Рыбий плавник

(1) грудные плавники (парные), (2) брюшные плавники (парные), (3) спинной плавник ,
(4) жировой плавник, (5) анальный плавник, (6) хвостовой (хвостовой) плавник.
Плавники — это подвижные придатки, выступающие из тела рыбы , которые взаимодействуют с водой, создавая тягу и помогая рыбе плавать . За исключением хвостового или хвостового плавника , плавники рыб не имеют прямой связи с позвоночником и поддерживаются только мышцами .
Рыбьи плавники представляют собой отличительные анатомические особенности с различной структурой в разных кладах : у лучепёрых рыб ( Actinopterygii ) плавники в основном состоят из костных шипов или лучей , покрытых тонким слоем бесчешуйной кожи ; у лопастноперых рыб ( Sarcopterygii ), таких как целаканты и двоякодышащие рыбы , плавники представляют собой короткие лучи, основанные вокруг мускулистой центральной почки , поддерживаемой суставчатыми костями ; у хрящевых рыб ( Chondrichthyes ) и бесчелюстных рыб ( Agnatha ) плавники представляют собой мясистые « ласты », поддерживаемые хрящевым скелетом.
Плавники в разных местах тела рыбы служат разным целям и делятся на две группы: среднесагиттальные непарные плавники и более латерально расположенные парные плавники . Непарные плавники преимущественно связаны с созданием линейного ускорения посредством колебательного движения , а также с обеспечением курсовой устойчивости ; в то время как парные плавники используются для создания ускорения , замедления при гребле, а также дифференциальной тяги или подъемной силы при повороте , всплытии на поверхность или нырянии и перекатывании . Плавники также можно использовать для других видов передвижения, помимо плавания, например, летучие рыбы используют грудные плавники для планирующего полета над поверхностью воды, а рыбы-лягушки и многие амфибии используют грудные и/или тазовые плавники для ползания . Плавники можно использовать и для других целей: реморы и бычки развили спинные плавники, похожие на присоски, для прикрепления к поверхности и «путешествия автостопом»; самцы акул и рыб-москитов используют модифицированный плавник для доставки спермы; акулы-молотилки используют хвостовой плавник, чтобы хлестать и оглушать добычу; рифовая рыба-камень в спинных плавниках есть шипы, которые впрыскивают яд в качестве защиты от хищников ; удильщики используют первый шип спинного плавника как удочку , чтобы приманить добычу; а спинороги избегают хищников , втискиваясь в расщелины кораллов и используя шипы в плавниках, чтобы закрепиться на месте.
Типы плавников
[ редактировать ]Плавники могут быть парными и непарными . Грудные и брюшные плавники парные, тогда как спинной, анальный и хвостовой плавники непарные и расположены вдоль средней линии тела. Для каждого типа плавников существует ряд видов рыб , у которых этот конкретный плавник был утерян в ходе эволюции (например, брюшные плавники у † Bobasatrania , хвостовой плавник у океанической солнечной рыбы ). У некоторых клад в ходе эволюции были приобретены дополнительные непарные плавники (например, дополнительные спинные плавники, жировой плавник). У некоторых † Acanthodii («колючих акул») между грудными и брюшными плавниками присутствуют одна или несколько пар «промежуточных» или «предбрюшных» шипов, но они не связаны с плавниками.
| Грудные плавники (плавники на руках) |

|
Парные грудные плавники расположены с каждой стороны, обычно сложенные сразу за жаберной крышкой, и гомологичны передним конечностям четвероногих четвероногих или верхним конечностям двуногих четвероногих.
|
|---|---|---|
| Брюшные/брюшные плавники (брюшные плавники) |

|
Парные брюшные или брюшные плавники — это брюшные плавники (от лат. venter «живот»), обычно расположенные на брюшной стороне ниже и позади грудных плавников, хотя во многих семействах рыб они могут располагаться перед грудными плавниками (например, у трески). Они гомологичны задним конечностям четвероногих четвероногих или нижним конечностям прямоногих четвероногих.
Брюшной плавник помогает рыбе подниматься или опускаться по воде, резко поворачиваться и быстро останавливаться.
|
| Спинной плавник (Спинные плавники) |
 Спинной плавник акулы
|
 Спинные плавники расположены на спине. У рыбы может быть до трех спинных плавников. Спинные плавники защищают рыбу от перекачивания, помогают ей совершать резкие повороты и остановки.
|
| Анальный/клоакальный конец | 
|
Анальный /клоакальный плавник расположен на вентральной поверхности позади ануса / клоаки .
|
| Жировой конец |  Жировой плавник форели
|
Жировой плавник — это мягкий мясистый плавник, расположенный на спине позади спинного плавника и сразу перед хвостовым плавником. Он отсутствует во многих семействах рыб, но встречается в девяти из 31 отряда эвтелеостовых ( Percopsiformes , Myctophiformes , Aulopiformes , Stomiiformes , Salmoniformes , Osmeriformes , Characiformes , Siluriformes и Argentiniformes ). [ 3 ] Известные представители этих отрядов — лососевые , харацидовые и сомовые .
Функция жирового плавника остается загадкой. Его часто отрезают, чтобы пометить рыбу, выращенную в заводских условиях, хотя данные 2005 года показали, что форель с удаленным жировым плавником имеет на 8% более высокую частоту ударов хвостом. [ 4 ] [ 5 ] Дополнительная информация, опубликованная в 2011 году, предполагает, что плавник может иметь жизненно важное значение для обнаружения и реагирования на такие раздражители, как прикосновение, звук и изменения давления. Канадские исследователи обнаружили в плавнике нейронную сеть, что указывает на то, что она, вероятно, выполняет сенсорную функцию, но до сих пор не уверены, каковы будут последствия ее удаления. [ 6 ] [ 7 ] Сравнительное исследование 2013 года показывает, что жировой плавник может развиваться двумя разными способами. Один из них — лососевидный, при котором жировой плавник развивается из личиночной плавниковой складки одновременно и таким же прямым образом, как и остальные срединные плавники. Другой способ - харациформный, при котором жировой плавник развивается поздно, после того как личиночная плавниковая складка уменьшилась и развились другие срединные плавники. Они утверждают, что существование харациформного типа развития предполагает, что жировой плавник не является «просто остатком личиночной плавниковой складки» и несовместим с мнением о том, что жировой плавник не имеет функции. [ 3 ] Исследования, опубликованные в 2014 году, показывают, что жировой плавник неоднократно эволюционировал в отдельных линиях . [ 8 ] |
| Хвостовой плавник (Хвостовой плавник) |
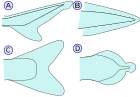  Гетероцеркальный хвостовой плавник (А)  Гомоцеркальный хвостовой плавник (С)
|
Хвостовой плавник — хвостовой плавник (от латинского cauda — хвост), расположенный на конце хвостового стебля. он используется для движения У большинства таксонов (см. Также передвижение тела и хвостового плавника ). Хвостовой плавник поддерживается позвонками осевого скелета и птеригиофорами (радиалами). В зависимости от взаимоотношений с осевым скелетом различают четыре типа хвостовых плавников (ХД):
(А) — Гетероцеркальный означает, что позвонки доходят до верхней доли хвоста, часто делая ее длиннее нижней доли (как у акул , † Placodermi , большинства стволовых Actinopterygii , а также осетров и веслоносов ). Однако внешняя форма гетероцеркальных хвостовых плавников также может казаться симметричной (например, † Birgeria , † Bobasatrania ). Гетероцеркальный – противоположность гипоцеркального.
(Б) - Протоцеркальный означает, что позвонки доходят до кончика хвоста, а хвост симметричен, но не расширен (как у первых рыб и круглоротых , а также у более примитивного предшественника у ланцетников ) (C) - Гомоцеркальный , при котором плавник обычно кажется внешне симметричным, но на самом деле позвонки заходят на очень короткое расстояние в верхнюю долю плавника. Однако гомоцеркальные хвостовые плавники также могут быть асимметричными (например, голубая летучая рыба ). Большинство современных рыб ( костистых рыб ) имеют гомоцеркальный хвост. Они бывают разных форм и могут выглядеть:
(D) - Дифицеркальный означает, что позвонки доходят до кончика хвоста, а хвост симметричен и расширен (как у бичира , двоякодышащих рыб , миног , целакантов и † Tarrasiiformes ). Большинство палеозойских рыб имели дифицеркальный гетероцеркальный хвост. [ 11 ]
|
| Хвостовой киль Финлеты |

|
Некоторые виды быстро плавающих рыб имеют горизонтальный хвостовой киль прямо перед хвостовым плавником. Подобно килю корабля, это боковой гребень на хвостовом стебле, обычно состоящий из щитков (см. Ниже), который обеспечивает устойчивость и поддержку хвостового плавника. Киль может быть одиночный парный, по одному с каждой стороны, или две пары сверху и снизу.
Плавники — это небольшие плавники, обычно расположенные позади спинного и анального плавников (у бичиров плавники есть только на спинной поверхности, а спинной плавник отсутствует). У некоторых рыб, таких как тунец или сайра , они лишены лучей, не убираются и находятся между последним спинным и/или анальным плавником и хвостовым плавником. |
Костные рыбы
[ редактировать ]
Костные рыбы ( Actinopterygii и Sarcopterygii ) образуют таксономическую группу, называемую Osteichthyes (или Euteleostomi , куда входят также наземные позвоночные ); их скелеты состоят в основном из костей, и их можно сравнить с хрящевыми рыбами (см. ниже), скелеты которых состоят в основном из хрящей (за исключением зубов , плавниковых шипов и зубчиков ).
Костные рыбы делятся на лучепёрых и лопастёрых . Большинство ныне живущих рыб — лучепёрые — чрезвычайно разнообразная и многочисленная группа, насчитывающая более 30 000 видов . Это самый крупный из существующих сегодня классов позвоночных животных, составляющий более 50% видов. [ 13 ] В далеком прошлом лопастноперые рыбы были в изобилии; однако в настоящее время существует только 8 видов.
У костистых рыб есть шипы на плавниках, называемые лепидотрихиями или «лучами» (из-за того, как шипы раскрываются). Обычно у них есть плавательные пузыри , которые позволяют рыбе изменять относительную плотность своего тела и, следовательно, плавучесть , поэтому она может тонуть или плавать без необходимости использовать плавники для плавания вверх и вниз. [ 14 ] Однако плавательный пузырь отсутствует у многих рыб, особенно у двоякодышащих рыб , у которых плавательный пузырь превратился в примитивные легкие . [ 15 ] которые могут иметь общее эволюционное происхождение с таковыми у их наземных родственников, четвероногих . [ 16 ] У костных рыб также есть пара крышек , которые перетягивают воду через жабры , что помогает им дышать без необходимости плыть вперед, чтобы направить воду в рот через жабры. [ 14 ]
лопастные плавники
[ редактировать ]


Лопастёрые рыбы образуют класс костистых рыб под названием Sarcopterygii. У них мясистые, лопастные , парные плавники, которые соединены с телом рядом костей. [ 17 ] Плавники лопастных рыб отличаются от плавников всех других рыб тем, что каждый из них имеет мясистый, лопастной, чешуйчатый стебель, отходящий от тела. Грудные и брюшные плавники имеют сочленения, напоминающие конечности четвероногих. Эти плавники превратились в ноги первых четвероногих наземных позвоночных ( амфибий ) в девонский период . Саркоптериги также обладают двумя спинными плавниками с отдельными основаниями, в отличие от одного спинного плавника у большинства лучепёрых рыб (за исключением некоторых костистых рыб ). Хвостовой плавник либо гетероцеркальный (только ископаемые таксоны ), либо дифицеркальный.
Целакант — один из видов живых лопастноперых рыб. Оба современных представителя этой группы, целакант Западной Индийского океана ( Latimeria chalumnae ) и индонезийский целакант ( Latimeria menadoensis ), встречаются в роде Latimeria . Считается, что целаканты примерно развились до своей нынешней формы около 408 миллионов лет назад, в раннем девоне. [ 18 ]
Передвижение целакантов уникально для их вида. Для передвижения латимерии чаще всего используют подъемы или спуски течения и дрейф. Они используют свои парные плавники, чтобы стабилизировать свое движение в воде. На дне океана их парные плавники не используются для каких-либо движений. Целаканты могут создавать тягу для быстрого старта, используя хвостовые плавники. Благодаря большому количеству плавников латимерии обладают высокой маневренностью и могут ориентировать свое тело практически в любом направлении в воде. Их видели стоящими на голове и плавающими животом вверх. Считается, что их ростральный орган помогает целакантам обеспечивать электровосприятие, которое помогает им передвигаться вокруг препятствий. [ 19 ]
Двоякодышащие рыбы также являются живыми лопастноперыми рыбами. Встречаются в Африке ( Protopterus ), Австралии ( Neoceratodus ) и Южной Америке ( Lepidosiren ). Двоякодышащие рыбы эволюционировали в девонский период. Генетические исследования и палеонтологические данные подтверждают, что двоякодышащие рыбы — ближайшие ныне живущие родственники наземных позвоночных . [ 20 ]
Расположение плавников и форма тела у лопастных рыб относительно консервативны. Тем не менее, есть несколько примеров из летописи окаменелостей , которые демонстрируют аберрантную морфологию , например, Allenypterus , Rebellatrix , Foreyia или тетраподоморфы .
Разнообразие плавников у лопастных рыб.
[ редактировать ]-

-
 Пятнистая двоякодышащая рыба Protopterus Dolloi
Пятнистая двоякодышащая рыба Protopterus Dolloi -
 Двоякодышащая рыба Квинсленда Neoceratodus forsteri
Двоякодышащая рыба Квинсленда Neoceratodus forsteri -

-

-

-

-

-
 Целакант Западной Индийского океана Latimeria chalumnae
Целакант Западной Индийского океана Latimeria chalumnae -










Лучевые плавники
[ редактировать ]
Лучеперые рыбы образуют класс костистых рыб под названием Actinopterygii. Их плавники содержат шипы или лучи. Плавник может содержать только колючие лучи, только мягкие лучи или комбинацию того и другого. Если присутствуют оба, колючие лучи всегда располагаются спереди . Шипы обычно жесткие и острые. Лучи обычно мягкие, гибкие, сегментированные и могут быть разветвленными. Эта сегментация лучей является главным отличием, отделяющим их от шипов; У некоторых видов шипы могут быть гибкими, но они никогда не будут сегментированы.
Шипы имеют разнообразное применение. У сома они используются как средство защиты; многие сомы обладают способностью выворачивать шипы наружу. Спинороги также используют шипы, чтобы запираться в расщелинах и не допускать их вытягивания.
Лепидотрихии обычно состоят из кости , но у ранних остихтиев , таких как Cheirolepis , также были дентин и эмаль . [ 21 ] Они сегментированы и представляют собой серию дисков, сложенных один на другой. Возможно, они произошли от кожных чешуек. [ 21 ] Считается, что генетической основой формирования плавниковых лучей являются гены, кодирующие выработку определенных белков. Было высказано предположение, что эволюция конечностей четвероногих у лопастных рыб связана с потерей этих белков. [ 22 ]
Разнообразие плавников лучепёрых рыб
[ редактировать ]-
 Рыболов Фанфин Колофрин Джордани
Рыболов Фанфин Колофрин Джордани -
 Блинная нетопырь Halieutichthys aculeatus
Блинная нетопырь Halieutichthys aculeatus -
 Стройная солнечная рыба Ranzania laevis
Стройная солнечная рыба Ranzania laevis -
 Фанфиш Pteraclis carolinus
Фанфиш Pteraclis carolinus -
 Прозрачный топорик Sternoptyxdiaphana
Прозрачный топорик Sternoptyxdiaphana -
 Серебряный грубиян Hoplostethus mediterraneus
Серебряный грубиян Hoplostethus mediterraneus -
 Хохлатая камбала Lophonectes Gallus
Хохлатая камбала Lophonectes Gallus -
 Рыба-нож Equetus lanceolatus
Рыба-нож Equetus lanceolatus -
 Атлантический помфрет Брама Брама
Атлантический помфрет Брама Брама -
 Атлантический затонувший корабль Polyprion americanus
Атлантический затонувший корабль Polyprion americanus -
 Звездчатый иглобрюх Arothron stellatus
Звездчатый иглобрюх Arothron stellatus -
 Морской дьявол, наблюдающий за звездами, Ceratias uranoscopus
Морской дьявол, наблюдающий за звездами, Ceratias uranoscopus -
 Риджхед Поромитра единороговый
Риджхед Поромитра единороговый -
 Тропическая двукрылая летучая рыба Exocoetus evolans
Тропическая двукрылая летучая рыба Exocoetus evolans -
 Огонь Benthocometesrobustus
Огонь Benthocometesrobustus -
 Крысохвост Trachonurus sulcatus
Крысохвост Trachonurus sulcatus -
 Рыба-тренога Bathypterois grallator
Рыба-тренога Bathypterois grallator -
 Гигантская рыба-весла Regalecus glesne
Гигантская рыба-весла Regalecus glesne -
 Короткоклювая копья Tetrapturus angustirostris
Короткоклювая копья Tetrapturus angustirostris -
 Рыба-нож-призрак Sternarchorhynchus oxyrhynchus
Рыба-нож-призрак Sternarchorhynchus oxyrhynchus -
 Ремора Ремора брахиптера
Ремора Ремора брахиптера -
 Скалистый прыгун с синей чертой Blenniella периофтальмус
Скалистый прыгун с синей чертой Blenniella периофтальмус -
 Нильский бишир Polypterus bichir
Нильский бишир Polypterus bichir -
 Прибрежная форель-головорез Oncorhynchus clarkii
Прибрежная форель-головорез Oncorhynchus clarkii -
 Африканский масляный сом Schilbe mystus
Африканский масляный сом Schilbe mystus -
 Морской угорь Leptocephalus conger
Морской угорь Leptocephalus conger

























Хрящевые рыбы
[ редактировать ]

Хрящевые рыбы образуют класс рыб Chondrichthyes. У них скелеты состоят из хрящей, а не из костей . В класс входят акулы , скаты и химеры .
Скелеты акульих плавников удлинены и поддерживаются мягкими несегментированными лучами, называемыми цератотрихиями, нитями эластичного белка, напоминающими роговой кератин в волосах и перьях. [ 23 ] Первоначально грудной и тазовый пояса, не содержащие кожных элементов, не соединялись. У более поздних форм каждая пара плавников стала соединяться вентрально посередине, когда развились лопаточно-коракоидная и лобково-седалищная перемычки. У скатов грудные плавники соединены с головой и очень гибки. Одной из основных характеристик, присутствующих у большинства акул, является гетероцеркальный хвост, который помогает в передвижении. [ 24 ] У большинства акул восемь плавников. Акулы могут отдаляться только от объектов, находящихся прямо перед ними, потому что их плавники не позволяют им двигаться в направлении хвоста вперед. [ 25 ]
В отличие от современных хрящевых рыб, представители стволовых хондрихтиевых линий (например, † климатииды и † диплакантиды ) [ 26 ] имели грудные кожные пластинки, а также кожные шипы, связанные с парными плавниками. Самым старым видом, демонстрирующим эти особенности, является † акантодий † Fanjingshania renovata. [ 27 ] из нижнего силура ( аэрона ) Китая. Fanjingshania обладает сложными грудными пластинами, состоящими из кожных чешуек, сросшихся с костной пластинкой, и плавниковыми шипами, полностью состоящими из кости. Шипы, связанные со спинными плавниками, редки среди современных хрящевых рыб, но присутствуют, например, у Heterodontus или Squalus . Шипы спинных плавников обычно развиты во многих группах ископаемых, например, у † Hybodontiformes , † Ctenacanthiformes или † Xenacanthida . У † Stethacanthus первый шип спинного плавника видоизменился, образовав комплекс шип-щетка.
Как и у большинства рыб, хвосты акул обеспечивают тягу, в результате чего скорость и ускорение зависят от формы хвоста. Форма хвостового плавника значительно различается у разных видов акул из-за их эволюции в разных средах обитания. Акулы обладают гетероцеркальным часть которого хвостовым плавником, спинная обычно заметно больше брюшной . акулы Это связано с тем, что позвоночный столб доходит до спинной части, обеспечивая большую площадь поверхности для мышц прикрепления . Это позволяет более эффективно передвигаться среди этих хрящевых рыб с отрицательной плавучестью . Напротив, у большинства костистых рыб имеется гомоцеркальный хвостовой плавник. [ 28 ]
Тигровые акулы имеют большую верхнюю лопасть , которая позволяет им двигаться медленно и резко увеличивать скорость. Тигровая акула должна иметь возможность легко извиваться и поворачиваться в воде во время охоты, чтобы поддерживать свой разнообразный рацион, в то время как белая акула , которая охотится на стайную рыбу, такую как скумбрия и сельдь , имеет большую нижнюю лопасть, которая помогает ей идти в ногу с ее быстрой скоростью. -плавающая добыча. [ 13 ] Другие приспособления хвоста помогают акулам ловить добычу более эффективно, например, акула-молотилка использует свою мощную удлиненную верхнюю лопасть для оглушения рыбы и кальмаров.
С другой стороны, скаты используют свои увеличенные грудные плавники для движения. Аналогично увеличенные грудные плавники можно обнаружить у вымерших † Petalodontiformes (например, † Belantsea , † Janassa , † Menaspis ), принадлежащих к Holocephali (рыба-крыса и их ископаемые родственники), или у † Aquilolamna ( Selachimorpha ) и † Squatinactis (Squatinactiformes). Некоторые хрящевые рыбы передвигаются по типу угря (например, Chlamydoselachus , † Thrinacoselache , [ 29 ] † Фебод [ 30 ] )
Разнообразие плавников хрящевых рыб.
[ редактировать ]-
 Мелкопятнистая кошачья акула Scyliorhinus canicula
Мелкопятнистая кошачья акула Scyliorhinus canicula -
 Большая белая акула Carcharodon carcharias
Большая белая акула Carcharodon carcharias -
 Обыкновенная молотилка Alopias vulpinus
Обыкновенная молотилка Alopias vulpinus -
 Крупнозубая рыба-пила Pristis perottetti
Крупнозубая рыба-пила Pristis perottetti -
 Мраморный электрический скат Torpedo marmorata
Мраморный электрический скат Torpedo marmorata -
 Скат Беннета Hemitrygon bennettii
Скат Беннета Hemitrygon bennettii -
 Плащеносная акула Chlamydoselachus anguineus
Плащеносная акула Chlamydoselachus anguineus -
 ксенакант † † senckenbergianus Lebachacanthus
ксенакант † † senckenbergianus Lebachacanthus -
 симморииформный † произвел † стетакантус
симморииформный † произвел † стетакантус -
 † † ктенакантид полосатое тело Водолея
† † ктенакантид полосатое тело Водолея -

-
 † Sibyrhynchus Iniopterygiformes † denisoni (Holocephali)
† Sibyrhynchus Iniopterygiformes † denisoni (Holocephali) -
 Кубинская химера Химера кубанская
Кубинская химера Химера кубанская -
 Американский слон Callorhinchus callorhynchus
Американский слон Callorhinchus callorhynchus














Акулий плавник
[ редактировать ]По данным Международного общества защиты животных , ежегодно около 100 миллионов акул убивают из-за их плавников. Этот акт известен как « акулий плавник» . [ 31 ] После того как плавники отрезают, изувеченных акул бросают обратно в воду и оставляют умирать.
В некоторых странах Азии акульи плавники являются кулинарным деликатесом, например, супом из акульих плавников . [ 32 ] В настоящее время обеспокоенность международного сообщества по поводу устойчивости и благополучия акул повлияла на потребление и доступность супа из акульих плавников во всем мире. [ 33 ] Добыча акульих плавников запрещена во многих странах.
Фин функции
[ редактировать ]Создание тяги
[ редактировать ]фольги в форме Плавники при движении создают тягу , подъем плавника приводит в движение воду или воздух и толкает плавник в противоположном направлении. Водные животные получают значительную тягу , двигая плавниками вперед и назад в воде. Часто хвостовой плавник используется , но некоторые водные животные создают тягу за счет грудных плавников . [ 34 ]



Кавитация возникает, когда отрицательное давление приводит к образованию в жидкости пузырьков (полостей), которые затем быстро и сильно разрушаются. Это может привести к значительным повреждениям и износу. [ 35 ] Кавитационные повреждения могут возникнуть в хвостовых плавниках мощных плавающих морских животных, таких как дельфины и тунцы. Кавитация чаще возникает вблизи поверхности океана, где давление окружающей воды относительно низкое. Даже если у них есть способность плавать быстрее, дельфинам, возможно, придется ограничить свою скорость, потому что схлопывание кавитационных пузырей на их хвосте слишком болезненно. [ 36 ] Кавитация также замедляет движение тунца, но по другой причине. В отличие от дельфинов, эти рыбы не чувствуют пузырей, поскольку имеют костистые плавники без нервных окончаний. Тем не менее, они не могут плыть быстрее, потому что кавитационные пузырьки создают вокруг их плавников паровую пленку, ограничивающую их скорость. На тунце были обнаружены повреждения, соответствующие кавитационным повреждениям. [ 36 ]
Скомбридовые рыбы (тунец, скумбрия и скумбрия) являются особенно эффективными пловцами. Вдоль края задней части их тела проходит линия небольших невыдвижных плавников без лучей, известных как плавники . Было много предположений о функции этих плавников. Исследования, проведенные в 2000 и 2001 годах Науэном и Лаудером, показали, что «плавники оказывают гидродинамическое воздействие на локальный поток во время устойчивого плавания» и что «самый задний плавник ориентирован на перенаправление потока в развивающийся хвостовой вихрь, что может увеличить тягу, создаваемую хвост плавающей скумбрии». [ 37 ] [ 38 ] [ 39 ]
У рыб несколько плавников, поэтому возможно, что один плавник может гидродинамически взаимодействовать с другим плавником. В частности, плавники, расположенные непосредственно перед хвостовым (хвостовым) плавником, могут находиться рядом с плавниками, которые могут напрямую влиять на динамику потока в хвостовом плавнике. В 2011 году исследователи, использующие методы объемной визуализации, смогли создать «первые мгновенные трехмерные изображения структур следа, создаваемых свободно плавающими рыбами». Они обнаружили, что «непрерывные удары хвоста привели к образованию связанной цепочки вихревых колец» и что «следы спинного и анального плавников быстро увлекаются следом хвостового плавника, примерно в течение времени последующего удара хвоста». [ 40 ]
Управление движением
[ редактировать ]Как только движение установлено, самим движением можно управлять с помощью других плавников. [ 34 ] [ 41 ]


Тело рифовых рыб часто имеет иную форму, чем у рыб открытой воды . Рыбы в открытой воде обычно созданы для скорости и имеют обтекаемую форму, как торпеды, чтобы минимизировать трение при движении в воде. Рифовые рыбы обитают в относительно ограниченных пространствах и сложных подводных ландшафтах коралловых рифов . Поскольку эта маневренность более важна, чем скорость по прямой, поэтому рыбы коралловых рифов развили тела, которые оптимизируют их способность бросаться и менять направление. Они перехитрили хищников, ныряя в трещины рифа или играя в прятки вокруг коралловых голов. [ 45 ] Грудные и тазовые плавники многих рифовых рыб, таких как рыба-бабочка , стрекоза и рыба-ангел , развились так, что они могут действовать как тормоза и обеспечивать сложные маневры. [ 47 ] Многие рифовые рыбы, такие как рыба-бабочка , рыба-стрекоза и рыба-ангел , развили тела, которые являются глубокими и сжатыми с боков, как блин, и могут помещаться в трещины в скалах. Их брюшные и грудные плавники развились по-разному, поэтому они действуют вместе с уплощенным телом, оптимизируя маневренность. [ 45 ] Некоторые рыбы, такие как рыба-фугу , рыба-филе и рыба-хобот , для плавания полагаются на грудные плавники и почти не используют хвостовые плавники. [ 47 ]
Воспроизведение
[ редактировать ]

Самцы хрящевых рыб (акулы и скаты), а также самцы некоторых живородящих лучепёрых рыб имеют плавники, которые были модифицированы и функционируют как интромиттивные органы , репродуктивные придатки, которые обеспечивают внутреннее оплодотворение . У лучепёрых рыб они называются гоноподиями или андроподиями , а у хрящевых рыб — класперами .
Гоноподии встречаются у самцов некоторых видов семейств Anablepidae и Poeciliidae . Это анальные плавники, которые были модифицированы и функционируют как подвижные интромиттивные органы и используются для оплодотворения самок молоком во время спаривания. Третий, четвертый и пятый лучи анального плавника самца сформированы в трубчатую структуру, в которую выбрасывается сперма рыбы. [ 50 ] Когда гонопий готов к спариванию, он становится прямым и направлен вперед в сторону самки. Самец вскоре вставляет орган в половое отверстие самки с помощью крючкообразных приспособлений, которые позволяют рыбе захватывать самку, чтобы обеспечить оплодотворение. Если самка остается неподвижной, а ее партнер касается ее отверстия своим гоноподием, она оплодотворяется. Сперма сохраняется в яйцеводе самки. Это позволяет самкам оплодотворять себя в любое время без дополнительной помощи самцов. У некоторых видов гоноподий может составлять половину общей длины тела. Иногда плавник слишком длинный, чтобы его можно было использовать, как у лирохвостых пород Xiphophorus helleri . У женщин, получающих гормональные препараты, могут развиться гоноподии. Они бесполезны для разведения.
Подобные органы со схожими характеристиками встречаются и у других рыб, например, андроподия у Hemirhamphodon или Goodeidae . [ 51 ] или гоноподий среднего триаса † Saurichthys , старейший известный пример живорождения лучепёрых рыб. [ 52 ]
Класперы встречаются у самцов хрящевых рыб . самки Они представляют собой заднюю часть брюшных плавников, которые также были модифицированы для выполнения функций интромиттирующих органов и используются для направления спермы в клоаку во время совокупления. Акт спаривания у акул обычно включает в себя поднятие одной из застежек, чтобы вода попала в сифон через определенное отверстие . Затем застежку вставляют в клоаку, где она раскрывается как зонтик, фиксируя свое положение. Затем сифон начинает сокращаться, вытесняя воду и сперму. [ 53 ] [ 54 ]
Другие функции
[ редактировать ]Другие виды использования плавников включают ходьбу и сидение на морском дне, скольжение по воде, охлаждение температуры тела, оглушение добычи, демонстрацию (отпугивание хищников, ухаживание), защиту (ядовитые шипы плавников, запирание между кораллами), заманивание добычи, и структуры прикрепления.
Парусник Индо-Тихоокеанского региона имеет выдающийся спинной плавник. Подобно скомброидам и другим марлиновым , во время плавания они оптимизируют себя, втягивая спинные плавники в бороздку на теле. [ 55 ] Огромный спинной плавник, или парус, парусника большую часть времени втянут. Парусники поднимают их, если хотят собрать стаю мелких рыб, а также после периодов высокой активности, предположительно, чтобы остыть. [ 55 ] [ 56 ]





У восточной летающей гурнарды большие грудные плавники, которые она обычно прижимает к телу, и они расширяются, когда им угрожают, чтобы отпугнуть хищников. Несмотря на свое название, это демерсальная рыба , а не летающая рыба, и она использует свои брюшные плавники, чтобы передвигаться по дну океана. [ 58 ] [ 59 ]
Плавники могут иметь адаптивное значение как сексуальное украшение. Во время ухаживания самка цихлиды демонстрирует Pelvicachromis taeniatus большой и визуально привлекающий внимание фиолетовый брюшной плавник . «Исследователи обнаружили, что самцы явно предпочитают самок с более крупным тазовым плавником и что у самок брюшные плавники растут более непропорционально, чем другие плавники». [ 60 ] [ 61 ]




Эволюция
[ редактировать ]Эволюция парных плавников
[ редактировать ]Существуют две преобладающие гипотезы, которые исторически обсуждались как модели эволюции парных плавников у рыб: теория жаберных дуг и теория боковых плавниковых складок. Первая, обычно называемая « гипотезой Гегенбаура », была выдвинута в 1870 году и предполагает, что «парные плавники произошли от жаберных структур». [ 63 ] Популярность этого подхода упала в пользу теории боковых плавниковых складок, впервые предложенной в 1877 году, которая предполагает, что парные плавники образовались из продольных боковых складок вдоль эпидермиса сразу за жабрами. [ 64 ] Обе гипотезы имеют слабую поддержку в летописи окаменелостей и в эмбриологии. [ 65 ] Однако недавние данные о закономерностях развития побудили пересмотреть обе теории, чтобы лучше объяснить происхождение парных плавников.
Классические теории
[ редактировать ]Карла Гегенбаура была введена в 1876 году. Концепция «архиптеригиума» [ 66 ] Его описали как жаберный луч или «соединенный хрящевой стебель», отходящий от жаберной дуги. Дополнительные лучи отходили вдоль дуги и от центрального жаберного луча. Гегенбаур предложил модель трансформационной гомологии , согласно которой все парные плавники и конечности позвоночных представляли собой трансформации архипттеригиума. Согласно этой теории, парные придатки, такие как грудные и брюшные плавники, должны были дифференцироваться из жаберных дуг и мигрировать назад. Однако эта гипотеза получила ограниченную поддержку в летописи окаменелостей как морфологически, так и филогенетически. [ 65 ] Кроме того, практически не было обнаружено признаков передне-задней миграции брюшных плавников. [ 67 ] Такие недостатки теории жаберных дуг привели к ее раннему упадку в пользу теории боковой плавниковой складки, предложенной Сент-Джорджем Джексоном Мивартом , Фрэнсисом Бальфуром и Джеймсом Кингсли Тэчером .
Теория боковых плавниковых складок предполагала, что парные плавники развиваются из боковых складок вдоль стенки тела рыбы. [ 64 ] Точно так же, как сегментация и почкование срединной плавниковой складки привели к образованию срединных плавников, аналогичный механизм сегментации и удлинения плавниковых зачатков из боковой плавниковой складки, как было высказано, привел к образованию парных грудных и брюшных плавников. Однако в летописи окаменелостей было мало свидетельств перехода от боковой складки к плавнику. [ 68 ] Кроме того, позже филогенетически было продемонстрировано, что грудные и брюшные плавники имеют разное эволюционное и механистическое происхождение. [ 65 ]
Эволюционная биология развития
[ редактировать ]Недавние исследования онтогенеза и эволюции парных придатков сравнили бесплавниковых позвоночных, таких как миноги , с хондриктисами , наиболее базальными из ныне живущих позвоночных с парными плавниками. [ 69 ] В 2006 году исследователи обнаружили, что та же генетическая программа, участвующая в сегментации и развитии срединных плавников, была обнаружена и в развитии парных придатков у кошачьих акул . [ 70 ] Хотя эти данные не подтверждают напрямую гипотезу боковой плавниковой складки, первоначальная концепция общего механизма эволюционного развития срединных парных плавников остается актуальной.
Аналогичное обновление старой теории можно найти в программировании развития хондритиковых жаберных дуг и парных придатков. В 2009 году исследователи из Чикагского университета продемонстрировали, что существуют общие механизмы формирования молекулярного паттерна на раннем этапе развития хондриховой жаберной дуги и парных плавников. [ 71 ] Подобные открытия побудили пересмотреть некогда развенчанную теорию жаберных дуг. [ 68 ]
От плавников до конечностей
[ редактировать ]Рыбы – предки всех млекопитающих, рептилий, птиц и земноводных. [ 72 ] В частности, наземные четвероногие (четвероногие животные) произошли от рыб и совершили свои первые набеги на сушу около 390 миллионов лет назад. [ 73 ] Для передвижения они использовали парные грудные и брюшные плавники. Грудные плавники превратились в передние ноги (руки в случае человека), а брюшные плавники — в задние ноги. [ 74 ] Большая часть генетического механизма, формирующего ходячие конечности четвероногих, уже присутствует в плавательном плавнике рыбы. [ 75 ] [ 76 ]
Аристотель признал различие между аналогичными и гомологичными структурами и сделал следующее пророческое сравнение: «Птицы чем-то напоминают рыб. У птиц крылья находятся в верхней части тела, а у рыб два плавника в передней части тела. У птиц ноги находятся на нижней части тела, а у большинства рыб есть вторая пара плавников в передней части тела. нижней части и возле передних плавников».
- Аристотель, О движении животных. [ 77 ]

В 2011 году исследователи из Университета Монаша в Австралии использовали примитивных, но все еще живых двоякодышащих рыб , «чтобы проследить эволюцию мышц брюшных плавников и выяснить, как развивались несущие нагрузку задние конечности четвероногих». [ 78 ] [ 79 ] Дальнейшие исследования в Чикагском университете показали, что двоякодышащие рыбы, ходящие по дну, уже развили характеристики походки наземных четвероногих. [ 80 ] [ 81 ]
В классическом примере конвергентной эволюции грудные конечности птерозавров , птиц и летучих мышей эволюционировали независимыми путями в летающие крылья. Даже летающие крылья имеют много общего с ходьбой ногами, и основные аспекты генетической модели грудных плавников сохранились. [ 82 ] [ 83 ]
Первые млекопитающие появились в триасовый период (между 251,9 и 201,4 миллиона лет назад). Несколько групп этих млекопитающих начали возвращаться в море, в том числе китообразные (киты, дельфины и морские свиньи). Недавний анализ ДНК показывает, что китообразные произошли от парнокопытных и что у них общий предок с гиппопотамом . [ 84 ] [ 85 ] Около 23 миллионов лет назад другая группа наземных млекопитающих, похожих на медведей, начала возвращаться в море. Это были печати . [ 86 ] То, что у китообразных и тюленей стало ходовыми конечностями, независимо друг от друга развилось в новые формы плавательных плавников. Передние конечности превратились в ласты , а задние конечности либо утратились (китообразные), либо также превратились в ласты (ластоногие). У китообразных хвост на конце обзавелся двумя плавниками, называемыми двуустами . [ 87 ] Рыбьи хвосты обычно вертикальны и движутся из стороны в сторону. Сосальщики китообразных расположены горизонтально и движутся вверх и вниз, поскольку шипы китообразных изгибаются так же, как и у других млекопитающих. [ 88 ] [ 89 ]

Ихтиозавры — древние рептилии, напоминающие дельфинов. Впервые они появились около 245 миллионов лет назад и исчезли около 90 миллионов лет назад.
«Эта морская рептилия, чьи наземные предки так сильно сблизились с рыбами, что у нее фактически развились спинной и хвостовой плавники для улучшения передвижения в воде. Эти структуры тем более примечательны, потому что они произошли из ничего — у предков наземных рептилий не было горба на спине. его спина или лезвие на хвосте служат предшественником». [ 90 ]
Биолог Стивен Джей Гулд сказал, что ихтиозавр был его любимым примером конвергентной эволюции . [ 91 ]
Плавники или ласты различной формы и в разных местах (конечности, тело, хвост) также развились у ряда других групп четвероногих, включая ныряющих птиц, таких как пингвины (измененные из крыльев), морские черепахи (передние конечности превратились в ласты), мозазавры. (конечности преобразованы в ласты) и морские змеи (вертикально расширенный, уплощенный хвостовой плавник).
Роботизированные плавники
[ редактировать ]
| Внешние видео | |
|---|---|
Использование плавников для движения водных животных может быть чрезвычайно эффективным. Было подсчитано, что некоторые рыбы могут достигать эффективности движения более 90%. [ 34 ] Рыба может ускоряться и маневрировать гораздо эффективнее, чем лодки или подводные лодки , и производить меньше волнений и шума в воде. Это привело к биомиметическим исследованиям подводных роботов, которые пытаются имитировать передвижение водных животных. [ 93 ] Примером может служить робот-тунец, созданный Институтом полевой робототехники для анализа и математического моделирования грозообразных движений . [ 94 ] В 2005 году в лондонском аквариуме Sea Life были представлены три роботизированные рыбы, созданные факультетом компьютерных наук Университета Эссекса . Рыбы были созданы, чтобы быть автономными, плавать и избегать препятствий, как настоящие рыбы. Их создатель утверждал, что пытался объединить «скорость тунца, ускорение щуки и навигационные навыки угря». [ 95 ] [ 96 ] [ 97 ]
AquaPenguin Festo , разработанный немецкой компанией , копирует обтекаемую форму и движение передних ласт пингвинов . [ 98 ] [ 99 ] Festo также разработала AquaRay , [ 100 ] АкваДжелли [ 101 ] и АйраКуда , [ 102 ] соответственно имитируя передвижение скатов, медуз и барракуд.
В 2004 году Хью Херр из Массачусетского технологического института создал прототип биомехатронной роботизированной рыбы с живым приводом , хирургическим путем пересадив роботу мышцы с лягушачьих лапок, а затем заставив робота плавать, подавая на мышечные волокна электрические импульсы. [ 103 ] [ 104 ]
Роботизированные рыбы предлагают некоторые исследовательские преимущества, такие как возможность исследовать отдельную часть конструкции рыбы отдельно от остальной рыбы. Однако это рискует чрезмерно упростить биологию, поэтому ключевые аспекты дизайна животных будут упущены из виду. Роботизированные рыбы также позволяют исследователям изменять один параметр, например гибкость или особый контроль движений. Исследователи могут напрямую измерять силы, что непросто сделать с живой рыбой. «Робототехнические устройства также облегчают трехмерные кинематические исследования и коррелирующий гидродинамический анализ, поскольку местоположение локомоторной поверхности может быть точно известно. И отдельные компоненты естественного движения (например, вынос или подъем машущего придатка) могут быть запрограммированы. отдельно, чего, безусловно, трудно добиться при работе с живым животным». [ 105 ]
См. также
[ редактировать ]- Головоногий плавник
- Движение плавников и ласт
- Передвижение рыбы
- Полидактилия у ранних четвероногих
- РобоТуна
- Суп из акульих плавников
- Компромиссы для передвижения в воздухе и воде
- Волнообразное передвижение
Ссылки
[ редактировать ]Цитаты
[ редактировать ]- ^ Стэнден, Э.М. (2009). «Мышечная активность и гидродинамическая функция брюшных плавников форели (Oncorhynchus mykiss) » . Журнал экспериментальной биологии . 213 (5): 831–841. дои : 10.1242/jeb.033084 . ПМИД 20154199 .
- ^ Джин Хелфман, Брюс Коллетт, Дуглас Фейси и Брайан Боуэн. (2009) Разнообразие рыб: биология, эволюция и экология. Джон Уайли и сыновья.
- ^ Jump up to: а б Бендер, Анке; Мориц, Тимо (1 сентября 2013 г.). «Остаток развития и новизна развития - различные способы формирования жировых плавников в ходе онтогенеза». Зоосистематика и эволюция . 89 (2): 209–214. дои : 10.1002/zoos.201300007 . ISSN 1860-0743 .
- ^ Тителл, Э. (2005). «Таинственный маленький толстый плавник» . Журнал экспериментальной биологии . 208 (1): v – vi. дои : 10.1242/jeb.01391 .
- ^ Раймхен, Т.Э.; Темпл, Северная Каролина (2004). «Гидродинамические и филогенетические аспекты жирового плавника рыб» . Канадский журнал зоологии . 82 (6): 910–916. дои : 10.1139/Z04-069 .
- ^ Темпл, Никола (18 июля 2011 г.). «Удаление форели, плавник лосося задевает нерв» . Космос . Архивировано из оригинала 12 января 2014 года.
- ^ Бакленд-Никс, Дж.А.; Гиллис, М.; Реймхен, Т.Э. (2011). «Нейронная сеть обнаружена в предполагаемом рудиментарном признаке: ультраструктуре жирового плавника лососевых» . Труды Королевского общества B: Биологические науки . 279 (1728): 553–563. дои : 10.1098/rspb.2011.1009 . ПМЦ 3234561 . ПМИД 21733904 .
- ^ Стюарт, Томас А.; Смит, В. Лео; Коутс, Майкл И. (2014). «Происхождение жировых плавников: анализ гомоплазии и серийной гомологии придатков позвоночных» . Труды Королевского общества B: Биологические науки . 281 (1781): 20133120. doi : 10.1098/rspb.2013.3120 . ПМЦ 3953844 . ПМИД 24598422 .
- ^ Хайман, Либби (1992). Сравнительная анатомия позвоночных Хаймана (3-е изд.). Издательство Чикагского университета. п. 210. ИСБН 978-0226870137 . Проверено 18 октября 2018 г.
- ^ Jump up to: а б Бро, Джеймс (1936). «Об эволюции костистых рыб в триасовый период». Биологические обзоры . 11 (3): 385–405. дои : 10.1111/j.1469-185X.1936.tb00912.x . S2CID 84992418 .
- ^ фон Зиттель К.А., Вудворд А.С. и Шлоссер М. (1932) Учебник палеонтологии, том 2, Macmillan and Company. Страница 13.
- ^ Коган, Романо (2016). «Переописание Saurichthys madagascariensis Piveteau, 1945 (Actinopterygii, ранний триас) с учетом раннего морфотипа заурихтиид». Журнал палеонтологии позвоночных . 36 (4): e1151886. Бибкод : 2016JVPal..36E1886K . дои : 10.1080/02724634.2016.1151886 . S2CID 87234436 .
- ^ Jump up to: а б Нельсон, Джозеф С. (1994). Рыбы мира . Нью-Йорк: Джон Уайли и сыновья. ISBN 978-0-471-54713-6 . OCLC 28965588 .
- ^ Jump up to: а б «Osteichthyes – Костистая рыба» . Журнал дикой природы для детей . PBS Нью-Гэмпшира. 2023 . Проверено 31 марта 2024 г.
- ^ Шпеер, БР (29 мая 2000 г.). «Знакомство с дипной — двоякодышащей рыбой» . Музей палеонтологии Калифорнийского университета . Проверено 31 марта 2024 г.
- ^ Бискотти, Массачусетс; Гердол, М.; Канапа, А.; Форкони, М.; Олмо, Э.; Паллавичини, А.; Баррука, М.; Шартл, М. (2016). «Транскриптом двоякодышащих рыб: взгляд на события молекулярной эволюции при переходе от воды к суше» . Научные отчеты . 6 : 21571. дои : 10.1038/srep21571 . ПМЦ 4764851 . ПМИД 26908371 . Искусство. № 21571.
- ^ Клак, JA (2002) Набирая силу . Университет Индианы
- ^ Йохансон, Зерина; Лонг, Джон А.; Талант, Джон А.; Жанвье, Филипп; Уоррен, Джеймс В. (2006). «Старейший целакант из раннего девона Австралии» . Письма по биологии . 2 (3): 443–46. дои : 10.1098/rsbl.2006.0470 . ПМК 1686207 . ПМИД 17148426 .
- ^ Фрике, Ганс; Райнике, Олаф; Хофер, Гериберт; Найтингейл, Вернер (1987). «Передвижение целаканта Latimeria Chalumnae в естественной среде». Природа . 329 (6137): 331–33. Бибкод : 1987Nature.329..331F . дои : 10.1038/329331a0 . S2CID 4353395 .
- ^ Такезаки, Н.; Нишихара, Х. (2017). «Поддержка двоякодышащих рыб как ближайшего родственника четвероногих путем использования медленно развивающихся лучепёрых рыб в качестве внешней группы» . Геномная биология и эволюция . 9 (1): 93–101. дои : 10.1093/gbe/evw288 . ПМЦ 5381532 . ПМИД 28082606 .
- ^ Jump up to: а б Зильберберг, Л.; Менье, Ф.Дж.; Лаурин, М. (2016). «Микроанатомическое и гистологическое исследование посткраниального дермального скелета девонского актиноптерига Cheirolepis canadensis » . Acta Palaeontologica Polonica . 61 (2): 363–376. дои : 10.4202/app.00161.2015 .
- ^ Чжан, Дж.; Ваг, П.; Гуай, Д.; Санчес-Пулидо, Л.; Падхи, Британская Колумбия; Корж, В.; Андраде-Наварро, Массачусетс; Акименко, М.А. (2010). «Потеря белков актинотрихии рыб и переход от плавника к конечностям». Природа . 466 (7303): 234–237. Бибкод : 2010Natur.466..234Z . дои : 10.1038/nature09137 . ПМИД 20574421 . S2CID 205221027 .
- ^ Гамлетт 1999 , с. 528.
- ^ Функция гетероцеркального хвоста у акул: количественная динамика следа во время устойчивого горизонтального плавания и вертикального маневрирования - Журнал экспериментальной биологии 205, 2365–2374 (2002)
- ^ «Скелет и органы акулы» . Архивировано из оригинала 5 августа 2010 года . Проверено 14 августа 2009 г.
- ^ Берроу, Кэрол (2021). Акантодии, стебель хондрихтиеса . Издательство Др. Фридрих Пфейль. ISBN 978-3-89937-271-7 . OCLC 1335983356 .
- ^ Андреев, Пламен С.; Сансом, Иван Дж.; Ли, Цян; Чжао, Вэньцзинь; Ван, Цзяньхуа; Ван, Чун-Чье; Пэн, Лицзянь; Цзя, Ляньтао; Цяо, Туо; Чжу, Мин (сентябрь 2022 г.). «Колючий хондрихт из нижнего силура Южного Китая» . Природа . 609 (7929): 969–974. Бибкод : 2022Nature.609..969A . дои : 10.1038/s41586-022-05233-8 . ISSN 1476-4687 . ПМИД 36171377 . S2CID 252570103 .
- ^ Майкл, Брайт. «Челюсти: естественная история акул» . Колумбийский университет. Архивировано из оригинала 24 декабря 2011 года . Проверено 29 августа 2009 г.
- ^ Гроган, Эйлин Д.; Лунд, Ричард (2008). «Базальные пластиножаберные, Thrinacoselache gracia n. gen и sp. (Thrinacodontidae, новое семейство) из известняка Медвежьего ущелья, серпухов, штат Монтана, США». Журнал палеонтологии позвоночных . 28 (4): 970–988. Бибкод : 2008JVPal..28..970G . дои : 10.1671/0272-4634-28.4.970 . S2CID 84735866 .
- ^ Фрей, Линда; Коутс, Майкл; Гинтер, Михал; Хайрапетян, Вачик; Рюклин, Мартин; Джерджен, Иван; Клюг, Кристиан (2019). «Ранний пластиножаберный Phoebodus : филогенетические отношения, экоморфология и новая временная шкала эволюции акул» . Труды Королевского общества B: Биологические науки . 286 (1912). дои : 10.1098/rspb.2019.1336 . ПМК 6790773 . ПМИД 31575362 . S2CID 203619135 .
- ^ Акулий плавник . Международное общество защиты животных.
- ^ Ваннучини С (1999). «Утилизация, маркетинг и торговля акулами» . Технический документ ФАО по рыболовству . 389 . Архивировано из оригинала 2 августа 2017 года . Проверено 21 января 2013 г.
- ^ «В Китае победа за сохранение дикой природы: граждан убедили отказаться от супа из акульих плавников — The Washington Post» . www.washingtonpost.com . Проверено 20 января 2017 г.
- ^ Jump up to: а б с Сфакиотакис, М; Лейн, DM; Дэвис, JBC (1999). «Обзор способов плавания рыб для передвижения в воде» (PDF) . Журнал IEEE океанической инженерии . 24 (2): 237–252. Бибкод : 1999IJOE...24..237S . CiteSeerX 10.1.1.459.8614 . дои : 10.1109/48.757275 . S2CID 17226211 . Архивировано из оригинала (PDF) 24 декабря 2013 года.
- ^ Франк, Жан-Пьер и Мишель, Жан-Мари (2004) Основы кавитации Спрингер. ISBN 9781402022326 .
- ^ Jump up to: а б Брагич, Екатерина (28 марта 2008 г.). «Дельфины плавают так быстро, что это причиняет боль» . Новый учёный . Проверено 31 марта 2008 г.
- ^ Науэн, JC; Лаудер, Г.В. (2001а). «Локомоция скомбридных рыб: визуализация обтекания хвостового стебля и плавников голавля скумбрии Scomber japonicus » . Журнал экспериментальной биологии . 204 (Часть 13): 2251–63. дои : 10.1242/jeb.204.13.2251 . ПМИД 11507109 .
- ^ Науэн, JC; Лаудер, Г.В. (2001б). «Трехмерный анализ кинематики плавников голавля голавля (Scomber japonicus) » . Биологический вестник . 200 (1): 9–19. дои : 10.2307/1543081 . JSTOR 1543081 . ПМИД 11249216 . S2CID 28910289 .
- ^ Науэн, JC; Лаудер, Г.В. (2000). «Локомоция скомбридных рыб: морфология и кинематика плавников голавля Scomber japonicus » (PDF) . Журнал экспериментальной биологии . 203 (Часть 15): 2247–59. дои : 10.1242/jeb.203.15.2247 . ПМИД 10887065 .
- ^ Фламманг, Бельгия; Лаудер, Г.В.; Трулин, ДР; Стрэнд, TE (2011). «Объемная визуализация передвижения рыб» . Письма по биологии . 7 (5): 695–698. дои : 10.1098/rsbl.2011.0282 . ПМК 3169073 . ПМИД 21508026 .
- ^ Фиш, FE; Лаудер, Г.В. (2006). «Пассивное и активное управление потоком плавающих рыб и млекопитающих». Ежегодный обзор механики жидкости . 38 (1): 193–224. Бибкод : 2006AnRFM..38..193F . doi : 10.1146/annurev.fluid.38.050304.092201 . S2CID 4983205 .
- ^ Магнусон Дж. Дж. (1978) «Передвижение скомбридных рыб: гидромеханика, морфология и поведение» в журнале «Физиология рыб» , том 7: «Передвижение», WS Hoar и DJ Randall (Eds) Academic Press. Стр. 240–308. ISBN 9780123504074 .
- ↑ Движение кораблей в море. Архивировано 25 ноября 2011 года на Wayback Machine. Проверено 22 ноября 2012 года.
- ^ Рана и Джоаг (2001) Классическая механика, стр. 391, Tata McGraw-Hill Education. ISBN 9780074603154 .
- ^ Jump up to: а б с Алевизон В.С. (1994) «Путеводитель для Рыб по экологии карибских рифов», издательство Gulf Publishing Company ISBN 1-55992-077-7
- ^ Лингхэм-Солиар, Т. (2005). «Спинной плавник белой акулы Carcharodon carcharias: динамический стабилизатор для быстрого плавания». Журнал морфологии . 263 (1): 1–11. дои : 10.1002/jmor.10207 . ПМИД 15536651 . S2CID 827610 .
- ^ Jump up to: а б Ихтиология Флоридский музей естественной истории . Проверено 22 ноября 2012 г.
- ^ Мастерсон, Дж. « Gambusia affinis » . Смитсоновский институт . Проверено 21 октября 2011 г.
- ^ Кунц, Альберт (1913). «Заметки о повадках, морфологии репродуктивных органов и эмбриологии живородящих рыб Gambusia affinis» . Бюллетень Бюро рыболовства США . 33 : 181–190.
- ^ Капур Б.Г. и Ханна Б. (2004) Справочник по ихтиологии, стр. 497–498, Springer Science & Business Media. ISBN 9783540428541 .
- ^ Хелфман Г., Коллетт Б.Б., Фейси Д.Х. и Боуэн Б.В. (2009) Разнообразие рыб: биология, эволюция и экология, с. 35, Уайли-Блэквелл. ISBN 978-1-4051-2494-2
- ^ Максвелл; и др. (2018). «Переоценка онтогенеза и репродуктивной биологии триасовых рыб Saurichthys (Actinopterygii, Saurichthyidae)». Палеонтология . 61 : 559–574. дои : 10.5061/dryad.vc8h5 .
- ^ «Системный словарь» . ФишБаза . Проверено 15 февраля 2013 г.
- ^ Хейнике, Мэтью П.; Нэйлор, Гэвин Дж. П.; Хеджес, С. Блэр (2009). Древо жизни: хрящевые рыбы (Chondrichthyes) . Издательство Оксфордского университета. п. 320. ИСБН 978-0191560156 .
- ^ Jump up to: а б Водная жизнь мира, стр. 332–333, Marshall Cavendish Corporation, 2000. ISBN 9780761471707 .
- ^ Обзор видов Dement J : Атлантический парусник ( Istiophorus albicans ). Архивировано 17 декабря 2010 г. на сайте Wayback Machinelittoralsociety.org . Проверено 1 апреля 2012 г.
- ^ Бертельсен Э. и Питч Т.В. (1998). Энциклопедия рыб . Сан-Диего: Академическая пресса. стр. 138–139. ISBN 978-0-12-547665-2 .
- ^ Пурпурный летающий гурнард, Dactyloptena orientalis (Cuvier, 1829), Австралийский музей . Обновлено: 15 сентября 2012 г. Дата обращения: 2 ноября 2012 г.
- ^ Фрёзе, Райнер; Поли, Дэниел (ред.) (2012). « Dactyloptena orientalis » в FishBase . Версия от ноября 2012 года.
- ^ Самки рыб выставляют напоказ плавники, чтобы привлечь партнера ScienceDaily . 8 октября 2010 г.
- ^ Балдауф, SA; Баккер, TCM; Гердер, Ф; Куллманн, Х; Тюнкен, Т (2010). «Выбор партнера-самца измеряет аллометрию женского орнамента у цихлиды» . Эволюционная биология BMC . 10 (1): 301. Бибкод : 2010BMCEE..10..301B . дои : 10.1186/1471-2148-10-301 . ПМЦ 2958921 . ПМИД 20932273 .
- ^ Шульц, Кен (2011) Полевой справочник Кена Шульца по морской рыбе, стр. 250, John Wiley & Sons. ISBN 9781118039885 .
- ^ Гудрич, Эдвин С. 1906. «Мемуары: заметки о развитии, структуре и происхождении средних и парных плавников рыб». Журнал клеточных наук, стр. 2-50 (198): 333–76.
- ^ Jump up to: а б Брэнд, Ричард А. (2008). «Происхождение и сравнительная анатомия грудной конечности» . Клиническая ортопедия и связанные с ней исследования . 466 (3): 531–42. дои : 10.1007/s11999-007-0102-6 . ПМК 2505211 . ПМИД 18264841 .
- ^ Jump up to: а б с Коутс, Мичиган (2003). «Эволюция парных плавников». Теория в биологических науках . 122 (2–3): 266–87. doi : 10.1078/1431-7613-00087 (неактивен 20 февраля 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на февраль 2024 г. ( ссылка ) - ^ Гегенбаур, К., Ф. Дж. Белл и Э. Рэй Ланкестер. 1878. Элементы сравнительной анатомии. Карл Гегенбаур... Тр. Ф. Джеффри Белла ... Версия перевода и предисловие, написанное Э. Рэем Ланкестером ... Лондон: Macmillan and Co.,.
- ^ Гудрич, Эдвин С. 1906. «Мемуары: заметки о развитии, структуре и происхождении средних и парных плавников рыб». Журнал клеточных наук , стр. 2-50 (198): 333–76.
- ^ Jump up to: а б Бегеманн, Геррит (2009). «Эволюционная биология развития». Рыбка данио . 6 (3): 303–4. дои : 10.1089/zeb.2009.0593 .
- ^ Коул, Николас Дж.; Карри, Питер Д. (2007). «Идеи акул: эволюционные модели развития плавников» . Динамика развития . 236 (9): 2421–31. дои : 10.1002/dvdy.21268 . ПМИД 17676641 . S2CID 40763215 .
- ^ Фрейтас, Рената; Чжан, Гуанцзюнь; Кон, Мартин Дж. (2006). «Доказательства того, что механизмы развития плавников развились в средней линии ранних позвоночных». Природа . 442 (7106): 1033–37. Бибкод : 2006Natur.442.1033F . дои : 10.1038/nature04984 . ПМИД 16878142 . S2CID 4322878 .
- ^ Гиллис, Дж.А.; Дан, РД; Шубин, Нью-Хэмпшир (2009). «Общие механизмы развития моделируют жаберную дугу позвоночных и парные плавниковые скелеты» . Труды Национальной академии наук . 106 (14): 5720–24. Бибкод : 2009PNAS..106.5720G . дои : 10.1073/pnas.0810959106 . ПМК 2667079 . ПМИД 19321424 .
- ^ «У первобытных рыб были рудиментарные пальцы» ScienceDaily , 23 сентября 2008 г.
- ^ «От плавников к конечностям и от воды к суше» The Harvard Gazzette , 25 ноября 2020 г., Гарвардский университет.
- ^ Холл, Брайан К. (2007) Плавники в конечности: эволюция, развитие и трансформация Издательство Чикагского университета. ISBN 9780226313375 .
- ^ Шубин, Нил (2009) Ваша внутренняя рыба: путешествие в 3,5-миллиардную историю человеческого тела . Винтажные книги. ISBN 9780307277459 . UCTV интервью
- ^ Клак, Дженнифер А. (2012) «От плавников к ногам» , Глава 6, страницы 187–260, в: Gaining Ground, Second Edition: The Origin and Evolution of Tetrapods , Indiana University Press. ISBN 9780253356758 .
- ^ Мур, Джон А. (1988). «[www.sicb.org/dl/saawok/449.pdf «Понимание природы — форма и функция»] Страница 485» . Американский зоолог . 28 (2): 449–584. дои : 10.1093/icb/28.2.449 .
- ↑ Двоякодышащая рыба дает представление о жизни на суше: «Люди — это просто видоизмененные рыбы» ScienceDaily , 7 октября 2011 г.
- ^ Коул, Нью-Джерси; Холл, TE; Дон, ЕК; Бергер, С; Бойсверт, Калифорния; и др. (2011). «Развитие и эволюция мышц брюшного плавника» . ПЛОС Биология . 9 (10): e1001168. дои : 10.1371/journal.pbio.1001168 . ПМК 3186808 . ПМИД 21990962 .
- ^ Маленький шаг для двоякодышащих рыб, большой шаг для эволюции ходьбы» ScienceDaily , 13 декабря 2011 г.
- ^ Кинг, Его Величество; Шубин, Нью-Хэмпшир; Коутс, Мичиган; Хейл, Мэн (2011). «Поведенческие доказательства эволюции ходьбы и прыжков у саркоптеригических рыб до наземного существования» . Труды Национальной академии наук . 108 (52): 21146–21151. Бибкод : 2011PNAS..10821146K . дои : 10.1073/pnas.1118669109 . ПМЦ 3248479 . ПМИД 22160688 .
- ^ Шубин Н.Г.; Табин, С; Кэрролл, С. (1997). «Окаменелости, гены и эволюция конечностей животных» (PDF) . Природа . 388 (6643): 639–648. Бибкод : 1997Natur.388..639S . дои : 10.1038/41710 . ПМИД 9262397 . S2CID 2913898 . Архивировано из оригинала (PDF) 16 сентября 2012 года.
- ^ Полет позвоночных: три решения Калифорнийского университета. Обновлено 29 сентября 2005 г.
- ^ «Ученые обнаружили недостающее звено между дельфином, китом и его ближайшим родственником, бегемотом» . Новости науки ежедневно . 25 января 2005 г. Архивировано из оригинала 4 марта 2007 г. Проверено 18 июня 2007 г.
- ^ Гейтси, Дж. (1 мая 1997 г.). «Большая поддержка ДНК клады китообразных/гиппопотамид: ген белка свертывания крови гамма-фибриноген» . Молекулярная биология и эволюция . 14 (5): 537–543. doi : 10.1093/oxfordjournals.molbev.a025790 . ПМИД 9159931 .
- ^ Флинн Дж.Дж., Финарелли Дж.А., Зер С., Сюй Дж., Недбал М.А. (2005). «Молекулярная филогения хищных животных (млекопитающих): оценка влияния увеличения выборки на разрешение загадочных взаимоотношений» . Систематическая биология . 54 (2): 317–337. дои : 10.1080/10635150590923326 . ПМИД 16012099 .
- ^ Фелтс WJL «Некоторые функциональные и структурные характеристики ласт и сосальщиков китообразных», страницы 255–275 в: Норрис К.С. (ред.) Киты, дельфины и морские свиньи , University of California Press.
- ^ Эволюция китов, Музей Калифорнийского университета . Проверено 27 ноября 2012 г.
- ^ Тьювиссен, JGM; Купер, Л.Н.; Джордж, Джей Си; Баджпай, С (2009). «От суши к воде: происхождение китов, дельфинов и морских свиней» (PDF) . Эво Эду Пропагандистская деятельность . 2 (2): 272–288. дои : 10.1007/s12052-009-0135-2 . S2CID 11583496 .
- ^ Мартилл Д.М. (1993). «Суповые субстраты: среда для исключительного сохранения ихтиозавров из сланцев Посидония (нижняя юра) в Германии». Каупиа – Дармштадтский вклад в естествознание , 2 : 77–97.
- ^ Гулд, Стивен Джей (1993) «Изогнутый из формы» в книге «Восемь поросят: размышления о естествознании» . Нортон, 179–94. ISBN 9780393311396 .
- ^ «Чарли: Робот-рыба ЦРУ» . Центральное разведывательное управление . 4 июня 2013 года. Архивировано из оригинала 16 августа 2013 года . Проверено 12 декабря 2016 г.
- ^ Ричард Мейсон. «Каков рынок рыб-роботов?» . Архивировано из оригинала 4 июля 2009 года.
- ^ Витун Джуварахавонг. «Рыба-робот» . Институт полевой робототехники. Архивировано из оригинала 4 ноября 2007 года . Проверено 25 октября 2007 г.
- ^ «Робот-рыба на базе ПК Gumstix и PIC» . Группа человекоцентрированной робототехники в Университете Эссекса . Проверено 25 октября 2007 г.
- ^ «Рыбы-роботы дебютируют в аквариуме» . cnn.com . CNN . 10 октября 2005 г. Архивировано из оригинала 26 ноября 2020 г. . Проверено 12 июня 2011 г.
- ^ Уолш, Доминик (3 мая 2008 г.). «Merlin Entertainments возглавила список лондонских достопримечательностей с покупкой аквариумов» . thetimes.co.uk . Таймс Лондона . Проверено 12 июня 2011 г.
- ^ Для Festo, Nature Shows the Way Control Engineering , 18 мая 2009 г.
- ^ Бионические пингвины летают в воде... и воздухе Gizmag , 27 апреля 2009 г.
- ↑ Festo AquaRay Robot Technovelgy , 20 апреля 2009 г.
- ↑ Робот-медуза AquaJelly от Festo Engineering TV , 12 июля 2012 г.
- ↑ Легкие роботы: летающий цирк Festo. Архивировано 19 сентября 2015 г. в Wayback Machine The Engineer , 18 июля 2011 г.
- ^ Огромный герр, Д. Роберт Дж. (октябрь 2004 г.). «Плавательный робот, приводимый в действие живой мышечной тканью» . Журнал нейроинженерии и реабилитации . 1 (1): 6. дои : 10.1186/1743-0003-1-6 . ПМК 544953 . ПМИД 15679914 .
- ^ Как работает биомехатроника HowStuffWorks / Проверено 22 ноября 2012 г.
- ^ Лаудер, Г.В. (2011). «Гидродинамика плавания: десять вопросов и технические подходы, необходимые для их решения» (PDF) . Эксперименты с жидкостями . 51 (1): 23–35. Бибкод : 2011ExFl...51...23L . дои : 10.1007/s00348-009-0765-8 . S2CID 890431 .
Библиография
[ редактировать ]- Гамлетт, Уильям К. (1999). Акулы, скаты и скаты: биология пластиножаберных рыб (1-е изд.). Издательство Университета Джонса Хопкинса. п. 56. ИСБН 978-0-8018-6048-5 .
Дальнейшее чтение
[ редактировать ]- Холл, Брайан К. (2007) Плавники в конечности: эволюция, развитие и трансформация Издательство Чикагского университета. ISBN 9780226313375 .
- Хелфман Г., Коллетт Б.Б., Фейси Д.Е. и Боуэн Б.В. (2009) «Функциональная морфология передвижения и питания», глава 8, стр. 101–116. В: Разнообразие рыб: биология , John Wiley & Sons. ISBN 9781444311907 .
- Лаудер, Г.В .; Науэн, JC; Друкер, Э.Г. (2002). «Экспериментальная гидродинамика и эволюция: функция срединных плавников лучепёрых рыб» . Интегр. Комп. Биол . 42 (5): 1009–1017. дои : 10.1093/icb/42.5.1009 . ПМИД 21680382 .
- Лаудер, Г.В.; Друкер, Э.Г. (2004). «Морфология и экспериментальная гидродинамика управляющих поверхностей плавников рыб» (PDF) . Журнал океанической инженерии . 29 (3): 556–571. Бибкод : 2004IJOE...29..556L . дои : 10.1109/joe.2004.833219 . S2CID 36207755 .
Внешние ссылки
[ редактировать ]- Гомология плавниковых лепидотрихий у остихтиевых рыб
- «Рыбий плавник» Сеть земной жизни
- Могут ли рыбы-роботы найти загрязнение? Как все работает . По состоянию на 30 января 2012 г.
| до |  | |
|---|---|---|
| Конечности | ||
| Крылья | ||
| Эволюция | ||
| Связанный | ||

| Базы данных органов управления : Национальные |
|---|