Липидный бислой


Липидный бислой (или фосфолипидный бислой ) представляет собой тонкую полярную мембрану, состоящую из двух слоев липидных молекул . Эти мембраны представляют собой плоские листы, образующие сплошной барьер вокруг всех клеток . Клеточные мембраны почти всех организмов и многих вирусов состоят из липидного бислоя, как и ядерная мембрана, окружающая ядро клетки , и мембраны мембраносвязанных органелл клетки. Липидный бислой — это барьер, который удерживает ионы , белки и другие молекулы там, где они необходимы, и предотвращает их диффузию в области, где их не должно быть. Липидные бислои идеально подходят для этой роли, хотя их всего несколько нанометров . ширина составляет [1] потому что они непроницаемы для большинства водорастворимых ( гидрофильных ) молекул. Бислои особенно непроницаемы для ионов, что позволяет клеткам регулировать концентрацию солей и pH путем транспортировки ионов через мембраны с помощью белков, называемых ионными насосами .
Биологические бислои обычно состоят из амфифильных фосфолипидов , которые имеют гидрофильную фосфатную головку и гидрофобный хвост, состоящий из двух цепей жирных кислот. Фосфолипиды с определенными головными группами могут изменять химический состав поверхности бислоя и могут, например, служить сигналами, а также «якорями» для других молекул в мембранах клеток. [2] Как и «головки», «хвосты» липидов также могут влиять на свойства мембраны, например, определяя фазу бислоя . Бислой может переходить в твердое фазовое состояние геля при более низких температурах, но подвергаться фазовому переходу в жидкое состояние при более высоких температурах, и химические свойства липидных хвостов влияют на то, при какой температуре это происходит. Упаковка липидов внутри бислоя также влияет на его механические свойства, в том числе на устойчивость к растяжению и изгибу. Многие из этих свойств были изучены с использованием искусственных «модельных» бислоев, изготовленных в лаборатории. Везикулы , образованные модельными бислоями, также использовались в клинических целях для доставки лекарств.
Структура биологических мембран обычно включает несколько типов молекул помимо фосфолипидов, составляющих бислой. Особенно важным примером в клетках животных является холестерин , который помогает укрепить бислой и уменьшить его проницаемость. Холестерин также помогает регулировать активность некоторых интегральных мембранных белков . Интегральные мембранные белки функционируют, когда включены в липидный бислой, и они прочно удерживаются на липидном бислое с помощью кольцевой липидной оболочки . Поскольку бислои определяют границы клетки и ее отсеков, эти мембранные белки участвуют во многих внутри- и межклеточных процессах передачи сигналов. Определенные виды мембранных белков участвуют в процессе слияния двух бислоев. Это слияние позволяет объединить две отдельные структуры, как при реакции время оплодотворения яйцеклетки акросомной сперматозоидом во или проникновения вируса в клетку. Поскольку липидные бислои хрупкие и невидимые в традиционном микроскопе, их изучение представляет собой сложную задачу. Эксперименты с бислоями часто требуют сложных методов, таких как электронная микроскопия и атомно-силовая микроскопия .
Структура и организация
[ редактировать ]Когда фосфолипиды подвергаются воздействию воды, они самособираются в двухслойный лист с гидрофобными хвостами, направленными к центру листа. Такое расположение приводит к образованию двух «листочков», каждый из которых представляет собой один молекулярный слой. Центр этого бислоя почти не содержит воды и исключает такие молекулы, как сахара или соли, которые растворяются в воде. Процесс сборки и обслуживания обусловлен агрегацией гидрофобных молекул (также называемой гидрофобным эффектом ). Этот сложный процесс включает в себя нековалентные взаимодействия, такие как силы Ван-дер-Ваальса , электростатические и водородные связи .
Анализ поперечного сечения
[ редактировать ]
Липидный бислой очень тонкий по сравнению с его латеральными размерами. Если типичную клетку млекопитающего (диаметром ~10 микрометров) увеличить до размера арбуза (~1 фут/30 см), то липидный бислой, составляющий плазматическую мембрану, будет примерно такой же толщины, как лист офисной бумаги. Несмотря на то, что бислой имеет толщину всего несколько нанометров, он состоит из нескольких различных химических областей по всему поперечному сечению. Эти регионы и их взаимодействие с окружающей водой за последние несколько десятилетий были охарактеризованы с помощью рентгеновской рефлектометрии . [4] рассеяние нейтронов , [5] и методы ядерного магнитного резонанса .
Первая область по обе стороны от бислоя представляет собой гидрофильную головную группу. Эта часть мембраны полностью гидратирована и обычно имеет толщину около 0,8–0,9 нм. В фосфолипидных бислоях фосфатная группа расположена внутри этой гидратированной области, примерно на 0,5 нм за пределами гидрофобного ядра. [6] В некоторых случаях гидратированная область может простираться гораздо дальше, например, в липидах с большой белковой или длинной сахарной цепью, привитой к головке. Одним из распространенных примеров такой модификации в природе является липополисахаридная оболочка на внешней мембране бактерий. [7] Это помогает сохранить водный слой вокруг бактерии и предотвратить обезвоживание.

Рядом с гидратированной областью находится промежуточная область, которая гидратирована лишь частично. Этот пограничный слой имеет толщину примерно 0,3 нм. На этом коротком расстоянии концентрация воды падает с 2М на стороне головной группы до почти нуля на стороне хвоста (сердцевины). [8] [9] Гидрофобное ядро бислоя обычно имеет толщину 3–4 нм, но это значение зависит от длины цепи и химического состава. [4] [10] Толщина ядра также существенно меняется в зависимости от температуры, особенно вблизи фазового перехода. [11]
Асимметрия
[ редактировать ]Во многих встречающихся в природе бислоях состав створок внутренней и внешней мембраны различен. В человеческих эритроцитах внутренний (цитоплазматический) листок состоит в основном из фосфатидилэтаноламина , фосфатидилсерина , фосфатидилинозитола и его фосфорилированных производных. Напротив, внешний (внеклеточный) листок основан на фосфатидилхолине , сфингомиелине и различных гликолипидах. [12] [13] [14] В некоторых случаях эта асимметрия основана на том, где в клетке образуются липиды, и отражает их первоначальную ориентацию. [15] Биологические функции липидной асимметрии изучены недостаточно, хотя ясно, что она используется в нескольких различных ситуациях. Например, когда клетка подвергается апоптозу , фосфатидилсерин, обычно локализованный в цитоплазматическом листке, переносится на внешнюю поверхность: там он распознается макрофагом, который затем активно уничтожает умирающую клетку.
Липидная асимметрия возникает, по крайней мере частично, из-за того, что большинство фосфолипидов синтезируются и первоначально встраиваются во внутренний монослой: те, которые составляют внешний монослой, затем транспортируются из внутреннего монослоя с помощью класса ферментов, называемых флипазами . [16] [17] Другие липиды, такие как сфингомиелин, по-видимому, синтезируются во внешнем листке. Флипазы являются членами более крупного семейства молекул, транспортирующих липиды, которое также включает флопазы, которые переносят липиды в противоположном направлении, и скрамблазы, которые хаотизируют распределение липидов по липидным бислоям (как в апоптотических клетках). В любом случае, как только липидная асимметрия установлена, она обычно не исчезает быстро, поскольку спонтанное переключение липидов между створками происходит чрезвычайно медленно. [18]
Эту асимметрию можно имитировать в лаборатории на моделях двухслойных систем. Определенные типы очень маленьких искусственных пузырьков автоматически становятся слегка асимметричными, хотя механизм возникновения этой асимметрии сильно отличается от механизма в клетках. [19] Используя два разных монослоя при Ленгмюра-Блоджетт. осаждении [20] или сочетание метода Ленгмюра-Блоджетт и отложения разрыва пузырька [21] также возможно синтезировать асимметричный планарный бислой. Эта асимметрия может со временем исчезнуть, поскольку липиды в поддерживаемых бислоях могут быть склонны к переворачиванию. [22] Однако сообщалось, что липидный переворот происходит медленнее, чем холестерин и другие более мелкие молекулы. [23] [24]
Сообщалось, что организация и динамика липидных монослоев в бислое взаимосвязаны. [25] [26] Например, введение препятствий в один монослой может замедлить латеральную диффузию в обоих монослоях. [25] Кроме того, разделение фаз в одном монослое может также вызвать разделение фаз в другом монослое, даже если другой монослой не может разделиться на фазы сам по себе. [26]
Фазы и фазовые переходы
[ редактировать ]
При данной температуре липидный бислой может существовать как в жидкой, так и в гелевой (твердой) фазе. Все липиды имеют характерную температуру, при которой они переходят (плавятся) из гелевой фазы в жидкую. В обеих фазах молекулы липидов не могут переворачиваться через бислой, но в бислоях жидкой фазы данный липид будет меняться местоположением со своим соседом миллионы раз в секунду. Этот обмен случайным блужданием позволяет липидам диффундировать и, таким образом, блуждать по поверхности мембраны. [27] В отличие от бислоев жидкой фазы, липиды в бислое гелевой фазы обладают меньшей подвижностью.
Фазовое поведение липидных бислоев во многом определяется силой притягивающих ван-дер-ваальсовых взаимодействий между соседними липидными молекулами. Липиды с более длинными хвостами имеют большую площадь для взаимодействия, что увеличивает силу этого взаимодействия и, как следствие, снижает подвижность липидов. Таким образом, при данной температуре липид с коротким хвостом будет более жидким, чем идентичный в остальном липид с длинным хвостом. [10] На температуру перехода также может влиять степень ненасыщенности липидных хвостов. Ненасыщенная двойная связь может вызвать перегиб алкановой цепи, нарушая липидную упаковку. Это нарушение создает дополнительное свободное пространство внутри бислоя, что обеспечивает дополнительную гибкость соседних цепей. [10] Пример этого эффекта можно отметить в повседневной жизни, поскольку сливочное масло, содержащее большой процент насыщенных жиров, при комнатной температуре остается твердым, а растительное масло, которое в основном ненасыщенное, является жидким.
Большинство природных мембран представляют собой сложную смесь различных липидных молекул. Если некоторые компоненты при данной температуре являются жидкими, а другие находятся в гелевой фазе, эти две фазы могут сосуществовать в пространственно разделенных областях, подобно айсбергу, плавающему в океане. Это разделение фаз играет решающую роль в биохимических явлениях, поскольку компоненты мембраны, такие как белки, могут разделяться на одну или другую фазу. [28] и, таким образом, быть локально концентрированным или активированным. Одним из особенно важных компонентов многих смешанофазных систем является холестерин , который модулирует проницаемость бислоя, механическую прочность и биохимические взаимодействия.
Химия поверхности
[ редактировать ]Хотя липидные хвосты в первую очередь модулируют фазовое поведение бислоя, именно головная группа определяет химический состав поверхности бислоя. Большинство природных бислоев состоят в основном из фосфолипидов , но сфинголипиды и стерины , такие как холестерин . важными компонентами также являются [29] Из фосфолипидов наиболее распространенной головной группой является фосфатидилхолин (ФХ), на который приходится около половины фосфолипидов в большинстве клеток млекопитающих. [30] ПК представляет собой цвиттер-ионную головную группу, так как он имеет отрицательный заряд фосфатной группы и положительный заряд амина, но, поскольку эти локальные заряды уравновешиваются, суммарный заряд отсутствует.
Другие головные группы также присутствуют в различной степени и могут включать фосфатидилсерин (PS), фосфатидилэтаноламин (PE) и фосфатидилглицерин (PG). Эти альтернативные головные группы часто наделяют определенными биологическими функциями, которые сильно зависят от контекста. Например, наличие ФС на внеклеточной мембране эритроцитов является маркером клеточного апоптоза . [31] тогда как PS в везикулах пластинки роста необходим для зарождения кристаллов гидроксиапатита и последующей минерализации кости. [32] [33] В отличие от ПК, некоторые другие головные группы несут суммарный заряд, который может изменить электростатические взаимодействия малых молекул с бислоем. [34]
Биологические роли
[ редактировать ]Сдерживание и разделение
[ редактировать ]Основная роль липидного бислоя в биологии заключается в отделении водных компартментов от их окружения. Без какого-либо барьера, отделяющего «я» от «чужого», трудно даже определить концепцию организма или жизни. Этот барьер принимает форму липидного бислоя у всех известных форм жизни, за исключением нескольких видов архей , которые используют специально адаптированный липидный монослой. [7] Было даже высказано предположение, что самой первой формой жизни могла быть простая липидная везикула , практически единственной биосинтетической способностью которой было производство большего количества фосфолипидов . [35] Способность липидного бислоя к разделению основана на том факте, что гидрофильные молекулы не могут легко пересекать ядро гидрофобного бислоя, как обсуждается ниже в разделе «Транспорт через бислой». Ядро, митохондрии и хлоропласты имеют два липидных бислоя, тогда как другие субклеточные структуры окружены одним липидным бислоем (например, плазматическая мембрана, эндоплазматическая сеть, аппарат Гольджи и лизосомы). См . Органелла . [36]
У прокариот есть только один липидный бислой — клеточная мембрана (также известная как плазматическая мембрана). У многих прокариот также есть клеточная стенка , но клеточная стенка состоит из белков или длинноцепочечных углеводов , а не из липидов. Напротив, эукариоты имеют ряд органелл, включая ядро , митохондрии , лизосомы и эндоплазматический ретикулум . Все эти субклеточные компартменты окружены одним или несколькими липидными бислоями и вместе обычно составляют большую часть площади бислоя, присутствующей в клетке. печени Например, в гепатоцитах плазматическая мембрана составляет только два процента от общей площади бислоя клетки, тогда как эндоплазматический ретикулум содержит более пятидесяти процентов, а митохондрии - еще тридцать процентов. [37]

Сигнализация
[ редактировать ]Наиболее знакомой формой клеточной передачи сигналов, вероятно, является синаптическая передача , при которой нервный импульс, достигший конца одного нейрона, передается соседнему нейрону посредством высвобождения нейротрансмиттеров . Эта передача становится возможной благодаря действию синаптических везикул , которые внутри клетки нагружены нейротрансмиттерами, которые высвобождаются позже. Эти нагруженные везикулы сливаются с клеточной мембраной на пресинаптическом конце, и их содержимое высвобождается в пространство за пределами клетки. Затем содержимое диффундирует через синапс к постсинаптическому терминалу.
Липидные бислои также участвуют в передаче сигналов, поскольку они служат домом для интегральных мембранных белков . Это чрезвычайно широкий и важный класс биомолекул. Подсчитано, что до трети протеома человека составляют мембранные белки. [38] Некоторые из этих белков связаны с внешней частью клеточной мембраны. Примером этого является белок CD59 , который идентифицирует клетки как «свои» и таким образом подавляет их разрушение иммунной системой. Вирус ВИЧ уклоняется от иммунной системы частично за счет прививки этих белков из мембраны хозяина на свою собственную поверхность. [37] Альтернативно, некоторые мембранные белки проникают через бислой и служат для передачи отдельных сигнальных событий снаружи внутрь клетки. Наиболее распространенным классом этого типа белков является рецептор, связанный с G-белком (GPCR). GPCR отвечают за большую часть способности клетки воспринимать свое окружение, и из-за этой важной роли около 40% всех современных лекарств нацелены на GPCR. [39]
В дополнение к процессам, опосредованным белками и растворами, липидные бислои также могут напрямую участвовать в передаче сигналов. Классическим примером этого является фосфатидилсерином , запускаемый фагоцитоз . В норме фосфатидилсерин распределен в клеточной мембране асимметрично и присутствует только на внутренней стороне. Во время запрограммированной гибели клеток белок, называемый скрамблазой, уравновешивает это распределение, отображая фосфатидилсерин на поверхности внеклеточного бислоя. Присутствие фосфатидилсерина затем запускает фагоцитоз для удаления мертвых или умирающих клеток.
Методы характеристики
[ редактировать ]
Липидный бислой — очень сложную для изучения структуру, поскольку он очень тонкий и хрупкий. Несмотря на эти ограничения, за последние семьдесят лет были разработаны десятки методов, позволяющих исследовать его структуру и функции.
Электрические измерения
[ редактировать ]Электрические измерения — это простой способ охарактеризовать важную функцию бислоя: его способность сегрегировать и предотвращать поток ионов в растворе. Прикладывая напряжение к бислою и измеряя результирующий ток, определяют сопротивление бислоя. Это сопротивление обычно довольно велико (10 8 Ом-см 2 или больше) [40] поскольку гидрофобное ядро непроницаемо для заряженных частиц. Наличие даже нескольких отверстий нанометрового размера приводит к резкому увеличению тока. [41] даже активность одиночных ионных каналов . Чувствительность этой системы такова, что можно разрешить [42]
Флуоресцентная микроскопия
[ редактировать ]
Липидный бислой невозможно увидеть с помощью традиционного микроскопа, поскольку он слишком тонкий, поэтому исследователи часто используют флуоресцентную микроскопию . Образец возбуждается светом одной длины волны и наблюдается при другой, так что будут видны только флуоресцентные молекулы с совпадающим профилем возбуждения и излучения.
Природный липидный бислой не является флуоресцентным, поэтому к некоторым молекулам бислоя необходимо присоединить хотя бы один флуоресцентный краситель. Разрешение обычно ограничено несколькими сотнями нанометров, что, к сожалению, намного больше, чем толщина липидного бислоя.
Электронная микроскопия
[ редактировать ]Электронная микроскопия дает изображение более высокого разрешения. В электронном микроскопе с образцом взаимодействует пучок сфокусированных электронов , а не луч света, как в традиционной микроскопии. В сочетании с методами быстрого замораживания электронная микроскопия также использовалась для изучения механизмов меж- и внутриклеточного транспорта, например, для демонстрации того, что экзоцитозные везикулы являются средством химического высвобождения в синапсах . [43]
Спектроскопия ядерного магнитного резонанса
[ редактировать ]31 Спектроскопия Р- ЯМР (ядерного магнитного резонанса) широко используется для исследования фосфолипидных бислоев и биологических мембран в нативных условиях. Анализ [44] из 31 Спектры P-ЯМР липидов могут предоставить широкий спектр информации об упаковке липидных бислоев, фазовых переходах (гелевая фаза, физиологическая жидкокристаллическая фаза, пульсирующие фазы, недвуслойные фазы), ориентации/динамике головных групп липидов и упругих свойствах чистого липида. двухслойный и в результате связывания белков и других биомолекул.
Атомно-силовая микроскопия
[ редактировать ]

Новым методом изучения липидных бислоев является атомно-силовая микроскопия (АСМ). Вместо использования луча света или частиц очень маленький заостренный кончик сканирует поверхность, вступая в физический контакт с бислоем и перемещаясь по нему, как игла проигрывателя пластинок. АСМ является многообещающим методом, поскольку он потенциально позволяет получать изображения с нанометровым разрешением при комнатной температуре и даже под водой или физиологическим буфером, в условиях, необходимых для естественного поведения бислоев. Используя эту возможность, АСМ использовалась для изучения динамического поведения бислоя, включая образование трансмембранных пор (отверстий). [45] и фазовые переходы в поддерживаемых бислоях. [46] Еще одним преимуществом является то, что АСМ не требует флуоресцентного или изотопного мечения липидов, поскольку кончик зонда механически взаимодействует с поверхностью бислоя. По этой причине одно и то же сканирование может отображать как липиды, так и связанные с ними белки, иногда даже с разрешением одной молекулы. [45] [47] АСМ также может исследовать механическую природу липидных бислоев. [48]
Интерферометрия двойной поляризации
[ редактировать ]Липидные бислои обладают высоким уровнем двойного лучепреломления , при этом показатель преломления в плоскости бислоя отличается от перпендикуляра на целых 0,1 единицы показателя преломления . Это использовалось для характеристики степени порядка и разрушения бислоев с использованием интерферометрии двойной поляризации для понимания механизмов взаимодействия белков.
Квантово-химические расчеты
[ редактировать ]Липидные бислои представляют собой сложные молекулярные системы со многими степенями свободы. Таким образом, атомистическое моделирование мембраны и, в частности, расчеты ее свойств ab initio сложны и требуют больших вычислительных затрат. Недавно были успешно проведены квантово-химические расчеты для оценки дипольных и квадрупольных моментов липидных мембран. [49]
Транспорт через бислой
[ редактировать ]Пассивная диффузия
[ редактировать ]Большинство полярных молекул имеют низкую растворимость в углеводородном ядре липидного бислоя и, как следствие, имеют низкие коэффициенты проницаемости через бислой. Этот эффект особенно выражен для заряженных частиц, которые имеют еще более низкие коэффициенты проницаемости, чем нейтральные полярные молекулы. [50] Анионы обычно имеют более высокую скорость диффузии через бислои, чем катионы . [51] [52] По сравнению с ионами молекулы воды на самом деле обладают относительно большой проницаемостью через бислой, о чем свидетельствует осмотическое набухание . Когда клетка или везикула с высокой внутренней концентрацией соли помещается в раствор с низкой концентрацией соли, она набухает и в конечном итоге лопается. Такой результат не наблюдался бы, если бы вода не могла сравнительно легко проходить через бислой. Аномально большая проницаемость воды через бислои до сих пор до конца не изучена и продолжает оставаться предметом активных дискуссий. [53] Маленькие незаряженные аполярные молекулы диффундируют через липидные бислои на много порядков быстрее, чем ионы или вода. Это относится как к жирам, так и к органическим растворителям, таким как хлороформ и эфир . Независимо от их полярного характера, более крупные молекулы диффундируют через липидные бислои медленнее, чем небольшие молекулы. [54]

Ионные насосы и каналы
[ редактировать ]Два специальных класса белков имеют дело с ионными градиентами, обнаруженными в клеточных и субклеточных мембранах в природных ионных каналах и ионных насосах . И насосы, и каналы представляют собой интегральные мембранные белки , которые проходят через бислой, но их роли совершенно различны. Ионные насосы — это белки, которые создают и поддерживают химические градиенты, используя внешний источник энергии для перемещения ионов против градиента концентрации в область с более высоким химическим потенциалом . Источником энергии может быть АТФ , как и в случае с Na. + -К + АТФаза . Альтернативно, источником энергии может быть другой уже существующий химический градиент, как в случае с Ca. 2+ / Что + антипортер . Именно благодаря действию ионных насосов клетки способны регулировать pH посредством перекачки протонов .
В отличие от ионных насосов, ионные каналы не создают химические градиенты, а скорее рассеивают их, чтобы выполнить работу или отправить сигнал. Вероятно, наиболее знакомым и наиболее изученным примером является потенциал-управляемый Na. + канал , обеспечивающий проведение потенциала действия по нейронам . Все ионные насосы имеют своего рода триггерный или «затворный» механизм. В предыдущем примере это было электрическое смещение, но другие каналы могут быть активированы путем связывания молекулярного агониста или посредством конформационного изменения в другом близлежащем белке. [55]

Эндоцитоз и экзоцитоз
[ редактировать ]Некоторые молекулы или частицы слишком велики или слишком гидрофильны, чтобы пройти через липидный бислой. Другие молекулы могут проходить через бислой, но их необходимо быстро транспортировать в таких больших количествах, что транспорт канального типа становится непрактичным. В обоих случаях эти типы грузов могут перемещаться через клеточную мембрану посредством слияния или отпочкования везикул . Когда везикула образуется внутри клетки и сливается с плазматической мембраной, высвобождая ее содержимое во внеклеточное пространство, этот процесс известен как экзоцитоз. В обратном процессе участок клеточной мембраны будет вдавливаться внутрь и в конечном итоге отщипываться, охватывая часть внеклеточной жидкости для транспортировки ее в клетку. Функционирование эндоцитоза и экзоцитоза зависит от совершенно разных молекулярных механизмов, но эти два процесса тесно связаны и не могут работать друг без друга. Основным механизмом этой взаимозависимости является большое количество задействованного липидного материала. [56] В типичной клетке область бислоя, эквивалентная всей плазматической мембране, проходит цикл эндоцитоза/экзоцитоза примерно за полчаса. [57] Если бы эти два процесса не уравновешивали друг друга, клетка либо раздулась бы до неуправляемых размеров, либо за короткое время полностью истощила бы свою плазматическую мембрану.

Экзоцитоз у прокариот : Мембранный везикулярный экзоцитоз , широко известный как торговля мембранными везикулами , процесс, получивший Нобелевскую премию (2013 год), традиционно считается прерогативой эукариотических клеток. [58] Однако этот миф был развеян открытием того, что нановезикулы, широко известные как везикулы наружной мембраны бактерий , выделяемые грамотрицательными микробами, перемещают бактериальные сигнальные молекулы в клетки-хозяева или клетки-мишени. [59] осуществлять многочисленные процессы в пользу секретирующего микроба, например, при инвазии в клетку-хозяина [60] и взаимодействие микроба с окружающей средой в целом. [61]
Электропорация
[ редактировать ]Электропорация — это быстрое увеличение проницаемости бислоя, вызванное применением сильного искусственного электрического поля через мембрану. Экспериментально электропорация используется для введения гидрофильных молекул в клетки. Это особенно полезный метод для больших сильно заряженных молекул, таких как ДНК , которые никогда не будут пассивно диффундировать через гидрофобное бислойное ядро. [62] По этой причине электропорация является одним из ключевых методов трансфекции , а также бактериальной трансформации . Было даже высказано предположение, что электропорация, возникающая в результате удара молнии , может быть механизмом естественного горизонтального переноса генов . [63]
Это увеличение проницаемости в первую очередь влияет на транспорт ионов и других гидратированных частиц, указывая на то, что механизм заключается в создании заполненных водой отверстий в нанометровом масштабе в мембране. Хотя электропорация и пробой диэлектрика происходят в результате применения электрического поля, механизмы их действия принципиально различны. При пробое диэлектрика материал барьера ионизируется, создавая проводящий путь. Таким образом, материальные изменения имеют химическую природу. Напротив, во время электропорации молекулы липидов не изменяются химически, а просто меняют положение, открывая пору, которая действует как проводящий путь через бислой, поскольку он заполнен водой.
Механика
[ редактировать ]
Липидные бислои представляют собой достаточно крупные структуры, обладающие некоторыми механическими свойствами жидкостей или твердых тел. Модуль сжатия площади K a , модуль изгиба K b и энергия края , можно использовать для их описания. Твердые липидные бислои также имеют модуль сдвига , но, как и любая жидкость, модуль сдвига для жидких бислоев равен нулю. Эти механические свойства влияют на функционирование мембраны. K a и K b влияют на способность белков и малых молекул внедряться в бислой, [64] [65] Было показано, что механические свойства двухслойных слоев изменяют функцию механически активированных ионных каналов. [66] Механические свойства бислоя также определяют, какие виды стресса клетка может выдержать, не разрываясь. Хотя липидные бислои легко сгибаются, большинство из них не могут растянуться более чем на несколько процентов, прежде чем разорвутся. [67]

Как обсуждалось в разделе «Структура и организация», гидрофобное притяжение липидных хвостов в воде является основной силой, удерживающей липидные бислои вместе. Таким образом, модуль упругости бислоя в первую очередь определяется тем, какая дополнительная площадь подвергается воздействию воды, когда молекулы липидов растягиваются. [68] Учитывая такое понимание задействованных сил, неудивительно, что исследования показали, что K a сильно зависит от осмотического давления. [69] но лишь слабо с длиной хвоста и ненасыщенностью. [10] трудно Поскольку задействованные силы настолько малы, экспериментально определить K a . Большинство методов требуют сложной микроскопии и очень чувствительного измерительного оборудования. [48] [70]
В отличие от K a , который является мерой того, сколько энергии необходимо для растяжения бислоя, K b является мерой того, сколько энергии необходимо для изгиба или изгиба бислоя. Формально модуль изгиба определяется как энергия, необходимая для деформации мембраны от ее внутренней кривизны до некоторой другой кривизны. Внутренняя кривизна определяется отношением диаметра головной группы к диаметру хвостовой группы. Для двухвостых PC-липидов это соотношение близко к единице, поэтому внутренняя кривизна почти равна нулю. Если конкретный липид имеет слишком большое отклонение от нулевой внутренней кривизны, он не образует бислой, а вместо этого образует другие фазы, такие как мицеллы или инвертированные мицеллы. Добавление небольших гидрофильных молекул, таких как сахароза, в ламеллярные липосомы со смешанными липидами , изготовленные из богатых галактолипидами тилакоидных мембран, дестабилизирует бислои и переходит в мицеллярную фазу. [71] Обычно K b не измеряется экспериментально, а рассчитывается на основе измерений K a и толщины бислоя, поскольку эти три параметра связаны между собой.
Это мера того, сколько энергии требуется, чтобы подвергнуть край бислоя воздействию воды, разрывая бислой или создавая в нем дыру. Происхождение этой энергии заключается в том, что создание такого интерфейса подвергает некоторые липидные хвосты воздействию воды, но точная ориентация этих пограничных липидов неизвестна. Есть некоторые свидетельства того, что могут сосуществовать как гидрофобные (с прямыми хвостами), так и гидрофильные (с загнутыми головками) поры. [72] [73]
Слияние
[ редактировать ]
Слияние — это процесс, при котором два липидных бислоя сливаются, образуя одну связанную структуру. Если это слияние происходит полностью через обе створки обоих бислоев, образуется заполненный водой мостик и растворы, содержащиеся в бислоях, могут смешиваться. Альтернативно, если в процессе слияния участвует только один листок от каждого бислоя, бислои называются полуслитыми. Слияние участвует во многих клеточных процессах, в частности у эукариот , поскольку эукариотическая клетка широко разделена липидными двухслойными мембранами. Экзоцитоз , оплодотворение яйцеклетки — вот лишь некоторые из многих эукариотических процессов, которые основаны на той путем активации спермы и транспорт отходов в лизозому или иной форме слияния. Даже проникновение патогенов можно контролировать путем слияния, поскольку многие вирусы с двухслойной оболочкой имеют специальные слитые белки для проникновения в клетку-хозяина.
В процессе слияния есть четыре основных этапа. [30] Во-первых, задействованные мембраны должны агрегировать, приближаясь друг к другу с точностью до нескольких нанометров. Во-вторых, два бислоя должны вступить в очень тесный контакт (в пределах нескольких ангстрем). Чтобы достичь такого тесного контакта, две поверхности должны стать хотя бы частично обезвоженными, поскольку обычно присутствующая связанная поверхностная вода вызывает сильное отталкивание бислоев. Присутствие ионов, в частности двухвалентных катионов, таких как магний и кальций, сильно влияет на этот этап. [74] [75] Одна из важнейших ролей кальция в организме – регулирование слияния мембран. В-третьих, в одной точке между двумя бислоями должна образоваться дестабилизация, локально искажающая их структуры. Точная природа этого искажения неизвестна. Одна из теорий состоит в том, что между двумя бислоями должен образоваться сильно изогнутый «стебель». [76] Сторонники этой теории считают, что она объясняет, почему фосфатидилэтаноламин, сильно изогнутый липид, способствует слиянию. [77] Наконец, на последнем этапе сплавления этот точечный дефект растет, и компоненты двух бислоев смешиваются и диффундируют от места контакта.
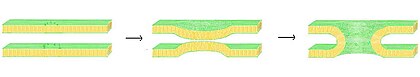

Ситуация еще больше усложняется при рассмотрении слияния in vivo , поскольку биологическое слияние почти всегда регулируется действием мембран-ассоциированных белков . Первыми из этих белков, которые были изучены, были слитые вирусные белки, которые позволяют вирусу с оболочкой вставлять свой генетический материал в клетку-хозяина (вирусы с оболочкой - это вирусы, окруженные липидным бислоем; некоторые другие имеют только белковую оболочку). Эукариотические клетки также используют слитые белки, наиболее изученными из которых являются SNARE . Белки SNARE используются для управления всем везикулярным внутриклеточным транспортом. Несмотря на годы исследований, многое еще неизвестно о функции этого класса белков. Фактически, до сих пор ведутся активные дебаты относительно того, связаны ли SNARE с ранней стыковкой или участвуют позже в процессе слияния, способствуя гемифузии. [79]
В исследованиях молекулярной и клеточной биологии часто желательно искусственно вызвать слияние. Добавление полиэтиленгликоля (ПЭГ) вызывает слияние без значительной агрегации или биохимического разрушения. Эта процедура сейчас широко используется, например, путем слияния В-клеток с клетками миеломы . [80] Полученная « гибридома в результате этой комбинации » экспрессирует желаемое антитело , что определяется участвующими В-клетками, но иммортализована благодаря компоненту меланомы. Слияние также можно вызвать искусственно посредством электропорации в процессе, известном как электрослияние. Считается, что это явление возникает из-за энергетически активных краев, образующихся во время электропорации, которые могут действовать как локальная точка дефекта, вызывающая рост стебля между двумя бислоями. [81]
Модельные системы
[ редактировать ]Липидные бислои можно создавать искусственно в лаборатории, чтобы позволить исследователям проводить эксперименты, которые невозможно провести с естественными бислоями. Их также можно использовать в области синтетической биологии для определения границ искусственных клеток . Эти синтетические системы называются модельными липидными бислоями. Существует множество различных типов модельных бислоев, каждый из которых имеет экспериментальные преимущества и недостатки. Они могут быть изготовлены как из синтетических, так и из натуральных липидов. Среди наиболее распространенных модельных систем можно выделить:
- Черные липидные мембраны (BLM)
- Поддерживаемые липидные бислои (SLB)
- Связанные двухслойные липидные мембраны (t-BLM)
- Везикулы
- Двухслойные интерфейсы капель (DIB)
Коммерческие приложения
[ редактировать ]На сегодняшний день наиболее успешным коммерческим применением липидных бислоев является использование липосом для доставки лекарств, особенно для лечения рака. (Обратите внимание: термин «липосома» по сути является синонимом « везикулы », за исключением того, что везикула — это общий термин для обозначения структуры, тогда как липосома относится только к искусственным, а не природным везикулам.) Основная идея липосомальной доставки лекарственного средства заключается в том, что лекарственное средство инкапсулируется в раствор внутри липосомы, а затем вводится пациенту. Эти загруженные лекарством липосомы проходят через систему, пока не связываются в целевом участке и не разрываются, высвобождая лекарство. Теоретически липосомы должны стать идеальной системой доставки лекарств, поскольку они могут изолировать практически любое гидрофильное лекарство, могут быть привиты молекулами для воздействия на определенные ткани и могут быть относительно нетоксичными, поскольку в организме есть биохимические пути расщепления липидов . [82]
Липосомы для доставки лекарств первого поколения имели простой липидный состав и имели ряд ограничений. Кровообращение в кровотоке было чрезвычайно ограничено как из-за почечной очистки, так и из -за фагоцитоза . Уточнение липидной композиции для настройки текучести, плотности поверхностного заряда и поверхностной гидратации привело к образованию везикул, которые адсорбируют меньше белков из сыворотки и, следовательно, менее легко распознаются иммунной системой . [83] Наиболее значительным достижением в этой области стало прививание полиэтиленгликоля (ПЭГ) на поверхность липосом для создания «скрытых» везикул, которые циркулируют в течение длительного времени без иммунного или почечного очищения. [84]
Первые стелс-липосомы были пассивно нацелены на опухолевые ткани. Поскольку опухоли вызывают быстрый и неконтролируемый ангиогенез, они особенно «протекают» и позволяют липосомам выходить из кровотока с гораздо большей скоростью, чем нормальная ткань. [85] Совсем недавно [ когда? ] Была предпринята работа по трансплантации антител или других молекулярных маркеров на поверхность липосом в надежде активно связывать их с определенным типом клеток или тканей. [86] Некоторые примеры этого подхода уже проходят клинические испытания. [87]
Еще одним потенциальным применением липидных бислоев является область биосенсоров . Поскольку липидный бислой является барьером между внутренней и внешней частью клетки, он также является местом обширной передачи сигнала. Исследователи на протяжении многих лет пытались использовать этот потенциал для разработки двухслойного устройства для клинической диагностики или обнаружения биотерроризма. Прогресс в этой области был медленным, и, хотя несколько компаний разработали автоматизированные системы обнаружения на основе липидов, они по-прежнему ориентированы на исследовательское сообщество. К ним относится компания Biacore (теперь GE Healthcare Life Sciences), которая предлагает одноразовый чип для использования липидных бислоев в исследованиях кинетики связывания. [88] и Nanion Inc., разработавшая автоматизированную систему зажима заплат . [89] Также разрабатываются и другие, более экзотические применения, такие как использование пор липидного бислоя мембраны для секвенирования ДНК компанией Oxford Nanolabs. На сегодняшний день эта технология не доказала свою коммерческую жизнеспособность.
Поддержанный липидный бислой (SLB), как описано выше, достиг коммерческого успеха в качестве метода скрининга для измерения проницаемости лекарств. Этот параллельный липидных коктейлей, которые , измеряет обнаружено , проницаемость анализа проницаемости искусственных мембран PAMPA как специально метод составленных тесно коррелируют с Caco-2 . культурами [90] [91] желудочно -кишечный тракт , [92] гематоэнцефалический барьер [93] и кожа. [94]
История
[ редактировать ]К началу двадцатого века ученые пришли к выводу, что клетки окружены тонким маслянистым барьером. [95] но структурная природа этой мембраны не была известна. Два эксперимента 1925 года заложили основу для заполнения этого пробела. Измеряя емкость растворов эритроцитов , Хьюго Фрике определил, что толщина клеточной мембраны составляет 3,3 нм. [96]
Хотя результаты этого эксперимента были точными, Фрике неверно истолковал данные, утверждая, что клеточная мембрана представляет собой один молекулярный слой. Профессор доктор Эверт Гортер [97] (1881–1954) и Ф. Грендель из Лейденского университета подошли к проблеме с другой точки зрения, распределив липиды эритроцитов в виде монослоя по впадине Ленгмюра-Блоджетт . Когда они сравнили площадь монослоя с площадью поверхности клеток, они обнаружили соотношение два к одному. [98] Более поздние анализы показали несколько ошибок и неверных предположений в этом эксперименте, но по счастливой случайности эти ошибки нивелировались, и на основе этих ошибочных данных Гортер и Грендель сделали правильный вывод: клеточная мембрана представляет собой липидный бислой. [30]
Эта теория была подтверждена с помощью электронной микроскопии в конце 1950-х годов. Хотя он не опубликовал первое электронно-микроскопическое исследование липидных бислоев. [99] Дж. Дэвид Робертсон был первым, кто предположил, что две темные электронно-плотные полосы представляют собой головные группы и связанные с ними белки двух расположенных рядом липидных монослоев. [100] [101] В этой работе Робертсон выдвинул концепцию «единичной мембраны». Это был первый случай, когда двухслойная структура была универсально приписана всем клеточным мембранам, а также органелл мембранам .
Примерно в то же время разработка модельных мембран подтвердила, что липидный бислой представляет собой стабильную структуру, способную существовать независимо от белков. «Нарисовав» раствор липида в органическом растворителе через отверстие, Мюллер и Рудин смогли создать искусственный бислой и определить, что он обладает латеральной текучестью, высоким электрическим сопротивлением и самовосстановлением в ответ на прокол. [102] все это свойства естественной клеточной мембраны. Несколько лет спустя Алек Бэнэм показал, что бислои в форме липидных пузырьков также могут образовываться, просто подвергая высушенный образец липида воздействию воды. [103] Это было важным достижением, поскольку оно продемонстрировало, что липидные бислои формируются спонтанно посредством самосборки и не требуют структурированной опорной структуры.
В 1977 году Кунитаке и Окахата изготовили полностью синтетическую двухслойную мембрану из одного органического соединения — бромида дидодецилдиметиламмония. [104] Это ясно показывает, что двухслойная мембрана была собрана за счет межмолекулярных сил .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Андерсен, Олаф С.; Кеппе, II, Роджер Э. (июнь 2007 г.). «Толщина бислоя и функция мембранного белка: энергетическая перспектива». Ежегодный обзор биофизики и биомолекулярной структуры . 36 (1): 107–130. doi : 10.1146/annurev.biophys.36.040306.132643 . ПМИД 17263662 . S2CID 6521535 .
- ^ Дивеча, Нуллин; Ирвин, Робин Ф (27 января 1995 г.). «Фосфолипидная сигнализация» . Клетка . 80 (2): 269–278. дои : 10.1016/0092-8674(95)90409-3 . ПМИД 7834746 . S2CID 14120598 .
- ^ Машаги и др. Гидратация сильно влияет на молекулярную и электронную структуру мембранных фосфолипидов. 136, 114709 (2012) «Журнал химической физики» . Архивировано из оригинала 15 мая 2016 года . Проверено 17 мая 2012 г.
- ^ Перейти обратно: а б Льюис Б.А., Энгельман Д.М. (май 1983 г.). «Толщина липидного бислоя линейно зависит от длины ацильной цепи в жидких фосфатидилхолиновых везикулах». Дж. Мол. Биол . 166 (2): 211–7. дои : 10.1016/S0022-2836(83)80007-2 . ПМИД 6854644 .
- ^ Заккай Дж., Блази Дж. К., Шенборн Б. П. (январь 1975 г.). «Нейтронографические исследования расположения воды в мембранах двухслойной модели лецитина» . Учеб. Натл. акад. наук. США . 72 (1): 376–380. Бибкод : 1975PNAS...72..376Z . дои : 10.1073/pnas.72.1.376 . ПМК 432308 . ПМИД 16592215 .
- ^ Нэгл Дж. Ф., Тристрам-Нэгл С. (ноябрь 2000 г.). «Строение липидных бислоев» . Биохим. Биофиз. Акта . 1469 (3): 159–95. дои : 10.1016/S0304-4157(00)00016-2 . ПМЦ 2747654 . ПМИД 11063882 .
- ^ Перейти обратно: а б Паркер Дж., Мэдиган М.Т., Брок Т.Д., Мартинко Дж.М. (2003). Брока биология микроорганизмов (10-е изд.). Энглвуд Клиффс, Нью-Джерси: Прентис Холл. ISBN 978-0-13-049147-3 .
- ^ Марш Д. (июль 2001 г.). «Полярность и профили проникновения в липидных мембранах» . Учеб. Натл. акад. наук. США . 98 (14): 7777–82. Бибкод : 2001PNAS...98.7777M . дои : 10.1073/pnas.131023798 . ПМК 35418 . ПМИД 11438731 .
- ^ Марш Д. (декабрь 2002 г.). «Профили мембранной водопроницаемости на спин-этикетках». Евро. Биофиз. Дж . 31 (7): 559–62. дои : 10.1007/s00249-002-0245-z . ПМИД 12602343 . S2CID 36212541 .
- ^ Перейти обратно: а б с д Равич В., Ольбрих К.К., Макинтош Т., Нидхэм Д., Эванс Э. (июль 2000 г.). «Влияние длины цепи и ненасыщенности на эластичность липидных бислоев» . Биофиз. Дж . 79 (1): 328–39. Бибкод : 2000BpJ....79..328R . дои : 10.1016/S0006-3495(00)76295-3 . ПМК 1300937 . ПМИД 10866959 .
- ^ Траубл Х., Хейнс Д.Х. (1971). «Изменение объема в ламелях липидного бислоя при фазовом переходе кристалл-жидкость-кристалл». хим. Физ. Липиды . 7 (4): 324–35. дои : 10.1016/0009-3084(71)90010-7 .
- ^ Бретчер М.С. (1 марта 1972 г.). «Асимметричная двухслойная структура липидов биологических мембран». Новая биология природы . 236 (61): 11–12. дои : 10.1038/newbio236011a0 . ПМИД 4502419 .
- ^ Верклей А.Дж., Цваал Р.Ф., Рулофсен Б., Комфуриус П., Кастелийн Д., ван Динен Л.Л. (октябрь 1973 г.). «Асимметричное распределение фосфолипидов в мембране эритроцитов человека. Комбинированное исследование с использованием фосфолипаз и электронной микроскопии с замораживанием». Биохим. Биофиз. Акта . 323 (2): 178–93. дои : 10.1016/0005-2736(73)90143-0 . ПМИД 4356540 .
- ^ Кунс, RT; Грин, Р.Дж.; Фрейзер, РА (2021). «Исследование состава головных групп липидов в эпителиальных мембранах: систематический обзор» . Мягкая материя . 17 (28): 6773–6786. Бибкод : 2021SMat...17.6773C . дои : 10.1039/D1SM00703C . ISSN 1744-683X . ПМИД 34212942 . S2CID 235708094 .
- ^ Белл Р.М., Баллас Л.М., Коулман Р.А. (1 марта 1981 г.). «Липидный топогенез» . Дж. Липид Рес . 22 (3): 391–403. дои : 10.1016/S0022-2275(20)34952-X . ПМИД 7017050 .
- ^ Бретчер М.С. (август 1973 г.). «Мембранная структура: некоторые общие принципы». Наука . 181 (4100): 622–629. Бибкод : 1973Sci...181..622B . дои : 10.1126/science.181.4100.622 . ПМИД 4724478 . S2CID 34501546 .
- ^ Ротман Дж. Э., Кеннеди Е. П. (май 1977 г.). «Быстрое трансмембранное движение вновь синтезированных фосфолипидов во время сборки мембраны» . Учеб. Натл. акад. наук. США . 74 (5): 1821–5. Бибкод : 1977PNAS...74.1821R . дои : 10.1073/pnas.74.5.1821 . ПМК 431015 . ПМИД 405668 .
- ^ Корнберг Р.Д., МакКоннелл Х.М. (март 1971 г.). «Переходы фосфолипидов внутрь и наружу в мембранах везикул». Биохимия . 10 (7): 1111–20. дои : 10.1021/bi00783a003 . ПМИД 4324203 .
- ^ Литман Б.Я. (июль 1974 г.). «Определение молекулярной асимметрии в поверхностном распределении фосфатидилэтаноламина в смешанных фосфолипидных везикулах». Биохимия . 13 (14): 2844–8. дои : 10.1021/bi00711a010 . ПМИД 4407872 .
- ^ Крейн Дж. М., Кисслинг В., Тамм Л. К. (февраль 2005 г.). «Измерение липидной асимметрии в плоских поддерживаемых бислоях с помощью флуоресцентной интерференционной контрастной микроскопии». Ленгмюр . 21 (4): 1377–88. дои : 10.1021/la047654w . ПМИД 15697284 .
- ^ Калб Э., Фрей С., Тамм Л.К. (январь 1992 г.). «Формирование поддерживаемых плоских бислоев путем слияния везикул с поддерживаемыми монослоями фосфолипидов». Биохим. Биофиз. Акта . 1103 (2): 307–16. дои : 10.1016/0005-2736(92)90101-Q . ПМИД 1311950 .
