Экосистема поверхности океана
| Морская среда обитания |
|---|
 |
| Прибрежные места обитания |
| Поверхность океана |
| Открытый океан |
| Морское дно |
Организмы, свободно живущие на поверхности океана, называемые нейстоном , включают ключевые организмы, такие как золотые водоросли Sargassum , составляющие Саргассово море , плавающие ракушки , морские улитки , голожаберные и книдарии . Многие экологически и экономически важные виды рыб живут за счет нейстона или полагаются на него. Виды на поверхности распределены неравномерно; Поверхность океана обеспечивает среду обитания для уникальных нейстонных сообществ и экорегионов, встречающихся только на определенных широтах и только в определенных океанских бассейнах. Но поверхность также находится на переднем крае изменения климата и загрязнения. Жизнь на поверхности океана соединяет миры. От мелководья до глубокого моря, от открытого океана до рек и озер – многочисленные наземные и морские виды зависят от поверхностной экосистемы и обитающих там организмов. [1]
Поверхность океана действует как оболочка между атмосферой вверху и водой внизу и содержит уникальную для этой среды экосистему. Эту залитую солнцем среду обитания можно определить как глубину примерно одного метра, поскольку почти половина УФ-B ослабляется в пределах этого первого метра. [2] Организмам здесь приходится бороться с волновым воздействием и уникальными химическими веществами. [3] [4] [5] и физические свойства. [6] Поверхность используется широким спектром видов, от различных рыб и китообразных до видов, которые передвигаются по океанскому мусору (так называемые стропила). [7] [8] [9]
Самое главное, что на поверхности обитает уникальное сообщество свободноживущих организмов, называемое нейстон (от греческого слова υεω, что означает одновременно плавать и плавать). Плавающие организмы также иногда называют плейстоном , хотя чаще используется нейстон. Несмотря на разнообразие и важность поверхности океана для соединения разрозненных сред обитания, а также риски, с которыми она сталкивается, о нейстонной жизни известно немного. [1]
Обзор
[ редактировать ]Нейстон является ключевым экологическим звеном, соединяющим экосистемы, начиная от коралловых рифов , островов, глубокого моря и даже пресноводных мест обитания. В северной части Тихого океана рацион головастых черепах на 80% состоит из добычи нейстона. [10] и почти 30% рациона лейсанского альбатроса составляет нейстон. [11] Разнообразные виды пелагических и рифовых рыб в молодом возрасте обитают на поверхности воды. [12] включая коммерчески важные виды рыб, такие как атлантическая треска , лосось и марлин . Нейстон может быть сконцентрирован в виде живых островов, полностью закрывающих морскую гладь, или разбросан по редким лугам на тысячи миль. Однако роль нейстонов, а во многих случаях и само их существование, часто упускается из виду. [1]
Одним из наиболее известных поверхностных экорегионов является Саргассово море , экологически особый регион, заполненный густыми нейстонными бурыми водорослями в Северной Атлантике. Множество экологически и коммерчески важных видов зависят от Саргассова моря, но нейстонная жизнь существует в каждом океанском бассейне и может играть аналогичную, хотя и непризнанную, роль в регионах по всей планете. Например, более 50 лет назад советский учёный А. И. Савилов охарактеризовал 7 нейстонных экорегионов в Тихом океане. [13] Каждый экорегион обладает уникальным сочетанием биотических и абиотических условий и содержит уникальное сообщество нейстонных организмов. Однако эти экорегионы в значительной степени забыты. [1]
Но есть и другая причина изучать нейстон: поверхность океана находится на передовой линии антропогенного воздействия: от изменения климата до загрязнения, от разливов нефти до пластика. Поверхность океана сильно пострадала от антропогенных изменений, и поверхностная экосистема, вероятно, уже кардинально отличается от той, что была всего несколько сотен лет назад. Например, до повсеместного строительства плотин, лесозаготовок и индустриализации в открытый океан могло попасть больше древесины. [14] тогда как пластик еще не был изобретен. А поскольку плавучая жизнь обеспечивает пищу и убежище для различных видов, изменения в поверхностной среде обитания вызовут изменения в других экосистемах и будут иметь последствия, которые в настоящее время не полностью поняты и не могут быть предсказаны. [1]
 Поверхность океана занимает 72% общей поверхности Земли. Их можно разделить на поверхности относительно мелких и богатых питательными веществами прибрежных районов над континентальными шельфами (светло-голубой) и поверхности более обширного и относительно глубокого, но бедного питательными веществами океана, лежащего за ними (темно-синий).
Поверхность океана занимает 72% общей поверхности Земли. Их можно разделить на поверхности относительно мелких и богатых питательными веществами прибрежных районов над континентальными шельфами (светло-голубой) и поверхности более обширного и относительно глубокого, но бедного питательными веществами океана, лежащего за ними (темно-синий).

| Внешние видео | |
|---|---|
«Незадолго до наступления темноты, когда они миновали огромный остров, заросший саргассовой травой, который вздымался и раскачивался в светлом море, как будто океан занимался любовью с чем-то под желтым одеялом, его маленькую леску перехватил дельфин». — Эрнест Хемингуэй, «Старик и море».

 Саргассово море
Саргассово море


Жизнь на поверхности океана (нейстон)
[ редактировать ]Вызывая образы поверхности открытого океана, воображение может представить себе бесконечное пустое пространство. Плоская линия, отделяющая синий цвет внизу от синего цвета вверху. Но на самом деле этот уникальный пограничный слой населяет множество разнообразных видов. Для разных организмов, занимающих разные ниши на поверхности океана, существует клубок терминов. Для обозначения всех из них здесь используется наиболее обширный термин нейстон. [1]
Нейстонные животные и растения живут, свисая с поверхности океана, словно подвешенные к крыше массивной пещеры, и не способны контролировать направление своего движения. Они считаются постоянными жителями поверхностного слоя. Многие роды распространены по всему миру. Многие организмы обладают морфологическими особенностями, позволяющими им оставаться на поверхности океана, причем наиболее заметными адаптациями являются поплавки. [1]
Поплавки (плейстон)
[ редактировать ]| Поплавки, иногда называемые плейстонами , — это организмы, обитающие на поверхности океана. [1] | ||
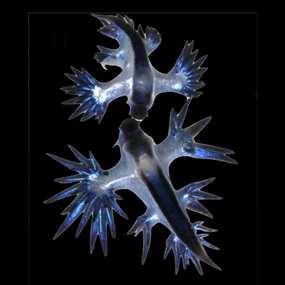
| Книдарийцы (медуза) | Веллелла , Порпита , Физалия и Актинекта. На поверхности океана обитают многочисленные плавающие книдарии (медузы), некоторые из которых известны (или печально известны), а другие редко встречаются. Такие виды, как Velella sp. (моряк по ветру) и Porpita sp. (синяя кнопка) занимают центральное место в поверхностной пищевой сети . В их тканях имеются симбиотические динофлагелляты, и, как и их двоюродные братья из бентосных кораллов, эти симбионты могут позволять им выживать в олиготрофных водах. Velella и Porpita — единственные два рода хондрофорной клады внутри Hydrozoa , которые, вероятно, эволюционировали конвергентно с другим нейстонным родом Hydrozoa: Physalia (португальский военный корабль). И Физалия , и Веллелла представляют собой «паруса», которые позволяют им путешествовать в зависимости от направления ветра. [15] Эти моряки, путешествующие по ветру, плавают у поверхности океана, свисая щупальцами внизу, в воде. Веллелла имеет приподнятый прозрачный «парус» на синем овальном диске. Внизу от диска свисают короткие бахромчатые щупальца. Движение осуществляется за счет ветра, ударяющего в парус. Некоторые Velella имеют правый парус, а некоторые — левый, что гарантирует, что их всех не унесет в одном направлении одновременно. [16] Физалии также используют висячие щупальца, которые служат морским якорем в открытом океане. [17] и нанеси мощное жало. Морские анемоны рода Actinecta встречаются редко, но они также плавают под водой на поверхности океана, подобно Porpita , но используют пузырьковый поплавок на педальном диске. [1]
| |
|---|---|---|
| Моллюски (морские улитки) | Моллюски Джантина , Реклюция и Главкус. Улитки, сплавляющиеся по пузырькам, Recluzia и Janthina строят плавающие плоты, погружая переднюю ногу в поверхность воды и окутывая захваченный воздух слоем слизи, образуя пузырь, который затем прилипают к плоту. Загадочная реклюция питается актинией Actinecta , оба вида коричнево-желтого цвета. Напротив, фиолетовые улитки Janthina охотятся на Velella , Porpita и иногда Physalia [3], хотя они не могут двигаться или охотиться. Вместо этого Джантина полагаются на пассивный контакт со своей добычей.Другие виды включают голожаберного Glaucus (синий морской дракон), который также питается плавающими гидрозойными животными. [18] и глотает воздух, чтобы оставаться на плаву. Существует несколько загадочных видов Glaucus . [19] и виды этого рода могут демонстрировать высокую степень региональной изоляции. [20] [1]
| |
| ракообразное |
Единственная по-настоящему нейстонная ракушка, Dosima fasccularis (ракушка-буй), живет на поверхности океана, сначала прикрепляясь к плавающим объектам в виде личинок (например, перьев) и выделяя воздушный подушеобразный поплавок, а не обычный твердый цемент, используемый другими ракушками. [21] [22] Этот поплавок позволяет ракушкам Досима в конечном итоге перерасти свой личиночный дом и дрейфовать самостоятельно. [1] | |
| Макроводоросли (водоросли) |
Нейстонные водоросли, такие как Sargassum fluitans и Sargassum natans, имеют многочисленные наполненные газом поплавки, которые остаются на поверхности океана. Эти водоросли создают среду обитания для множества видов, связанных с саргассумами , особенно на западной окраине североатлантического субтропического круговорота , известного как Саргассово море . [23] В Тихом океане род водорослей Turbinaria размножается плавающими фронтами. [24] Кроме того, на поверхности было обнаружено более 20 видов водорослей и восемь видов морской травы. [7] [1] | |
