Паракринная сигнализация
В клеточной биологии паракринная передача сигналов — это форма клеточной передачи сигналов , тип клеточной коммуникации , при котором клетка вырабатывает сигнал, вызывающий изменения в соседних клетках, изменяя поведение этих клеток. Сигнальные молекулы, известные как паракринные факторы, диффундируют на относительно короткое расстояние (локальное действие), в отличие от передачи сигналов клеткам эндокринными факторами , гормонами , которые перемещаются на значительно большие расстояния через систему кровообращения ; юкстакринные взаимодействия ; и аутокринная сигнализация . Клетки, продуцирующие паракринные факторы, секретируют их в непосредственную внеклеточную среду. Затем факторы перемещаются в близлежащие клетки, в которых градиент полученного фактора определяет результат. Однако точное расстояние, на которое могут распространяться паракринные факторы, не установлено.
Хотя паракринная передача сигналов вызывает разнообразный набор ответов в индуцированных клетках, большинство паракринных факторов используют относительно упорядоченный набор рецепторов и путей. Фактически, известно, что разные органы тела – даже у разных видов – используют одинаковый набор паракринных факторов в дифференцированном развитии. [1] Высококонсервативные рецепторы и пути могут быть организованы в четыре основных семейства на основе сходных структур: факторов роста фибробластов семейство Hedgehog (FGF), семейство , семейство Wnt и суперсемейство TGF-β . Связывание паракринного фактора с соответствующим рецептором инициирует каскады передачи сигнала , вызывая различные ответы.
Паракринные факторы побуждают компетентных ответчиков
[ редактировать ]Чтобы паракринные факторы успешно вызывали ответ в принимающей клетке, эта клетка должна иметь соответствующие рецепторы, доступные на клеточной мембране для приема сигналов, также известные как компетентность . Кроме того, отвечающая клетка также должна обладать способностью механически индуцироваться.
Семейство факторов роста фибробластов (FGF)
[ редактировать ]Хотя семейство паракринных факторов FGF имеет широкий спектр функций, основные результаты подтверждают идею о том, что они в первую очередь стимулируют пролиферацию и дифференцировку. [2] [3] Для выполнения множества разнообразных функций FGF могут подвергаться альтернативному сплайсингу или даже иметь разные инициирующие кодоны для создания сотен различных изоформ FGF . [4]
Одна из наиболее важных функций рецепторов FGF (FGFR) связана с развитием конечностей. В этой передаче сигналов участвуют девять различных альтернативно сплайсированных изоформ рецептора. [5] Fgf 8 и Fgf 10 — два важнейших игрока в развитии конечностей. При зарождении передних конечностей и росте конечностей у мышей аксиальные (продольные) сигналы от промежуточной мезодермы производят Tbx 5, который впоследствии передает той же мезодерме сигнал о выработке Fgf 10. Затем Fgf 10 сигнализирует эктодерме о начале производства Fgf 8, который также стимулирует выработку Fgf 10. Удаление Fgf 10 приводит к появлению мышей без конечностей. [6]
Кроме того, паракринная передача сигналов Fgf важна для развивающихся глаз цыплят. МРНК fgf8 сетчатке локализуется нейральной глазного бокала . в Эти клетки контактируют с клетками внешней эктодермы, которые в конечном итоге станут хрусталиком. [4]
Фенотип и выживаемость мышей после нокаута некоторых генов FGFR: [5]
| Нокаутный ген FGFR | Выживание | Фенотип |
|---|---|---|
| ФГФ 1 | Жизнеспособный | Неясно |
| ФГФ 3 | Жизнеспособный | Внутреннее ухо, скелетная (хвостовая) дифференцировка |
| Рисунок 4 | Смертельный | Пролиферация внутренней клеточной массы |
| ФГФ 8 | Смертельный | Дефект гаструляции , развитие ЦНС, развитие конечностей |
| ФГФ 10 | Смертельный | Развитие нескольких органов (включая конечности, тимус, гипофиз) |
| ФГФ 17 | Жизнеспособный | Развитие мозжечка |
Путь рецепторной тирозинкиназы (RTK)
[ редактировать ]Паракринная передача сигналов через факторы роста фибробластов и соответствующие рецепторы использует рецепторный тирозиновый путь. Этот сигнальный путь был тщательно изучен с использованием глаз дрозофилы и рака человека. [7]