- ^ Lin WC, Blanchette CD, Ratto TV, Longo ML (январь 2006 г.). «Липидная асимметрия в липидных бислоях, поддерживаемых DLPC/DSPC: комбинированное исследование АСМ и флуоресцентной микроскопии» . Биофиз. Дж . 90 (1): 228–37. Бибкод : 2006BpJ....90..228L . дои : 10.1529/biophysj.105.067066 . ПМК 1367021 . ПМИД 16214871 .
- ^ Перес-Салас, Урсула; Поркар, Лайонел; Гарг, Сумит; Да, Мануэла А.А.; Левитан, Ирена (октябрь 2022 г.). «Эффективные параметры, контролирующие перенос стерола: исследование малоуглового рассеяния нейтронов с временным разрешением» . Журнал мембранной биологии . 255 (4–5): 423–435. дои : 10.1007/s00232-022-00231-3 . ISSN 1432-1424 . ПМИД 35467109 . S2CID 248375027 .
- ^ Гарг, С.; Поркар, Л.; Вудка, AC; Батлер, PD; Перес-Салас, У. (20 июля 2011 г.). «Измерения неинвазивного рассеяния нейтронов показывают более медленный транспорт холестерина в модельных липидных мембранах» . Биофизический журнал . 101 (2): 370–377. Бибкод : 2011BpJ...101..370G . дои : 10.1016/j.bpj.2011.06.014 . ISSN 1542-0086 . ПМК 3136766 . ПМИД 21767489 .
- ^ Перейти обратно: а б Девералл, Миранда А.; Гарг, Сумит; Людтке, Карин; Джордан, Райнер; Рюэ, Юрген; Науманн, Кристоф А. (12 августа 2008 г.). «Трансбислойное соединение затрудненной диффузии липидов в связанных с полимером фосфолипидных бислоях» . Мягкая материя . 4 (9): 1899–1908. Бибкод : 2008SMat....4.1899D . дои : 10.1039/B800801A . ISSN 1744-6848 .
- ^ Перейти обратно: а б Гарг, Сумит; Рюэ, Юрген; Людтке, Карин; Джордан, Райнер; Науманн, Кристоф А. (15 февраля 2007 г.). «Регистрация доменов в липидных смесях, имитирующих плоты, изученных с использованием связанных с полимером липидных бислоев» . Биофизический журнал . 92 (4): 1263–1270. Бибкод : 2007BpJ....92.1263G . doi : 10.1529/biophysj.106.091082 . ISSN 0006-3495 . ПМЦ 1783876 . ПМИД 17114215 .
- ^ Берг, Ховард К. (1993). Случайные блуждания по биологии (Расширенное издание в мягкой обложке). Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-00064-0 .
- ^ Дитрих С., Воловик З.Н., Леви М., Томпсон Н.Л., Джейкобсон К. (сентябрь 2001 г.). «Разделение Thy-1, GM1 и аналогов сшитых фосфолипидов на липидные рафты, восстановленные в монослоях модельной мембраны на носителе» . Учеб. Натл. акад. наук. США . 98 (19): 10642–7. Бибкод : 2001PNAS...9810642D . дои : 10.1073/pnas.191168698 . ПМЦ 58519 . ПМИД 11535814 .
- ^ Альбертс, Брюс (2017). «Глава 10: Мембранные структуры» . Молекулярная биология клетки . Гирляндная наука. ISBN 9781317563747 .
- ^ Перейти обратно: а б с Йигл, Филип (1993). Мембраны клеток (2-е изд.). Бостон: Академическая пресса. ISBN 978-0-12-769041-4 .
- ^ Фадок В.А., Брэттон Д.Л., Фраш С.С., Уорнер М.Л., Хенсон П.М. (июль 1998 г.). «Роль фосфатидилсерина в распознавании апоптотических клеток фагоцитами» . Гибель клеток отличается . 5 (7): 551–62. дои : 10.1038/sj.cdd.4400404 . ПМИД 10200509 .
- ^ Андерсон Х.К., Гаримелла Р., Тейг С.Э. (январь 2005 г.). «Роль матричных везикул в развитии пластинки роста и биоминерализации» . Передний. Биосци . 10 (1–3): 822–37. дои : 10.2741/1576 . ПМИД 15569622 .
- ^ Инес Э.Д., Хайлер А.В. (январь 1987 г.). «Осаждение фосфата кальция в водных суспензиях фосфатидилсеринсодержащих анионных липосом». Кальциф. Ткань Int . 40 (1): 43–8. дои : 10.1007/BF02555727 . ПМИД 3103899 . S2CID 26435152 .
- ^ Ким Дж., Мосиор М., Чунг Л.А., Ву Х., Маклафлин С. (июль 1991 г.). «Связывание пептидов с основными остатками с мембранами, содержащими кислые фосфолипиды» . Биофиз. Дж . 60 (1): 135–48. Бибкод : 1991BpJ....60..135K . дои : 10.1016/S0006-3495(91)82037-9 . ПМК 1260045 . ПМИД 1883932 .
- ^ Кох А.Л. (1984). «Первобытные клетки: возможные механизмы генерации энергии и деления клеток». Дж. Мол. Эвол . 21 (3): 270–7. дои : 10.1007/BF02102359 . ПМИД 6242168 . S2CID 21635206 .
- ^ «5.1 Структура клеточной мембраны | Науки о жизни | Токийский университет» . Архивировано из оригинала 22 февраля 2014 года . Проверено 10 ноября 2012 г.
- ^ Перейти обратно: а б Альбертс, Брюс (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-4072-0 .
- ^ Мартелли П.Л., Фариселли П., Касадио Р. (2003). «Подход машинного обучения ENSEMBLE для предсказания всех альфа-мембранных белков» . Биоинформатика . 19 (Приложение 1): i205–11. doi : 10.1093/биоинформатика/btg1027 . ПМИД 12855459 .
- ^ Филмор Д. (2004). «Это мир GPCR». Современное открытие лекарств . 11 : 24–9.
- ^ Монталь М., Мюллер П. (декабрь 1972 г.). «Формирование бимолекулярных мембран из липидных монослоев и исследование их электрических свойств» . Учеб. Натл. акад. Наука . 69 (12): 3561–6. Бибкод : 1972PNAS...69.3561M . дои : 10.1073/pnas.69.12.3561 . ПМК 389821 . ПМИД 4509315 .
- ^ Меликов К.С., Фролов В.А., Щербаков А, Самсонов А.В., Чизмаджев Ю.А., Черномордик Л.В. (апрель 2001 г.). «Индуцированные напряжением непроводящие предпоры и метастабильные одиночные поры в немодифицированном плоском липидном бислое» . Биофиз. Дж . 80 (4): 1829–36. Бибкод : 2001BpJ....80.1829M . дои : 10.1016/S0006-3495(01)76153-X . ПМК 1301372 . ПМИД 11259296 .
- ^ Неер Э., Сакманн Б. (апрель 1976 г.). «Одноканальные токи, зарегистрированные с мембраны денервированных мышечных волокон лягушки» . Природа . 260 (5554): 799–802. Бибкод : 1976Natur.260..799N . дои : 10.1038/260799a0 . ПМИД 1083489 . S2CID 4204985 .
- ^ Хойзер Дж. Э., Риз Т. С., Деннис М. Дж., Ян Ю, Ян Л., Эванс Л. (май 1979 г.). «Экзоцитоз синаптических везикул, захватываемый быстрым замораживанием и коррелирующий с высвобождением квантового медиатора» . Дж. Клеточная Биол . 81 (2): 275–300. дои : 10.1083/jcb.81.2.275 . ПМК 2110310 . ПМИД 38256 .
- ^ Дубинский М.А., Лесовой Д.М., Дубовский П.В., Чупин В.В., Арсеньев А.С. (июнь 2006 г.). «Моделирование 31 Спектры P-ЯМР магнитоориентированных фосфолипидных липосом: новое аналитическое решение». Solid State Nucl Magn Reson . 29 (4): 305–311. doi : 10.1016/j.ssnmr.2005.10.009 . PMID 16298110 . [ мертвая ссылка ]
- ^ Перейти обратно: а б с Ройтер, Юрий; Орнатская Марина; Раммохан, Аравинд Р.; Балакришнан, Джитендра; Хейне, Дэвид Р.; Минько, Сергей (2008). «Взаимодействие наночастиц с липидной мембраной». Нано-буквы . 8 (3): 941–944. Бибкод : 2008NanoL...8..941R . дои : 10.1021/nl080080l . ПМИД 18254602 .
- ^ Токумасу Ф., Джин А.Дж., Дворжак Дж.А. (2002). «Фазовое поведение липидной мембраны, выясненное в реальном времени с помощью атомно-силовой микроскопии в контролируемой среде» . Журнал электронной микроскопии . 51 (1): 1–9. дои : 10.1093/jmicro/51.1.1 . ПМИД 12003236 .
- ^ Рихтер Р.П., Бриссон А. (2003). «Характеристика липидных бислоев и белковых сборок, закрепленных на шероховатых поверхностях, с помощью атомно-силовой микроскопии». Ленгмюр . 19 (5): 1632–40. дои : 10.1021/la026427w . S2CID 56532332 .
- ^ Перейти обратно: а б Стельтенкамп С., Мюллер М.М., Дезерно М., Хеннестал С., Стейнем С., Яншофф А. (июль 2006 г.). «Механические свойства перекрывающих поры липидных бислоев, исследованные с помощью атомно-силовой микроскопии» . Биофиз. Дж . 91 (1): 217–26. Бибкод : 2006BpJ....91..217S . doi : 10.1529/biophysj.106.081398 . ПМК 1479081 . ПМИД 16617084 .
- ^ Алиреза Машаги и др., Гидратация сильно влияет на молекулярную и электронную структуру мембранных фосфолипидов. Дж. Хим. Физ. 136, 114709 (2012) «Журнал химической физики» . Архивировано из оригинала 15 мая 2016 года . Проверено 17 мая 2012 г.
- ^ Чакрабарти AC (1994). «Проницаемость мембран для аминокислот и модифицированных аминокислот: механизмы транслокации». Аминокислоты . 6 (3): 213–29. дои : 10.1007/BF00813743 . ПМИД 11543596 . S2CID 24350029 .
- ^ Хаузер Х., Филлипс М.К., Стаббс М. (октябрь 1972 г.). «Ионная проницаемость фосфолипидных бислоев». Природа . 239 (5371): 342–4. Бибкод : 1972Natur.239..342H . дои : 10.1038/239342a0 . ПМИД 12635233 . S2CID 4185197 .
- ^ Папахаджопулос Д., Уоткинс Дж. К. (сентябрь 1967 г.). «Фосфолипидные модельные мембраны. II. Проницаемость гидратированных жидких кристаллов». Биохим. Биофиз. Акта . 135 (4): 639–52. дои : 10.1016/0005-2736(67)90095-8 . ПМИД 6048247 .
- ^ Паула С., Волков А.Г., Ван Хук А.Н., Хейнс Т.Х., Димер Д.В. (январь 1996 г.). «Проникновение протонов, ионов калия и небольших полярных молекул через фосфолипидные бислои в зависимости от толщины мембраны» . Биофиз. Дж . 70 (1): 339–48. Бибкод : 1996BpJ....70..339P . дои : 10.1016/S0006-3495(96)79575-9 . ПМЦ 1224932 . ПМИД 8770210 .
- ^ Сян, Техас, Андерсон Б.Д. (июнь 1994 г.). «Взаимосвязь между размером проникающего вещества и проницаемостью в липидных бислойных мембранах». Дж. Член. Биол . 140 (2): 111–22. дои : 10.1007/bf00232899 . ПМИД 7932645 . S2CID 20394005 .
- ^ Гуо Э., Маккиннон Р. (декабрь 2005 г.). «Принципы селективного транспорта ионов в каналах и насосах». Наука . 310 (5753): 1461–5. Бибкод : 2005Sci...310.1461G . дои : 10.1126/science.1113666 . ПМИД 16322449 . S2CID 16323721 .
- ^ Гундельфингер Э.Д., Кессельс М.М., Квалманн Б. (февраль 2003 г.). «Временная и пространственная координация экзоцитоза и эндоцитоза». Нат. Преподобный мол. Клеточная Биол . 4 (2): 127–39. дои : 10.1038/nrm1016 . ПМИД 12563290 . S2CID 14415959 .
- ^ Штейнман Р.М., Броди С.Е., Кон З.А. (март 1976 г.). «Мембранный поток при пиноцитозе. Стереологический анализ» . Дж. Клеточная Биол . 68 (3): 665–87. дои : 10.1083/jcb.68.3.665 . ПМК 2109655 . ПМИД 1030706 .
- ^ YashRoy RC (1999) «Экзоцитоз у прокариотов» и его роль в инвазии сальмонеллы . ICAR NEWS - Информационный бюллетень по науке и технологиям , (октябрь-декабрь), том. 5(4), стр. 18. https://www.researchgate.net/publication/230822402_'Exocytosis_in_prokaryotes'_and_its_role_in_Salmonella_invasion?ev=prf_pub
- ^ YashRoy RC (1993)Электронно-микроскопические исследования поверхностных пилей и везикул Salmonella 3,10:r:- организмов. Индийский журнал Anim Sci 63, 99–102. https://www.researchgate.net/publication/230817087_Electron_microscope_studies_of_surface_pilli_and_vesicles_of_Salmonella_310r-_organisms?ev=prf_pub
- ^ YashRoy RC (1998)Открытие везикулярного экзоцитоза у прокариот и его роль в инвазии сальмонеллы . Современная наука , том. 75(10), стр. 1062-1066. https://www.researchgate.net/publication/230793568_Discovery_of_vesicle_exocytosis_in_prokaryotes_and_its_role_in_Salmonella_invasion?ev=prf_pub
- ^ ЯшРой RC (1998). «Экзоцитоз грамотрицательных бактерий при инвазии сальмонелл в эпителий подвздошной кишки курицы» . Индийский журнал птицеводства . 33 (2): 119–123.
- ^ Нойманн Э., Шефер-Риддер М., Ван Ю., Хофшнайдер П.Х. (1982). «Перенос генов в клетки лиомы мышей путем электропорации в сильных электрических полях» . ЭМБО Дж . 1 (7): 841–5. дои : 10.1002/j.1460-2075.1982.tb01257.x . ПМЦ 553119 . ПМИД 6329708 .
- ^ Деманеш С., Бертолла Ф., Буре Ф. и др. (август 2001 г.). «Лабораторные доказательства переноса генов в почве, опосредованного молнией» . Прил. Окружающая среда. Микробиол . 67 (8): 3440–4. Бибкод : 2001ApEnM..67.3440D . дои : 10.1128/АЕМ.67.8.3440-3444.2001 . ПМК 93040 . ПМИД 11472916 .
- ^ Гарсия М.Л. (июль 2004 г.). «Ионные каналы: ожидания ворот». Природа . 430 (6996): 153–5. Бибкод : 2004Natur.430..153G . дои : 10.1038/430153а . ПМИД 15241399 . S2CID 4427370 .
- ^ Макинтош Т.Дж., Саймон С.А. (2006). «Роль свойств двухслойного материала в функции и распределении мембранных белков». Анну. Преподобный Биофиз. Биомол. Структурировать . 35 (1): 177–98. doi : 10.1146/annurev.biophys.35.040405.102022 . ПМИД 16689633 .
- ^ Сучина Т.М., Лента С.Е., Кеппе Р.Э., Андерсен О.С., Сакс Ф., Готлиб П.А. (июль 2004 г.). «Двуслойно-зависимое ингибирование механочувствительных каналов нейроактивными пептидными энантиомерами». Природа . 430 (6996): 235–40. Бибкод : 2004Natur.430..235S . дои : 10.1038/nature02743 . ПМИД 15241420 . S2CID 4401688 .
- ^ Халлетт Ф.Р., Марш Дж., Никель Б.Г., Вуд Дж.М. (февраль 1993 г.). «Механические свойства везикул. II. Модель осмотического набухания и лизиса» . Биофиз. Дж . 64 (2): 435–42. Бибкод : 1993BpJ....64..435H . дои : 10.1016/S0006-3495(93)81384-5 . ПМЦ 1262346 . ПМИД 8457669 .
- ^ Боал, Дэвид Х. (2001). Механика клетки . Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 978-0-521-79681-1 .
- ^ Рутковски Калифорния, Уильямс Л.М., Хейнс Т.Х., Cummins HZ (июнь 1991 г.). «Эластичность синтетических фосфолипидных везикул, полученных методом фотонной корреляционной спектроскопии». Биохимия . 30 (23): 5688–96. дои : 10.1021/bi00237a008 . ПМИД 2043611 .
- ^ Эванс Э., Генрих В., Людвиг Ф., Равич В. (октябрь 2003 г.). «Спектроскопия динамического натяжения и прочность биомембран» . Биофиз. Дж . 85 (4): 2342–50. Бибкод : 2003BpJ....85.2342E . дои : 10.1016/S0006-3495(03)74658-X . ПМЦ 1303459 . ПМИД 14507698 .
- ^ YashRoy RC (1994)Дестабилизация ламеллярной дисперсии липидов тилакоидных мембран сахарозой. Biochimica et Biophysical Acta , vol. 1212, стр. 129-133. https://www.researchgate.net/publication/15042978_Destabilisation_of_lamellar_dispersion_of_thylakoid_membrane_lipids_by_sucrose?ev=prf_pub
- ^ Уивер Ю.С., Чизмаджев Ю.А. (1996). «Теория электропорации: обзор». Биоэлектрохимия и биоэнергетика . 41 (2): 135–60. дои : 10.1016/S0302-4598(96)05062-3 .
- ^ Зейди, Махди; Ким, Чун Ил (2018). «Влияние внутримембранной вязкости на морфологию липидной мембраны: полное аналитическое решение» . Научные отчеты . 8 (1): 12845. Бибкод : 2018NatSR...812845Z . дои : 10.1038/s41598-018-31251-6 . ISSN 2045-2322 . ПМК 6110749 . ПМИД 30150612 .
- ^ Папахаджопулос Д., Нир С., Дюзгунес Н. (апрель 1990 г.). «Молекулярные механизмы кальций-индуцированного слияния мембран». Дж. Биоэнергетика. Биомембр . 22 (2): 157–79. дои : 10.1007/BF00762944 . ПМИД 2139437 . S2CID 1465571 .
- ^ Левентис Р., Ганье Дж., Фуллер Н., Рэнд Р.П., Сильвиус Дж.Р. (ноябрь 1986 г.). «Слияние, индуцированное двухвалентными катионами, и латеральная сегрегация липидов в везикулах фосфатидилхолина и фосфатидной кислоты». Биохимия . 25 (22): 6978–87. дои : 10.1021/bi00370a600 . ПМИД 3801406 .
- ^ Маркин В.С., Козлов М.М., Боровягин В.Л. (октябрь 1984 г.). «К теории слияния мембран. Стеблевой механизм». Генерал физиол. Биофиз . 3 (5): 361–77. ПМИД 6510702 .
