ИРАК4
| ИРАК4 | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | IRAK4 , IPD1, IRAK-4, NY-REN-64, REN64, киназа 4, связанная с рецептором интерлейкина 1, IMD67 | ||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 606883 ; МГИ : 2182474 ; Гомологен : 41109 ; GeneCards : IRAK4 ; ОМА : IRAK4 — ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||




IRAK-4 (киназа 4, связанная с рецептором интерлейкина-1) из семейства IRAK представляет собой протеинкиназу, участвующую в передаче сигналов врожденных иммунных ответов от Toll-подобных рецепторов . Он также поддерживает передачу сигналов от рецепторов Т-клеток. IRAK4 содержит доменные структуры, аналогичные структурам IRAK1 , IRAK2 , IRAK3 и Pelle. IRAK4 уникален по сравнению с IRAK1, IRAK2 и IRAKM тем, что он действует выше других IRAK, но по этому признаку он больше похож на Pelle. IRAK4 имеет важное клиническое применение.
Животные без IRAK-4 более восприимчивы к вирусам и бактериям, но полностью устойчивы к заражению ЛПС .
История
[ редактировать ]Первая киназа, ассоциированная с рецептором IL-1 (IRAK), была обнаружена в 1994 году в ходе экспериментов с линиями Т-хелперных клеток мышей D10N и EL-4. [ 5 ] Два года спустя был клонирован первый экспериментальный член этого семейства киназ IRAK1. [ 6 ] В 2002 году в результате поиска в базе данных Национального центра биотехнологической информации в попытке распознать новых членов семейства IRAK была идентифицирована последовательность кДНК человека, которая кодировала пептид, имеющий значительную гомологию с IRAK1. Было обнаружено, что эта последовательность кДНК имеет пять аминокислотных замен по сравнению с IRAK1 и получила название IRAK4. [ 7 ]
IRAK4 был предложен как гомолог млекопитающих гена Pelle, обнаруженного у Drosophila melanogaster, и было высказано предположение, что для его функционирования при активации NF-κB требуется его киназная активность . Это также было предложено Ли и др. что он может функционировать выше других IRAK и, возможно, вызывать каскад событий фосфорилирования благодаря своей функции киназы IRAK1. [ 7 ] Эта идея о каскаде событий фосфорилирования была подтверждена исследованием, в котором нокаут IRAK4 у мышей показал более тяжелый фенотип, чем другие эксперименты с нокаутом IRAK, и передача сигналов через рецептор Toll/IL-1 (TIR) практически устраняется. [ 7 ]
В 2007 году было обнаружено, что активность IRAK4 необходима для активации сигнальных путей, которые приводят к митоген-активируемым протеинкиназам (MAPK) или иммунным ответам, опосредованным Toll-подобным рецептором (TLR) , но не является существенной для Т-клеточного рецептора (TCR). ) сигнализация, как было первоначально предложено. [ 8 ]
Структура белка
[ редактировать ]IRAK4 представляет собой треонин/сериновую протеинкиназу , состоящую из 460 аминокислот, которая содержит как киназный домен, так и домен смерти . [ 7 ] Его киназный домен демонстрирует типичную двудольную структуру киназ, при этом N-концевая доля состоит из пятинитевого антипараллельного бета-листа и одной альфа-спирали . С-концевая доля состоит в основном из нескольких альфа-спиралей. [ 9 ] В N-конце IRAK4 также содержится удлинение из двадцати аминокислот, что является уникальным для IRAK4 среди киназ, даже внутри семейства IRAK. [ 10 ] Там, где встречаются две доли, находится сайт связывания АТФ, прикрытый тирозиновым привратником. Считается, что тирозин в качестве «привратника» уникален для киназ семейства IRAK. [ 9 ] Белок также содержит три сайта аутофосфорилирования, мутация каждого из которых приводит к снижению киназной активности IRAK4. [ 11 ]
структура аутофосфорилирования петли активации , при которой петля активации Thr345 одного мономера находится в активном центре другого мономера в кристалле (PDB: 4U9A, 4U97). Определена [ 12 ] [ 13 ]
Функция, механизм, сигнальный путь
[ редактировать ]Члены рецептора интерлейкина-1 ( Il-1R ) и суперсемейства Toll-подобных рецепторов имеют общий внутрицитоплазматический домен рецептора Toll-IL-1 ( TIR ), который опосредует рекрутирование комплекса киназы, ассоциированной с рецептором интерлейкина-1 (IRAK) через TIR. -содержащие молекулы-адаптеры. Сигнальный путь TIR-IRAK, по-видимому, имеет решающее значение для защитного иммунитета против конкретных бактерий, но является избыточным против большинства других микроорганизмов . [ 14 ] IRAK4 считается «главным IRAK» в семействе IRAK млекопитающих, поскольку это единственный компонент сигнального пути IL-1/TLR, который абсолютно необходим для его функционирования. Когда один из этих путей стимулируется, клетка начинает вырабатывать провоспалительные сигналы и запускать врожденные иммунные действия. Потеря IRAK4 или его внутренней киназной активности может полностью остановить передачу сигналов по этим путям. [ 15 ]
IRAK4 участвует в путях передачи сигнала, стимулируемых клеточными рецепторами, принадлежащими к суперсемейству рецепторов Toll/Интерлейкин-1. Toll-подобные рецепторы (TLR) стимулируются за счет распознавания патоген-ассоциированных молекулярных структур (PAMPS) , тогда как члены семейства IL-1R стимулируются цитокинами . [ 16 ] Оба играют важную роль в иммунном ответе. Связывание лиганда вызывает конформационные изменения внутриклеточного домена, что позволяет рекрутировать каркасные белки. Один из этих белков, MyD88 , использует свои домены смерти для рекрутирования, ориентации и активации IRAK4. Затем IRAK2 может быть фосфорилирован и соединяется с IRAK4 и MyD88, образуя миддосомный комплекс, который далее фосфорилирует и рекрутирует IRAK1 . [ 17 ] Миддосомный комплекс и IRAK1 рекрутируют и активируют фактор 6, связанный с рецептором TNF (TRAF6) , убиквитиновую протеинлигазу. [ 7 ] TRAF6 может полиубиквитинировать IKK-γ, а также сам себя, что рекрутирует TGF-β-активированную киназу 1 (TAK1), чтобы активировать ее способность фосфорилировать IKK-β . Оба этих пути разрушают IKKγ, что высвобождает NFκB и освобождает его для транслокации в ядро. Кроме того, TAK1 может активировать JNK , индуцируя путь киназы MAP , который приводит к индуцированной AP-1 экспрессии гена. [ 8 ] Вместе AP-1 и NFκB приводят к усилению транскрипции цитокинов, производству молекул адгезии и высвобождению вторичных посредников инфекции. [ 17 ]
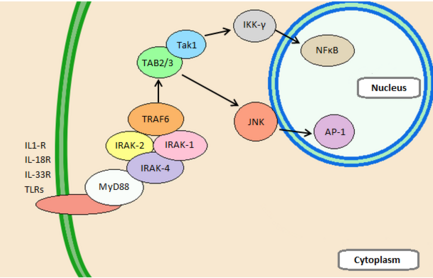
Центральное место во всех этих сигнальных путях занимает киназа IRAK4. Результаты показывают, что IRAK4 является важнейшим компонентом реакции животного на IL-1 . Было обнаружено, что животные с дефицитом этой киназы не способны распознавать вирусные и бактериальные захватчики и полностью устойчивы к летальным дозам липополисахарида (ЛПС). [ 16 ] Это связано с функцией IRAK4 как структурного белка и киназы. Обе эти функции необходимы для образования комплекса миддосом. Кроме того, было показано, что IRAK4 абсолютно необходим для передачи сигналов TLR. Мыши с дефицитом IRAK4 имеют сильно нарушенную способность продуцировать IL-6 , TNF-α и IL-12 в ответ на лиганды TLR. Однако стоит отметить, что, несмотря на свою важность для многих путей передачи иммунных сигналов, IRAK4, по-видимому, не участвует в передаче сигналов TCR. [ 8 ]
Клиническое значение
[ редактировать ]Есть три компонента доказательств, которые иллюстрируют участие IRAK4 в передаче сигналов TLR. Во-первых, IRAK4 является начальной киназой рядом с рецептором TLR, которая активирует нижестоящие эффекторы, такие как цитокины и хемокины, в воспалительном каскаде . [ 7 ] Во-вторых, делеция гена IRAK4 приводит к различным дефектам цитокинового ответа и, наконец, у пациентов с дефицитом IRAK4 наблюдается дефектный иммунитет в ответ на IL-1, IL-8 и другие лиганды, связывающие TLR. [ 16 ] Учитывая положение IRAK4 после этих сигнальных событий, он является важной мишенью лекарственной терапии при различных воспалительных заболеваниях, включая ревматоидный артрит, воспалительные заболевания кишечника и другие аутоиммунные заболевания. [ 17 ]
Рак простаты
[ редактировать ]Важная область исследований, которая в настоящее время изучается [ кем? ] какую роль ген IRAK4 может играть в развитии рака простаты. Существует несколько взаимодействующих факторов, которые приводят к развитию этого заболевания, однако генетическая предрасположенность к хроническому воспалению считается одним из наиболее важных. Было обнаружено, что мутации в гене IRAK4 могут привести к нарушению передачи сигналов TLR и в конечном итоге привести к усилению врожденных иммунных ответов и, следовательно, к усилению воспалительной реакции. Со временем это может привести к развитию рака простаты. [ 18 ]
Меланома
[ редактировать ]Еще одно интересное применение гена IRAK4 было обнаружено в исследовании с участием пациентов с меланомой человека. Это исследование показало, что у пациентов с меланин-клеточными опухолями наблюдается повышение уровня фосфорилирования IRAK4. IRAK4 Ингибирование siRNA у мышей продемонстрировало большую запрограммированную гибель клеток (PCD) и замедлило рост опухоли. [ 17 ]
IRAK4 имеет более высокие уровни в некоторых линиях меланомы. Снижая активность IRAK4, можно будет идентифицировать новые химиотерапевтические агенты для лечения пациентов с поздней стадией меланомы, для которых не существует эффективного лечения. [ 19 ]
Рак поджелудочной железы
[ редактировать ]На модели мышей введение IRAK4 уменьшало передачу воспалительных сигналов, после чего Т-клетки начали атаковать опухоли, а иммунотерапия стала более эффективной. [ 20 ]
Цель по наркотикам
[ редактировать ]Общая проблема в отношении лекарственной терапии или нокдауна IRAK4 заключается в том, что его отсутствие приведет к невыносимым побочным эффектам, учитывая, что IRAK4 играет чрезвычайно центральную роль в сигнальном пути TLR. [ 15 ] Было обнаружено, что у детей с дефицитом IRAK4 снижен иммунитет к некоторым специфическим бактериальным инфекциям, но не к вирусным, паразитарным или другим микробным инфекциям. Однако по мере того, как эти дети становятся взрослыми и материнские антитела больше не присутствуют, восприимчивость к инфекциям становится редкостью. В одном исследовании у всех обследованных пациентов старше 14 лет с дефицитом IRAK4 не было зарегистрировано никаких серьезных бактериальных инфекций. Это может означать, что на более поздних этапах жизни ингибирование IRAK4 может принести пользу против определенных заболеваний, сохраняя при этом иммунитет. [ 21 ]
Следующим шагом в этой области исследований является создание безопасных ингибиторов IRAK4. Был достигнут скромный прогресс в разработке некоторых потенциальных ингибиторов IRAK4, механизм действия которых заключается в блокировании его тирозин-зависимого сайта связывания АТФ. По состоянию на 2007 год [update] Все потенциальные лекарства находятся на ранних доклинических стадиях разработки. [ 22 ]
К 2019 году начались ранние клинические испытания ингибитора IRAK4. [ 23 ] Более того, средства, разрушающие белок IRAK4, недавно вступили в клинические испытания, в первую очередь от компании Kymera Therapeutics. [ 24 ]
Ингибиторы
[ редактировать ]- Эмавусертиб (CA-4948)
- Зимловисертиб (PF-06650833)
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000198001 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000059883 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Мартин М., Бёль Г.Ф., Эрикссон А., Реш К., Бригелиус-Флоэ Р. (июль 1994 г.). «Интерлейкин-1-индуцированная активация протеинкиназы, соосаждающейся с рецептором интерлейкина-1 типа I в Т-клетках». Европейский журнал иммунологии . 24 (7): 1566–1571. дои : 10.1002/eji.1830240717 . ПМИД 8026518 . S2CID 25609420 .
- ^ Цао З, Хензель В.Дж., Гао X (февраль 1996 г.). «IRAK: киназа, связанная с рецептором интерлейкина-1». Наука . 271 (5252): 1128–1131. Бибкод : 1996Sci...271.1128C . дои : 10.1126/science.271.5252.1128 . ПМИД 8599092 . S2CID 42977425 .
- ^ Перейти обратно: а б с д и ж Ли С., Стрелов А., Фонтана Э.Дж., Веше Х. (апрель 2002 г.). «IRAK-4: новый член семейства IRAK со свойствами IRAK-киназы» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (8): 5567–5572. Бибкод : 2002PNAS...99.5567L . дои : 10.1073/pnas.082100399 . ПМК 122810 . ПМИД 11960013 .
- ^ Перейти обратно: а б с Каваго Т., Сато С., Юнг А., Ямамото М., Мацуи К., Като Х. и др. (май 2007 г.). «Основная роль белка IRAK-4 и его киназной активности в иммунных ответах, опосредованных Toll-подобными рецепторами, но не в передаче сигналов TCR» . Журнал экспериментальной медицины . 204 (5): 1013–1024. дои : 10.1084/jem.20061523 . ПМК 2118579 . ПМИД 17485511 .
- ^ Перейти обратно: а б Ван З., Лю Дж., Судом А., Эйрес М., Ли С., Веше Х. и др. (декабрь 2006 г.). «Кристаллические структуры киназы IRAK-4 в комплексе с ингибиторами: серин/треониновая киназа с тирозином в качестве привратника» . Структура . 14 (12): 1835–1844. дои : 10.1016/j.str.2006.11.001 . ПМИД 17161373 .
- ^ Куглстаттер А., Вилласенор А.Г., Шоу Д., Ли С.В., Цинг С., Ню Л. и др. (март 2007 г.). «На переднем крае: структуры киназы 4, ассоциированной с рецептором IL-1, обнаруживают новые особенности и множественные конформации» . Журнал иммунологии . 178 (5): 2641–2645. дои : 10.4049/jimmunol.178.5.2641 . ПМИД 17312103 .
- ^ Ченг Х., Аддона Т., Кешишян Х., Дальстранд Э., Лу С., Дорш М. и др. (январь 2007 г.). «Регуляция активности киназы IRAK-4 посредством аутофосфорилирования внутри петли активации». Связь с биохимическими и биофизическими исследованиями . 352 (3): 609–616. дои : 10.1016/j.bbrc.2006.11.068 . ПМИД 17141195 .
- ^ Феррао Р., Чжоу Х., Шан Ю., Лю К., Ли К., Шоу Д.Э. и др. (сентябрь 2014 г.). «Димеризация и транс-аутофосфорилирование IRAK4 индуцируются сборкой миддосомы» . Молекулярная клетка . 55 (6): 891–903. doi : 10.1016/j.molcel.2014.08.006 . ПМЦ 4169746 . ПМИД 25201411 .
- ^ Сюй К., Малецка К.Л., Финк Л., Джордан Э.Дж., Даффи Э., Коландер С. и др. (декабрь 2015 г.). «Идентификация трехмерных структур комплексов аутофосфорилирования в кристаллах протеинкиназ» . Научная сигнализация . 8 (405): сс13. дои : 10.1126/scisignal.aaa6711 . ПМЦ 4766099 . ПМИД 26628682 .
- ^ Ку CL, Ян К., Бустаманте Дж., Пуэль А., фон Бернут Х., Сантос О.Ф. и др. (февраль 2005 г.). «Наследственные нарушения передачи сигналов Toll-подобных рецепторов человека: иммунологические последствия». Иммунологические обзоры . 203 : 10–20. дои : 10.1111/j.0105-2896.2005.00235.x . ПМИД 15661018 . S2CID 21786295 .
- ^ Перейти обратно: а б Ван З., Веше Х., Стивенс Т., Уокер Н., Йе У.К. (1 января 2009 г.). «Ингибиторы ИРАК-4 при воспалении» . Актуальные темы медицинской химии . 9 (8): 724–737. дои : 10.2174/156802609789044407 . ПМК 3182414 . ПМИД 19689377 .
- ^ Перейти обратно: а б с Сузуки Н., Сузуки С., Дункан Г.С., Миллар Д.Г., Вада Т., Мирцос С. и др. (апрель 2002 г.). «Серьезное нарушение передачи сигналов интерлейкина-1 и Toll-подобных рецепторов у мышей, лишенных IRAK-4». Природа . 416 (6882): 750–756. Бибкод : 2002Natur.416..750S . дои : 10.1038/nature736 . ПМИД 11923871 . S2CID 4428621 .
- ^ Перейти обратно: а б с д и Чаудхари Д., Робинсон С., Ромеро Д.Л. (январь 2015 г.). «Последние достижения в открытии низкомолекулярных ингибиторов киназы 4, связанной с рецептором интерлейкина-1 (IRAK4), в качестве терапевтической мишени при воспалениях и онкологических заболеваниях». Журнал медицинской химии . 58 (1): 96–110. дои : 10.1021/jm5016044 . ПМИД 25479567 .
- ^ Сунь Дж., Виклунд Ф., Сюй Ф.К., Бельтер К., Чжэн С.Л., Йоханссон Дж.Э. и др. (март 2006 г.). «Взаимодействие вариантов последовательностей киназы 4, связанной с рецептором интерлейкина-1, и кластера генов толл-подобного рецептора 6-1-10 увеличивает риск рака простаты». Эпидемиология рака, биомаркеры и профилактика . 15 (3): 480–485. doi : 10.1158/1055-9965.EPI-05-0645 . ПМИД 16537705 . S2CID 25190194 .
- ^ Шривастава Р., Гэн Д., Лю Ю., Чжэн Л., Ли З., Джозеф М.А. и др. (декабрь 2012 г.). «Усиление терапевтического ответа при меланоме путем ингибирования IRAK-1,-4» . Исследования рака . 72 (23): 6209–6216. дои : 10.1158/0008-5472.CAN-12-0337 . ПМЦ 3677596 . ПМИД 23041547 .
- ^ Сомани В., Чжан Д., Додиавала П.Б., Ландер В.Е., Лю Х, Кан Ли и др. (март 2022 г.). «Передача сигналов IRAK4 вызывает устойчивость к иммунотерапии контрольных точек при аденокарциноме протоков поджелудочной железы» . Гастроэнтерология . 162 (7): 2047–2062. дои : 10.1053/j.gastro.2022.02.035 . ПМЦ 9387774 . ПМИД 35271824 .
- ^ Пикард С., Пуэль А., Боннет М., Ку С.Л., Бустаманте Дж., Ян К. и др. (март 2003 г.). «Гноеродные бактериальные инфекции у людей с дефицитом IRAK-4». Наука . 299 (5615): 2076–2079. Бибкод : 2003Sci...299.2076P . дои : 10.1126/science.1081902 . ПМИД 12637671 . S2CID 22438404 .
- ^ Ку CL, фон Бернут Х., Пикард С., Чжан С.Ю., Чанг Х.Х., Ян К. и др. (октябрь 2007 г.). «Избирательная предрасположенность к бактериальным инфекциям у детей с дефицитом IRAK-4: в противном случае IRAK-4-зависимые TLR являются избыточными в защитном иммунитете» . Журнал экспериментальной медицины . 204 (10): 2407–2422. дои : 10.1084/jem.20070628 . ПМК 2118442 . ПМИД 17893200 .
- ^ Curis, Inc. (2019). «Курис впервые продемонстрировал, что нацеливание на IRAK4 у пациентов с рецидивирующей/рефрактерной неходжкинской лимфомой приводит к противораковой активности в продолжающемся исследовании фазы 1» . ООО «Ассоциация PR Newswire».
- ^ Маллард А. (ноябрь 2020 г.). «Деградатор IRAK4 приобретет врожденный иммунитет» . Природная биотехнология . 38 (11): 1221–1223. дои : 10.1038/s41587-020-0724-8 . ПМИД 33144727 . S2CID 226249972 .
Внешние ссылки
[ редактировать ]- IRAK4 + белок, + человек Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
- Обзор всей структурной информации, доступной в PDB для UniProt : Q9NWZ3 (киназа 4, связанная с рецептором интерлейкина-1 человека) на PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q8R4K2 (киназа 4, связанная с рецептором интерлейкина-1 мыши) на PDBe-KB .