| Микроорганизмы (бактерии, простейшие и т. д.) |  триходезмиума Цветение Разнообразные микроорганизмы населяют поверхностную экосистему океана. [25] [26] [27] [28] [10] и может играть значительную роль в газообмене между океаном и атмосферой. [29] Многие из этих организмов были идентифицированы в микрослое морской поверхности , который, в зависимости от определения, простирается на 100–1000 мкм ниже поверхности океана. [25] Поверхность океана обладает уникальными химическими и физическими свойствами, которые могут способствовать концентрации видов, специально адаптированных к этим условиям. Например, бактерионейстон, обитающий в микрослое морской поверхности, часто бывает ярко окрашенным, [30] возможно, в качестве защиты от солнечной радиации. В поверхностном микрослое могут в значительной степени доминировать гетеротрофные организмы, включая как бактерии, так и микроэукариоты, которые используют преимущества связанных с поверхностью соединений. [27] Другие виды могут выходить за пределы микрослоя морской поверхности, но все еще связаны с поверхностью, включая экологически важную цианобактерию Trichodesmium . [31] Тем не менее, как и в случае с более крупными организмами, поверхностные микроорганизмы, как правило, мало изучены. [10] [1] | |















Эпинеустон
[ редактировать ]| Эпинеустон — это организмы, которые используют поверхностное натяжение воды, чтобы удерживаться на поверхности океана. [1] | ||
| Насекомые |  Агрегация морских конькобежцев [32] Морских насекомых очень мало. [33] Единственными настоящими насекомыми открытого океана являются Halobates . На поверхности воды обитают эпинейстонные организмы, а в открытом океане все эпинейстонные виды относятся к роду насекомых Halobates . Известные как «морские конькобежцы», Halobates sp. охотятся на другие нейстонные виды и зоопланктон, пойманные в ловушкуповерхность. [34] Галобаты откладывают яйца на различные предметы, в том числе на плавающие перья, дерево, пластик и т. д. [35] и что необычно для пелагических моллюсков, таких как Atlanta turriculata . [34] [1] | |
|---|---|---|
Гипонейстон
[ редактировать ]| Гипонейстон — это мобильные организмы, живущие непосредственно под поверхностью воды. [1] | ||
| Копеподы |  На поверхности океана можно встретить самые разнообразные веслоногие ракообразные. [36] [37] [38] [39] [40] [41] [42] Некоторые нейстонические копеподы обладают замечательными адаптациями, особенно среди веслоногих понтеллид . Адаптации понтеллид включают специализированные структуры прикрепления к поверхности, [43] синяя пигментация. [36] [40] и даже поведение в полете, чтобы спастись от хищников. [41] Копеподы Sapphirinidae часто также связаны с поверхностью, [37] а некоторые виды имеют невероятную структурную окраску. [42] Как и во многих морских экосистемах, копеподы представляют собой основной источник пищи для множества нейстонных и обитающих на поверхности видов. [1] Микрослой морской поверхности (SML) на границе раздела воздух-море представляет собой особую, недостаточно изученную среду обитания по сравнению с недрами, и копеподы, важные компоненты пищевых сетей океана, разработали ключевые приспособления для использования этой ниши. [40] Охватывающая океан SML образует границу между атмосферой и гидросферой . Несмотря на толщину менее одного миллиметра, SML имеет совершенно другие физико-химические и биологические характеристики, чем подстилающая вода (ULW). [44] SML обеспечивает биогенный желатиновый каркас. [4] и обычно обогащен органическими веществами, [45] гетеротрофные микроорганизмы [26] а также организмы более высокого трофического уровня. [46] [40] Среди таксонов зоопланктона, обитающих в пределах SML, в тропических районах всех океанов часто отмечаются нейстонные копеподы (тип Arthropoda, класс Crustacea) семейства Pontellidae. [47] [48] [49] SML считается сложной или даже экстремальной средой обитания, поскольку организмы подвергаются воздействию переменных температур и высокой интенсивности солнечного и ультрафиолетового (УФ) излучения. [50] Копеподы — самые многочисленные многоклеточные существа на Земле. [51] и демонстрируют впечатляющую краткосрочную адаптацию к стрессовым факторам окружающей среды, например, подавление клеточной реакции на тепловой стресс. [52] Учитывая их важную роль в морских пищевых цепях и функционировании экосистем, [53] Знание пределов толерантности веслоногих ракообразных к абиотическим факторам имеет важное значение, если мы хотим сделать надежные прогнозы последствий глобальных изменений в Мировом океане. Последствия вызванного климатом потепления (и закисления) на экосистему SML и веслоногих ракообразных, обитающих в нейстоне, хотя на сегодняшний день почти не изучены, могут быть особенно драматичными. [40] Особенностью многих копепод понтеллид является их синяя окраска, которая встречается и у другого поверхностного мезозоопланктона. [54] Окраска обусловлена комплексом пигмента астаксантина и каротинопротеина. [55] Астаксантин может производиться из пищевых источников, и было обнаружено, что он является основным каротиноидом в четырех различных родах синепигментированных веслоногих ракообразных, а также в Oikopleura dioica класса Appendularia, что указывает на конвергентную эволюцию этого признака у разных обитателей нейстона. [56] Были разработаны различные теории, объясняющие значение синей окраски веслоногих, включая защиту от сильного солнечного и / или УФ-излучения. [57] [58] маскировка от визуальных хищников, кормящихся в самых верхних слоях воды [54] а также распознавание особей своего вида, когда они встречаются вместе с копеподами, имеющими окраску на основе зеленого флуоресцентного белка (GFP). [59] [40] | |
|---|---|---|
| изоподы |  Изопода Idotea Metallica Idotea Metallica — это замечательная изопода , связанная с поверхностью , которую можно найти либо плавающей вверх тормашками на поверхности океана. [1] или прикреплены к плавающим обломкам или нейстону (например, к пузырчатым плотам Джантины ). Обычно это связано с обломками. [60] и способен активно переплывать от одного плавучего объекта к другому. Этот вид якобы встречается по всему миру в теплых водах, хотя, как и в случае со многими видами, связанными с поверхностью, информация о его генетическом разнообразии скудна. Из-за смены течений он часто переносится в более северные регионы. Для людей, прибывающих в летние месяцы в Гельголанд (Германия; Северное море), основная термическая ниша составляет 16 °C, при этом самая низкая переносимая температура, вероятно, составляет около 13 °C. [61] Однако к этим пределам термоустойчивости следует относиться с осторожностью: как и многие нейстонные виды, I. Metallica плохо изучен, и неясно, действительно ли это один вид или множество загадочных видов. [1] | |
| Креветка |
Несколько видов креветок связаны с плавающими саргассумами , а также их можно встретить плавающими на поверхности, в том числе Latreutes fucorum и Hippolyte coerulescens . Нейстонические креветки демонстрируют удивительное разнообразие цветовых узоров. [62] включая обычный нейстонический синий цвет с хромофорами, которые могут реагировать на изменение условий освещения. [63] [1] | |
| Рыба |
Удивительное разнообразие рыб проводит свою молодую жизнь на поверхности океана. В этот список вошли многие известные, экологически и экономически важные виды из самых разных мест обитания. Пелагические виды рыб включают некоторые анчоусы , махи-махи , марлинов , рыбу-меч , янтаря и атлантическую скумбрию . Хорошо известные и экологически важные донные рыбы в молодом возрасте общаются с поверхностью, в том числе такие виды: левоглазая камбала , морская собачка , бычок , морские коньки , морские драконы и игла . К глубоководным рыбам с поверхностными личинками относятся рыбы-гадюки и рыбы-фонарики . Многие угри , как рифовые, донные, так и глубоководные, в ночное время мигрируют в поверхностный слой в виде личинок. [64] Но хотя поверхность океана может показаться странной средой обитания для личинок глубоководных рыб, они далеко не самые необычные. Разнообразные рыбы, которые мигрируют в пресную воду во взрослом возрасте (либо в качестве среды обитания, либо нерестилища), в молодом возрасте полагаются на нейстон. Годовалые и сеголетки лососей разных видов поедают нейстонную добычу в северном Калифорнийском течении . [65] Американские европейцы выплывают из своих пресноводных рек и сходятся в середине Северной Атлантики, чтобы нереститься в Саргассовом море. Некоторые рыбы всю жизнь занимают поверхность океана и даже способны парить над волнами, в том числе летучие рыбы и полуклювы . Другие часто посещают поверхность океана, в том числе такие виды, как солнечная рыба и гигантские акулы . [1] | |
| Головоногие моллюски |
Хотя ни одно головоногие не привязано к поверхностному слою постоянно, некоторые из них часто посещают поверхностную среду обитания и приспособлены к ее использованию. осьминогов Самки -аргонавтов ( Argonauta spp.) опускают свой похожий на бумагу панцирь в воздух, улавливая пузырьки газа, которые затем используют для поддержания плавучести. [66] [67] Разнообразные летающих кальмаров виды Ommastrephidae и Onychoteuthidae могут выпрыгивать из воды и парить на впечатляющие расстояния, некоторые могут достигать высоты более трех метров, а другие могут плавать на расстояния до 55 метров. [68] | |





Рафтинговые организмы
[ редактировать ]| Сплавляющиеся виды живут либо прикрепленными к нейстонным организмам (например, ракушкам, которые поселяются на раковинах Джантины), либо к неодушевленным отбросам. | ||
| ракушки (инкрустирует) |
Сплавляющиеся виды живут либо прикрепленными к нейстонным организмам (например, ракушкам , которые поселяются на Джантины раковинах ), либо к неодушевленным отбросам. Некоторые рафтинговые виды эволюционировали, чтобы жить на мусоре на поверхности океана, например, гладкая ракушка-гусиная шея Lepas anatifera , в то время как другие могут быть прибрежными видами, которые селятся на прибрежных плавающих обломках и затем переносятся течениями в открытый океан. Несколько отличных обзоров посвящены биологии стропил, в том числе плавающему основанию стропил. [7] рафтинг-сообщество, [8] и биогеографические и эволюционные последствия рафтинга. [9] [1] | |
|---|---|---|

Поверхностный микрослой
[ редактировать ]
(II) Отдельные микробные сообщества, перерабатывающие растворенные и твердые органические вещества. [71]
(III) Максимальное воздействие солнечной радиации запускает фотохимические реакции и образование радикалов. [72]
Микрослой морской поверхности (СМЛ) является границей между атмосферой и океаном, покрывая около 70% поверхности Земли. При эксплуатационной толщине от 1 до 1000 мкм SML обладает физико-химическими и биологическими свойствами, которые заметно отличаются от нижележащих вод. Недавние исследования теперь показывают, что SML в значительной степени покрывает океан, и данные показывают, что это среда, обогащенная биопленками, с отдельными микробными сообществами. Благодаря своему уникальному положению на границе воздух-море, SML играет центральную роль в ряде глобальных биогеохимических и климатических процессов. [69]
Микрослой морской поверхности (СМЛ) является границей между атмосферой и океаном, покрывая около 70% поверхности Земли. SML имеет физико-химические и биологические свойства, которые заметно отличаются от подстилающих вод. Благодаря своему уникальному положению на границе воздух-море, SML играет центральную роль в ряде глобальных биогеохимических и климатических процессов. Несмотря на то, что SML был известен в течение последних шести десятилетий, он часто оставался в отдельной исследовательской нише, прежде всего потому, что не считалось, что он существует в типичных океанических условиях. Недавние исследования теперь показывают, что SML в значительной степени покрывает океан. [73] подчеркивая его глобальную значимость как пограничного слоя, связывающего два основных компонента системы Земли – океан и атмосферу. [69]

В 1983 году Зибурт предположил, что SML представляет собой гидратированный гелеобразный слой, образованный сложной смесью углеводов, белков и липидов. [71] В последние годы его гипотеза была подтверждена, и научные данные указывают на то, что SML представляет собой биопленочную среду, обогащенную агрегатами, с отдельными микробными сообществами. [74] В 1999 году Эллисон и др. подсчитано, что 200 Тг С в год −1 накапливается в SML, аналогично скорости осаждения углерода на морском дне океана, хотя накопленный углерод в SML, вероятно, имеет очень короткое время пребывания. [75] Хотя общий объем микрослоя очень мал по сравнению с объемом океана, Карлсон в своей плодотворной статье 1993 года предположил, что в SML могут происходить уникальные межфазные реакции, которые могут не происходить в подстилающей воде или происходить там с гораздо меньшей скоростью. [70] Поэтому он предположил, что SML играет важную роль в диагенезе углерода в верхних слоях океана. [70] Свойства биопленки и максимально возможное воздействие солнечного излучения приводят к интуитивному предположению, что SML представляет собой биохимический микрореактор. [76] [69]
Исторически сложилось так, что SML представляет собой микросреду обитания, состоящую из нескольких слоев, отличающихся своими экологическими, химическими и физическими свойствами, с общей эксплуатационной толщиной от 1 до 1000 мкм. В 2005 году Хантер определил SML как «микроскопическую часть поверхностного океана, которая находится в контакте с атмосферой и может иметь физические, химические или биологические свойства, значительно отличающиеся от свойств соседних подземных вод». [77] Он избегает определенного диапазона толщины, поскольку он сильно зависит от интересующей особенности. Толщина 60 мкм была измерена на основе внезапных изменений pH. [78] и может быть значимо использован для изучения физико-химических свойств SML. При такой толщине СМЛ представляет собой ламинарный слой, свободный от турбулентности и сильно влияющий на газообмен между океаном и атмосферой. Как среда обитания нейстона (поверхностных организмов, от бактерий до более крупных сифонофоров), толщина SML в некотором смысле зависит от интересующего организма или экологических особенностей. В 2005 году Зайцев описал SML и связанный с ним приповерхностный слой (до 5 см) как инкубатор или питомник для яиц и личинок широкого спектра водных организмов. [37] [69]
Определение Хантера включает все взаимосвязанные слои от ламинарного слоя до питомника без явной ссылки на определенные глубины. [79] В 2017 году Вурл и др. предложил, чтобы определение Хантера было подтверждено с помощью переработанной парадигмы SML, которая включает его глобальное присутствие, свойства, подобные биопленке, и роль питомника. Новая парадигма выводит SML в новый и более широкий контекст, актуальный для многих наук об океане и климате. [69]
По данным Вурла и др. м СМЛ никогда не может быть лишена органики из-за обилия поверхностно-активных веществ (например, ПАВ) в верхних слоях океана. [73] и явление поверхностного натяжения на границе раздела воздух-жидкость. [80] SML аналогичен тепловому пограничному слою, а дистанционное зондирование температуры поверхности моря показывает повсеместные аномалии между оболочкой поверхности моря и температурой объема. [81] Несмотря на это, различия в обоих случаях обусловлены разными процессами. Обогащение, определяемое как отношение концентрации аналита в SML к основной объемной воде, десятилетиями использовалось в качестве доказательства существования SML. Следовательно, истощение органических веществ в SML является спорным; однако вопрос обогащения или истощения, вероятно, будет зависеть от толщины SML (которая меняется в зависимости от состояния моря); [82] включая потери из-за морских брызг, концентрацию органических веществ в основной воде, [73] и ограничения методов отбора проб для сбора тонких слоев. [83] Обогащение поверхностно-активных веществ, изменение температуры и солености поверхности моря служат универсальными индикаторами присутствия СМЛ. Организмы, возможно, менее подходят в качестве индикаторов SML, поскольку они могут активно избегать SML и/или суровые условия SML могут привести к сокращению их популяций. Однако толщина SML остается «рабочей» в полевых экспериментах, поскольку толщина собранного слоя определяется методом отбора проб. Достижения в технологии отбора проб SML необходимы для улучшения нашего понимания того, как SML влияет на взаимодействие воздуха и моря. [69]
Поверхностные пятна
[ редактировать ]
Слики — это извилистые линии гладкой воды на поверхности океана, которые являются повсеместными прибрежными объектами по всему миру. [85] Различные механизмы могут вызывать образование пятен, включая приливные и мысные фронты, и, как следствие, подземные волны, называемые внутренними волнами . [86] Внутренние волновые пятна образуются, когда внутренние волны взаимодействуют с крутым рельефом морского дна и создают области схождения и расхождения на поверхности океана. [87] Накопление органических материалов ( поверхностно-активных веществ ) на поверхности изменяет поверхностное натяжение, создавая гладкий вид, напоминающий масляное пятно . [88] Сходящийся поток может накапливать плотные скопления планктона, включая личинок рыб и беспозвоночных, на поверхности океана или под ней. [89] [90] [91] [92] [93] [94] [95] [96]
Поверхностные пятна являются центром многочисленных трофических и личиночных связей, которые лежат в основе функционирования морской экосистемы. [96] Жизнь многих морских организмов начинается у поверхности океана. Из плавучих яиц вылупляются планктонные личинки, которые развиваются и рассеиваются в океане на несколько недель или месяцев, прежде чем превратиться в молодь и в конечном итоге найти подходящую среду обитания для взрослых особей. [97] Пелагическая личиночная стадия объединяет популяции и служит источником новых взрослых особей. Океанические процессы, влияющие на судьбу личинок, оказывают глубокое влияние на пополнение популяции, связь и структуру экосистемы. [98] Хотя это важный этап жизни, по состоянию на 2021 год знания об экологии и поведении личинок ограничены. [96] Понимание биофизических взаимодействий, которые управляют выживанием и транспортировкой личинок рыб, имеет важное значение для прогнозирования и управления морскими экосистемами, а также рыболовством, которое они поддерживают. [99] [100] [96]

На диаграмме показано: (1) Личиночные и молодые стадии рыб из многих океанических местообитаний собираются в пятна, чтобы извлечь выгоду из плотных скоплений добычи (2 — фитопланктон, 3 — зоопланктон, 4 — личинки беспозвоночных, 5 — икра и 6, насекомые). Увеличение перекрытия хищник-жертва в пятнах увеличивает поток энергии, которая распространяется вверх по пищевой сети (пунктирные синие линии показывают трофические связи), увеличивая энергию, доступную хищникам более высокого трофического уровня (значки, обведенные синим цветом), включая людей. Более 100 видов рыб развиваются и растут в поверхностных питомниках перед переходом во взрослое состояние (сплошные белые линии, расходящиеся наружу) в коралловых рифах (7–12), эпипелагических (13–15) и глубоководных (16–17) океанах. места обитания. Во взрослом состоянии эти таксоны (значки, обведенные белым) играют важные экологические функции и обеспечивают рыбными ресурсами местное население. Например, прибрежные стайные рыбы (7, ставрида скумбрия) являются важной пищевой и наживочной рыбой для человека. Планктоноядные рыбы (8, некоторые стрекозы и спинороги) передают энергию от зоопланктона рифовым хищникам, таким как джеки (9), [101] которые обеспечивают контроль над рифами сверху вниз [102] и являются важным объектом для прибрежных рыбаков-любителей. [103] Пасущиеся (10, голавли) помогают предотвратить зарастание коралловых рифов макроводорослями. [104] Криптобентосные рыбы, такие как собачки (11) и донные макроракообразные (12, креветки, ротоногие, крабы), составляют большую часть потребляемой биомассы на рифах. [105] [106] В пелагическом океане летучие рыбы (13) передают энергию и питательные вещества от зоопланктона пелагическим хищникам, таким как махи-махи (14) и марлиновые (15), оба из которых используют пятна в качестве среды обитания. Личинки мезопелагических рыб, таких как рыба-фонарь (16) и батидемерсальные рыбы-треноги (17), используют эти поверхностные горячие точки перед тем, как спуститься в глубоководную среду обитания взрослых особей. [96]
Распределение добычи и хищников в океане неоднородно. [107] [108] Выживание личинок зависит от наличия добычи, хищничества и транспортировки в подходящую среду обитания, причем все это зависит от условий океана. [109] Океанские процессы, которые вызывают сходящиеся потоки, такие как фронты, внутренние волны и водовороты, могут структурировать планктон, усиливать перекрытие хищников и добычи и влиять на распространение личинок. [89] [110] [111] [112] [113] [114] [115] [116] Конвергентные характеристики также могут привести к каскаду эффектов, которые в конечном итоге влияют на структуру пищевой сети и повышают продуктивность экосистемы. [117] [96]
История жизни
[ редактировать ]
Истории жизни соединяют разрозненные экосистемы; Виды, обитающие на поверхности на одном этапе жизненного цикла, могут занимать глубоководные районы, бентос, рифы или пресноводные экосистемы на другом. Поверхность океана используют разнообразные виды рыб. [119] либо как взрослые особи, либо как среда обитания для яиц и молоди. Напротив, виды, плавающие на поверхности океана на одной стадии жизненного цикла, часто (хотя и не всегда) имеют пелагические личиночные стадии. Веллелла и Порпита выпускают медуз (медуз), [120] мало что известно и хотя о Porpita medusae , Velella medusae, возможно, могла погрузиться в более глубокие воды. [120] или остаются у поверхности, где получают питательные вещества от зооксантелл. [118] У Янтины есть пелагические личинки велигера, [121] и Physalia могут выделять репродуктивные кластеры, которые дрейфуют в толще воды. Галобаты откладывают яйца на различные объекты, в том числе на плавающие. [34] и пелагические раковины улиток. [122] [1]
Все виды, находящиеся на пелагических стадиях, в конечном итоге должны вернуться на поверхность. У Velella и Porpita личинки, образующиеся в результате полового размножения медуз, развивают небольшие поплавки, которые выносят их на поверхность. [123] [124] Для личинок Janthina переход к поверхностной жизни включает в себя деградацию их глаз и преддверной системы и в то же время образование внешней структуры, которая, как сообщается, представляет собой либо небольшой парашют из слизи, либо скопление пузырьков, которые они выносят на поверхность. [125] [126] Молодые Halobates могут вылупиться как над поверхностью, так и под ней, а для тех, кто находится ниже, поверхностное натяжение оказывается огромным барьером. может потребоваться Нимфам Halobates несколько часов, чтобы прорваться через поверхностную пленку. [34] Несмотря на трудности выхода на поверхность, временная пелагическая жизнь может принести пользу. [1]


(г) эти глубоководные места обитания могут позволить им использовать встречные течения для перемещения в направлении, противоположном поверхностным течениям (гипотеза, предложенная для Веллеллы ) [129]
Связности поверхностных экосистем океана может способствовать история жизни обитающих там видов. Одна из гипотез состоит в том, что у видов есть пелагические стадии, позволяющие «покинуть» области поверхностного стока и повторно заселить области поверхностного источника, где одна стадия жизненного цикла дрейфует под воздействием поверхностных течений в одном направлении, а пелагическая стадия либо остается географически локализованной. [130] или дрейфует в противоположном направлении. [131] Однако некоторые поверхностные виды, такие как эндемичные виды Саргассова моря, могут оставаться географически изолированными на протяжении всей своей жизненной истории. Хотя эти гипотезы интригуют, неизвестно, влияет ли история жизни на распределение популяций/видов для большинства нейстонных видов и каким образом. Понимание того, как история жизни варьируется в зависимости от вида, является важнейшим компонентом оценки как связности, так и сохранения нейстонных экосистем. [1]
Морские брызги
[ редактировать ]
Поток переносимых по воздуху микроорганизмов кружит над планетой над погодными системами, но под коммерческими воздушными путями. [132] Некоторые странствующие микроорганизмы переносятся земными пыльными бурями, но большинство происходит из морских микроорганизмов, содержащихся в морских брызгах . В 2018 году ученые сообщили, что сотни миллионов вирусов и десятки миллионов бактерий ежедневно оседают на каждом квадратном метре планеты. [133] [134]
Эти переносимые по воздуху микроорганизмы составляют часть аэропланктона . это крошечные формы жизни, которые плавают и дрейфуют в воздухе, увлекаемые потоками ветра Аэропланктон — ; они являются атмосферным аналогом океанического планктона . Большинство живых существ, составляющих аэропланктон, имеют размеры от очень маленьких до микроскопических , и многие из них сложно идентифицировать из-за их крошечных размеров. Ученые собирают их для изучения в ловушки и сметают сетями с самолетов , воздушных змеев или воздушных шаров. [135]
Экологическая роль переносимых по воздуху цианобактерий и микроводорослей понятна лишь частично. Присутствуя в воздухе, цианобактерии и микроводоросли могут способствовать зарождению льда и образованию облачных капель . Цианобактерии и микроводоросли также могут влиять на здоровье человека. [136] [137] [138] [139] [140] [141] В зависимости от размера передающиеся по воздуху цианобактерии и микроводоросли могут вдыхаться человеком и расселяться в различных отделах дыхательной системы, приводя к формированию или обострению многочисленных заболеваний и недугов, например, аллергии, дерматита, ринита. [138] [142] [143] [144]
См. также
[ редактировать ]- Экология морских личинок
- Топография поверхности океана
- Поверхностный слой
- Морские брызги
- Морской воздух
- Исследование нижней атмосферы поверхности океана
- Совместное исследование глобальных океанских потоков
- Региональная система моделирования океана
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и Хелм, Ребекка Р. (28 апреля 2021 г.). «Таинственная экосистема на поверхности океана» . ПЛОС Биология . 19 (4). Публичная научная библиотека (PLoS): e3001046. дои : 10.1371/journal.pbio.3001046 . ISSN 1545-7885 . ПМК 8081451 . ПМИД 33909611 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 . - ^ Флейшманн, Эстер М. (1989). «Измерение и проникновение ультрафиолетового излучения в тропическую морскую воду» . Лимнология и океанография . 34 (8): 1623–1629. Бибкод : 1989LimOc..34.1623F . дои : 10.4319/lo.1989.34.8.1623 . S2CID 86478743 .
- ^ Харди, Джей Ти (1982). «Микрослой морской поверхности: биология, химия и антропогенное обогащение». Прогресс в океанографии . 11 (4): 307–328. Бибкод : 1982Proce..11..307H . дои : 10.1016/0079-6611(82)90001-5 .
- ^ Jump up to: а б Вурл, Оливер; Холмс, Майкл (2008). «Студенистая природа микрослоя морской поверхности». Морская химия . 110 (1–2): 89–97. Бибкод : 2008Март.110...89Вт . дои : 10.1016/j.marchem.2008.02.009 .
- ^ Канлифф, Майкл; Мюррелл, Дж. Колин (2009). «Микрослой морской поверхности представляет собой студенистую биопленку» . Журнал ISME . 3 (9): 1001–1003. дои : 10.1038/ismej.2009.69 . ПМИД 19554040 . S2CID 32923256 .
- ^ Вурл, Оливер; Экау, Вернер; Лендинг, Уильям М.; Заппа, Кристофер Дж. (2017). «Микрослой морской поверхности в меняющемся океане – перспектива» . Элемента: Наука об антропоцене . 5 . дои : 10.1525/elementa.228 .
- ^ Jump up to: а б с Тиль, М.; Гутов, Л. (2005). «I. Плавающие субстраты» . В Гибсоне, Робин (ред.). Океанография и морская биология: ежегодный обзор . Бока-Ратон, Флорида: CRC Press. ISBN 978-0-203-50781-0 .
- ^ Jump up to: а б Тиль, М.; Гутов, Л. (2005). «II. Сплавные организмы и сообщества» . В Гибсоне, Робин (ред.). Океанография и морская биология: ежегодный обзор . Бока-Ратон, Флорида: CRC Press. ISBN 978-0-203-50781-0 .
- ^ Jump up to: а б Тиль, М.; Гутов, Л. (2005). «III. Биогеографические и эволюционные последствия» . В Гибсоне, Робин (ред.). Океанография и морская биология: ежегодный обзор . Бока-Ратон, Флорида: CRC Press. ISBN 978-0-203-50781-0 .
- ^ Jump up to: а б с Ральф, Янина (2019). «Вирионеустон: обзор вирусно-бактериальных ассоциаций на границе раздела воздух-вода» . Вирусы . 11 (2): 191. дои : 10.3390/v11020191 . ПМК 6410083 . ПМИД 30813345 .
- ^ «Экология питания гавайских морских птиц» , Монографии дикой природы , 85 : 3-71. Уайли.
- ^ Гоув, Джеймисон М.; Уитни, Джонатан Л.; Макманус, Маргарет А.; Леки, Джоуи; Карвальо, Фелипе К.; Линч, Дженнифер М.; Ли, Дживэй; Нойбауэр, Филипп; Смит, Кэтрин А.; Фиппс, Яна Э.; Кобаяши, Дональд Р.; Балагсо, Карла Б.; Контрерас, Эмили А.; Мануэль, Марк Э.; Меррифилд, Марк А.; Половина, Джеффри Дж.; Аснер, Грегори П.; Мейнард, Джеффри А.; Уильямс, Гарет Дж. (2019). «Пластмасса размером с добычу вторгается в питомники личинок рыб» . Труды Национальной академии наук . 116 (48): 24143–24149. Бибкод : 2019PNAS..11624143G . дои : 10.1073/pnas.1907496116 . ПМЦ 6883795 . ПМИД 31712423 .
- ^ Савилов, А.И. (1969) "Плейстон Тихого океана". В Зенкевиче, Луизиана (ред.) Биология Тихого океана: Часть 2. Глубоководная донная фауна .
- ^ Ли, Хеджон; Гали, Валье; Фэн, Сяоцзюань; Понтон, Камило; Гали, Альберт; Франс-Ланор, Кристиан; Фикинс, Сара Дж. (2019). «Устойчивое деревянное захоронение в Бенгальском конусе за последние 19 млн лет» . Труды Национальной академии наук . 116 (45): 22518–22525. Бибкод : 2019PNAS..11622518L . дои : 10.1073/pnas.1913714116 . ПМК 6842586 . ПМИД 31636189 .
- ^ Феррер, Луис; Пастор Ане (2017). «Португальский военный корабль: Унесенные ветром». Региональные исследования в области морских наук . 14 : 53–62. Бибкод : 2017RSMS...14...53F . дои : 10.1016/j.rsma.2017.05.004 .
- ^ Браун, Дж. (2019) Velella velella : Моряк на ветру в коллекциях музеев Виктории, Австралия. Доступ 4 декабря 2021 г. Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Иосилевский Г.; Вейс, Д. (2009). «Гидродинамика плавания португальского военного корабля Physalia physalis» . Журнал интерфейса Королевского общества . 6 (36): 613–626. дои : 10.1098/rsif.2008.0457 . ПМК 2696138 . ПМИД 19091687 .
- ^ Биери, Роберт (1966). «Предпочтения и темпы питания улиток, Ianthina Prolongata, ракушек, Lepas Anserifera, голожаберников, Glaucus Atlanticus и Fiona Pinnata, а также пищевой сети в морском Нейстоне» . Публикации Сетоской морской биологической лаборатории . 14 (2): 161–170. дои : 10.5134/175429 .
- ^ Черчилль, Селия К.К.; Вальдес, Анхель; — Фойхил, Диармайд (2014). «Молекулярная и морфологическая систематика нейстонных голожаберных (Mollusca: Gastropoda: Glaucidae: Glaucus) с описаниями трех новых загадочных видов». Систематика беспозвоночных . 28 (2): 174. дои : 10.1071/IS13038 . S2CID 84010907 .
- ^ Черчилль, Селия К.К.; Вальдес, Анхель; — Фойхил, Диармайд (2014). «Афро-Евразия и Америка представляют собой барьеры для потока генов для космополитического нейстонного голожаберника Glaucus atlanticus». Морская биология . 161 (4): 899–910. дои : 10.1007/s00227-014-2389-7 . S2CID 84153330 .
- ^ Жеден, Ванесса; Ковалев Александр; Горб, Станислав Н.; Клепал, Вальтрауд (2015). «Характеристика плавучести цементного поплавка у ракушки на стебле Dosima fasccularis (Crustacea, Cirripedia)» . Фокус на интерфейсе . 5 (1). дои : 10.1098/rsfs.2014.0060 . ПМЦ 4275874 . ПМИД 25657839 .
- ^ Костон-Клементс, Л., Сеттл, Л.Р., Хосс, Д.Э. и Кросс, Ф.А. (1991) Использование среды обитания саргассумов морскими беспозвоночными и позвоночными, обзор . Технический меморандум NOAA, том 296, NMFS-SEFSC-296.
- ^ Стюарт, Ханна Луиза (2006). «Онтогенетические изменения плавучести, прочности на разрыв, растяжимости и репродуктивных инвестиций у дрейфующей макроводоросли Turbinaria Ornata (Phaeophyta)1». Журнал психологии . 42 : 43–50. дои : 10.1111/j.1529-8817.2006.00184.x . S2CID 84580325 .
- ^ Jump up to: а б Маршалл, Гарольд Г.; Бурхардт, Любомира (2005). «Нейстон: его определение с историческим обзором его концепции и структуры сообщества». Архив гидробиологии . 164 (4): 429–448. дои : 10.1127/0003-9136/2005/0164-0429 .
- ^ Jump up to: а б Франклин, Марк П.; Макдональд, Ян Р.; Борн, Дэвид Г.; Оуэнс, Николас Дж. П.; Апстиль-Годдард, Роберт К.; Мюррелл, Дж. Колин (2005). «Бактериальное разнообразие в бактерионейстоне (микрослое поверхности моря): Бактерионейстон сквозь зеркало». Экологическая микробиология . 7 (5): 723–736. дои : 10.1111/j.1462-2920.2004.00736.x . ПМИД 15819854 .
- ^ Jump up to: а б Зибурт, Джон МакН. ; Уиллис, Паула-Жан; Джонсон, Кеннет М.; Берни, Кертис М.; Лавуа, Деннис М.; Хинга, Кеннет Р.; Кэрон, Дэвид А.; Френч, Фредерик В.; Джонсон, Пол В.; Дэвис, Пол Г. (1976). «Растворенное органическое вещество и гетеротрофный микронейстон в поверхностных микрослоях Северной Атлантики». Наука . 194 (4272): 1415–1418. Бибкод : 1976Sci...194.1415M . дои : 10.1126/science.194.4272.1415 . ПМИД 17819279 . S2CID 24058391 .
- ^ Тейлор, Джо Д.; Канлифф, Майкл (2014). «Высокопроизводительное секвенирование выявляет разнообразие нейстонных и планктонных микробов-эукариот в прибрежных водах». Журнал психологии . 50 (5): 960–965. дои : 10.1111/jpy.12228 . ПМИД 26988649 . S2CID 1205582 .
- ^ Апстиль-Годдард, Роберт К.; Фрост, Томас; Генри, Гордон Р.; Франклин, Марк; Мюррелл, Дж. Колин; Оуэнс, Николас Дж. П. (2003). «Бактерионейстонный контроль воздушно-водяного обмена метана, определяемый с помощью лабораторного газообменного резервуара» . Глобальные биогеохимические циклы . 17 (4): 1108. Бибкод : 2003GBioC..17.1108U . дои : 10.1029/2003GB002043 . S2CID 97712481 .
- ^ Цыбан, А.В. (1971). «Морской бактерионейстон». Журнал Океанографического общества Японии . 27 (2): 56–66. дои : 10.1007/BF02109331 . S2CID 198202161 .
- ^ Капоне, Дуглас Г.; Зер, Джонатан П.; Паерл, Ханс В.; Бергман, Биргитта; Карпентер, Эдвард Дж. (1997). «Триходезмий, морская цианобактерия глобального значения». Наука . 276 (5316): 1221–1229. дои : 10.1126/science.276.5316.1221 .
- ^ Икава, Теруми; Нозоэ, Юичи; Ямасита, Нацуко; Нисимура, Намико; и др. (2018). «Исследование распространения двух находящихся под угрозой исчезновения морских конькобежцев Halobates matsumurai Esaki и Asclepios shiranui (Esaki) (Hemiptera: Gerridae: Halobatinae) с особым упором на их стратегии борьбы с приливными течениями» . Психика: журнал энтомологии . 2018 . Хиндави Лимитед: 1–7. дои : 10.1155/2018/3464829 . ISSN 0033-2615 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Почему в море так мало насекомых? Немецкая волна , 9 июля 2018.
- ^ Jump up to: а б с д Андерсен, Нью-Мексико; Ченг, Л. (2005). «Морские насекомые галобатные (Heteroptera: Gerridae)» . В Гибсоне, Робин (ред.). Океанография и морская биология: ежегодный обзор . Бока-Ратон, Флорида: CRC Press. ISBN 978-0-203-50781-0 . OCLC 664909565 .
- ^ Ченг, Л. (1985). «Биология Halobates (Heteroptera: Gerridae)» . Ежегодный обзор энтомологии . 30 (1). Годовые обзоры: 111–135. doi : 10.1146/annurev.en.30.010185.000551 . ISSN 0066-4170 . S2CID 86774669 .
- ^ Jump up to: а б Херринг, П.Дж. (1965). «Синий пигмент океанической копеподы, живущей на поверхности». Природа . 205 (4966): 103–104. Бибкод : 1965Natur.205..103H . дои : 10.1038/205103a0 . S2CID 85081097 .
- ^ Jump up to: а б с Зайцев Ю. (1997). «Нейстон морей и океанов». В Лиссе П.С. (ред.). Поверхность моря и глобальные изменения . Кембридж, Нью-Йорк: Издательство Кембриджского университета. стр. 371–382. ISBN 978-0-521-56273-7 . OCLC 34933503 .
- ^ Янора, А.; Сантелла, Л. (1991). «Диапаузные эмбрионы нейстонного копепода Anomalocera patersoni». Морская биология . 108 (3): 387–394. дои : 10.1007/BF01313647 . S2CID 85058107 .
- ^ Чон, Хён Кён; Су, Хэ-Лип; Юн, Ян Хо; Чой, Им Хо; Со, Хо Ён (2008). «Первые находки двух нейстонических веслоногих каланоидов, Pontella securifer и P. Sinica (Calanoida, pontellidae) в Южном море, Корея». Журнал науки об океане . 43 (2): 91–100. Бибкод : 2008OSJ....43...91J . дои : 10.1007/BF03020585 . S2CID 84647702 .
- ^ Jump up to: а б с д и ж Ральф, Янина; Рибас-Рибас, Мариана; Браун, Скотт М.; Мустафа, Нур Или Хамиза; и др. (31 июля 2018 г.). «Синяя пигментация нейстонных копепод способствует эксплуатации богатой добычей ниши на границе воздуха и моря» . Научные отчеты . 8 (1). Springer Science and Business Media LLC: 11510. Бибкод : 2018NatSR...811510R . дои : 10.1038/s41598-018-29869-7 . ISSN 2045-2322 . ПМК 6068160 . ПМИД 30065353 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б Светличный Леонид; Ларсен, Пол С.; Кьёрбо, Томас (2017). «Плавай и летай. Стратегия побега нейстонных и планктонных копепод» . Журнал экспериментальной биологии . 221 (Часть 2). дои : 10.1242/jeb.167262 . ПМИД 29191859 . S2CID 26677839 .
- ^ Jump up to: а б Че, Дж.; Нисида, С. (1994). «Покровная ультраструктура и окраска радужных копепод семейства Sapphirinidae (Copepoda: Poecilostomatoida)». Морская биология . 119 (2): 205–210. дои : 10.1007/BF00349558 . S2CID 85268406 .
- ^ Янора, А.; Миральто, А.; Вануччи, С. (1992). «Структура поверхностного прикрепления: уникальный тип покровного образования у нейстонических копепод». Морская биология . 113 (3): 401–407. дои : 10.1007/BF00349165 . S2CID 84911783 .
- ^ Ангел, Аня; Банге, Герман В.; Канлифф, Майкл; Берроуз, Сюзанна М.; Фридрихс, Гернот; Гальгани, Луиза; Херрманн, Хартмут; Херткорн, Норберт; Джонсон, Мартин; Лисс, Питер С.; Куинн, Патрисия К.; Шартау, Маркус; Соловьев Александр; Столле, Кристиан; Апстиль-Годдард, Роберт К.; Ван Пинкстерен, Мануэла; Ценкер, Бирте (2017). «Жизненная кожа океана: к комплексному пониманию микрослоя морской поверхности» . Границы морской науки . 4 . дои : 10.3389/fmars.2017.00165 . hdl : 10026.1/16046 .
- ^ Зибурт, Джон МакН.; Уиллис, Паула-Жан; Джонсон, Кеннет М.; Берни, Кертис М.; Лавуа, Деннис М.; Хинга, Кеннет Р.; Кэрон, Дэвид А.; Френч, Фредерик В.; Джонсон, Пол В.; Дэвис, Пол Г. (1976). «Растворенное органическое вещество и гетеротрофный микронейстон в поверхностных микрослоях Северной Атлантики». Наука . 194 (4272): 1415–1418. Бибкод : 1976Sci...194.1415M . дои : 10.1126/science.194.4272.1415 . ПМИД 17819279 . S2CID 24058391 .
- ^ Бродер, Ричард Д. (1989). «Нейстонное питание молоди лососевых рыб в прибрежных водах северо-восточной части Тихого океана». Канадский журнал зоологии . 67 (8): 1995–2007. дои : 10.1139/z89-284 .
- ^ Генрих, АК (1971). «О приповерхностном планктоне восточной части южной части Тихого океана». Морская биология . 10 (4): 290–294. дои : 10.1007/BF00368087 . S2CID 85738413 .
- ^ Генрих, АК (2010). «Влияние муссонного климата на распространение веслоногих ракообразных нейстона в северо-восточной части Индийского океана». Океанология . 50 (4): 549–555. Бибкод : 2010Ocgy...50..549H . дои : 10.1134/S0001437010040119 . S2CID 128770397 .
- ^ Тернер, Дж.Т., Коллард, С.Б., Райт, Дж.К., Митчелл, Д.В. и Стил, П. (1979) «Летнее распределение копепод понтеллид в нейстоне восточной части континентального шельфа Мексиканского залива». Бюллетень морской науки , 29 (3): 287–297.
- ^ Маки, Джеймс С. (2003). «Нейстонская микробиология: жизнь на границе воздух-вода». Энциклопедия экологической микробиологии . дои : 10.1002/0471263397.env234 . ISBN 0471263397 .
- ^ Хьюмс, А.Г. (1994) «Сколько веслоногих ракообразных?» В кн.: Экология и морфология веслоногих ракообразных . Развитие гидробиологии (ред. Феррари Ф.Д. и Брэдли Б.П.), Vol. 102 Спрингер, Дордрехт, 1–7.
- ^ Ральф, Янина; Петерс, Жанна; Мояно, Марта; Пожалуйста, Оле; Клауссен, Карстен; Пек, Майрон А. (2017). «Краткосрочные молекулярные и физиологические реакции на тепловой стресс у неритических копепод Acartia tonsa и Eurytemora affinis». Сравнительная биохимия и физиология. Часть A: Молекулярная и интегративная физиология . 203 : 348–358. дои : 10.1016/j.cbpa.2016.11.001 . ПМИД 27825870 .
- ^ Моклин, Дж (1998). Биология каланоидов веслоногих . Сан-Диего: Академическая пресса. ISBN 978-0-08-057956-6 . OCLC 276935882 .
- ^ Jump up to: а б Херринг, П.Дж. (1967) «Пигменты планктона на поверхности моря». В: Симп. Зоол. Соц. Лонд , 19 : 215–235).
- ^ Загальский, П.Ф.; Херринг, Питер Дж. (1972). «Исследования каротинопротеина, выделенного из копепод, Labidocera acutifrons, и его связи с каротинопротеинами десятиногих ракообразных и другими полиенсвязывающими белками». Сравнительная биохимия и физиология. Часть B: Сравнительная биохимия . 41 (2): 397–415. дои : 10.1016/0305-0491(72)90043-0 .
- ^ Моджиб, Назия; Амад, Маан; Тимма, Манджула; Альданондо, Нароа; Кумаран, Манде; Иригойен, Ксавье (2014). «Метаболическое профилирование каротиноидов и анализ генома транскриптома выявили функциональную эквивалентность между синепигментированными копеподами и аппендикуляриями» . Молекулярная экология . 23 (11): 2740–2756. дои : 10.1111/mec.12781 . hdl : 10754/550807 . ПМИД 24803335 . S2CID 20245858 .
- ^ Херринг, П.Дж. (1965). «Синий пигмент океанической копеподы, живущей на поверхности». Природа . 205 (4966): 103–104. Бибкод : 1965Natur.205..103H . дои : 10.1038/205103a0 . S2CID 85081097 .
- ^ Карамухо, Мария-Хосе; Де Карвальо, Карла CCR; Сильва, Сорая Дж.; Карман, Кевин Р. (2012). «Диетические каротиноиды регулируют содержание астаксантина в веслоногих ракообразных и модулируют их восприимчивость к ультрафиолетовому излучению и токсичности меди» . Морские наркотики . 10 (12): 998–1018. дои : 10.3390/md10050998 . ПМЦ 3397456 . ПМИД 22822352 .
- ^ Шагин Дмитрий А.; Барсова Екатерина Владимировна; Янушевич Юрий Григорьевич; Фрадков Аркадий Ф.; Лукьянов Константин А.; Лабас Юлий А.; Семенова Татьяна Н.; Угальде, Хуан А.; Мейерс, Энн; Нуньес, Хосе М.; Виддер, Эдит А.; Лукьянов Сергей А.; Мац, Михаил В. (2004). «GFP-подобные белки как повсеместное надсемейство многоклеточных животных: эволюция функциональных особенностей и структурной сложности». Молекулярная биология и эволюция . 21 (5): 841–850. дои : 10.1093/molbev/msh079 . ПМИД 14963095 .
- ^ Абелло, Пере; Герао, Гильермо; Кодина, Мериткселл (2004). «Распространение нейстонических изопод Idotea Metallica по отношению к фронтальным структурам шельф-склона» . Журнал биологии ракообразных . 24 (4): 558–566. дои : 10.1651/C-2505 . S2CID 85806315 .
- ^ Гутов, Ларс; Франке, Хайнц-Дитер (2001). «О нынешнем и возможном будущем статусе нейстонической изоподы Idotea Metallica Bosc в Северном море: лабораторное исследование» (PDF) . Журнал морских исследований . 45 (1): 37–44. Бибкод : 2001JSR....45...37G . дои : 10.1016/S1385-1101(00)00058-7 .
- ^ Хакер, С.Д. и Мадин, Л.П. (1991) «Почему архитектура и цвет среды обитания важны для креветок, живущих в пелагической Саргассуме: использование камуфляжа и мимикрии частей растений». Серия достижений морской экологии , Ольдендорф, 70 (2): 143-155.
- ^ Хакер, СД; Мадин, LP (1991). «Почему архитектура и цвет среды обитания важны для креветок, живущих в пелагической Саргассуме: использование камуфляжа и мимикрии частей растений» . Серия «Прогресс в области морской экологии» . 70 : 143–155. Бибкод : 1991MEPS...70..143H . дои : 10.3354/meps070143 .
- ^ Миллер, Майкл (2009). «Экология ангиллиформных лептоцефалов: замечательные прозрачные личинки рыб поверхностного слоя океана». Монографии по аква-бионаукам . 2 (4). дои : 10.5047/absm.2009.00204.0001 .
- ^ Бродер, Р.Д., Пул, С.С. и Миллер, Т.В. (2013) «Селективность добычи молоди лосося по нейстоническому мезозоопланктону в северном Калифорнийском течении». Комиссия по анадромным рыбам северной части Тихого океана , Технический отчет, 9 : 104-108.
- ^ Финн, Джулиан К.; Норман, Марк Д. (2010). «Оболочка аргонавта: контроль плавучести с помощью газа у пелагического осьминога» . Труды Королевского общества B: Биологические науки . 277 (1696): 2967–2971. дои : 10.1098/rspb.2010.0155 . ПМК 2982015 . ПМИД 20484241 .
- ^ Далл WH (1869) «Заметки об аргонавте». Американский натуралист , 3 (5): 236–239.
- ^ Масия, С. (2004). «Новые наблюдения за воздушно-реактивным движением (полетом) кальмаров с обзором предыдущих отчетов». Журнал исследований моллюсков . 70 (3): 297–299. дои : 10.1093/моллюс/70.3.297 .
- ^ Jump up to: а б с д и ж г Вурл, Оливер; Экау, Вернер; Лендинг, Уильям М.; Заппа, Кристофер Дж. (1 января 2017 г.). Деминг, Джоди В.; Боуман, Джефф (ред.). «Микрослой морской поверхности в меняющемся океане – перспектива» . Элемента: Наука об антропоцене . 5 . Издательство Калифорнийского университета. дои : 10.1525/elementa.228 . ISSN 2325-1026 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б с Карлсон, Дэвид Дж. (1993). «Ранний диагенез органического вещества: реакция на границе раздела воздух-море». Органическая геохимия . Темы геобиологии. Том. 11. С. 255–268. дои : 10.1007/978-1-4615-2890-6_12 . ISBN 978-1-4613-6252-4 .
- ^ Jump up to: а б Зибурт, Джон МакН. (1983). «Микробиологические и органо-химические процессы в поверхностных и смешанных слоях». Воздушно-морской обмен газов и частиц . стр. 121–172. дои : 10.1007/978-94-009-7169-1_3 . ISBN 978-94-009-7171-4 .
- ^ Зафириу, Оливер К. (1986). «Фотохимия и микрослой морской поверхности: естественные процессы и потенциал как метода». Динамические процессы в химии верхнего слоя океана . стр. 129–135. дои : 10.1007/978-1-4684-5215-0_11 . ISBN 978-1-4684-5217-4 .
- ^ Jump up to: а б с Вурл, О.; Вурл, Э.; Миллер, Л.; Джонсон, К.; Вагл, С. (2011). «Формирование и глобальное распространение микрослоев морской поверхности» . Биогеонауки . 8 (1): 121–135. Бибкод : 2011BGeo....8..121W . дои : 10.5194/bg-8-121-2011 .
- ^ Канлифф, Майкл; Энгель, Аня; Фрка, Саня; Гашпарович, Блаженка; Гитарт, Карлос; Мюррелл, Дж. Колин; Солтер, Мэтью; Столле, Кристиан; Апстиль-Годдард, Роберт; Вурл, Оливер (2013). «Микрослои морской поверхности: единая физико-химическая и биологическая перспектива границы раздела воздух-океан». Прогресс в океанографии . 109 : 104–116. Бибкод : 2013Proce.109..104C . дои : 10.1016/j.pocean.2012.08.004 .
- ^ Эллисон, Дж. Барни; Так, Адриан Ф.; Вайда, Вероника (1999). «Атмосферная обработка органических аэрозолей» . Журнал геофизических исследований: Атмосфера . 104 (Д9): 11633–11641. Бибкод : 1999JGR...10411633E . дои : 10.1029/1999JD900073 .
- ^ Лисс, PS (1997). «Фотохимия микрослоя морской поверхности». Поверхность моря и глобальные изменения . Кембридж, Нью-Йорк: Издательство Кембриджского университета. стр. 383–424. ISBN 978-0-521-56273-7 . OCLC 34933503 .
- ^ Хантер, К.А. (1977) Химия микрослоя морской поверхности Университет Восточной Англии. Школа экологических наук.
- ^ Чжан, Чжэнбинь (2003). «Прямое определение толщины микрослоя морской поверхности с использованием pH-микроэлектрода в исходном месте». Наука в Китае. Серия Б. 46 (4): 339. дои : 10.1360/02yb0192 .
- ^ Лисс, PS (1997). «Химия микрослоя морской поверхности». Поверхность моря и глобальные изменения . Кембридж, Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-511-52502-5 . OCLC 34933503 .
- ^ Левич В.Г. (1962) Физико-химическая гидродинамика , Prentice Hall International.
- ^ Шлюссель, Питер; Эмери, Уильям Дж.; Грассль, Хартмут; Маммен, Теодор (1990). «О объемной разности температур кожи и ее влиянии на спутниковое дистанционное зондирование температуры поверхности моря». Журнал геофизических исследований . 95 (C8): 13341. Бибкод : 1990JGR....9513341S . дои : 10.1029/JC095iC08p13341 . hdl : 21.11116/0000-0004-BC37-B .
- ^ Карлсон, Дэвид Дж. (1982). «Полевая оценка методов отбора проб микрослоев пластин и сит». Морская химия . 11 (3): 189–208. Бибкод : 1982Март..11..189C . дои : 10.1016/0304-4203(82)90015-9 .
- ^ Канлифф М., Вурл О. (2014) Руководство по передовым методам изучения поверхности океана , Плимутские периодические публикации Ассоциации морской биологии Соединенного Королевства.
- ^ Патрик, Паула; Вайдберг, Николас; Гошен, Уэйн С.; Джексон, Дженнифер М.; Маккуэйд, Кристофер Д.; Порри, Франческа (31 мая 2021 г.). «Структура сообщества личинок рыб на прибрежных фронтах и влияние изменчивости окружающей среды» . Границы экологии и эволюции . 9 . Фронтирс Медиа С.А. дои : 10.3389/fevo.2021.684502 . hdl : 10037/23945 . ISSN 2296-701X . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Апель, Джон Р.; Бирн, Х. Майкл; Прони, Джон Р.; Чарнелл, Роберт Л. (1975). «Наблюдения океанических внутренних и поверхностных волн со спутника, посвященного ресурсам Земли». Журнал геофизических исследований . 80 (6): 865–881. Бибкод : 1975JGR....80..865A . дои : 10.1029/JC080i006p00865 .
- ^ Кингсфорд, MJ (1990). «Линейные океанографические особенности: в центре внимания исследования процессов набора персонала». Австралийская экология . 15 (4): 391–401. дои : 10.1111/j.1442-9993.1990.tb01465.x .
- ^ Климак, Джоди; Легг, Соня ; Алфорд, Мэтью; Буйсман, Мартен; Пинкель, Роберт; Нэш, Джонатан (2012). «Прямое разрушение внутренних волн при крутом рельефе» . Океанография . 25 (2): 150–159. дои : 10.5670/oceanog.2012.50 .
- ^ Энгель, Аня; Банге, Герман В.; Канлифф, Майкл; Берроуз, Сюзанна М.; и др. (2017). «Жизненная кожа океана: к комплексному пониманию микрослоя морской поверхности» . Границы морской науки . 4 . дои : 10.3389/fmars.2017.00165 . hdl : 10026.1/16046 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Jump up to: а б Шанкс, Алабама (1983). «Поверхностные пятна, связанные с приливно-отливными внутренними волнами, могут переносить пелагические личинки донных беспозвоночных и рыб к берегу» . Серия «Прогресс в области морской экологии» . 13 : 311–315. Бибкод : 1983MEPS...13..311S . дои : 10.3354/meps013311 .
- ^ Джиллетт, Дж. Б. и Зелдис, Дж. Р. (1985) «Аэрофотонаблюдения за неоднородностью поверхности планктонных ракообразных». Бык. март. наук. , 37 : 609–619.
- ^ Кингсфорд, MJ; Чоат, Дж. Х. (1986). «Влияние поверхностных пятен на распространение и перемещение мелкой рыбы по берегу». Морская биология . 91 (2): 161–171. дои : 10.1007/BF00569432 . S2CID 83769659 .
- ^ л. Шанкс, Алан; г. Райт, Уильям (1987). «Перенос карповых, мегалоп и гаммарид к берегу, опосредованный внутренними волнами, и коррелирующие прибрежные различия в скорости оседания приливных ракушек». Журнал экспериментальной морской биологии и экологии . 114 : 1–13. дои : 10.1016/0022-0981(87)90135-3 .
- ^ Шанкс, А.Л. (1988) «Дальнейшая поддержка гипотезы о том, что внутренние волны могут вызывать перенос личинок беспозвоночных и рыб к берегу» . Рыба. Бык. , 86 : 703–714.
- ^ Кингсфорд, MJ; Волански, Э.; Чоат, Дж. Х. (1991). «Влияние приливно-отливных фронтов и ленгмюровских циркуляций на распределение и перемещение расселяющихся рыб вокруг кораллового рифа». Морская биология . 109 : 167–180. дои : 10.1007/BF01320244 . S2CID 86057295 .
- ^ Вайдберг, Н.; Лобон, К.; Лопес, Э.; Гарсиа Флорес, Л.; Фернандес Руэда, MdP; Ларжье, Дж.; Акунья, JL (2014). «Влияние прибрежных поверхностных пятен на распределение меропланктона: роль поведения личинок» . Серия «Прогресс в области морской экологии» . 506 : 15–30. Бибкод : 2014MEPS..506...15W . дои : 10.3354/meps10777 . hdl : 10651/28404 .
- ^ Jump up to: а б с д и ж г Уитни, Джонатан Л.; Гоув, Джеймисон М.; Макманус, Маргарет А.; и др. (04.02.2021). «Поверхностные пятна являются пелагическими рассадниками разнообразной океанской фауны» . Научные отчеты . 11 (1). Springer Science and Business Media LLC: 3197. Бибкод : 2021NatSR..11.3197W . дои : 10.1038/s41598-021-81407-0 . ISSN 2045-2322 . ПМЦ 7862242 . ПМИД 33542255 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Лейс Дж. М., Маккормик М. И. (2002). «Биология, поведение и экология пелагической, личиночной стадии рыб коралловых рифов». В продаже П (ред.). Рыбы коралловых рифов: динамика и разнообразие в сложной экосистеме . Амстердам: Академическая пресса. стр. 171–199. ISBN 978-0-12-373609-3 . OCLC 53963482 .
- ^ Коуэн РК (2002). «Океанографические влияния на распространение и удержание личинок и их последствия для связи популяций». В продаже П (ред.). Рыбы коралловых рифов: динамика и разнообразие в сложной экосистеме . Амстердам: Академическая пресса. стр. 149–170. ISBN 978-0-12-373609-3 . OCLC 53963482 .
- ^ Доэрти, Питер; Фаулер, Тони (1994). «Эмпирический тест ограничения пополнения коралловых рифовых рыб». Наука . 263 (5149): 935–939. Бибкод : 1994Sci...263..935D . дои : 10.1126/science.263.5149.935 . ПМИД 17758633 . S2CID 30258297 .
- ^ Армсворт, Пол Р. (2002). «Ограничение пополнения, регулирование численности и связь личинок в метапопуляциях рифовых рыб». Экология . 83 (4): 1092. doi : 10.1890/0012-9658(2002)083[1092:RLPRAL]2.0.CO;2 . ISSN 0012-9658 .
- ^ Хобсон, Э.С. (1991) «Трофические взаимоотношения рыб, специализирующихся на питании зоопланктерами над коралловыми рифами» . В: Экология рыб коралловых рифов , Academic Press, стр. 69-95.
- ^ Боаден, А.Е.; Кингсфорд, MJ (2015). «Хищники определяют структуру сообщества рыб коралловых рифов» . Экосфера . 6 (4): 1–33. дои : 10.1890/ES14-00292.1 .
- ^ Гаффни, Р. (2004) «Оценка состояния любительского рыболовства улуа на Гавайях и рекомендации по будущему управлению». Департамент земельных и природных ресурсов Гавайских островов , Технический отчет Отдела водных ресурсов 20–02, 1–42.
- ^ Дауни, РА; Бэбкок, Колорадо; Томсон, ДП; Вандерклифт, Массачусетс (2013). «На плотность растительноядных рыб и интенсивность травоядности влияет близость к коралловым рифам» . Серия «Прогресс в области морской экологии» . 482 : 217–225. Бибкод : 2013MEPS..482..217D . дои : 10.3354/meps10250 .
- ^ Пэрриш, JD (1989). «Рыбные сообщества взаимодействующих мелководных местообитаний тропических океанических регионов» . Серия «Прогресс в области морской экологии» . 58 : 143–160. Бибкод : 1989MEPS...58..143P . дои : 10.3354/meps058143 .
- ^ Брандл, Саймон Дж.; Мораис, Ренато А.; Кейси, Джордан М.; Парравичини, Валериано; Торнабене, Люк; Гоутли, Кристофер HR; Коте, Изабель М.; Болдуин, Кэрол С.; Шьеткатте, Нина, доктор медицинских наук; Беллвуд, Дэвид Р. (2019). «Ответ на комментарий «Демографическая динамика мельчайших морских позвоночных способствует функционированию экосистемы коралловых рифов» » . Наука . 366 (6472). дои : 10.1126/science.aaz1301 . ПМИД 31857447 . S2CID 209424415 .
- ^ Уд, ЭД (1997). «Закономерности и тенденции роста и смертности костистых рыб на личиночной стадии». Журнал биологии рыб . 51 : 52–83. дои : 10.1111/j.1095-8649.1997.tb06093.x .
- ^ Хори, ЛР; Макгоуэн, Дж.А.; Вибе, PH (1978). «Закономерности и процессы во временно-пространственных масштабах распределения планктона». Пространственная структура планктонных сообществ . стр. 277–327. дои : 10.1007/978-1-4899-2195-6_12 . ISBN 978-1-4899-2197-0 .
- ^ Летчер, Б.Х.; Райс, Дж.А.; Краудер, Л.Б.; Роуз, Калифорния (1996). «Изменчивость выживаемости личинок рыб: разделение компонентов с помощью обобщенной индивидуальной модели». Канадский журнал рыболовства и водных наук . 53 (4): 787–801. дои : 10.1139/f95-241 .
- ^ Пинеда, Хесус (1994). «Внутренние приливные волны в прибрежной зоне: фронты теплой воды, гравитационные течения в сторону моря и перенос нейстонных личинок на берег». Журнал морских исследований . 52 (3): 427–458. дои : 10.1357/0022240943077046 .
- ^ Шанкс, Алан Л.; Ларжье, Джон; Бринк, Лаура; Брубейкер, Джон; Хофф, Райан (2000). «Демонстрация переноса личинок беспозвоночных на берег путем движения апвеллингового фронта к берегу» . Лимнология и океанография . 45 (1): 230–236. Бибкод : 2000LimOc..45..230S . дои : 10.4319/lo.2000.45.1.0230 . S2CID 83672860 .
- ^ Гарланд, Элизабет Д.; Циммер, Шерил Энн; Ленц, Стивен Дж. (2002). «Распределение личинок в водах внутреннего шельфа: роль ветровых поперечных шельфовых течений и суточных вертикальных миграций». Лимнология и океанография . 47 (3): 803–817. Бибкод : 2002LimOc..47..803G . дои : 10.4319/lo.2002.47.3.0803 . S2CID 86452791 .
- ^ Спонагл, Су; Ли, Томас; Курафалу, Василики; Пинкард, Дина (2005). «Фронтальные водовороты Флоридского течения и заселение коралловых рифовых рыб». Лимнология и океанография . 50 (4): 1033–1048. Бибкод : 2005LimOc..50.1033S . дои : 10.4319/lo.2005.50.4.1033 . S2CID 16048164 .
- ^ Грир, Адам Т.; Коуэн, Роберт К.; Гиганд, Седрик М.; Заяц, Джонатан А.; Тан, Дороти (2014). «Роль внутренних волн во взаимодействии личинок рыб с потенциальными хищниками и добычей». Прогресс в океанографии . 127 : 47–61. Бибкод : 2014Proce.127...47G . дои : 10.1016/j.pocean.2014.05.010 .
- ^ Шульзицки, Кэтрин; Спонагл, Су; Хауф, Марта; Уолтер, Кристен; д'Алессандро, Эван К.; Коуэн, Роберт К. (2015). «Близкие контакты с водоворотами: океанографические особенности ускоряют рост личинок рифовых рыб во время их путешествия к рифу» . Письма по биологии . 11 (1). дои : 10.1098/rsbl.2014.0746 . ПМЦ 4321146 . ПМИД 25631227 .
- ^ Шульзицки, Кэтрин; Спонагл, Су; Хауф, Марта; Уолтер, Кристен Д.; Коуэн, Роберт К. (2016). «Встреча с мезомасштабными водоворотами повышает выживаемость личинок коралловых рифовых рыб» . Труды Национальной академии наук . 113 (25): 6928–6933. Бибкод : 2016PNAS..113.6928S . дои : 10.1073/pnas.1601606113 . ПМЦ 4922168 . ПМИД 27274058 .
- ^ Вудсон, К. Брок; Литвин, Стивен Ю. (2015). «Океанические фронты стимулируют морское рыболовство и биогеохимический цикл» . Труды Национальной академии наук . 112 (6): 1710–1715. Бибкод : 2015PNAS..112.1710W . дои : 10.1073/pnas.1417143112 . ПМК 4330775 . ПМИД 25624488 .
- ^ Jump up to: а б Ларсон, Рональд Дж. (1980). «Медуза Velella velella (Линней, 1758) (Hydrozoa, Chondrophorae)». Журнал исследований планктона . 2 (3): 183–186. дои : 10.1093/планкт/2.3.183 .
- ^ Шторканова, Хана; Ореска, Сабина; Спиритович, Майя; Гержманкова, Барбора; Бубова, Кристина; Комарц, Мартин; Павелка, Карел; Венцовский, Иржи; Дистлер, Йорг Х.В.; Сенольт, Ладислав; Бечварж, Радим; Томчик, Михал (2021). «Уровни Hsp90 в плазме у пациентов с системной склеродермией и связь с поражением легких и кожи: поперечное и продольное исследование» . Научные отчеты . 11 (1): 1. Бибкод : 2021НатСР..11....1С . дои : 10.1038/s41598-020-79139-8 . ПМЦ 7791137 . ПМИД 33414495 .
- ^ Jump up to: а б Бринкманн-Фосс, А. (1970) Anthomedusae, Athecatae: (Hydrozoa, Cnidaria) Средиземноморья. 1. Случилось . Зоологическая станция.
- ^ Лаурсен, Д. (1953) Род Ianthina: монография . Океанографическая экспедиция вокруг света 1928–30 годов Фонда Карлсберга и предыдущие экспедиции «Дана». CA Рейтцелс.
- ^ Миллер Андерсен Н., Ченг Л. (2010) «Морские насекомоногие (Heteroptera: Gerridae)». В: Океанография и морская биология . CRC Press, 119–179.
- ^ Лелуп Э. (1929) «Исследование анатомии и развития Velella spirans Forsk» . Льеж.
- ^ Дельсман, ХК (1923) Вклад в Порпиты историю развития « » . Треубия, 3 : 243–266.
- ^ Уилсон, Дуглас П.; Уилсон, М. Элисон (1956). «Вклад в биологию Ianthina janthina (L.)» (PDF) . Журнал Морской биологической ассоциации Соединенного Королевства . 35 (2): 291–305. дои : 10.1017/S0025315400010146 . S2CID 83752461 .
- ^ Лалли, Кэрол (1989). Пелагические улитки: биология голопланктонных брюхоногих моллюсков . Стэнфорд, Калифорния: Издательство Стэнфордского университета. ISBN 978-0-8047-1490-7 . OCLC 18256759 .
- ^ Гауэр, JFR; Кинг, Ю.А. (2011). «Распределение плавающих саргассумов в Мексиканском заливе и Атлантическом океане, нанесенное на карту с помощью MERIS». Международный журнал дистанционного зондирования . 32 (7): 1917–1929. Бибкод : 2011IJRS...32.1917G . дои : 10.1080/01431161003639660 . S2CID 130180590 .
- ^ Вольтерек Р. (1904) «О развитии Веллеллы из глубокой личинки». Рыбак .
- ^ Савилов, А.И. (1969) "Плейстон Тихого океана". Биология Тихого океана , 264–353.
- ^ Биери, Р. (19770 «Экологическое значение сезонного появления и скорости роста Velella (Hydrozoa)». Публикации Сетоской морской биологической лаборатории , 24 (1–3): 63-76.
- ^ Савилов, А.И. (1969) "Плейстон Тихого океана". В: Биология Тихого океана , страницы 264–353.
- ^ Живые бактерии движутся в воздушных потоках Земли Журнал Smithsonian , 11 января 2016 г.
- ^ Роббинс, Джим (13 апреля 2018 г.). «Триллионы и триллионы вирусов падают с неба каждый день» . Нью-Йорк Таймс . Проверено 14 апреля 2018 г.
- ^ Рече, Изабель; Д'Орта, Гаэтано; Младенов, Наталья; Вингет, Даниэль М; Саттл, Кертис А. (29 января 2018 г.). «Скорость осаждения вирусов и бактерий над пограничным слоем атмосферы» . Журнал ИСМЕ . 12 (4): 1154–1162. дои : 10.1038/s41396-017-0042-4 . ПМК 5864199 . ПМИД 29379178 .
- ^ AC Hardy и PS Milne (1938) Исследования распространения насекомых воздушными потоками. Журнал экологии животных, 7(2):199-229.
- ^ Депре, Вивианер.; Хаффман, Дж. Алекс; Берроуз, Сюзанна М.; Хуз, Коринна; Сафатов, Александрс.; Буряк, Галина; Фрелих-Новойский, Жанин; Эльберт, Вольфганг; Андреэ, Мейнрато.; Пёшль, Ульрих; Йенике, Рупрехт (2012). «Первичные биологические аэрозольные частицы в атмосфере: обзор» . Теллус Б: Химическая и физическая метеорология . 64 : 15598. Бибкод : 2012TellB..6415598D . дои : 10.3402/tellusb.v64i0.15598 . S2CID 98741728 .
- ^ Вишневска, К.; Левандовска, Австралия; Сливиньска-Вильчевска, С. (2019). «Важность цианобактерий и микроводорослей, присутствующих в аэрозолях, для здоровья человека и окружающей среды – обзорное исследование» . Интернационал окружающей среды . 131 : 104964. doi : 10.1016/j.envint.2019.104964 . ПМИД 31351382 .
- ^ Jump up to: а б Мустака-Гуни, Мария; Кормас, К.А.; Мустака-Гуни, М. (2011). «Распространение переносимых по воздуху водорослей и цианобактерий и связанные с ними последствия для здоровья» . Границы бионауки . 3 (2): 772–787. дои : 10.2741/e285 . ПМИД 21196350 .
- ^ Бернштейн, И.Леонард; Сафферман, Роберт С. (1966). «Чувствительность кожи и слизистой бронхов к зеленым водорослям». Журнал аллергии . 38 (3): 166–173. дои : 10.1016/0021-8707(66)90039-6 . ПМИД 5223702 .
- ^ Хуз, К.; Мёлер, О. (2012). «Гетерогенное зародышеобразование льда в атмосферных аэрозолях: обзор результатов лабораторных экспериментов» . Химия и физика атмосферы . 12 (20): 9817–9854. Бибкод : 2012ACP....12.9817H . дои : 10.5194/acp-12-9817-2012 .
- ^ Тессон, Сильви В.М.; Шантл-Темкив, Тина (2018). «Активность нуклеации льда и успех эолового расселения воздушных и водных микроводорослей» . Границы микробиологии . 9 : 2681. дои : 10.3389/fmicb.2018.02681 . ПМК 6240693 . ПМИД 30483227 .
- ^ Шарма, Навин Кумар; Рай, Ашвани К. (2008). «Аллергенность воздушно-капельных цианобактерий Phormidium хрупкого и Nostoc muscorum». Экотоксикология и экологическая безопасность . 69 (1): 158–162. дои : 10.1016/j.ecoenv.2006.08.006 . ПМИД 17011621 .
- ^ Левандовска, Анита Урсула; Сливиньска-Вильчевска, Сильвия; Возничка, Доминика (2017). «Идентификация цианобактерий и микроводорослей в аэрозолях различного размера в воздухе над южной частью Балтийского моря». Бюллетень о загрязнении морской среды . 125 (1–2): 30–38. Бибкод : 2017МартПБ.125...30Л . дои : 10.1016/j.marpolbul.2017.07.064 . ПМИД 28823424 .
- ^ Доммерг, Орельен; Амато, Пьер; Тинья-Перье, Роми; Маганд, Оливье; Толлот, Альбан; Жоли, Мюриэл; Бувье, Летиция; Селлегри, Карин; Фогель, Тимоти; Сонке, Йерун Э.; Джаффрезо, Жан-Люк; Андраде, Маркос; Морено, Изабель; Лабушань, Каспер; Мартин, Линвилл; Чжан, Цянгун; Лароз, Кэтрин (2019). «Методы исследования глобального атмосферного микробиома» . Границы микробиологии . 10 : 243. дои : 10.3389/fmicb.2019.00243 . ПМК 6394204 . ПМИД 30967843 . Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .