Глубоководное сообщество
Эта статья нуждается в дополнительных цитатах для проверки . ( январь 2021 г. ) |


Глубоководное сообщество — это любое сообщество организмов , связанных общей средой обитания в глубоком море . Глубоководные сообщества остаются в значительной степени неисследованными из-за технологических и логистических проблем и затрат, связанных с посещением этого отдаленного биома . Из-за уникальных проблем (особенно высокого атмосферного давления , экстремальных температур и отсутствия света ) долгое время считалось, что в этой враждебной среде существует мало жизни. Однако с XIX века исследования показали, что значительное биоразнообразие в глубоководных водах существует .
Тремя основными источниками энергии и питательных веществ для глубоководных сообществ являются морской снег , падение китов и хемосинтез в гидротермальных жерлах и холодных просачиваниях .
История
[ редактировать ]До XIX века учёные предполагали, что жизнь в глубоком океане немногочисленна. В 1870-х годах сэр Чарльз Уивилл Томсон и его коллеги на борту экспедиции «Челленджер» обнаружили множество глубоководных существ самых разных типов.
Первое открытие какого-либо глубоководного хемосинтетического сообщества, включающего высших животных, было неожиданно сделано в гидротермальных жерлах восточной части Тихого океана в ходе геологических исследований (Corliss et al., 1979). [ 1 ] Два учёных, Дж. Корлисс и Дж. ван Андел, впервые стали свидетелями плотных хемосинтезирующих слоев моллюсков с подводного аппарата DSV Alvin 17 февраля 1977 года, после их неожиданного открытия с помощью телекамеры с дистанционным управлением двумя днями ранее. [ 1 ]
— Бездна Челленджера самая глубокая исследуемая точка из всех океанов Земли; он расположен на южном конце Марианской впадины недалеко от группы Марианских островов . Впадина названа в честь HMS Challenger , исследователи которого сделали первые записи ее глубины 23 марта 1875 года на станции 225 . Заявленная глубина составила 4475 саженей (8184 метра) на основе двух отдельных зондирований. В 1960 году Дон Уолш и Жак Пиккар спустились на дно Бездны Челленджера на «Триест» батискафе . На такой огромной глубине была замечена маленькая, похожая на камбалу рыбка, удалявшаяся от прожектора батискафа.
Японский аппарат с дистанционным управлением (ROV) Kaiko стал вторым судном, достигшим дна Бездны Челленджера в марте 1995 года. Nereus , гибридный аппарат с дистанционным управлением (HROV) Океанографического института Вудс-Хоул , является единственным транспортным средством, способным исследовать океан. Глубины более 7000 метров. 31 мая 2009 года Нерей достиг глубины 10 902 метра в Бездне Челленджера. [ 2 ] [ 3 ] 1 июня 2009 года гидролокационное картирование Бездны Челленджера с помощью системы Simrad EM120 многолучевой гидролокационной батиметрической на борту научно-исследовательского судна Kilo Moana показало, что максимальная глубина составляет 10 971 метр (6,817 миль). Гидроакустическая система использует фазовое и амплитудное определение дна с точностью не хуже 0,2% от глубины воды (на этой глубине это погрешность около 22 метров). [ 3 ] [ 4 ]
Среда
[ редактировать ]Тьма
[ редактировать ]
Океан можно представить как разделенный на различные зоны в зависимости от глубины и наличия или отсутствия солнечного света . Почти все формы жизни в океане зависят от фотосинтетической деятельности фитопланктона и других морских растений по преобразованию углекислого газа в органический углерод , который является основным строительным блоком органического вещества . Фотосинтез, в свою очередь, требует энергии солнечного света для запуска химических реакций, в результате которых образуется органический углерод. [ 5 ]
Слой водной толщи , до которого проникает солнечный свет, называется фотозоной . можно Фотическую зону разделить на две разные вертикальные области. Самая верхняя часть фотической зоны, где достаточно света для поддержки фотосинтеза фитопланктона и растений, называется эвфотической зоной (также называемой эпипелагической зоной или поверхностной зоной ). [ 6 ] Нижняя часть фотической зоны, где интенсивность света недостаточна для фотосинтеза, называется дисфотической зоной (дисфотический в переводе с греческого означает «плохо освещенный»). [ 7 ] Дисфотическую зону называют также мезопелагической зоной или сумеречной зоной . [ 8 ] Его самая нижняя граница находится на термоклине 12 ° C (54 ° F), который в тропиках обычно находится на высоте от 200 до 1000 метров. [ 9 ]
Эвфотическая зона определяется несколько условно как простирающаяся от поверхности до глубины, где интенсивность света составляет примерно 0,1–1% от поверхностного солнечного излучения , в зависимости от времени года , широты и степени мутности воды . [ 6 ] [ 7 ] В самой чистой океанской воде эвфотическая зона может простираться на глубину около 150 метров. [ 6 ] или редко, до 200 метров. [ 8 ] Растворенные вещества и твердые частицы поглощают и рассеивают свет, а в прибрежных районах высокая концентрация этих веществ приводит к быстрому ослаблению света с глубиной. В таких районах глубина эвфотической зоны может составлять всего несколько десятков метров или меньше. [ 6 ] [ 8 ] Дисфотическая зона, где интенсивность света значительно меньше 1% поверхностной освещенности, простирается от основания эвфотической зоны примерно до 1000 метров. [ 9 ] От дна фотической зоны до морского дна простирается афотическая зона — область вечной тьмы. [ 8 ] [ 9 ]
Поскольку средняя глубина океана составляет около 3688 метров, [ 10 ] фотозона представляет собой лишь небольшую часть общего объема океана. Однако благодаря способности к фотосинтезу фотическая зона обладает наибольшим биоразнообразием и биомассой среди всех океанических зон. Здесь происходит почти вся первичная продукция океана. Любые формы жизни, присутствующие в афотической зоне, должны либо быть способны двигаться вверх через толщу воды в фотическую зону для питания, либо должны полагаться на опускание материала сверху . [ 5 ] или должны найти другой источник энергии и питания, как это происходит у хемосинтезирующих архей, обитающих вблизи гидротермальных источников и холодных выходов .
Гипербаричность
[ редактировать ]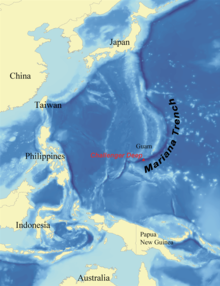
Эти животные эволюционировали , чтобы выдерживать экстремальное давление субфотических зон . Давление увеличивается примерно на одну атмосферу каждые десять метров. Чтобы справиться с давлением, многие рыбы довольно мелкие, обычно не превышающие 25 см в длину. Кроме того, ученые обнаружили, что чем глубже обитают эти существа, тем более студенистой их плоть и более минимальной является структура их скелета. Эти существа также удалили все лишние полости, которые могли бы разрушиться под давлением, например плавательные пузыри. [ 11 ]
Давление является величайшим фактором окружающей среды, действующим на глубоководные организмы. В глубоком море, хотя большая часть морских глубин находится под давлением от 200 до 600 атм, диапазон давления составляет от 20 до 1000 атм. Давление играет большую роль в распространении глубоководных организмов. До недавнего времени людям не хватало подробной информации о прямом воздействии давления на большинство глубоководных организмов, поскольку практически все организмы, выловленные из глубин моря, достигали поверхности мертвыми или умирающими. С появлением ловушек со специальной камерой для поддержания давления неповрежденные крупные многоклеточные животные стали извлекаться из морских глубин в хорошем состоянии. Некоторые из них используются в экспериментальных целях, и мы получаем больше знаний о биологических эффектах давления.
Температура
[ редактировать ]Двумя областями наибольшего и наиболее быстрого изменения температуры в океанах являются зона перехода между поверхностными и глубокими водами, термоклин, и переход между глубоководным дном и потоками горячей воды в гидротермальных жерлах. Мощность термоклинов варьируется от нескольких сотен метров до почти тысячи метров. Ниже термоклина водная масса глубокого океана холодная и гораздо более однородная. Наиболее сильны термоклины в тропиках, где температура эпипелагиали обычно превышает 20 °С. От основания эпипелагиали температура падает на несколько сотен метров до 5–6 °C на высоте 1000 метров. Он продолжает снижаться к низу, но темпы гораздо медленнее. Ниже 3000–4000 м вода изотермична . На любой заданной глубине температура практически не меняется в течение длительных периодов времени. Сезонных изменений температуры нет, как и годовых изменений. Ни одна другая среда обитания на Земле не имеет такой постоянной температуры.
Гидротермальные источники представляют собой прямой контраст с постоянной температурой. В этих системах температура воды, выходящей из труб «черного курильщика», может достигать 400 °C (от кипения ее удерживает высокое гидростатическое давление), а в пределах нескольких метров она может снова опуститься до 2–4 °С. [ 12 ]
Соленость
[ редактировать ]
Соленость постоянна на всей глубине глубокого моря. Из этого правила есть два заметных исключения:
- В Средиземном море потери воды за счет испарения значительно превышают поступления от осадков и речного стока. Из-за этого соленость в Средиземноморье выше, чем в Атлантическом океане . [ 13 ] Испарение особенно велико в его восточной половине, из-за чего в этом районе снижается уровень воды и увеличивается соленость. [ 14 ] Возникающий в результате градиент давления выталкивает относительно прохладную воду с низкой соленостью из Атлантического океана через бассейн. Эта вода нагревается и становится более соленой по мере продвижения на восток, затем опускается в районе Леванта и циркулирует на запад, чтобы вылиться обратно в Атлантику через Гибралтарский пролив . [ 15 ] Конечным результатом этого является то, что в Гибралтарском проливе существует поверхностное течение холодной воды с более низкой соленостью из Атлантики, направленное на восток, и одновременное течение на запад теплой соленой воды из Средиземного моря в более глубоких зонах. Вернувшись в Атлантику, эта химически отличающаяся средиземноморская промежуточная вода может сохраняться на протяжении тысяч километров от своего источника. [ 16 ]
- Бассейны с рассолом представляют собой большие участки рассола на морском дне . Эти бассейны представляют собой водоемы, соленость которых в три-пять раз превышает соленость окружающего океана. Для глубоководных рассолов источником соли является растворение крупных соли отложений в результате соляной тектоники . Рассол часто содержит высокие концентрации метана, обеспечивающего энергией хемосинтезирующих , экстремофилов обитающих в этом специализированном биоме . Известно также, что залежи рассола существуют на континентальном шельфе Антарктики , где источником рассола является соль, исключенная при формировании морского льда . Глубоководные и антарктические рассолы могут быть токсичными для морских животных. Бассейны с рассолом иногда называют озерами на морском дне, потому что плотный рассол создает галоклин , который нелегко смешивается с вышележащей морской водой. Высокая соленость повышает плотность рассола, что создает четкую поверхность и береговую линию бассейна. [ 17 ]
Глубоководное море , или глубокий слой, — самый нижний слой океана, существующий ниже термоклина, на глубине 1000 саженей (1800 м) и более. Самая глубокая часть глубокого моря — Марианская впадина, расположенная в западной части северной части Тихого океана. Это также самая глубокая точка земной коры. Его максимальная глубина составляет около 10,9 км, что превышает высоту Эвереста . В 1960 году Дон Уолш и Жак Пикар достигли дна Марианской впадины на «Триест» батискафе . Давление составляет около 11 318 метрических тонн-сил на квадратный метр (110,99 МПа или 16 100 фунтов на квадратный дюйм ).
Зоны
[ редактировать ]Мезопелагический
[ редактировать ]
Мезопелагическая зона представляет собой верхнюю часть средней водной зоны и простирается на глубину от 200 до 1000 метров (от 660 до 3280 футов) ниже уровня моря . В просторечии это известно как «сумеречная зона», поскольку свет все еще может проникать в этот слой, но он слишком низок, чтобы поддерживать фотосинтез. Однако ограниченное количество света все же позволяет организмам видеть, а существа с чувствительным зрением могут обнаруживать добычу, общаться и ориентироваться с помощью зрения. У организмов этого слоя большие глаза, что позволяет максимально увеличить количество света в окружающей среде. [ 18 ]
Большинство мезопелагических рыб совершают ежедневные вертикальные миграции , перемещаясь ночью в эпипелагическую зону, часто следуя за подобными миграциями зоопланктона, и возвращаясь на глубину в целях безопасности в течение дня. [ 19 ] [ 20 ] : 585 Эти вертикальные миграции часто происходят на большие вертикальные расстояния и осуществляются с помощью плавательного пузыря . Плавательный пузырь надувается, когда рыба хочет двигаться вверх, а, учитывая высокое давление в мезопелагической зоне, это требует значительных затрат энергии. По мере подъема рыбы давление в плавательном пузыре должно регулироваться, чтобы предотвратить его разрыв. Когда рыба хочет вернуться на глубину, плавательный пузырь сдувается. [ 21 ] Некоторые мезопелагические рыбы совершают ежедневные миграции через термоклин , где температура колеблется от 10 до 20 ° C (от 18 до 36 ° F), тем самым демонстрируя значительную устойчивость к изменению температуры. [ 20 ] : 590
Мезопелагические рыбы обычно не имеют защитных шипов и используют цвет и биолюминесценцию , чтобы маскировать их от других рыб. Засадные хищники темные, черные или красные. Поскольку более длинные красные длины волн света не достигают морских глубин, красный цвет эффективно действует так же, как черный. Мигрирующие формы используют с контрастными оттенками серебристые цвета . На животе у них часто имеются фотофоры, излучающие слабый свет. Для хищника снизу, смотрящего вверх, эта биолюминесценция маскирует силуэт рыбы. Однако у некоторых из этих хищников есть желтые линзы, которые фильтруют окружающий свет (с недостатком красного цвета), оставляя видимой биолюминесценцию. [ 22 ]
Батиал
[ редактировать ]
Батиальная зона представляет собой нижнюю часть средней водной зоны и охватывает глубины от 1000 до 4000 метров (от 3300 до 13 100 футов). [ 23 ] Свет не достигает этой зоны, поэтому она получила прозвище «полночная зона»; из-за недостатка света она менее густонаселена, чем эпипелагическая зона, хотя и значительно крупнее. [ 24 ] Рыбам трудно жить в этой зоне из-за давящего давления, низких температур 4 °C (39 °F), низкого уровня растворенного кислорода и недостатка питательных веществ. [ 20 ] : 585 То немногое энергии, которое имеется в батипелагической зоне, фильтруется сверху в виде детрита, фекалий и редких беспозвоночных или мезопелагических рыб. [ 20 ] : 594 Около 20% пищи, происходящей из эпипелагиали, попадает в мезопелагическую зону, но лишь около 5% фильтруется в батипелагическую зону. [ 25 ] Рыбы, обитающие там, возможно, имеют уменьшенные или полностью утраченные жабры, почки, сердце и плавательные пузыри, имеют слизистую, а не чешуйчатую кожу, а также слабое скелетное и мускулистое строение. [ 20 ] : 587 Отсутствие окостенения является адаптацией к экономии энергии, когда еды не хватает. [ 26 ] Большинство животных, обитающих в батиальной зоне, — беспозвоночные ( морские губки , головоногие и иглокожие) . За исключением очень глубоких участков океана, батиальная зона обычно достигает донной зоны морского дна . [ 24 ]
Бездонный и разговорный
[ редактировать ]
Абиссальная зона остается в вечной темноте на глубине от 4000 до 6000 метров (от 13 000 до 20 000 футов). [ 23 ] Единственные организмы, населяющие эту зону, — это хемотрофы и хищники, способные выдерживать огромное давление, иногда достигающее 76 мегапаскалей (750 атм; 11 000 фунтов на квадратный дюйм). Зона хадал (названная в честь Аида , греческого бога подземного мира) — это зона, предназначенная для самых глубоких траншей в мире, достигающих глубины ниже 6000 метров (20 000 футов). Самая глубокая точка хадальной зоны — Марианская впадина , глубина которой составляет 10 911 метров (35 797 футов), а давление составляет 110 мегапаскалей (1100 атм; 16 000 фунтов на квадратный дюйм). [ 27 ] [ 28 ] [ 29 ]
Источники энергии
[ редактировать ]
Морской снег
[ редактировать ]Верхняя фотическая зона океана заполнена взвешенными органическими веществами (ВОВ), особенно в прибрежных районах и зонах апвеллинга. Однако большинство ПОМ маленькие и легкие. Этим частицам могут потребоваться сотни или даже тысячи лет, чтобы оседать через толщу воды в глубокие глубины океана. Этой временной задержки достаточно для того, чтобы частицы реминерализовались и были поглощены организмами пищевой сети .
В глубоком Саргассовом море ученые из Океанографического института Вудс-Хоул (WHOI) обнаружили так называемый морской снег , в котором ПОМ переупаковываются в гораздо более крупные частицы, которые тонут с гораздо большей скоростью и падают, как снег. [ 30 ]
Из-за скудности пищи организмы, живущие на дне и на дне, обычно ведут себя условно-патогенно. У них есть особые приспособления к этой экстремальной среде: быстрый рост, эффектный механизм расселения личинок и способность использовать «переходной» пищевой ресурс. Типичным примером являются двустворчатые моллюски , сверлящие древесину, которые проникают в древесину и другие растительные остатки и питаются органическими веществами из останков.
Периодическое поверхностное цветение
[ редактировать ]Иногда внезапный доступ к питательным веществам у поверхности приводит к цветению планктона, водорослей или животных, таких как сальпы , которых становится настолько много, что они полностью погружаются на дно, не потребляясь другими организмами. Эти короткие порции питательных веществ, достигающие морского дна, могут превышать годы содержания морского снега и быстро потребляются животными и микробами. Отходы становятся частью глубоководных отложений и в течение многих лет перерабатываются животными и микробами, питающимися илом. [ 31 ]
Кит падает
[ редактировать ]Для глубоководной экосистемы смерть кита является важнейшим событием. Мертвый кит может принести на дно сотни тонн органики. Сообщество падения китов проходит три стадии: [ 32 ]
- Стадия мобильного падальщика: Крупные и подвижные глубоководные животные прибывают на место практически сразу после падения китов на дно. Амфиподы , крабы , полярные акулы и миксины — падальщики.
- Оппортунистическая стадия: прибывают организмы, которые колонизируют кости и окружающие их отложения, загрязненные органическими веществами туши и любых других тканей, оставленных падальщиками. Один род — Osedax , [ 33 ] трубчатый червь. Личинка рождается без пола. Окружающая среда определяет пол личинки. Когда личинка поселяется на китовой кости, она превращается в самку; когда личинка поселяется на самке или внутри нее, она превращается в карликового самца. Одна самка оседакса может нести в своем яйцеводе более 200 особей мужского пола.
- Сульфофильная стадия: на этой стадии происходит дальнейшее разложение костей и сульфатредукция морской воды. Бактерии создают богатую сульфидами среду, аналогичную гидротермальным источникам. Сюда заселяются полиноиды, двустворчатые моллюски, брюхоногие моллюски и другие серолюбивые существа.
Хемосинтез
[ редактировать ]Гидротермальные источники
[ редактировать ]
Гидротермальные источники были открыты в 1977 году учеными из Океанографического института Скриппса. На данный момент все обнаруженные гидротермальные источники расположены на границах плит: Восточно-Тихоокеанской, Калифорнийской, Срединно-Атлантического хребта, Китайской и Японской.
Новый материал океанского бассейна образуется в таких регионах, как Срединно-Атлантический хребет, по мере того, как тектонические плиты отходят друг от друга. Скорость распространения плит составляет 1–5 см/год. Холодная морская вода циркулирует вниз через трещины между двумя плитами и нагревается, проходя через горячую породу. Минералы и сульфиды растворяются в воде при взаимодействии с породой. В конце концов, горячие растворы исходят из активного разлома под морским дном, создавая гидротермальный источник.
Хемосинтез бактерий обеспечивает энергию и органическое вещество для всей пищевой сети в жерловых экосистемах. Эти вентиляционные отверстия выделяют очень большое количество химических веществ, которые эти бактерии могут превращать в энергию. Эти бактерии также могут расти независимо от хозяина и создавать маты бактерий на морском дне вокруг гидротермальных источников, где они служат пищей для других существ. Бактерии являются ключевым источником энергии в пищевой цепи. Этот источник энергии создает большие популяции в районах вокруг гидротермальных источников, что дает ученым легкую остановку для исследований. Организмы также могут использовать хемосинтез для привлечения добычи или партнера. [ 34 ] Гигантские трубчатые черви могут вырасти до 2,4 м (7 футов 10 дюймов) в высоту из-за богатства питательных веществ. В гидротермальных жерлах обнаружено более 300 новых видов. [ 35 ]
Гидротермальные источники представляют собой целые экосистемы, независимые от солнечного света, и могут быть первым свидетельством того, что Земля может поддерживать жизнь без Солнца.
Холодные просачивания
[ редактировать ]Холодное просачивание (иногда называемое холодным жерлом) — это область дна океана , где происходит просачивание сероводорода , метана и других богатых углеводородами жидкостей, часто в виде лужи рассола .
Экология
[ редактировать ]Глубоководные пищевые сети сложны, и аспекты этой системы плохо изучены. Обычно взаимодействие хищник-жертва на глубине определяется путем прямого наблюдения (вероятно, с дистанционно управляемых подводных аппаратов ), анализа содержимого желудка и биохимического анализа. Анализ содержимого желудка является наиболее распространенным методом, но для некоторых видов он ненадежен. [ 36 ]
В глубоководных пелагических экосистемах у берегов Калифорнии в трофической сети преобладают глубоководные рыбы , головоногие моллюски , студенистый зоопланктон и ракообразные . В период с 1991 по 2016 год 242 уникальных пищевых взаимоотношения между 166 видами хищников и жертв продемонстрировали, что студенистый зоопланктон оказывает экологическое воздействие, аналогичное воздействию крупных рыб и кальмаров. наркомедузы , сифонофоры (семейства Physonectae ), гребневики и головоногие моллюски в порядке убывания. Наибольшее разнообразие добычи потребляли [ 36 ] Каннибализм зафиксирован у кальмаров рода Gonatus . [ 37 ]
Глубоководная добыча полезных ископаемых имеет серьезные последствия для океанских экосистем. Разрушение среды обитания, нарушение слоев отложений и шумовое загрязнение угрожают морским видам. [ 38 ] Важное биоразнообразие может быть утрачено с непредсказуемыми последствиями для пищевой цепи. Кроме того, могут выделяться токсичные металлы и химические вещества, что приводит к долгосрочному загрязнению морской воды. [ 39 ] Это поднимает вопросы об устойчивости и экологических издержках такой деятельности.
Глубоководные исследования
[ редактировать ]
Люди исследовали менее 4% дна океана, и при каждом погружении обнаруживаются десятки новых видов глубоководных существ. Подводная лодка DSV Alvin, принадлежащая ВМС США и управляемая Океанографическим институтом Вудс-Хоул (WHOI) в Вудс-Хоул, штат Массачусетс, является примером типа корабля, используемого для исследования глубокой воды. Эта 16-тонная подводная лодка способна выдерживать экстремальное давление и легко маневрирует, несмотря на свой вес и размеры.
Чрезвычайная разница в давлении между морским дном и поверхностью делает выживание существ на поверхности практически невозможным; это затрудняет углубленное исследование, поскольку большую часть полезной информации можно найти только пока существа живы. Последние разработки позволили ученым рассмотреть этих существ более внимательно и в течение более длительного времени. Морской биолог Джеффри Дрейзен нашел решение: ловушку для рыбы под давлением. Он захватывает глубоководное существо и медленно доводит его внутреннее давление до уровня поверхности по мере того, как существо поднимается на поверхность, в надежде, что существо сможет приспособиться. [ 40 ]
Другая научная группа из Университета Пьера и Марии Кюри разработала устройство захвата, известное как PERISCOP , которое поддерживает давление воды при ее поверхности, тем самым удерживая образцы под давлением во время подъема. Это позволяет проводить тщательное исследование поверхности без каких-либо возмущений давления, влияющих на образец. [ 41 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Служба управления минеральными ресурсами региона OCS Мексиканского залива (ноябрь 2006 г.). «Продажи нефти и газа в аренду OCS в Мексиканском заливе: 2007–2012 гг. Продажи в западной зоне планирования 204, 207, 210, 215 и 218. Продажи в зоне центрального планирования 205, 206, 208, 213, 216 и 222. Проект воздействия на окружающую среду Заявление, том I: главы 1–8 и приложения». Министерство внутренних дел США, Служба управления минеральными ресурсами, регион OCS Мексиканского залива, Новый Орлеан. стр. 3-27. PDF , заархивировано 26 марта 2009 г. в Wayback Machine.
- ^ «Робот-субмарина достигает самой глубокой глубины океана» . Новости Би-би-си . 3 июня 2009 года . Проверено 3 июня 2009 г.
- ^ Jump up to: а б Морской центр Гавайского университета (4 июня 2009 г.). «Ежедневные отчеты НИС KILO MOANA за июнь и июль 2009 г.» . Гонолулу, Гавайи: Гавайский университет. Архивировано из оригинала 19 сентября 2009 года . Проверено 24 июня 2010 г.
- ^ Морской центр Гавайского университета (4 июня 2009 г.). «Инвентаризация научного оборудования на борту НИС KILO MOANA» . Гонолулу, Гавайи: Гавайский университет. Архивировано из оригинала 13 июня 2010 года . Проверено 18 июня 2010 г.
- ^ Jump up to: а б К.Л. Смит-младший; Х.А. Руль; Би Джей Бетт; ДСМ Биллетт; Р. С. Лэмпитт; Р.С. Кауфманн (17 ноября 2009 г.). «Климат, круговорот углерода и глубоководные экосистемы» . ПНАС . 106 (46): 19211–19218. Бибкод : 2009PNAS..10619211S . дои : 10.1073/pnas.0908322106 . ПМК 2780780 . ПМИД 19901326 .
- ^ Jump up to: а б с д Хорхе Сирке (1997). «II. Пределы продуктивности моря» (PDF) . В Эдварде А. Лоусе (ред.). Эль-Ниньо и промысел перуанского анчоуса (серия: Программа обучения глобальным изменениям) . Том. 9. Саусалито: Университетские научные книги. стр. 118–121. дои : 10.1023/А:1008801515441 . ISBN 978-0-935702-80-4 . S2CID 29314639 . Архивировано из оригинала (PDF) 10 июня 2011 года . Проверено 18 июня 2010 г.
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Jump up to: а б «Фотическая зона» . Британская энциклопедия . 2010 . Проверено 18 июня 2010 г.
- ^ Jump up to: а б с д Жананда Кол (2004). «Сумеречная океанская (дисфотическая) зона» . EnchantedLearning.com . Проверено 18 июня 2010 г.
- ^ Jump up to: а б с Кен О. Бюсселер; Карл Х. Ламборг; Филип В. Бойд; Фиби Дж. Лам; и др. (27 апреля 2007 г.). «Возвращение к потоку углерода через сумеречную зону океана». Наука . 316 (5824): 567–570. Бибкод : 2007Sci...316..567B . CiteSeerX 10.1.1.501.2668 . дои : 10.1126/science.1137959 . ПМИД 17463282 . S2CID 8423647 .
- ^ Национальное управление океанических и атмосферных исследований (2 декабря 2008 г.). «Насколько глубок океан?» . Вашингтон, округ Колумбия: Национальное управление океанических и атмосферных исследований . Проверено 19 июня 2010 г.
- ^ «Глубокое море на MarineBio.org – биология океана, морская жизнь, морские существа, охрана морской среды» . Архивировано из оригинала 6 января 2009 г. Проверено 8 мая 2011 г.
- ^ Нибаккен, Джеймс В. Морская биология: экологический подход. Пятое издание. Бенджамин Каммингс, 2001. с. 136–141.
- ^ Пол Р. Пине (1996). Приглашение к океанографии (3-е изд.). Сент-Пол, Миннесота: West Publishing Co., с. 202. ИСБН 978-0-314-06339-7 .
- ^ Пинет 1996, с. 206.
- ^ Пинет 1996, стр. 206–207.
- ^ Пинет 1996, с. 207.
- ^ Исследование NOAA бассейна с рассолом
- ^ «Средневодная зона» . Водная жизнь мира . Том. 6. Тэрритаун, Нью-Йорк: Корпорация Маршалл Кавендиш. 2001. стр. 340–341. ISBN 978-0-7614-7176-9 .
- ^ Боун, Квентин; Мур, Ричард (2008). Биология рыб . Гирляндная наука. п. 38. ISBN 978-0-203-88522-2 .
- ^ Jump up to: а б с д и Мойл, ПБ; Чех, Джей-Джей (2004). Рыбы, Введение в ихтиологию (5-е изд.). Бенджамин Каммингс. ISBN 978-0-13-100847-2 .
- ^ Дуглас, Э.; Фридл, В.; Пиквелл, Г. (1976). «Рыбы в зонах с минимумом кислорода: характеристики оксигенации крови». Наука . 191 (4230): 957–9. Бибкод : 1976Sci...191..957D . дои : 10.1126/science.1251208 . ПМИД 1251208 .
- ^ Мунц, WRA (2009). «О желтых линзах у мезопелагических животных». Журнал Морской биологической ассоциации Соединенного Королевства . 56 (4): 963–976. дои : 10.1017/S0025315400021019 . S2CID 86353657 .
- ^ Jump up to: а б «Батипелагическая зона» . Слои океана . Национальная метеорологическая служба. Архивировано из оригинала 7 февраля 2017 года . Проверено 1 января 2021 г.
{{cite web}}: CS1 maint: bot: исходный статус URL неизвестен ( ссылка ) - ^ Jump up to: а б Эниг, CC (1997). Исследования морского бентоса . Испанский институт океанографии (на испанском языке). Мадрид: Министерство сельского хозяйства. стр. 23–33. ISBN 978-84-491-0299-8 .
- ^ Райан, Пэдди (21 сентября 2007 г.). «Глубоководные существа: Батипелагическая зона» . Те Ара – Энциклопедия Новой Зеландии . Проверено 4 сентября 2016 г.
- ^ Янси, Пол Х.; Герринджер, Маккензи Э.; Дразен, Джеффри С.; Роуден, Эшли А.; Джеймисон, Алан (25 марта 2014 г.). «Морские рыбы могут быть биохимически ограничены в заселении самых глубоких океанских глубин» . Труды Национальной академии наук . 111 (12): 4461–4465. Бибкод : 2014PNAS..111.4461Y . дои : 10.1073/pnas.1322003111 . ISSN 0027-8424 . ПМЦ 3970477 . ПМИД 24591588 .
- ^ «Исследователь океана NOAA: История: Цитаты: зондирования, морское дно и геофизика» . НОАА, Управление по исследованию и исследованию океана . Проверено 4 сентября 2016 г.
- ^ Смит, Крейг Р.; де Лео, Фабио К.; Бернардино, Анджело Ф.; Свитман, Эндрю К.; Арбису, Педро Мартинес (2008). «Глубинное ограничение пищи, структура экосистемы и изменение климата» (PDF) . Тенденции экологии и эволюции . 23 (9): 518–528. дои : 10.1016/j.tree.2008.05.002 . ПМИД 18584909 . Архивировано из оригинала (PDF) 20 июля 2011 г. Проверено 4 сентября 2016 г.
- ^ Виноградова Н.Г. (1997). «Зоогеография абиссальной и хадальской зон». Биогеография океанов . Достижения морской биологии. Том. 32. С. 325–387. дои : 10.1016/S0065-2881(08)60019-X . ISBN 978-0-12-026132-1 .
- ^ «Морской снег и фекальные гранулы» .
- ^ Пир и голод на бездонной равнине.
- ^ Шана Гоффреди, Необычная донная фауна, связанная с падением китов в каньоне Монтерей, Калифорния, Deep-Sea Research, 1295–1304, 2004 г.
- ^ Ной К. Уайтман, Между китовой костью и глубоким синим морем: происхождение карликовых самцов трубчатых червей, питающихся китовыми костями, Молекулярная экология, 4395–4397, 2008 г.
- ^ Хемосинтез
- ^ Ботос, Соня. «Жизнь на гидротермальном источнике» .
- ^ Jump up to: а б Чой, К. Анела; Хэддок, Стивен HD; Робисон, Брюс Х. (6 декабря 2017 г.). «Структура глубоководной пелагической пищевой сети, выявленная наблюдениями за питанием in situ » . Учеб. Р. Сок. Б. 284 (1868): 20172116. doi : 10.1098/rspb.2017.2116 . ПМК 5740285 . ПМИД 29212727 .
- ^ Кляйн, Джоанна (19 декабря 2017 г.). «Что что ест: Путеводитель по глубоководным обедам для сухопутных жителей» . Нью-Йорк Таймс . ISSN 0362-4331 . Архивировано из оригинала 20 декабря 2017 года . Проверено 20 декабря 2017 г.
- ^ Довер, CL (2017). «Воздействие глубоководной добычи полезных ископаемых на окружающую среду». Наука . 359 (6377): 34–38.
- ^ Левин, Луизиана (2020). «Глубоководная добыча полезных ископаемых: оценка воздействия на окружающую среду». Ежегодный обзор морской науки . 12 :19–43.
- ^ Новая ловушка может безопасно вытащить глубоководную рыбу из темноты
- ^ Левер А.М. (31 июля 2008 г.). «Живая рыба, пойманная на рекордной глубине» . Новости Би-би-си . Проверено 18 февраля 2011 г.
Дальнейшее чтение
[ редактировать ]- Куприянова Е.К.; Винн, О.; Тейлор, PD; Шопф, Дж.В.; Кудрявцев А.Б.; Бэйли-Брок, Дж. (2014). «Серпулиды, живущие глубоко: известковые трубчатые черви за бездной». Глубоководные исследования . Часть I. 90 : 91–104. Бибкод : 2014DSRI...90...91K . дои : 10.1016/j.dsr.2014.04.006 .