- ^ Черномордик Л.В., Козлов М.М. (2003). «Белко-липидное взаимодействие при слиянии и делении биологических мембран» . Анну. Преподобный Биохим . 72 (1): 175–207. doi : 10.1146/annurev.biochem.72.121801.161504 . ПМИД 14527322 .
- ^ Георгиев, Данко Д.; Глейзбрук, Джеймс Ф. (2007). «Субнейронная обработка информации уединенными волнами и случайными процессами». В Лышевском, Сергей Эдвард (ред.). Справочник по нано- и молекулярной электронике . Серия «Нано и микроинженерия». ЦРК Пресс. С. 17 –1–17–41. дои : 10.1201/9781315221670-17 . ISBN 978-0-8493-8528-5 . S2CID 199021983 .
- ^ Чен Ю.А., Шеллер Р.Х. (февраль 2001 г.). «SNARE-опосредованное слияние мембран». Нат. Преподобный мол. Клеточная Биол . 2 (2): 98–106. дои : 10.1038/35052017 . ПМИД 11252968 . S2CID 205012830 .
- ^ Кёлер Г., Мильштейн С. (август 1975 г.). «Непрерывные культуры слитых клеток, секретирующих антитела заданной специфичности». Природа . 256 (5517): 495–7. Бибкод : 1975Natur.256..495K . дои : 10.1038/256495a0 . ПМИД 1172191 . S2CID 4161444 .
- ^ Джордан, Кэрол А.; Нойманн, Эберхард; Мейсон Соверши, Артур Э. (1989). Электропорация и электрослияние в клеточной биологии . Нью-Йорк: Пленум Пресс. ISBN 978-0-306-43043-5 .
- ^ Иммордино М.Л., Досио Ф., Кэттель Л. (2006). «Стелс-липосомы: обзор фундаментальной науки, обоснования и клинических применений, существующих и потенциальных» . Инт Дж Наномед . 1 (3): 297–315. дои : 10.2217/17435889.1.3.297 . ПМЦ 2426795 . ПМИД 17717971 .
- ^ Чонн А., Семпл СК, Каллис П.Р. (15 сентября 1992 г.). «Ассоциация белков крови с большими однослойными липосомами in vivo. Связь со временем циркуляции» . Ж. Биол. Хим . 267 (26): 18759–65. дои : 10.1016/S0021-9258(19)37026-7 . ПМИД 1527006 .
- ^ Борис Э.Х., Винтерхальтер М., Фредерик П.М., Валлнер Дж.Дж., Ласич Д.Д. (1997). «Стелс-липосомы: от теории к продукту». Обзоры расширенной доставки лекарств . 24 (2–3): 165–77. дои : 10.1016/S0169-409X(96)00456-5 .
- ^ Маэда Х., Сава Т., Конно Т. (июль 2001 г.). «Механизм адресной доставки макромолекулярных препаратов в опухоль, включая эффект ЭПР при солидной опухоли и клинический обзор прототипа полимерного препарата SMANCS». J Контрольный релиз . 74 (1–3): 47–61. дои : 10.1016/S0168-3659(01)00309-1 . ПМИД 11489482 .
- ^ Лопес Д.Е., Менезеш Д.Е., Кирхмайер М.Дж., Ганье Дж.Ф. (1999). «Клеточный транспорт и цитотоксичность липосомального доксорубицина, нацеленного на анти-CD19, в клетках В-лимфомы». Журнал исследований липосом . 9 (2): 199–228. дои : 10.3109/08982109909024786 .
- ^ Мацумура Ю., Гото М., Муро К. и др. (март 2004 г.). «Фаза I и фармакокинетическое исследование MCC-465, доксорубицина (DXR), инкапсулированного в иммунолипосомы ПЭГ, у пациентов с метастатическим раком желудка» . Энн. Онкол . 15 (3): 517–25. дои : 10.1093/annonc/mdh092 . ПМИД 14998859 .
- ^ [1] [ постоянная мертвая ссылка ] . Biacore Inc. Проверено 12 февраля 2009 г.
- ^ Нанионные технологии. Автоматический зажим патчей. Архивировано 31 марта 2010 года в Wayback Machine . Проверено 28 февраля 2010 г. (PDF)
- ^ Бермехо, М.; Авдеев, А.; Руис, А.; Налда, Р.; Руэлл, Дж.А.; Цинман О.; Гонсалес, И.; Фернандес, К.; Санчес, Г.; Гарригес, ТМ; Мерино, В. (2004). «PAMPA - модель абсорбции лекарственного средства in vitro 7. Сравнение проницаемости фторхинолонов для крыс in situ, Caco-2 и PAMPA». Европейский журнал фармацевтических наук . 21 (4): 429–41. дои : 10.1016/j.ejps.2003.10.009 . ПМИД 14998573 .
- ^ Авдеев, А.; Артурссон, П.; Нойхофф, С.; Лазорова Л.; Гросё, Дж.; Тавелин, С. (2005). «Проницаемость Caco-2 слабоосновных лекарств, предсказанная с помощью метода двойного стока PAMPA pKa (поток)». Европейский журнал фармацевтических наук . 24 (4): 333–49. дои : 10.1016/j.ejps.2004.11.011 . ПМИД 15734300 .
- ^ Авдеев, А.; Нильсен, ЧП; Цинман, О. (2004). «ПАМПА - модель 11 абсорбции лекарственного средства in vitro. Подбор толщины слоя неперемешиваемой воды in vivo путем индивидуального перемешивания лунок в микротитровальных планшетах». Европейский журнал фармацевтических наук . 22 (5): 365–74. дои : 10.1016/j.ejps.2004.04.009 . ПМИД 15265506 .
- ^ Дагенайс, К.; Авдеев, А.; Цинман О.; Дадли, А.; Беливо, Р. (2009). «Проницаемость гематоэнцефалического барьера у мышей с дефицитом P-гликопротеина in situ и ее прогнозирование с использованием комбинированной модели PAMPA» . Европейский журнал фармацевтических наук . 38 (2): 121–37. дои : 10.1016/j.ejps.2009.06.009 . ПМЦ 2747801 . ПМИД 19591928 .
- ^ Синько, Б.; Кёкёси, Дж.; Авдеев, А.; Такач-Новак, К. (2009). «Исследование PAMPA эффекта новых аналогов церамидов, повышающих проницаемость». Химия и биоразнообразие . 6 (11): 1867–74. дои : 10.1002/cbdv.200900149 . ПМИД 19937821 . S2CID 27395246 .
- ^ Леб Дж. (декабрь 1904 г.). «Новейшее развитие биологии» . Наука . 20 (519): 777–786. Бибкод : 1904Sci....20..777L . дои : 10.1126/science.20.519.777 . ПМИД 17730464 .
- ^ Фрике Х (1925). «Электрическая емкость суспензий с особым учетом крови» . Журнал общей физиологии . 9 (2): 137–52. дои : 10.1085/jgp.9.2.137 . ПМК 2140799 . ПМИД 19872238 .
- ^ Доорен Л.Дж., Видеманн Л.Р. (1986). «О бимолекулярных слоях липидов на хромоцитах крови». Журнал Европейского журнала педиатрии . 145 (5): 329. doi : 10.1007/BF00439232 . ПМИД 3539619 . S2CID 36842138 .
- ^ Гортер Э., Грендель Ф (1925). «О бимолекулярных слоях липидов на хромоцитах крови» . Журнал экспериментальной медицины . 41 (4): 439–43. дои : 10.1084/jem.41.4.439 . ПМК 2130960 . ПМИД 19868999 .
- ^ Сьёстранд Ф.С., Андерссон-Седергрен Э., Дьюи М.М. (апрель 1958 г.). «Ультраструктура вставочных дисков сердечной мышцы лягушки, мыши и морской свинки». Дж. Ультраструктур. Рез . 1 (3): 271–87. дои : 10.1016/S0022-5320(58)80008-8 . ПМИД 13550367 .
- ^ Робертсон Дж. Д. (1960). «Молекулярная структура и контактные взаимоотношения клеточных мембран». Прог. Биофиз. Мол. Биол . 10 : 343–418. ПМИД 13742209 .
- ^ Робертсон Дж. Д. (1959). «Ультраструктура клеточных мембран и их производных». Биохим. Соц. Симп . 16 :3–43. ПМИД 13651159 .
- ^ Мюллер П., Рудин Д.О., Тьен Х.Т., Уэскотт У.К. (июнь 1962 г.). «Восстановление структуры клеточной мембраны in vitro и превращение ее в возбудимую систему». Природа . 194 (4832): 979–80. Бибкод : 1962Natur.194..979M . дои : 10.1038/194979a0 . ПМИД 14476933 . S2CID 2110051 .
- ^ Бангэм, AD ; Хорн, RW (1964). «Отрицательное окрашивание фосфолипидов и их структурная модификация поверхностно-активными веществами, наблюдаемая в электронном микроскопе». Журнал молекулярной биологии . 8 (5): 660–668. дои : 10.1016/S0022-2836(64)80115-7 . ПМИД 14187392 .
- ^ Кунитаке Т (1977). «Полностью синтетическая двухслойная мембрана». Дж. Ам. хим. Соц . 99 (11): 3860–3861. дои : 10.1021/ja00453a066 .
Внешние ссылки
[ редактировать ]- LIPIDAT Обширная база данных физических свойств липидов.
- Структура жидких липидных бислоев. Архивировано 11 апреля 2011 г. в Wayback Machine Simulations и ссылки на публикации, связанные со структурой поперечного сечения липидных бислоев.