Остракодерма


Остракодермы ( букв. « раковинные шкуры » ) — бронированные бесчелюстные рыбы палеозойской эры . Этот термин сегодня не часто появляется в классификациях, поскольку он парафилетичен (за исключением челюстных рыб ) (может также быть полифилетическим, если анаспиды ближе к круглоротым ) и, таким образом, не соответствует одной эволюционной линии. [1] Тем не менее, этот термин до сих пор используется как неофициальный способ группировки бронированных бесчелюстных рыб.
Новшеством остракодерм стало использование жабр не для питания, а исключительно для дыхания . Более ранние хордовые животные с предшественниками жабр использовали их как для дыхания, так и для питания. [2] У остракодерм были отдельные жаберные глоточные мешки по бокам головы, которые были постоянно открыты и не имели защитной крышки . В отличие от беспозвоночных , которые используют ресничные движения для перемещения пищи, остракодермы использовали свою мускулистую глотку для создания всасывания, которое втягивало в рот мелкую и медленно движущуюся добычу.
Швейцарский анатом Луи Агассис получил несколько окаменелостей костистых панцирных рыб из Шотландии в 1830-х годах. Ему было трудно их классифицировать, так как они не походили ни на одно живое существо. Сначала он сравнил их с современными панцирными рыбами, такими как сом и осетр , но позже понял, что у них отсутствуют подвижные челюсти. Следовательно, в 1844 году он классифицировал их как новую группу, названную «остракодермы», что означает «покрытые панцирем» (от греческого ὄστρακον óstrakon + δέρμα dérma ). [3]
Головы остракодерм покрыты костным щитком. Они являются одними из первых существ с костлявыми головами. Микроскопические слои этого щита кажутся биологам-эволюционистам «словно состоящими из маленьких зубоподобных структур». [4] Нил Шубин пишет: «Разрежьте кость черепа [остракодермы]… рассмотрите ее под микроскопом и… вы обнаружите практически ту же структуру, что и в наших зубах. Там есть слой эмали и даже слой пульпы. Весь щиток состоит из тысяч маленьких зубов, сросшихся вместе. Этот костный череп — один из самых ранних в летописи окаменелостей — полностью состоит из маленьких . зубов использовать по-новому, чтобы защитить их». [4]
Остракодермы существовали в двух основных группах: более примитивных гетеростраканах и цефаласпидах . Цефаласпиды были более продвинутыми, чем гетеростраканы, поскольку у них были боковые стабилизаторы для лучшего контроля над плаванием.
Долгое время считалось, что птераспидоморфы и телодонты были единственными остракодермами с парными ноздрями, в то время как другие группы имели только одну срединную ноздрю. С тех пор выяснилось, что даже если у галеаспиданов есть только одно наружное отверстие, у них есть два внутренних носовых органа. [5] [6]
После появления челюстных рыб (плакодерм, акантодий, акул и др.) около 420 млн лет назад большинство видов остракодерм подверглось упадку, и последние остракодермы вымерли в конце девона . Более поздние исследования показывают, что рыбы с челюстями имели гораздо меньшее отношение к исчезновению остракодерм, чем предполагалось ранее, поскольку они сосуществовали без заметного сокращения в течение примерно 30 миллионов лет. [7]
Подкласс Ostracodermi был помещен в отдел Agnatha вместе с существующим подклассом Cyclostomata , который включает миног и миксин .
Основные группы
[ редактировать ]| Основные группы остракодерм | |||
|---|---|---|---|
| Группа | Сорт | Изображение | Описание |
| Цефаласпидо- морфи | Цефаласпидоморфы или цефаласпиды («головные щитки»), как и большинство современных рыб, были очень хорошо бронированы. Особенно хорошо был развит головной щит, защищающий голову, жабры и передний отдел внутренностей. Корпуса в большинстве форм также были хорошо бронированы. Головной щит имел ряд борозд по всей поверхности, образующих обширный орган боковой линии . Глаза были довольно маленькими и располагались на вершине головы. не было Собственной челюсти . Ротовое отверстие было окружено небольшими пластинками, делавшими губы гибкими, но лишенными возможности кусать. [8] Большинство биологов считают этот таксон вымершим, но это название иногда используется при классификации миног, поскольку когда-то считалось, что миноги связаны с цефаласпидами. Если включить миног, это расширит известный ареал этой группы с силурийского и девонского периодов до наших дней. | ||
| † Галеаспида (вымерший) |  | Галеаспиды («шлем-щитки») имеют на голове массивный костный щиток. Галеаспида обитала на мелководье, в пресной воде и в морской среде в силурийский и девонский период (от 430 до 370 миллионов лет назад) на территории современного Южного Китая, Тибета и Вьетнама. На первый взгляд их морфология больше похожа на морфологию Heterostraci , чем на Osteostraci , а один вид, Tujiaaspis Vividus , имел парные плавники. [9] Galeaspida считаются более близкими родственниками Osteostraci на основании более близкого сходства морфологии черепной коробки . | |
| † Питуриаспида (вымерший) |  | Pituriaspida (« питури -щитки») — небольшая группа вымерших панцирных бесчелюстных рыб с огромными носообразными рострумами , обитавших в морских дельтовых средах среднего девона Австралии (около 390 млн лет назад). Они известны только двум видам: Pituriaspis doylei и Neeyambaspis enigmatica, обнаруженным в одном месте из песчаника в бассейне Джорджина в Западном Квинсленде, Австралия . | |
| † Остеостраки (вымерший) |  | Остеостраки («костяные ракушки») жили на территории современной Северной Америки, Европы и России от среднего силура до позднего девона . С анатомической точки зрения остеостраки, особенно девонские виды, были одними из самых продвинутых из всех известных бесчелюстных. Это связано с развитием парных плавников и их сложной черепной анатомией. Остестраки были больше похожи на миног, чем на челюстных позвоночных, поскольку имели две пары полукружных каналов во внутреннем ухе, в отличие от трех пар, обнаруженных во внутреннем ухе челюстных позвоночных. Считается, что они являются сестринской группой питуриаспидов. Вместе эти два таксона бесчелюстных позвоночных составляют сестринскую группу челюстноротых. Несколько синапоморфий подтверждают эту гипотезу, например наличие: склеротических косточек, парных грудных плавников, кожного скелета с тремя слоями (базальный слой изопедина, средний слой губчатой кости и поверхностный слой дентина) и надхрящевой кости. . [10] | |
| Другой группы | Другие группы | ||
| † Птераспидо- морфи (вымерший) |  | † Птераспидоморфы («крылощитовидные формы») имеют обширную защиту головы. У многих были гипоцеркальные хвосты, которые создавали подъемную силу и облегчали передвижение по воде их бронированных тел, покрытых кожной костью. У них также были сосущие части рта, и некоторые виды, возможно, жили в пресной воде. Таксон содержит подгруппы Heterostraci , Astraspida , Arandaspida . | |
| † Телодонти (вымерший) |  | Thelodonti («слабозубые») — группа мелких вымерших бесчелюстных рыб с характерной чешуей вместо больших пластин панциря. Существует много споров о том, является ли группа палеозойских рыб, известная как Thelodonti (ранее целолепиды), [11] ) представляют собой монофилетическую группу или разрозненные стволовые группы основных линий бесчелюстных и челюстных рыб . Телодонты объединены обладателями « телодонтовой чешуи ». Этот определяющий признак не обязательно является результатом общего происхождения, поскольку он мог быть развит независимо разными группами . Таким образом, обычно считается, что телодонты представляют собой полифилетическую группу. [12] хотя по этому вопросу нет твердого согласия; если они монофилетичны, нет убедительных доказательств того, каким было их предковое состояние. [13] : 206 Телодонты были морфологически очень похожи и, вероятно, тесно связаны с рыбами классов Heterostraci и Anaspida , отличаясь главным образом покрытием из характерных мелких колючих чешуек. Эти чешуйки легко рассеялись после смерти; их небольшой размер и устойчивость сделали их наиболее распространенными ископаемыми позвоночных своего времени. [14] [15] Рыбы жили как в пресноводной, так и в морской среде, впервые появившись в ордовике и исчезнув во время франско-фаменского вымирания в позднем девоне . В основном они были донными обитателями, питавшимися отложениями, хотя есть основания полагать, что некоторые виды перешли в толщу воды и стали свободно плавающими организмами. | |
| † Анаспида (вымерший) | 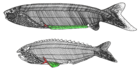 | Анаспида («без щитов») — вымершая группа примитивных бесчелюстных позвоночных, живших в силурийский и девонский периоды. [16] Анаспиды были небольшими морскими бесчелюстными, у которых не было тяжелого костного щита и парных плавников, но они имели поразительный сильно гипоцеркальный хвост. Впервые они появились в раннем силуре и процветали до позднего девона . [17] где большинство видов, за исключением миног , вымерло из-за экологических потрясений, произошедших в то время. | |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Бентон, Майкл (2009) Палеонтология позвоночных, издание 3, стр. 44, John Wiley & Sons. ISBN 9781405144490 .
- ^ Уокер; Лием (1994). Функциональная анатомия позвоночных: эволюционная перспектива (2-е изд.). Издательство колледжа Сандерса. ISBN 0-03-096846-1 .
«В глоточных мешках протохордовых нет жабр, как у рыб; скорее, [фарангеальные] щели протохордовых являются частью их механизма питания». - стр. 32 «Вода всасывается в глотку... Стенка глотки перфорирована множеством вертикально вытянутых глоточных щелей... Клетки эндостиля дна глотки выделяют слизь, которая улавливает мельчайшие частицы пищи». - стр. 35 «Амфиоксус также получает кислород и выделяет углекислый газ из воды, текущей через глотку, хотя жабр нет». - стр. 35
- ^ Мэйси, Джон Г. (1996). Открытие ископаемых рыб (иллюстрированное ред.). Нью-Йорк: Генри Холт и компания. п. 37 . ISBN 9780805043662 .
- ^ Jump up to: а б Шубин, Нил (2009). Ваша внутренняя рыба: путешествие в 3,5-миллиардную историю человеческого тела (переиздание). Нью-Йорк: Книги Пантеона. стр. 85–86. ISBN 9780307277459 .
- ^ «Ископаемая рыба показывает, как развивались челюсти — Planet Earth Online» . Архивировано из оригинала 3 июля 2012 г. Проверено 11 апреля 2014 г.
- ^ GEOL 331 Палеонтология беспозвоночных
- ^ Конструкция челюстей позвоночных заблокирована рано.
- ^ Моралес, Эдвин Х. Колберт, Майкл (1991). Эволюция позвоночных: история позвоночных животных во времени (4-е изд.). Нью-Йорк: Вили-Лисс. ISBN 978-0-471-85074-8 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мертвая рыба вдохнула новую жизнь в эволюционное происхождение плавников и конечностей.
- ^ Сансом, РС (2009). «Филогения, классификация и полярность признаков Osteostraci (Vertebrata)». Журнал систематической палеонтологии . 7 : 95–115. дои : 10.1017/S1477201908002551 . S2CID 85924210 .
- ^ Тернер, С.; Тарлинг, Д.Х. (1982). «Распространение телодонта и других бесчелюстных как тест континентальных реконструкций нижнего палеозоя». Палеогеография, Палеоклиматология, Палеоэкология . 39 (3–4): 295–311. Бибкод : 1982PPP....39..295T . дои : 10.1016/0031-0182(82)90027-X .
- ^ Сарджант, Уильям Энтони С.; Л. Б. Холстед (1995). Окаменелости позвоночных и эволюция научных концепций: сочинения в честь Беверли Холстед . ISBN 978-2-88124-996-9 .
- ^ Донохью, ПК, П.Л. Фори и Р.Дж. Олдридж (2000). «Сродство конодонтов и филогения хордовых». Биологические обзоры Кембриджского философского общества . 75 (2): 191–251. дои : 10.1111/j.1469-185X.1999.tb00045.x . ПМИД 10881388 . S2CID 22803015 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тернер, С. (1999). «Комплексы телодонтов от раннего силура до раннего девона и их возможное экологическое значение». В А. Ж. Буко; Дж. Лоусон (ред.). Палеосообщества, Международная программа геологической корреляции 53, Проект «Экостратиграфия», Заключительный отчет . Издательство Кембриджского университета . стр. 42–78.
- ^ Ранний и средний силур. Видеть Казлев М.А., Уайт Т. (6 марта 2001 г.). «Телодонти» . Палеос.com . Архивировано из оригинала 28 октября 2007 года . Проверено 30 октября 2007 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Альберг, Пер Эрик (2001). Основные события ранней эволюции позвоночных: палеонтология, филогения, генетика и развитие . Вашингтон, округ Колумбия: Тейлор и Фрэнсис. п. 188. ИСБН 0-415-23370-4 .
- ^ Холл, Брайан Кейт; Ханкен, Джеймс (1993). Череп . Чикаго: Издательство Чикагского университета. п. 131. ИСБН 0-226-31568-1 .