Связывание FGF с FGFR фосфорилирует холостую киназу и активирует путь RTK. Этот путь начинается на поверхности клеточной мембраны, где лиганд связывается со своим специфическим рецептором. Лиганды, которые связываются с RTK, включают факторы роста фибробластов , эпидермальные факторы роста, тромбоцитарные факторы роста и фактор стволовых клеток . [7] Это димеризует трансмембранный рецептор до другого RTK-рецептора, что вызывает аутофосфорилирование и последующее конформационное изменение гомодимеризованного . рецептора Это конформационное изменение активирует спящую киназу каждой RTK на остатке тирозина. В связи с тем, что рецептор проходит через мембрану из внеклеточной среды, через липидный бислой и в цитоплазму , связывание рецептора с лигандом также вызывает трансфосфорилирование цитоплазматического домена рецептора. [8]
Адаптерный белок (такой как SOS) распознает фосфорилированный тирозин на рецепторе. Этот белок действует как мост, соединяющий RTK с промежуточным белком (таким как GNRP), запуская внутриклеточный сигнальный каскад. В свою очередь, промежуточный белок стимулирует GDP-связанный Ras к активированному GTP-связанному Ras. GAP в конечном итоге возвращает Ras в неактивное состояние. Активация Ras может инициировать три сигнальных пути ниже Ras: путь киназы Ras → Raf → MAP, путь киназы PI3 и путь Ral. Каждый путь приводит к активации факторов транскрипции, которые проникают в ядро и изменяют экспрессию генов. [9]

RTK-рецептор и рак
[ редактировать ]Было показано, что паракринная передача сигналов факторов роста между соседними клетками усугубляет канцерогенез . Фактически, мутантные формы одной RTK могут играть причинную роль в самых разных типах рака. Kit Протоонкоген кодирует рецептор тирозинкиназы, лигандом которого является паракринный белок, называемый фактором стволовых клеток (SCF), который важен для кроветворения (образования клеток в крови). [10] Рецептор Kit и родственные ему тирозинкиназные рецепторы на самом деле являются ингибирующими и эффективно подавляют активацию рецептора. Мутантные формы рецептора Kit, которые активируются конститутивно, независимо от лигандов, обнаруживаются при различных раковых новообразованиях. [11]
Путь RTK и рак
[ редактировать ]Исследования рака щитовидной железы пролили свет на теорию о том, что паракринная передача сигналов может способствовать созданию микроокружения опухоли. Транскрипция хемокинов усиливается, когда Ras находится в состоянии, связанном с GTP. Затем хемокины высвобождаются из клетки и могут свободно связываться с другой близлежащей клеткой. Паракринная передача сигналов между соседними клетками создает эту петлю положительной обратной связи. Таким образом, конститутивная транскрипция активированных белков создает идеальную среду для возникновения опухолей. [12] , множественное связывание лигандов с рецепторами RTK чрезмерно стимулирует путь Ras-Raf-MAPK, который сверхэкспрессирует митогенную Фактически и инвазивную способность клеток. [13]
JAK-STAT путь
[ редактировать ]В дополнение к пути RTK факторы роста фибробластов также могут активировать сигнальный путь JAK-STAT . Вместо ковалентно связанных тирозинкиназных доменов рецепторы Jak-STAT образуют нековалентные комплексы с тирозинкиназами класса Jak ( янус-киназы ). Эти рецепторы связываются с эритропоэтином (важным для эритропоэза ), тромбопоэтином (важным для образования тромбоцитов ) и интерфероном (важным для обеспечения функции иммунных клеток). [14]
После димеризации рецепторов цитокинов после связывания лиганда JAK трансфосфорилируют друг друга. Образующиеся фосфотирозины привлекают белки STAT. Белки STAT димеризуются и проникают в ядро, действуя в качестве факторов транскрипции и изменяя экспрессию генов. [14] В частности, STAT транскрибируют гены, которые способствуют пролиферации и выживанию клеток, такие как myc. [15]
Фенотип и выживаемость мышей после нокаута некоторых генов JAK или STAT: [16]
| Нокаут Джин | Выживание | Фенотип |
|---|---|---|
| Як1 | Смертельный | Неврологический дефицит |
| Як2 | Смертельный | Сбой в эритропоэзе |
| Стат1 | Жизнеспособный | карликовости человека и краниосиностоза Синдромы |
| Стат3 | Смертельный | Тканеспецифические фенотипы |
| Стат4 | Жизнеспособный | дефектная дифференцировка Th1, управляемая IL-12, повышенная восприимчивость к внутриклеточным патогенам |
Аберрантный путь JAK-STAT и мутации костей
[ редактировать ]Сигнальный путь JAK-STAT играет важную роль в развитии конечностей, в частности, в его способности регулировать рост костей посредством паракринной передачи сигналов цитокинов. Однако мутации в этом пути связаны с тяжелыми формами карликовости: танатофорной дисплазией (летальной) и ахондроплазионной карликовостью (жизнеспособной). [17] Это связано с мутацией гена Fgf , вызывающей преждевременную и конститутивную активацию транскрипционного фактора Stat1 . Деление хондроцитов преждевременно прекращается, что приводит к летальной карликовости. Клетки пластинок роста костей ребер и конечностей не транскрибируются. Таким образом, неспособность грудной клетки расширяться препятствует дыханию новорожденного. [18]
Путь JAK-STAT и рак
[ редактировать ]Исследование паракринной передачи сигналов через путь JAK-STAT выявило его потенциал в активации инвазивного поведения эпителиальных клеток яичников . Этот эпителиально- мезенхимальный переход очень очевиден при метастазах . [19] Паракринная передача сигналов через путь JAK-STAT необходима при переходе от стационарных эпителиальных клеток к подвижным мезенхимальным клеткам, которые способны вторгаться в окружающие ткани. Было обнаружено, что только путь JAK-STAT индуцирует миграцию клеток. [20]
Семья ёжиков
[ редактировать ]Семейство белков Hedgehog участвует в индукции типов клеток, создании границ тканей и формировании паттерна и обнаружено во всех двусторонних организмах. Белки ежей были впервые обнаружены и изучены у дрозофилы . Белки Hedgehog производят ключевые сигналы для установления строения конечностей и тела плодовых мух, а также гомеостаза взрослых тканей, участвующих в позднем эмбриогенезе и метаморфозе . по крайней мере три гомолога У позвоночных были обнаружены ежа «Drosophila»: звуковой еж, пустынный еж и индийский еж. Sonic hedgehog ( SHH ) играет различную роль в развитии позвонков, опосредуя передачу сигналов и регулируя организацию центральной нервной системы, конечностей и сомитов полярности . Пустынный еж ( DHH ) экспрессируется в клетках Сертоли, участвующих в сперматогенезе . Индийский еж ( IHH ) экспрессируется в кишечнике и хрящах, что играет важную роль в постнатальном росте костей. [21] [22] [23]
Сигнальный путь ежа
[ редактировать ]

Члены семейства белков Hedgehog действуют путем связывания с трансмембранным « патченным » рецептором, который связан с « сглаживанным сигнал Hedgehog » белком, с помощью которого может передаваться . В отсутствие Hedgehog рецептор Patched ингибирует действие Smoothened. Ингибирование Smoothened приводит к тому, что белковый комплекс Cubitus Interruptus (Ci), Fused и Cos, прикрепленный к микротрубочкам, остается интактным. В этой конформации белок Ci расщепляется, так что часть белка может проникнуть в ядро и действовать как репрессор транскрипции . В присутствии Ежика Patched больше не подавляет Smoothened. Тогда активный белок Smoothened способен ингибировать PKA и Slimb, так что белок Ci не расщепляется. Этот интактный белок Ci может проникать в ядро, связываться с белком CPB и действовать как активатор транскрипции , индуцируя экспрессию генов реакции Hedgehog. [23] [24] [25]
Сигнальный путь ежа и рак
[ редактировать ]Сигнальный путь Hedgehog имеет решающее значение для правильного формирования и ориентации тканей во время нормального развития большинства животных. Белки Hedgehog индуцируют пролиферацию клеток в одних клетках и дифференцировку в других. Аберрантная активация пути Hedgehog связана с несколькими типами рака , базальноклеточной карциномой в частности с . Эта неконтролируемая активация белков Hedgehog может быть вызвана мутациями сигнального пути, которые будут независимыми от лигандов , или мутацией, вызывающей сверхэкспрессию белка Hedgehog, которая будет зависеть от лигандов. Кроме того, было показано, что индуцированная терапией активация пути Hedgehog необходима для прогрессирования опухолей простаты после андрогенной депривационной терапии . [26] Эта связь между сигнальным путем Hedgehog и раком человека может обеспечить возможность терапевтического вмешательства в качестве лечения таких видов рака. Сигнальный путь Hedgehog также участвует в нормальной регуляции популяций стволовых клеток и необходим для нормального роста и регенерации поврежденных органов. Это может обеспечить еще один возможный путь онкогенеза через путь Hedgehog. [27] [28] [29]
Семья WNT
[ редактировать ]Семейство белков Wnt включает большое количество цистеином богатых гликопротеинов . Белки Wnt активируют каскады сигнальной трансдукции по трем различным путям: каноническому пути Wnt , неканоническому пути полярности плоских клеток (PCP) и неканоническому пути Wnt/Ca. 2+ путь. Белки Wnt, по-видимому, контролируют широкий спектр процессов развития и считаются необходимыми для контроля ориентации веретена , полярности клеток, кадгерин-опосредованной адгезии и раннего развития эмбрионов у многих различных организмов. Текущие исследования показали, что дерегуляция передачи сигналов Wnt играет роль в формировании опухоли, поскольку на клеточном уровне белки Wnt часто регулируют пролиферацию клеток , морфологию клеток, подвижность клеток и судьбу клеток. [30]
Канонический сигнальный путь Wnt
[ редактировать ]
В каноническом пути белки Wnt связываются со своим трансмембранным рецептором семейства белков Frizzled . Связывание Wnt с белком Frizzled активирует белок Disheveled . В активном состоянии белок Disheveled ингибирует активность фермента киназы гликогенсинтазы 3 ( GSK3 ). Обычно активный GSK3 предотвращает диссоциацию β-катенина с белком APC , что приводит к деградации β-катенина . Таким образом, ингибирование GSK3 позволяет β-катенину диссоциировать от APC, накапливаться и перемещаться в ядро. В ядре β-катенин связывается с транскрипционным фактором Lef/Tcf , который уже действует на ДНК как репрессор, ингибируя транскрипцию генов, с которыми он связывается. Связывание β-катенина с Lef/Tcf действует как активатор транскрипции, активируя транскрипцию Wnt-зависимых генов. [31] [32] [33]
Неканонические сигнальные пути Wnt
[ редактировать ]Неканонические пути Wnt обеспечивают путь передачи сигнала для Wnt, который не включает β-катенин . В неканонических путях Wnt влияет на актин и микротрубочковый цитоскелет , а также на транскрипцию генов .
Путь неканонической планарной клеточной полярности (PCP)
[ редактировать ]
Неканонический путь PCP регулирует морфологию , деление и движение клеток . Белки Wnt снова связываются с Frizzled и активируют его, так что Frizzled активирует белок Disheveled, который привязан к плазматической мембране через белок Prickle и трансмембранный белок Stbm. Активный Disheveled активирует RhoA GTPase через связанный с Disheveled активатор морфогенеза 1 (Daam1) и белок Rac . Активный RhoA способен вызывать изменения цитоскелета путем активации Roh-ассоциированной киназы (ROCK) и напрямую влиять на транскрипцию генов. Active Rac может напрямую вызывать изменения цитоскелета и влиять на транскрипцию генов посредством активации JNK. [31] [32] [33]
Неканонический Wnt/Ca 2+ путь
[ редактировать ]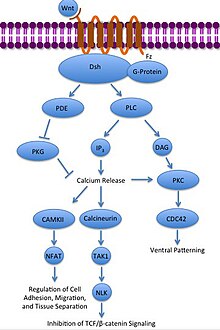
Неканонический Wnt/Ca 2+ Путь регулирует внутриклеточный уровень кальция . Wnt снова привязывается и активируется к Frizzled. Однако в этом случае активированный Frizzled заставляет связанный G-белок активировать фосфолипазу (PLC), которая взаимодействует с PIP 2 и расщепляет его на DAG и IP 3 . Затем IP 3 может связываться с рецептором эндоплазматической сети, высвобождая внутриклеточные запасы кальция и индуцируя кальций-зависимую экспрессию генов. [31] [32] [33]
Сигнальные пути Wnt и рак
[ редактировать ]Сигнальные пути Wnt имеют решающее значение для передачи сигналов между клетками во время нормального развития и эмбриогенеза и необходимы для поддержания взрослой ткани, поэтому нетрудно понять, почему нарушение сигнальных путей Wnt может способствовать дегенеративным заболеваниям и раку человека .
Сигнальные пути Wnt сложны, включают множество различных элементов и, следовательно, имеют множество мишеней для неправильной регуляции. Мутации, которые вызывают конститутивную активацию сигнального пути Wnt, приводят к образованию опухолей и раку. Аберрантная активация пути Wnt может привести к усилению пролиферации клеток. Текущие исследования сосредоточены на действии сигнального пути Wnt, регулирующем выбор стволовых клеток для пролиферации и самообновления. Такое действие передачи сигналов Wnt в возможном контроле и поддержании стволовых клеток может обеспечить возможное лечение рака, демонстрирующего аберрантную передачу сигналов Wnt. [34] [35] [36]
Суперсемейство TGF-β
[ редактировать ]« TGF » (трансформирующий фактор роста) представляет собой семейство белков, включающее 33 члена, которые кодируют димерные секретируемые полипептиды, регулирующие развитие. [37] Многие процессы развития находятся под его контролем, включая гаструляцию, осевую симметрию тела, морфогенез органов и гомеостаз тканей у взрослых. [38] Все лиганды TGF-β связываются с рецепторами типа I или типа II, образуя гетеротетрамные комплексы. [39]
Путь TGF-β
[ редактировать ]Путь TGF-β регулирует многие клеточные процессы в развивающихся эмбрионах и взрослых организмах, включая рост клеток , дифференцировку , апоптоз и гомеостаз . У человека и других млекопитающих существует пять типов рецепторов типа II и семь типов рецепторов типа I. Эти рецепторы известны как «киназы двойной специфичности», поскольку их цитоплазматический киназный домен обладает слабой тирозинкиназной активностью, но сильной серин / треониновой киназной активностью. [40] Когда лиганд суперсемейства TGF-β связывается с рецептором типа II, он рекрутирует рецептор типа I и активирует его путем фосфорилирования остатков серина или треонина его «GS»-бокса. [41] При этом образуется активационный комплекс, который затем может фосфорилировать белки SMAD.

путь SMAD
[ редактировать ]Существует три класса SMAD:
Примеры SMAD в каждом классе: [42] [43] [44]
| Сорт | SMAD |
|---|---|
| Р-СМАД | SMAD1 , SMAD2 , SMAD3 , SMAD5 и SMAD8/9 |
| Ко-СМАД | SMAD4 |
| Я-СМАД | SMAD6 и SMAD7 |
Суперсемейство TGF-β активирует членов семейства SMAD , которые действуют как факторы транскрипции. В частности, рецептор типа I, активированный рецептором типа II, фосфорилирует R-SMAD , которые затем связываются с ко-SMAD, SMAD4 . R-SMAD/Co-SMAD образует комплекс с импортином и поступает в ядро, где они действуют как факторы транскрипции и либо повышают, либо подавляют экспрессию целевого гена.
Специфические лиганды TGF-β приводят к активации SMAD2/3 или SMAD1/5 R-SMAD . Например, когда активин , Nodal или лиганд TGF-β связываются с рецепторами, фосфорилированный рецепторный комплекс может активировать SMAD2 и SMAD3 посредством фосфорилирования. Однако когда лиганд BMP связывается с рецепторами, фосфорилированный рецепторный комплекс активирует SMAD1 и SMAD5 . Затем комплексы Smad2/3 или Smad1/5 образуют димерный комплекс с SMAD4 и становятся факторами транскрипции . Хотя в этом пути участвует множество R-SMAD , есть только один ко-SMAD, SMAD4 . [45]
Не-SMAD путь
[ редактировать ]Сигнальные белки, не относящиеся к Smad, способствуют ответам пути TGF-β тремя способами. Во-первых, сигнальные пути, отличные от Smad, фосфорилируют Smad. Во-вторых, Smads напрямую передают сигнал другим путям, напрямую связываясь с другими сигнальными белками, такими как киназы. Наконец, рецепторы TGF-β напрямую фосфорилируют белки, не относящиеся к Smad. [46]
Члены суперсемейства TGF-β
[ редактировать ]1. Семейство TGF-β
[ редактировать ]В это семейство входят TGF-β1 , TGF-β2 , TGF-β3 и TGF-β5. Они участвуют в положительной и отрицательной регуляции деления клеток , формировании внеклеточного матрикса между клетками, апоптозе и эмбриогенезе . Они связываются с рецептором TGF-β типа II (TGFBRII).
TGF-β1 стимулирует синтез коллагена и фибронектина и ингибирует деградацию внеклеточного матрикса . В конечном итоге он увеличивает выработку внеклеточного матрикса эпителиальными клетками . [39] Белки TGF-β регулируют эпителий, контролируя, где и когда они разветвляются, образуя протоки почек, легких и слюнных желез. [39]
2. Семейство костных морфогенетических белков (BMP).
[ редактировать ]Первоначально было обнаружено, что члены семейства BMP индуцируют костеобразование , как следует из их названия. Однако BMP очень многофункциональны и могут также регулировать апоптоз , миграцию клеток , клеток деление и дифференцировку . Они также определяют переднюю/заднюю ось, вызывают рост и регулируют гомеостаз . [37]
BMP связываются с костным морфогенетическим белковым рецептором типа II (BMPR2). Некоторыми белками семейства BMP являются BMP4 и BMP7 . BMP4 способствует образованию костей, вызывает гибель клеток или сигнализирует о формировании эпидермиса , в зависимости от ткани, на которую он действует. BMP7 имеет решающее значение для развития почек, синтеза сперматозоидов и поляризации нервной трубки. И BMP4 , и BMP7 регулируют стабильность и процессинг зрелых лигандов, включая деградацию лигандов в лизосомах. [37] BMP действуют путем диффузии из клеток, которые их создают. [47]
Другие члены суперсемейства TGF-β
[ редактировать ]- Семья Vg1
- Семья активинов
- Участвует в эмбриогенезе и остеогенезе.
- Регулирует уровень инсулина и гипофиза , половых желез и гипоталамуса. гормонов
- Факторы выживания нервных клеток
- 3 активина: активин А , активин В и активин AB .
- Глиальный нейротрофический фактор (GDNF)
- почек и кишечника. нейронов Необходим для дифференцировки
- Мюллеровский ингибирующий фактор
- Участвует в определении пола млекопитающих.
- Узловой
- Связывается с рецептором активина A типа 2B.
- Образует рецепторный комплекс с рецептором активина A типа 1B или с рецептором активина A типа 1C . [48]
- Факторы роста и дифференциации (GDF)
Сводная таблица сигнального пути TGF-β
[ редактировать ]| Лиганд суперсемейства TGF Beta | Рецептор типа II | Рецептор I типа | R-SMAD | Ко-СМАД | Лигандные ингибиторы |
|---|---|---|---|---|---|
| Активин А | ACVR2A | ACVR1B (АЛК4) | СМАД2 , СМАД3 | SMAD4 | Фоллистатин |
| GDF1 | ACVR2A | ACVR1B (АЛК4) | СМАД2 , СМАД3 | SMAD4 | |
| GDF11 | ACVR2B | ACVR1B (ALK4), TGFβRI (ALK5) | СМАД2 , СМАД3 | SMAD4 | |
| Костные морфогенетические белки | БМПР2 | БМПР1А (АЛК3), БМПР1Б (АЛК6) | СМАД1 СМАД5 , СМАД8 | SMAD4 | Ноггин , Чордин , ДЭН |
| Узловой | ACVR2B | ACVR1B (ALK4), ACVR1C (ALK7) | СМАД2 , СМАД3 | SMAD4 | Левша |
| TGFβs | TGFβRII | TGFβRI (ALK5) | СМАД2 , СМАД3 | SMAD4 | LTBP1 , THBS1 , Декорин |
Примеры
[ редактировать ]Фактор роста и факторы свертывания крови являются паракринными сигнальными агентами. Местное действие передачи сигналов факторов роста играет особенно важную роль в развитии тканей. Кроме того, ретиноевая кислота , активная форма витамина А , функционирует паракринным образом, регулируя экспрессию генов во время эмбрионального развития у высших животных. [49] У насекомых аллатостатин контролирует рост посредством паракринного действия на аллаты. [ нужна ссылка ]
У зрелых организмов паракринная передача сигналов участвует в реакциях на аллергены , восстановлении тканей, образовании рубцовой ткани и свертывании крови . [ нужна ссылка ] Гистамин — это паракринный препарат, который выделяется иммунными клетками бронхиального дерева. Гистамин вызывает сокращение гладкомышечных клеток бронхов, сужая дыхательные пути. [50]
См. также
[ редактировать ]- цАМФ-зависимый путь
- Перекрестные помехи (биология)
- Липидная сигнализация
- Местный гормон - либо паракринный гормон, либо гормон, действующий как паракринно, так и эндокринно.
- Сигнальный путь МАРК
- Netpath – тщательно подобранный ресурс о путях передачи сигналов у человека.
- Паракринный регулятор
Ссылки
[ редактировать ]- ^ «Паракринные факторы» . Проверено 27 июля 2018 г.
- ^ Господирович, Д.; Феррара, Н.; Швайгерер, Л.; Нойфельд, Г. (1987). «Структурная характеристика и биологические функции фактора роста фибробластов». Эндокринные обзоры . 8 (2): 95–114. дои : 10.1210/edrv-8-2-95 . ПМИД 2440668 .
- ^ Рифкин, Дэниел Б.; Москателли, Дэвид (1989). «Последние разработки в клеточной биологии основного фактора роста фибробластов» . Журнал клеточной биологии . 109 (1): 1–6. дои : 10.1083/jcb.109.1.1 . JSTOR 1613457 . ПМК 2115467 . ПМИД 2545723 .
- ^ Jump up to: Перейти обратно: а б Лаппи, Дуглас А. (1995). «Нацеливание на опухоль через рецепторы фактора роста фибробластов». Семинары по биологии рака . 6 (5): 279–88. дои : 10.1006/scbi.1995.0036 . ПМИД 8562905 .
- ^ Jump up to: Перейти обратно: а б Сюй, Дж.; Сюй, Дж; Колвин, Дж.С.; МакИвен, генеральный директор; Макартур, Калифорния; Кулиер, Ф; Гао, Г; Гольдфарб, М (1996). «Рецепторная специфичность семейства факторов роста фибробластов» . Журнал биологической химии . 271 (25): 15292–7. дои : 10.1074/jbc.271.25.15292 . ПМИД 8663044 .
- ^ Логан, М. (2003). «Палец или палец ноги: молекулярная основа идентичности конечностей» . Разработка . 130 (26): 6401–10. дои : 10.1242/dev.00956 . ПМИД 14660539 .
- ^ Jump up to: Перейти обратно: а б Фантл, Венди Дж; Джонсон, Дэниел Э; Уильямс, Льюис Т. (1993). «Передача сигналов рецепторными тирозинкиназами». Ежегодный обзор биохимии . 62 : 453–81. дои : 10.1146/annurev.bi.62.070193.002321 . ПМИД 7688944 .
- ^ Ярден, Йосеф; Ульрих, Аксель (1988). «Тирозинкиназы рецептора фактора роста». Ежегодный обзор биохимии . 57 : 443–78. дои : 10.1146/annurev.bi.57.070188.002303 . ПМИД 3052279 .
- ^ Кац, Майкл Э; Маккормик, Фрэнк (1997). «Передача сигнала от нескольких эффекторов Ras». Текущее мнение в области генетики и развития . 7 (1): 75–9. дои : 10.1016/S0959-437X(97)80112-8 . ПМИД 9024640 .
- ^ Жебо, Кристина М.; Уильямс, Дэвид А.; Гейсслер, Эдвин Н.; Броуди, Вирджиния К.; Мартин, Фрэнсис Х.; Аткинс, Гарри Л.; Сюй, Роу-Инь; Биркетт, Нил К.; Окино, Кеннет Х.; Мердок, Дуглас К.; Якобсен, Фредерик В.; Лэнгли, Кейт Э.; Смит, Кент А.; Такейш, Такаши; Каттанах, Брюс М.; Галли, Стивен Дж.; Саггс, Сидни В. (1990). «Фактор стволовых клеток кодируется в локусе SI мыши и является лигандом тирозинкиназного рецептора c-kit». Клетка . 63 (1): 213–24. дои : 10.1016/0092-8674(90)90302-У . ПМИД 1698556 . S2CID 39924379 .
- ^ Рённстранд, Л. (2004). «Передача сигнала через рецептор фактора стволовых клеток / c-Kit». Клеточные и молекулярные науки о жизни . 61 (19–20): 2535–48. дои : 10.1007/s00018-004-4189-6 . ПМИД 15526160 . S2CID 2602233 .
- ^ Мелилло, Роза Марина; Кастеллоне, Мария Доменика; Гуарино, Валентина; Де Фалько, Валентина; Чирафичи, Анна Мария; Сальваторе, Джулиана; Кайаццо, Фиорина; Басоло, Фульвио; Джаннини, Риккардо; Крухоффер, Могенс; Орнтофт, Торбен; Фуско, Альфредо; Санторо, