , морской еж неправильной формы ( Irregularia ). -
 Phyllacanthus Imperialis , цидароидный морской еж ( Cidaroidea )
Phyllacanthus Imperialis , цидароидный морской еж ( Cidaroidea )



Описание
[ редактировать ]
Размер ежей обычно варьируется от 3 до 10 см (от 1 до 4 дюймов), но самые крупные виды могут достигать 36 см (14 дюймов). [ 4 ] У них жесткое, обычно сферическое тело с подвижными шипами, которые дали этому классу название Echinoidea (от греческого ἐχῖνος ehinos «позвоночник»). [ 5 ] Название «еж» — старое слово, обозначающее «ежа» , на которого похожи морские ежи; их архаично называли морскими ежами . [ 6 ] [ 7 ] Название происходит от старофранцузского herichun , от латинского ericius («еж»). [ 8 ]
Как и другие иглокожие, ранние личинки морского ежа обладают двусторонней симметрией. [ 9 ] но по мере взросления у них развивается пятикратная симметрия. Это наиболее очевидно у «обычных» морских ежей, у которых тела примерно сферической формы с пятью частями одинакового размера, расходящимися от их центральных осей. Рот находится у основания животного, анус - наверху; нижняя поверхность описывается как «оральная», а верхняя — как «аборальная». [ а ] [ 2 ]
Однако некоторые морские ежи, в том числе песчаные доллары, имеют овальную форму с четко выраженными передним и задним концами, что придает им некоторую степень двусторонней симметрии. У этих ежей верхняя поверхность тела слегка куполообразная, а нижняя плоская, а бока лишены трубчатых ножек. Эта «неправильная» форма тела развилась, чтобы позволить животным рыться в песке или других мягких материалах. [ 4 ]
Системы
[ редактировать ]Опорно-двигательный аппарат
[ редактировать ]
Внутренние органы заключены в твердую оболочку или панцирь, состоящую из сросшихся пластинок карбоната кальция, покрытых тонкой дермой и эпидермисом . Тест называется эндоскелетом, а не экзоскелетом, хотя он охватывает почти весь еж. Это потому, что он покрыт тонким слоем мышц и кожи; морским ежам также не нужно линять, как это делают беспозвоночные с настоящим экзоскелетом, вместо этого пластинки, образующие панцирь, растут так же, как и животное.
Тесто жесткое и делится на пять амбулакральных бороздок, разделенных пятью более широкими межамбулакральными областями. Каждая из этих десяти продольных колонн состоит из двух наборов пластин (всего 20 колонн). Амбулакральные пластинки имеют пары крошечных отверстий, через которые проходят ножки трубки. [ 10 ]
Все пластинки покрыты округлыми бугорками, к которым прикреплены шипы. Шипы используются для защиты и передвижения и бывают самых разных форм. [ 11 ] Внутренняя поверхность яичка выстлана брюшиной . [ 4 ] Морские ежи преобразуют водный диоксид углерода с помощью каталитического процесса с участием никеля в часть теста, содержащую карбонат кальция. [ 12 ]

У большинства видов имеются две серии шипов: первичные (длинные) и вторичные (короткие), распределенные по поверхности тела, причем самые короткие на полюсах и самые длинные на экваторе. Колючки обычно полые и цилиндрические. Сокращение мышечной оболочки, покрывающей яичко, заставляет позвоночник наклоняться в ту или иную сторону, а внутренняя оболочка из коллагеновых волокон может обратимо меняться с мягкой на жесткую, что может фиксировать позвоночник в одном положении. Среди шипов расположены несколько видов педицеллярий — подвижных стебельчатых структур с челюстями. [ 2 ]
Морские ежи передвигаются при ходьбе, используя свои многочисленные гибкие трубчатые ножки, подобно морским звездам; У обычных морских ежей нет любимого направления прогулок. [ 13 ] Ножки трубки выступают через пары пор в тесте и управляются водной сосудистой системой ; это работает за счет гидравлического давления , позволяя морскому ежу закачивать воду в ножки трубки и из нее. Во время передвижения ногам трубки помогают шипы, с помощью которых можно толкать тело вперед или поднимать тест с субстрата. Передвижение обычно связано с кормлением: красный морской еж ( Mesocentrotus franciscanus ) преодолевает около 7,5 см (3 дюйма) в день, когда еды достаточно, и до 50 см (20 дюймов) в день, когда ее нет. Перевернутый морской еж может выпрямиться, постепенно прикрепляя и отсоединяя трубчатые ножки и манипулируя шипами, чтобы перевернуть тело в вертикальное положение. [ 2 ] Некоторые виды зарываются в мягкий осадок с помощью шипов, а Paracentrotus lividus использует челюсти, чтобы зарываться в мягкие камни. [ 14 ]
-
 Тест Echinus esculentus , обычного морского ежа.
Тест Echinus esculentus , обычного морского ежа. -
 Тест черного морского ежа с бугорками и амбулакральными пластинками (справа)
Тест черного морского ежа с бугорками и амбулакральными пластинками (справа) -
 На внутренней поверхности теста справа видны пятиугольные межамбулакральные пластины, а слева — отверстия для ножек трубки.
На внутренней поверхности теста справа видны пятиугольные межамбулакральные пластины, а слева — отверстия для ножек трубки. -

-
 Тест Phyllacanthus Imperialis , цидароидного морского ежа. Для них характерны большие бугорки с крупными радиолами.
Тест Phyllacanthus Imperialis , цидароидного морского ежа. Для них характерны большие бугорки с крупными радиолами. -
 Крупный план теста, показывающий амбулакральную бороздку с двумя рядами пар пор между двумя областями интерамбулакров (зеленый). Бугорки неперфорированные.
Крупный план теста, показывающий амбулакральную бороздку с двумя рядами пар пор между двумя областями интерамбулакров (зеленый). Бугорки неперфорированные. -
![Крупный план апикального диска цидароидного морского ежа: 5 отверстий представляют собой гонопоры, а центральное - анус («перипрокт»). Самая большая половая пластинка — мадрепорит.[15]](//upload.wikimedia.org/wikipedia/commons/thumb/9/9e/Sea_Urchin_Shell_detail.jpg/270px-Sea_Urchin_Shell_detail.jpg) Крупный план апикального диска цидароидного морского ежа: 5 отверстий представляют собой гонопоры, а центральное - анус («перипрокт»). Самая крупная половая пластинка — мадрепорит . [ 15 ]
Крупный план апикального диска цидароидного морского ежа: 5 отверстий представляют собой гонопоры, а центральное - анус («перипрокт»). Самая крупная половая пластинка — мадрепорит . [ 15 ]






![Крупный план апикального диска цидароидного морского ежа: 5 отверстий представляют собой гонопоры, а центральное - анус («перипрокт»). Самая большая половая пластинка — мадрепорит.[15]](http://upload.wikimedia.org/wikipedia/commons/thumb/9/9e/Sea_Urchin_Shell_detail.jpg/270px-Sea_Urchin_Shell_detail.jpg)
Питание и пищеварение
[ редактировать ]
Рот расположен в центре поверхности рта у обычных ежей или ближе к одному концу у неправильных ежей. Он окружен губами из более мягких тканей с многочисленными маленькими костными кусочками. Эта область, называемая перистомом, также включает пять пар видоизмененных трубчатых ножек и, у многих видов, пять пар жабр. [ 4 ] Челюстной аппарат состоит из пяти прочных стреловидных пластинок, известных как пирамиды, вентральная поверхность каждой из которых имеет зубную ленту с твердым зубцом, направленным к центру рта. Специализированные мышцы контролируют выдвижение аппарата и действие зубов, животное может хватать, царапать, тянуть и рвать. [ 2 ] Было обнаружено, что структура рта и зубов настолько эффективна при захвате и измельчении, что подобные структуры были протестированы для использования в реальных приложениях. [ 16 ]
На верхней поверхности яичка у аборального полюса находится мембрана — перипрокт , окружающая задний проход . Перипрокт содержит различное количество твердых пластинок, пять из которых, генитальные пластинки, содержат гонопоры, а одна модифицирована и содержит мадрепорит , который используется для баланса водной сосудистой системы. [ 2 ]

Рот большинства морских ежей состоит из пяти зубов или пластинок карбоната кальция с мясистой, похожей на язык структурой внутри. Весь жевательный орган известен как фонарь Аристотеля из Аристотеля описания в его «Истории животных» (перевод Д'Арси Томпсона ):
... у ежа есть то, что мы обычно называем головой и ртом внизу, и место для выхода остатков вверху. У ежа также есть пять полых зубов внутри, а в середине этих зубов мясистое вещество, служащее языком . Далее идет пищевод , а затем желудок , разделенный на пять частей и наполненный выделениями, причем все пять частей соединяются у анального отверстия, где оболочка перфорирована для выхода... На самом деле ротовой аппарат еж непрерывен от одного конца до другого, но по внешнему виду это не так, а похоже на роговой фонарь с оставленными роговыми стеклами.
Однако недавно было доказано, что это неправильный перевод. Фонарь Аристотеля на самом деле относится к форме морских ежей, которые похожи на древние лампы времен Аристотеля. [ 17 ] [ 18 ]
Сердечные ежи необычны тем, что у них нет фонаря. Вместо этого рот окружен ресничками , которые тянут нити слизи, содержащей частицы пищи, к бороздкам вокруг рта. [ 4 ]
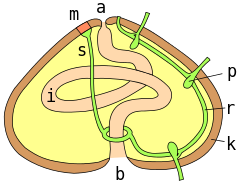
а = анус ; м = мадрепорит ; s = водоносный канал; r = радиальный канал; p = подиальная ампула; k = испытательная стена; я = кишечник ; б = рот
Фонарь, если он есть, окружает ротовую полость и глотку . В верхней части фонаря глотка открывается в пищевод, который спускается по внешней стороне фонаря и соединяется с тонкой кишкой и единственной слепой кишкой . Тонкая кишка проходит полный круг внутри теста, прежде чем присоединиться к толстой кишке, которая завершает еще один круг в противоположном направлении. Из толстой кишки прямая кишка поднимается к анусу. Несмотря на названия, тонкий и толстый кишечник морских ежей ни в коей мере не гомологичен одноименным структурам позвоночных. [ 4 ]
Пищеварение происходит в кишечнике, при этом слепая кишка вырабатывает дополнительные пищеварительные ферменты . Дополнительная трубка, называемая сифоном, проходит вдоль большей части кишечника и открывается в него с обоих концов. Он может участвовать в резорбции воды из пищи. [ 4 ]
Кровообращение и дыхание
[ редактировать ]
Водная сосудистая система ведет вниз от мадрепорита через тонкий каменный канал к кольцевому каналу, который окружает пищевод. Отсюда через каждую амбулакральную область проходят радиальные каналы, заканчивающиеся небольшим щупальцем, которое проходит через амбулакральную пластинку возле аборального полюса. От этих радиальных каналов отходят латеральные каналы, заканчивающиеся ампулами. Отсюда две трубки проходят через пару пор на пластине и заканчиваются ножками трубки. [ 2 ]
Морские ежи обладают гемальной системой со сложной сетью сосудов в брыжейках вокруг кишечника, но о функционировании этой системы мало что известно. [ 2 ] Однако основная кровеносная жидкость заполняет общую полость тела, или целом . Эта целомическая жидкость содержит фагоцитирующие целомоциты, которые перемещаются по сосудистой и гемальной системам и участвуют во внутреннем транспорте и газообмене. Целомоциты играют важную роль в свертывании крови , но также собирают отходы и активно выводят их из организма через жабры и ножки трубок. [ 4 ]
У большинства морских ежей пять пар внешних жабр, прикрепленных к перистомиальной мембране вокруг рта. Эти тонкостенные выступы полости тела являются основными органами дыхания у тех ежей, которые ими обладают. Жидкость может перекачиваться через внутреннюю часть жабр мышцами, связанными с фонарем, но это не обеспечивает непрерывного потока и происходит только тогда, когда у животного мало кислорода. Трубчатые ножки также могут действовать как органы дыхания и являются основными местами газообмена у сердечных ежей и песчаных долларов, у которых нет жабр. Внутренняя часть каждого основания трубки разделена перегородкой, которая уменьшает диффузию между входящим и выходящим потоками жидкости. [ 2 ]
Нервная система и органы чувств
[ редактировать ]Нервная система морских ежей имеет сравнительно простое устройство. Не имея настоящего мозга, нервный центр представляет собой большое нервное кольцо, окружающее рот прямо внутри фонаря. От нервного кольца пять нервов расходятся под радиальными каналами водной сосудистой системы и разветвляются на множество более тонких нервов, иннервирующих ножки трубки, шипы и педицеллярии . [ 4 ]
Морские ежи чувствительны к прикосновениям, свету и химическим веществам. В эпителии имеется множество чувствительных клеток, особенно в шипах, педицелляриях и ножках трубки, а также вокруг рта. [ 2 ] Хотя у них нет глаз или глазных пятен (за исключением диадематид , которые могут преследовать угрозу своими шипами), все тело большинства обычных морских ежей может функционировать как сложный глаз. [ 19 ] Как правило, морских ежей отрицательно привлекает свет, и они стремятся спрятаться в расщелинах или под предметами. У большинства видов, за исключением карандашных ежей имеются статоцисты , в шаровидных органах, называемых сферидиями, . Это стебельчатые структуры, расположенные в пределах амбулакральных областей; их функция — помогать в гравитационной ориентации. [ 4 ]
История жизни
[ редактировать ]Воспроизведение
[ редактировать ]Морские ежи раздельнополы , имеют отдельные мужской и женский пол, хотя внешне никаких отличительных особенностей не видно. Помимо своей роли в размножении, гонады также являются органами хранения питательных веществ и состоят из двух основных типов клеток: половых клеток и соматических клеток , называемых питательными фагоцитами. [ 20 ] У обычных морских ежей пять гонад, лежащих под межамбулакральными областями панциря, тогда как у неправильных форм в основном четыре, причем самая задняя гонада отсутствует; у сердечных ежей их три или два. Каждая гонада имеет единственный проток, поднимающийся от верхнего полюса и открывающийся в гонопор, лежащий в одной из половых пластинок, окружающих анус. У некоторых роющих песчаных долларов есть удлиненный сосочек, который позволяет высвобождать гаметы над поверхностью осадка. [ 2 ] Гонады покрыты мышцами под брюшиной, что позволяет животному выдавливать свои гаметы через проток в окружающую морскую воду, где оплодотворение . происходит [ 4 ]
Разработка
[ редактировать ]
На раннем этапе развития эмбрион морского ежа претерпевает 10 циклов клеточного деления . [ 21 ] В результате образуется один слой эпителия , окутывающий бластоцель . Затем эмбрион начинает гаструляцию , многочастный процесс, который радикально перестраивает его структуру путем инвагинации с образованием трех зародышевых листков , включая эпителиально-мезенхимальный переход ; первичные мезенхимные клетки перемещаются в бластоцель [ 22 ] и стать мезодермой . [ 23 ] Было высказано предположение, что эпителиальной полярности вместе с полярностью плоских клеток может быть достаточно для запуска гаструляции у морских ежей. [ 24 ]

личинки Необычной особенностью развития морского ежа является замена двусторонней симметрии пятикратной симметрией взрослой особи. При дроблении выделяются мезодерма и мелкие микромеры. В конце гаструляции клетки этих двух типов образуют целомические сумки. На личиночных стадиях взрослый зачаток вырастает из левого целомического мешка; после метаморфоза этот зачаток вырастает и становится взрослым. Животно -растительная ось устанавливается до оплодотворения яйцеклетки. Орально-аборальная ось определяется на ранних стадиях дробления, а левая-правая ось появляется на поздней стадии гаструлы. [ 25 ]
Жизненный цикл и развитие
[ редактировать ]
В большинстве случаев икра самки свободно плавает в море, но некоторые виды удерживают ее шипами, обеспечивая большую степень защиты. Неоплодотворенная яйцеклетка встречается со свободно плавающей спермой, выделяемой самцами, и бластулы всего за 12 часов развивается в свободно плавающий эмбрион . Первоначально представляющая собой простой клубок клеток, бластула вскоре трансформируется в конусообразную личинку эхиноплютеуса . У большинства видов эта личинка имеет 12 удлиненных рук, покрытых полосками ресничек, которые захватывают частицы пищи и транспортируют их ко рту. У некоторых видов бластула содержит запасы питательного желтка и не имеет рук, поскольку не нуждается в питании. [ 4 ]
Для завершения развития личинки требуется несколько месяцев, причем переход во взрослую форму начинается с образования тестовых пластинок в ювенильном зачатке, который развивается на левой стороне личинки и его ось перпендикулярна оси личинки. Вскоре личинка опускается на дно и превращается в молодого ежа. всего за час [ 2 ] У некоторых видов взрослые особи достигают максимального размера примерно за пять лет. [ 4 ] Пурпурный еж становится половозрелым через два года и может прожить двадцать. [ 26 ]
Долголетие
[ редактировать ]Первоначально считалось, что красные морские ежи живут от 7 до 10 лет, но недавние исследования показали, что они могут жить более 100 лет. Было обнаружено, что канадским красным ежам около 200 лет. [ 27 ] [ 28 ]
Экология
[ редактировать ]Трофический уровень
[ редактировать ]Морские ежи питаются в основном водорослями , поэтому они в основном травоядны , но могут питаться морскими огурцами и широким спектром беспозвоночных, таких как мидии , полихеты , губки , хрупкие звезды и криноидеи, что делает их всеядными и потребителями различных трофических свойств. уровни . [ 29 ]
Хищники, паразиты и болезни
[ редактировать ]Массовая смертность морских ежей впервые была зарегистрирована в 1970-х годах, но до появления аквакультуры болезни морских ежей были мало изучены. В 1981 году бактериальная «пятнистая болезнь» вызвала почти полную смертность молодых особей Pseudocentrotus depressus и Hemicentrotus pulcherrimus , культивируемых в Японии; болезнь рецидивировала в последующие годы. Ее разделили на прохладноводную «весеннюю» болезнь и жарководную «летнюю» форму. [ 30 ] Другое заболевание, болезнь лысого морского ежа , вызывает потерю шипов и повреждения кожи и, как полагают, имеет бактериальное происхождение. [ 31 ]
Взрослые морские ежи обычно хорошо защищены от большинства хищников своими сильными и острыми шипами, которые у некоторых видов могут быть ядовитыми. [ 32 ] Маленькая морская рыба-еж обитает среди игл таких ежей, как Diadema ; молодые особи питаются педицелляриями и сферидиями, взрослые самцы выбирают трубчатые ножки, а взрослые самки отходят, чтобы питаться яйцами креветок и моллюсками. [ 33 ]
Морские ежи — один из любимых продуктов многих омаров , крабов , спинорогов , калифорнийских овчарок , каланов и волчьих угрей (которые специализируются на морских ежах). Все эти животные обладают особыми приспособлениями (зубы, клешни, когти) и силой, позволяющей им преодолевать превосходные защитные свойства морских ежей. Оставленные без внимания хищников, ежи опустошают окружающую среду, создавая то, что биологи называют бесплодным ежом , лишенным макроводорослей и связанной с ними фауны . [ 34 ] Морские ежи пасутся на нижних стеблях водорослей, в результате чего водоросли уносятся и погибают. Утрата среды обитания и питательных веществ, обеспечиваемых лесами водорослей, приводит к глубоким каскадным воздействиям на морскую экосистему. Морские выдры вернулись в Британскую Колумбию , что значительно улучшило здоровье прибрежной экосистемы. [ 35 ]
-
 Волчий угорь — узкоспециализированный хищник морских ежей.
Волчий угорь — узкоспециализированный хищник морских ежей. -
 Морская выдра питается фиолетовым морским ежом .
Морская выдра питается фиолетовым морским ежом . -
 Мраморный каменный краб ( Carpilius convexus ) атакует морского ежа-грифельного карандаша ( Heterocentrotus mamillatus )
Мраморный каменный краб ( Carpilius convexus ) атакует морского ежа-грифельного карандаша ( Heterocentrotus mamillatus ) -
 Губан . доедает остатки поврежденной решетки Tripneustes
Губан . доедает остатки поврежденной решетки Tripneustes




Защита от хищников
[ редактировать ]
Шипы , длинные и острые у некоторых видов, защищают ежа от хищников . Некоторые тропические морские ежи, такие как Diadematidae , Echinothuriidae и Toxopneustidae, имеют ядовитые шипы. Другие существа также используют эту защиту; крабы, креветки и другие организмы прячутся среди шипов и часто принимают окраску своего хозяина. Некоторые крабы семейства Dorippidae носят в клешнях морских ежей, морские звезды, острые раковины или другие защитные предметы. [ 36 ]
Педицеллярии [ 37 ] являются хорошим средством защиты от эктопаразитов, но не панацеей, поскольку некоторые из них фактически ими питаются. [ 38 ] Гемальная система защищает от эндопаразитов. [ 39 ]
Ареал и среда обитания
[ редактировать ]Морские ежи обитают в большинстве мест обитания на морском дне, начиная с литорали и вниз, на чрезвычайно широком диапазоне глубин. [ 40 ] Некоторые виды, например Cidaris abyssicola , могут обитать на глубине нескольких километров. Многие роды встречаются только в абиссальной зоне , в том числе множество цидароидов , большинство родов семейства Echinothuriidae и «кактусовые ежи» Dermechinus . Одно из самых глубокоживущих семейств — Pourtalesiidae . [ 41 ] странные морские ежи неправильной формы в форме бутылки, которые обитают только в зоне хадала и были собраны на глубине 6850 метров под поверхностью Зондской впадины . [ 42 ] Тем не менее, это делает морского ежа классом иглокожих, живущих на меньшей глубине, по сравнению с хрупкими звездами , морскими звездами и криноидеями , которые остаются в изобилии на глубине ниже 8000 м (26 250 футов), а также морскими огурцами , которые были зарегистрированы на глубине 10 687 м (35 100 футов). [ 42 ]
Плотность населения варьируется в зависимости от среды обитания: на бесплодных территориях популяция более плотная, чем на зарослях водорослей . [ 43 ] [ 44 ] Даже в этих бесплодных районах наибольшая плотность наблюдается на мелководье. Популяции обычно обитают на более глубокой воде, если присутствует волновое воздействие. [ 44 ] Плотность снижается зимой, когда штормы заставляют их искать защиту в трещинах и вокруг более крупных подводных сооружений. [ 44 ] Галечный еж ( Colobocentrotus atratus ), обитающий на открытых береговых линиях, особенно устойчив к воздействию волн. Это один из немногих морских ежей, способных выжить в течение многих часов без воды. [ 45 ]
Морских ежей можно встретить во всех климатических условиях: от теплых морей до полярных океанов. [ 40 ] личинки полярного морского ежа Sterechinus neumayeri используют энергию в метаболических процессах в двадцать пять раз эффективнее, чем большинство других организмов. Было обнаружено, что [ 46 ] Несмотря на свое присутствие почти во всех морских экосистемах, большинство видов обитает на побережьях умеренного и тропического поясов, от поверхности до нескольких десятков метров в глубину, вблизи фотосинтетической пищи. источников [ 40 ]
-
 Пурпурные морские ежи во время отлива в Калифорнии . Они роют углубление в скале, чтобы днем прятаться от хищников.
Пурпурные морские ежи во время отлива в Калифорнии . Они роют углубление в скале, чтобы днем прятаться от хищников. -
 Dermechinus horridus , абиссальный вид, обитающий на глубине тысяч метров.
Dermechinus horridus , абиссальный вид, обитающий на глубине тысяч метров. -
 Антарктический морской еж ( Sterechinus neumayeri ) обитает в замерзших морях.
Антарктический морской еж ( Sterechinus neumayeri ) обитает в замерзших морях. -
 Форма галечного ежа позволяет ему держаться на избитых волнами скалах.
Форма галечного ежа позволяет ему держаться на избитых волнами скалах.




Эволюция
[ редактировать ]Ископаемая история
[ редактировать ]
Самые ранние окаменелости ежовых рыб относятся к среднему ордовику ( около 465 млн лет назад ). [ 47 ] [ 48 ] [ 49 ] Существует богатая летопись окаменелостей, их твердые образцы состоят из пластин кальцита , сохранившихся в породах всех периодов с тех пор. [ 50 ] У некоторых хорошо сохранившихся экземпляров имеются шипы, но обычно остается только панцирь. Изолированные шипы часто встречаются в окаменелостях. Некоторые юрские и меловые Cidaroida имели очень тяжелые булавовидные шипы. [ 51 ]
Большинство ископаемых морских ежей палеозойской эры являются неполными и состоят из изолированных шипов и небольших скоплений разбросанных пластинок раздробленных особей, в основном в девонских и каменноугольных породах. Мелководные известняки ордовика и славятся ежами силура Эстонии . [ 52 ] Палеозойские ежи, вероятно, обитали в относительно спокойных водах. Из-за своего тонкого тела они наверняка не выжили бы в волнообразных прибрежных водах, населенных многими современными ежами. [ 52 ] В конце палеозойской эры ежи практически вымерли, и из пермского периода было известно всего шесть видов. Только две линии пережили массовое вымирание этого периода и дошли до триаса : род Miocidaris , который дал начало современным cidaroida (карандашным ежам), и предок, который дал начало эвхиноидам . К верхнему триасу их численность вновь увеличилась. Цидароиды очень мало изменились со времен позднего триаса и являются единственной сохранившейся группой палеозойских ежей. [ 52 ]
Эвхиноиды диверсифицировались в новые линии в юрском и меловом возникли первые неправильные ежи ( Atelostomata ). периодах, и из них в ранней юре [ 53 ]
Некоторые морские ежи, такие как Микрастер из мела мелового периода, служат зональными или индексными окаменелостями. Поскольку они многочисленны и быстро развивались, они позволяют геологам датировать окружающие породы. [ 54 ]
В палеогеновом и неогеновом периодах ( около 66–2,6 млн лет назад) возникли песчаные доллары (Clypeasteroida). Их характерные уплощенные раковины и крошечные шипы были приспособлены к жизни на рыхлом песке или под ним на мелководье, и они широко распространены в виде окаменелостей в известняках и песчаниках южной Европы. [ 52 ]






Филогения
[ редактировать ]Внешний
[ редактировать ]Ехиноиды — вторичноротые животные, как и хордовые . Анализ 219 генов всех классов иглокожих, проведенный в 2014 году, дает следующее филогенетическое древо . [ 55 ] Приблизительные даты разветвления основных клад указаны в миллионах лет назад (млн лет назад).
| Искатель |
| ||||||||||||





Внутренний
[ редактировать ]Филогения морских ежей следующая: [ 56 ] [ 57 ]
| Эхинодея | |
| 450 млн лет назад |

Филогенетическое исследование 2022 года представляет другую топологию филогенетического дерева Euechinoidea . Irregularia — сестринская группа эхинацеи (в том числе Salenioida ), образующая общую кладу Carinacea , базальные группы Aspidodiadematoida , Diadematoida , Echinothurioida , Micropygoida и Pedinoida входят в общую базальную кладу Aulodonta . [ 58 ]
Отношение к людям
[ редактировать ]Травмы
[ редактировать ]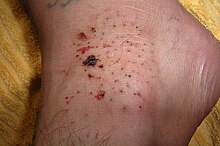
Травмы морского ежа — это колотые раны, нанесенные хрупкими и хрупкими шипами животного. [ 59 ] Они являются частым источником травм для океанских пловцов, особенно вдоль прибрежной поверхности, где присутствуют кораллы и неподвижные морские ежи. Их укусы различаются по степени тяжести в зависимости от вида. Их шипы могут быть ядовитыми или вызывать инфекцию. Также могут возникнуть гранулема и окрашивание кожи естественным красителем внутри морского ежа. Проблемы с дыханием могут указывать на серьезную реакцию морского ежа на токсины. [ 60 ] Они наносят болезненную рану, когда проникают в кожу человека, но сами по себе не опасны, если их сразу полностью удалить; если оставить его на коже, могут возникнуть дальнейшие проблемы. [ 61 ]
Наука
[ редактировать ]Морские ежи являются традиционными модельными организмами в биологии развития . Это использование зародилось в 1800-х годах, когда их эмбриональное развитие стало легко наблюдать с помощью микроскопии. Прозрачность яиц ежа позволила использовать их для наблюдения за тем, как сперматозоиды на самом деле оплодотворяют яйцеклетки . [ 62 ] Их продолжают использовать для исследований эмбрионов, поскольку пренатальное развитие продолжает требовать тестирования на смертельные заболевания. Морские ежи используются в исследованиях долголетия для сравнения молодых и старых представителей этого вида, особенно на предмет их способности регенерировать ткани по мере необходимости. [ 63 ] Ученые из Университета Сент-Эндрюс обнаружили у морских ежей генетическую последовательность, область «2А», которая, как ранее считалось, принадлежала только таким вирусам, как вирус ящура . [ 64 ] Совсем недавно Эрик Х. Дэвидсон и Рой Джон Бриттен выступили за использование ежей в качестве модельного организма из-за их легкой доступности, высокой плодовитости и долгой продолжительности жизни. Помимо эмбриологии , ежи предоставляют возможность исследовать цис-регуляторные элементы . [ 65 ] Океанография проявила интерес к мониторингу здоровья ежей и их популяций как способа оценки общего закисления океана . [ 66 ] температуры и экологические последствия.
Эволюционное расположение организма и уникальная эмбриология с пятикратной симметрией были основными аргументами в предложении секвенировать его геном . Важно отметить, что ежи действуют как ближайшие живые существа по отношению к хордовым и, таким образом, представляют интерес, поскольку могут пролить свет на эволюцию позвоночных . [ 67 ] Геном Strongylocentrotus purpuratus был завершен в 2006 году и установил гомологию между морским ежом и генами, связанными с иммунной системой позвоночных . Морские ежи кодируют по меньшей мере 222 гена Toll-подобных рецепторов и более 200 генов, связанных с семейством Nod-подобных рецепторов, обнаруженных у позвоночных. [ 68 ] Это увеличивает его полезность в качестве ценного модельного организма для изучения эволюции врожденного иммунитета . Секвенирование также показало, что, хотя считалось, что некоторые гены доступны только позвоночным, существовали также инновации, которые ранее никогда не наблюдались за пределами классификации хордовых, такие как иммунные факторы транскрипции PU.1 и SPIB . [ 67 ]
В качестве еды
[ редактировать ]
Гонады . как самцов, так и самок морских ежей, иногда эвфемизированные как «икра» морского ежа или «кораллы» [ 69 ] являются кулинарными деликатесами во многих частях мира, особенно в Японии. [ 70 ] [ 71 ] [ 72 ] В Японии морской еж известен как уни ( うに ) , а его гонады (единственные мясистые съедобные части животного) могут стоить до 40 000 йен (360 долларов США) за килограмм; [ 73 ] их подают сырыми в виде сашими или в суши , с соевым соусом и васаби . Япония импортирует большие объемы продукции из США, Южной Кореи и других производителей. Япония потребляет 50 000 тонн ежегодно, что составляет более 80% мирового производства. [ 74 ] Спрос Японии на морских ежей вызвал обеспокоенность по поводу чрезмерного вылова рыбы. [ 75 ]
Морских ежей обычно едят фаршированными рисом в традиционном блюде око-око среди народа сама-баджау на Филиппинах . [ 76 ] Когда-то их добывали прибрежные малайские общины Сингапура, которые называли их джани . [ 77 ] В Новой Зеландии Evechinus chromoticus , известный как кина на языке маори , представляет собой деликатес, который традиционно едят сырым. Хотя новозеландские рыбаки хотели бы экспортировать их в Японию, их качество слишком варьируется. [ 78 ]
В средиземноморской кухне Paracentrotus lividus часто едят сырым или с лимоном. [ 79 ] и известен как риччи в итальянском меню, где его иногда используют в соусах для пасты. Им также можно ароматизировать омлеты , яичницу , уху , [ 80 ] майонез , соус бешамель для тарталеток, [ 81 ] булли для суфле , [ 82 ] или голландский соус, чтобы приготовить рыбный соус. [ 83 ]
На тихоокеанском побережье Северной Америки Strongylocentrotus franciscanus получил высокую оценку Юэлла Гиббонса ; Strongylocentrotus purpuratus также едят. [ 70 ] коренные американцы в Калифорнии едят морских ежей. Известно, что [ 84 ] Побережье Южной Калифорнии известно как источник высококачественной рыбы : дайверы собирают морских ежей из зарослей на глубине до 24 м/80 футов. [ 85 ] По состоянию на 2013 год штат ограничивал эту практику 300 лицензиями ныряльщиков морских ежей. [ 85 ] Хотя съедобный Strongylocentrotus droebachiensis встречается в Северной Атлантике, его едят редко. Тем не менее, морские ежи (называемые уутук на Алутик ) обычно едят коренное население Аляски вокруг острова Кадьяк . Его обычно экспортируют, в основном в Японию . [ 86 ] В Вест-Индии грифельных ежей . едят [ 70 ]
В чилийской кухне его подают сырым с лимоном, луком и оливковым маслом.
-
 Японский уни-дон , или миска риса с икрой морского ежа.
Японский уни-дон , или миска риса с икрой морского ежа. -
 Японские нигиридзуси с икрой морского ежа
Японские нигиридзуси с икрой морского ежа -
 Икра морского ежа ( уни ) сашими
Икра морского ежа ( уни ) сашими -
 Жареный рис с морским ежом (海胆, хйидан ), подается в Китае.
Жареный рис с морским ежом (海胆, хйидан ), подается в Китае.




Аквариум
[ редактировать ]
Некоторые виды морских ежей, такие как грифельный еж ( Eucidaris tribuloides ), обычно продаются в аквариумных магазинах. Некоторые виды эффективны в борьбе с нитчатыми водорослями и являются хорошим дополнением к аквариуму с беспозвоночными . [ 88 ]
Фольклор
[ редактировать ]Народная традиция Дании и южной Англии считала окаменелости морских ежей молниями, способными отразить вред от молнии или колдовства, как апотропный символ . [ 89 ] Другая версия предполагала, что это были окаменелые яйца змей, способные защитить от болезней сердца и печени, ядов и ранений в бою, и, соответственно, их носили в качестве амулетов . Согласно легенде, они были созданы по волшебству из пены, образуемой змеями в середине лета. [ 90 ]
Пояснительные примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ «Сеть разнообразия животных - Echinoidea» . Зоологический музей Мичиганского университета . Проверено 26 августа 2012 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Руперт, Эдвард Э.; Фокс, Ричард С.; Барнс, Роберт Д. (2004). Зоология беспозвоночных: функционально-эволюционный подход (7-е изд.). Бельмонт, Калифорния: Томсон-Брукс/Коул. стр. 896–906 . ISBN 0030259827 . ОСЛК 53021401 .
- ^ Кро, А.; Ханссон, Х. (2013). « Ехинодея (Леске, 1778)» . ВОРМС . Всемирный реестр морских видов . Проверено 4 января 2014 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м Барнс, Роберт Д. (1982). Зоология беспозвоночных . Филадельфия, Пенсильвания: Холт-Сондерс Интернэшнл. стр. 961–981. ISBN 0-03-056747-5 .
- ^ Гилл, Майкл. «Таксономическая этимология EEOB 111» . Проверено 13 марта 2018 г.
- ^ Райт, Энн. 1851. Наблюдательный глаз, или Письма к детям о трех низших отделах животной жизни. Лондон: Джаррольд и сыновья, с. 107.
- ^ Сойер, Алексис . 1853. Пантрофеон, или История еды и ее приготовления: с древнейших времен мира. Бостон: Тикнор, Рид и Филдс, с. 245.
- ^ «еж (сущ.)» . Интернет-словарь этимологии . Проверено 13 марта 2018 г.
- ^ Стачан и Рид, Молекулярная генетика человека , «Что делает нас людьми», стр. 381 .
- ^ «Справочник ежовых рыб - Музей естественной истории» . www.nhm.ac.uk. Музей естественной истории, Великобритания . Проверено 28 декабря 2022 г.
- ^ «Справочник ежовых рыб - Музей естественной истории» . www.nhm.ac.uk. Музей естественной истории, Великобритания . Проверено 28 декабря 2022 г.
- ^ «Морские ежи открывают многообещающую альтернативу улавливанию углерода» . Гизмаг. 4 февраля 2013 года . Проверено 5 февраля 2013 г.
- ^ Казуя Ёсимура, Томоаки Икетани и Тацуо Мотокава, «Отдают ли обычные морские ежи предпочтение, в какой части тела они ориентируются вперед при ходьбе?», Морская биология , том. 159, № 5, 2012, с. 959–965.
- ^ Будуреск, Чарльз Ф.; Верлак, Марк (2006). «13: Экология Paracentrotus lividus » . В Лоуренсе, Джон М. (ред.). Съедобные морские ежи: биология и экология . Эльзевир. п. 243. ИСБН 978-0-08-046558-6 .
- ^ «Апикальный диск и перипрокт» . Музей естественной истории, Лондон . Проверено 2 ноября 2019 г.
- ^ Коготь, вдохновленный пастью морских ежей, может зачерпывать марсианскую почву.
- ^ Вулсиаду, Элени; Чинтироглу, Харитон (2008). «Фонарь Аристотеля у иглокожих: древняя загадка» (PDF) . Тетради по морской биологии . 49 (3). Биологическая станция Роскофф: 299–302.
- ^ Чой, Чарльз К. (29 декабря 2010 г.). «У морских ежей, жующих камни, зубы самозатачивающиеся» . Национальные географические новости . Архивировано из оригинала 31 декабря 2010 года . Проверено 12 ноября 2017 г.
- ^ Найт, К. (2009). «Морские ежи используют все тело как глаз» . Журнал экспериментальной биологии . 213 (2): i – ii. дои : 10.1242/jeb.041715 .
- Чарльз К. Чой (28 декабря 2009 г.). «Тело морского ежа — это один большой глаз» . LiveScience (Пресс-релиз).
- ^ Гайтан-Эпития, доктор юридических наук; Санчес, Р.; Брюнинг, П.; Карденас, Л. (2016). «Функциональное понимание транскриптома семенников съедобного морского ежа Loxechinus albus» . Научные отчеты . 6 : 36516. Бибкод : 2016NatSR...636516G . дои : 10.1038/srep36516 . ПМК 5090362 . ПМИД 27805042 .
- ^ А. Гайон, А. Скудери; Д. Пеллегрини; Д. Сартори (2013). «Воздействие мышьяка влияет на развитие эмбриона морского ежа Paracentrotus lividus (Ламарк, 1816)». Бюллетень загрязнения окружающей среды и токсикологии . 39 (2): 124–8. дои : 10.3109/01480545.2015.1041602 . ПМИД 25945412 . S2CID 207437380 .
- ^ Коминами, Тецуя; Таката, Хироми (2004). «Гаструляция эмбриона морского ежа: модельная система для анализа морфогенеза однослойного эпителия». Развитие, рост и дифференциация . 46 (4): 309–26. дои : 10.1111/j.1440-169x.2004.00755.x . ПМИД 15367199 . S2CID 23988213 .
- ^ Шук, Д; Келлер, Р. (2003). «Механизмы, механика и функции эпителиально-мезенхимальных переходов в раннем развитии» . Механизмы развития . 120 (11): 1351–83. дои : 10.1016/j.mod.2003.06.005 . ПМИД 14623443 . S2CID 15509972 . ; Катов, Хидеки; Солурш, Майкл (1980). «Ультраструктура ингрессии первичных мезенхимных клеток у морского ежа Lytechinus pictus». Журнал экспериментальной зоологии . 213 (2): 231–246. Бибкод : 1980JEZ...213..231K . дои : 10.1002/jez.1402130211 . ; Балинский, Б.И. (1959). «Электромикроскопическое исследование механизмов адгезии клеток бластулы и гаструлы морского ежа». Экспериментальные исследования клеток . 16 (2): 429–33. дои : 10.1016/0014-4827(59)90275-7 . ПМИД 13653007 . ; Герцлер, Польша; Макклей, доктор медицинских наук (1999). «альфаSU2, эпителиальный интегрин, который связывает ламинин в эмбрионе морского ежа» . Биология развития . 207 (1): 1–13. дои : 10.1006/dbio.1998.9165 . ПМИД 10049560 . ; Финк, Р.Д.; Макклей, доктор медицинских наук (1985). «Три изменения в распознавании клеток сопровождают проникновение первичных мезенхимных клеток морского ежа». Биология развития . 107 (1): 66–74. дои : 10.1016/0012-1606(85)90376-8 . ПМИД 2578117 . ; Бурдсал, Калифорния; Альегро, MC; Макклей, доктор медицинских наук (1991). «Тканеспецифические временные изменения в адгезии клеток к эхинонектину у эмбриона морского ежа». Биология развития . 144 (2): 327–34. дои : 10.1016/0012-1606(91)90425-3 . ПМИД 1707016 . ; Миллер-младший; Макклей, доктор медицинских наук (1997). «Характеристика роли кадгерина в регулировании адгезии клеток во время развития морских ежей» . Биология развития . 192 (2): 323–39. дои : 10.1006/dbio.1997.8740 . ПМИД 9441671 . ; Миллер-младший; Макклей, доктор медицинских наук (1997). «Изменения в структуре бета-катенина, связанного с слипчивыми соединениями, сопровождают морфогенез эмбриона морского ежа» . Биология развития . 192 (2): 310–22. дои : 10.1006/dbio.1997.8739 . ПМИД 9441670 . ; Анстром, Дж. А. (1989). «Клетки первичной мезенхимы морского ежа: ингрессия происходит независимо от микротрубочек». Биология развития . 131 (1): 269–75. дои : 10.1016/S0012-1606(89)80058-2 . ПМИД 2562830 . ; Анстром, Дж. А. (1992). «Микрофиламенты, изменения формы клеток и образование первичной мезенхимы у эмбрионов морских ежей». Журнал экспериментальной зоологии . 264 (3): 312–22. дои : 10.1002/jez.1402640310 . ПМИД 1358997 .
- ^ Ниссен, Сайлас Бойе; Ронхильд, Стивен; Трусина, Ала; Снеппен, Ким (27 ноября 2018 г.). «Теоретический инструмент, соединяющий полярности клеток с развитием устойчивой морфологии» . электронная жизнь . 7 : е38407. дои : 10.7554/eLife.38407 . ПМК 6286147 . ПМИД 30477635 .
- ^ Уорнер, Джейкоб Ф.; Лайонс, Дейдра К.; Макклей, Дэвид Р. (2012). «Левая-правая асимметрия в эмбрионе морского ежа: BMP и асимметричное происхождение взрослой особи» . ПЛОС Биология . 10 (10): е1001404. дои : 10.1371/journal.pbio.1001404 . ПМЦ 3467244 . ПМИД 23055829 .
- ^ Уорли, Алиса (2001). « Стронгилоцентротус пурпуратус » . Сеть разнообразия животных . Проверено 5 декабря 2016 г.
- ^ «Красный морской еж» .
- ^ Томас А. Эберт и Джон Р. Саутон (2003). «Красные морские ежи ( Strongylocentrotus franciscanus ) могут жить более 100 лет: подтверждение с помощью атомной бомбы 14 углерод» (PDF) . Fishery Bulletin . 101 (4): 915–922.
- ^ Баумиллер, Томаш К. (2008). «Экологическая морфология криноидей». Ежегодный обзор наук о Земле и планетах . 36 : 221–49. Бибкод : 2008AREPS..36..221B . doi : 10.1146/annurev.earth.36.031207.124116 .
- ^ Лоуренс, Джон М. (2006). Съедобные морские ежи: биология и экология . Эльзевир. стр. 167–168. ISBN 978-0-08-046558-6 .
- ^ Жангу, Мишель (1987). «Болезни иглокожих. I. Возбудители микроорганизмов и протистанов» . Болезни водных организмов . 2 : 147–162. дои : 10.3354/dao002147 .
- ^ «Защита – шипы» . Справочник Ежиных . Музей естественной истории .
- ^ Сакашита, Хироко (1992). «Половой диморфизм и пищевые привычки прилипалы Diademichthys lineatus и ее зависимость от морского ежа-хозяина». Экологическая биология рыб . 34 (1): 95–101. Бибкод : 1992EnvBF..34...95S . дои : 10.1007/BF00004787 . S2CID 32656986 .
- ^ Терборг, Джон; Эстес, Джеймс А. (2013). Трофические каскады: хищники, жертвы и меняющаяся динамика природы . Остров Пресс. п. 38. ISBN 978-1-59726-819-6 .
- ^ «Водные виды, находящиеся под угрозой – Профиль видов – Морская выдра» . Рыбное хозяйство и океаны Канады . Архивировано из оригинала 23 января 2008 г. Проверено 29 ноября 2007 г.
- ^ Тиль, Мартин; Уотлинг, Лес (2015). Образ жизни и биология питания . Издательство Оксфордского университета. стр. 200–202. ISBN 978-0-19-979702-8 .
- ^ «Защита – педицеллярии» . Справочник Ежиных . Музей естественной истории .
- ^ Хироко Сакашита, «Половой диморфизм и пищевые привычки прилипалы Diademichthys lineatus и ее зависимость от морского ежа-хозяина», Экологическая биология рыб, том. 34, № 1, 1994, с. 95–101
- ^ Жангу, М. (1984). «Болезни иглокожих» (PDF) . Морские исследования Гельголанда . 37 (1–4): 207–216. Бибкод : 1984HM.....37..207J . дои : 10.1007/BF01989305 . S2CID 21863649 . Проверено 23 марта 2018 г.
- ^ Перейти обратно: а б с Кро, Андреас (2010). «Филогения и классификация постпалеозойских морских ежей» . Журнал систематической палеонтологии . 8 (2): 147–212. Бибкод : 2010JSPal...8..147K . дои : 10.1080/14772011003603556 . .
- ^ Ма, Кристофер (12 апреля 2011 г.). «Размеры и виды самых странных из странных: глубоководные ежи-пурталезииды» . Эчиноблог . .
- ^ Перейти обратно: а б Ма, Кристофер (8 апреля 2014 г.). «Какие самые глубокие из известных иглокожих?» . Эчиноблог . Проверено 22 марта 2018 г. .
- ^ Мэттисон, Дж. Э.; Трент, доктор юридических наук; Шанкс, Алабама; Акин, ТБ; Пирс, Дж. С. (1977). «Перемещение и пищевая активность красных морских ежей ( Strongylocentrotus franciscanus ), прилегающих к лесу из водорослей». Морская биология . 39 (1): 25–30. Бибкод : 1977МарБи..39...25М . дои : 10.1007/BF00395589 . S2CID 84338735 .
- ^ Перейти обратно: а б с Конар, Бренда (январь 2000 г.). «Влияние среды обитания на популяции морских ежей» . Написано в Сент-Пит-Бич, Флорида. В Холлоке, Памела; Френч, Ллин (ред.). Дайвинг ради науки: материалы 20-го ежегодного научного симпозиума по дайвингу, 11–15 октября 2000 г. Нахант, Массачусетс: Американская академия подводных наук. OCLC 45371343 . Архивировано из оригинала 15 апреля 2013 года . Проверено 7 января 2011 г.
- ^ КрисМ (21 апреля 2008 г.). «Эчиноблог» . echinoblog.blogspot.com .
- ^ Антарктический морской еж демонстрирует удивительную энергоэффективность в условиях глубокой заморозки природы, 15 марта 2001 г., Университет штата Делавэр. Проверено 22 марта 2018 г.
- ^ Боттинг, Джозеф П.; Мьюир, Люси А. (март 2012 г.). «Фауна и экология слоя голотурий, Лландриндод, Уэльс, Великобритания (дарривилий, средний ордовик), и древнейшие сочлененные голотурии» . Электронная палеонтология . 15 (1): 1–28. дои : 10.26879/272 . S2CID 55716313 .
- ^ Томпсон, Джеффри Р.; Коттон, Лаура Дж.; Кандела, Ив; Кучер, Манфред; Райх, Майк; Боттьер, Дэвид Дж. (14 апреля 2022 г.). «Ордовикская диверсификация морских ежей: систематика Bothriocidaroida (Echinodermata: Echinoidea)» . Журнал систематической палеонтологии . 19 (20): 1395–1448. дои : 10.1080/14772019.2022.2042408 . S2CID 248192052 . Проверено 29 октября 2022 г.
- ^ «Ехиноиды» . Британская геологическая служба. 2017 . Проверено 14 марта 2018 г.
- ^ «Справочник ежовых рыб | Введение» . Музей естественной истории . Проверено 16 марта 2018 г.
- ^ «Справочник ежовых рыб | Шипы» . Музей естественной истории . Проверено 16 марта 2018 г.
- ^ Перейти обратно: а б с д Киркалди, Дж. Ф. (1967). Окаменелости в цвете . Лондон: Бландфорд Пресс. стр. 161–163.
- ^ Шульц, Heinke AG (2015). Echinoidea: с пентамерной симметрией . Вальтер де Грюйтер. стр. 36 и далее, раздел 2.4. ISBN 978-3-11-038601-1 .
- ^ Уэллс, Х.Г. ; Хаксли, Джулиан ; Уэллс, врач общей практики (1931). Наука жизни . стр. 346–348.
- ^ Телфорд, MJ; Лоу, CJ; Кэмерон, CB; Ортега-Мартинес, О.; Аронович, Дж.; Оливери, П.; Копли, Р.Р. (2014). «Филогеномный анализ отношений классов иглокожих поддерживает Asterozoa» . Труды Королевского общества B: Биологические науки . 281 (1786): 20140479. doi : 10.1098/rspb.2014.0479 . ПМК 4046411 . ПМИД 24850925 .
- ^ Планета, Пол Дж.; Зиглер, Александр; Шредер, Лейф; Огуррек, Мальте; Фабер, Корнелиус; Стах, Томас (2012). «Эволюция новой конструкции мышц у морских ежей (Echinodermata: Echinoidea)» . ПЛОС ОДИН . 7 (5): e37520. Бибкод : 2012PLoSO...737520Z . дои : 10.1371/journal.pone.0037520 . ПМЦ 3356314 . ПМИД 22624043 .
- ^ Кро, Андреас; Смит, Эндрю Б. (2010). «Филогения и классификация постпалеозойских морских ежей» . Журнал систематической палеонтологии . 8 (2): 147–212. Бибкод : 2010JSPal...8..147K . дои : 10.1080/14772011003603556 .
- ^ Кох, Николас Монджардино; Томпсон, Джеффри Р.; Хили, Эйвери С; МакКоуин, Марина Ф; Армстронг, Фрэнсис; Коппард, Саймон Э; Агилера, Фелипе; Бронштейн, Омри; Кро, Андреас; Муи, Рич; Роуз, Грег В. (22 марта 2022 г.). «Филогеномный анализ разнообразия морских ежей побуждает к переоценке их окаменелостей» . электронная жизнь . 11 : е72460. дои : 10.7554/eLife.72460 . ПМК 8940180 . ПМИД 35315317 .
- ^ Джеймс, Уильям Д.; Бергер, Тимоти Г.; и др. (2006). Болезни кожи Эндрюса: клиническая дерматология . Сондерс Эльзевир. п. 431. ИСБН 0-7216-2921-0 .
- ^ Галлахер, Скотт А. «Отравление иглокожих» . Электронная медицина . Проверено 12 октября 2010 г.
- ^ Мэтью Д. Гаргус; Дэвид К. Морохаши (2012). «Средство от морского ежа, охлаждающее позвоночник» . Медицинский журнал Новой Англии . 30 (19): 1867–1868. дои : 10.1056/NEJMc1209382 . ПМИД 23134402 .
- ^ «Взгляд морского ежа» . Станция визуализации микроскопа . Эксплораториум. Архивировано из оригинала 12 марта 2017 г. Проверено 07 марта 2018 г.
- ^ Боднар, Андреа Г.; Коффман, Джеймс А. (01 августа 2016 г.). «Поддержание регенерации соматических тканей с возрастом у коротко- и долгоживущих видов морских ежей» . Стареющая клетка . 15 (4): 778–787. дои : 10.1111/acel.12487 . ISSN 1474-9726 . ПМЦ 4933669 . ПМИД 27095483 .
- ^ Роулстон, К.; Люк, Джорджия; де Фелипе, П.; Руан, Л.; Коуп, Дж.; Николсон, Дж.; Суходуб А.; Тилснер, Дж.; Райан, доктор медицины (2016). « 2A-подобные сигнальные последовательности, опосредующие трансляционное перекодирование: новая форма нацеливания на два белка» (PDF) . Трафик . 17 (8): 923–39. дои : 10.1111/tra.12411 . ПМЦ 4981915 . ПМИД 27161495 .
- ^ «Проект генома морского ежа» . sugp.caltech.edu . Архивировано из оригинала 20 декабря 2016 г. Проверено 5 декабря 2016 г.
- ^ «Стэнфорд ищет секрет морских ежей, позволяющих выжить при закислении океана | Стэнфордский пресс-релиз» . news.stanford.edu . 08.04.2013. Архивировано из оригинала 20 декабря 2016 г. Проверено 5 декабря 2016 г.
- ^ Перейти обратно: а б Содергрен, Э; Вайншток, генеральный директор; Дэвидсон, Э.Х.; и др. (10 ноября 2006 г.). «Геном морского ежа Strongylocentrotus purpuratus» . Наука . 314 (5801): 941–952. Бибкод : 2006Sci...314..941S . дои : 10.1126/science.1133609 . ПМК 3159423 . ПМИД 17095691 .
- ^ Раст, Дж.П.; Смит, Л.К.; Лоза-Колл, М; Хибино, Т; Литман, Г.В. (2006). «Геномный взгляд на иммунную систему морского ежа» . Наука . 314 (5801): 952–6. Бибкод : 2006Sci...314..952R . дои : 10.1126/science.1134301 . ПМК 3707132 . ПМИД 17095692 .
- ^ Лаура Роджерс-Беннетт, «Экология Strongylocentrotus franciscanus и Strongylocentrotus purpuratus » в книге Джона М. Лоуренса, Съедобные морские ежи: биология и экология , с. 410
- ^ Перейти обратно: а б с Дэвидсон, Алан (2014) Оксфордский справочник по еде . Издательство Оксфордского университета , 3-е издание. стр. 730–731.
- ^ Джон М. Лоуренс, «Кухня из икры морских ежей» в книге Джона М. Лоуренса, Съедобные морские ежи: биология и экология.
- ^ « Восстание морского ежа », Франц Лидз , июль 2014 г., Смитсоновский институт.
- ^ Мейси, Ричард (9 ноября 2004 г.). «Маленькие мальчишки, которые могут потребовать королевскую цену» . Сидней Морнинг Геральд .
- ^ Затыльный, Джейн (6 сентября 2018 г.). «В поисках ежа: кулинарный квест» . Журнал Хакай . Проверено 10 сентября 2018 г.
- ^ «Промысел морских ежей и чрезмерный вылов рыбы», Тематические исследования TED 296 Американского университета. , полный текст
- ^ «Приготовление око-око, деликатеса из самских морских ежей» . Кауман Сама Онлайн . 27 июня 2013 года . Проверено 3 июня 2023 г.
- ^ Хир Джохари (октябрь – декабрь 2021 г.). «Роль собирательства в малайской кухне» . БиблиоАзия . Том. 17, нет. 3. Совет национальной библиотеки, Сингапур. стр. 20–23.
- ^ Васильев, Мэгги (2 марта 2009 г.). «морские ежи» . Те Ара: Энциклопедия Новой Зеландии .
- ^ для Апулии, Италия: Touring Club Italiano, Путеводитель по гастрономической Италии , 1984, стр. 314; для Александрии, Египет: Клаудия Роден , Книга о ближневосточной кухне , стр. 183
- ^ Алан Дэвидсон , Средиземноморские морепродукты , с. 270
- ^ Гастрономический Ларусс [ нужна страница ]
- ^ Курнонский , Кухня и вина Франции , новое издание, 1974, с. 248
- ^ Дэвидсон, Алан (2014) Оксфордский справочник по еде . Издательство Оксфордского университета , 3-е издание. п. 280
- ^ Мартин, Р.Э.; Картер, EP; Флик, Дж.Дж.; Дэвис, LM (2000). Справочник по морской и пресноводной продукции . Тейлор и Фрэнсис. п. 268. ИСБН 978-1-56676-889-4 . Проверено 3 декабря 2014 г.
- ^ Перейти обратно: а б Лам, Фрэнсис (14 марта 2014 г.). «Калифорнийские ныряльщики с морскими ежами, интервью Фрэнсиса Лама» . Приятного аппетита . Проверено 26 марта 2017 г.
- ^ Клейман, Дена (3 октября 1990 г.). «Презираемый дома морской еж штата Мэн — звезда в Японии» . Нью-Йорк Таймс . п. С1.
- ^ «Амулет | LIN-B37563» . Схема переносного антиквариата . Проверено 14 марта 2018 г.
- ^ Таллок, Джон Х. (2008). Ваш первый морской аквариум: все об устройстве морского аквариума, включая кондиционирование, уход, выбор рыб и беспозвоночных и многое другое . Образовательная серия Бэррона. п. 63 . ISBN 978-0-7641-3675-7 .
- ^ Макнамара, Кен (2012). «Коллекционеры доисторических окаменелостей» . Геологическое общество . Проверено 14 марта 2018 г.
- ^ Маррен, Питер ; Мэби, Ричард (2010). Баги Британника . Чатто и Виндус. стр. 469–470. ISBN 978-0-7011-8180-2 .
Внешние ссылки
[ редактировать ]- Ссылка на Всемирный реестр морских видов : Echinoidea Leske, 1778 ( +список видов )
- Проект генома морского ежа
- Ассоциация сборщиков морских ежей – Калифорния , также: (604) 524-0322.
- Справочник ежовых рыб из Музея естественной истории .
- Виртуальный еж в Стэнфорде
- Калифорнийская комиссия по морским ежам
- Знакомство с Echinoidea в UCMP Беркли
Основные промыслового рыболовства группы видов | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Дикий |
|    | ||||||||||||||||||
| Выращенный | ||||||||||||||||||||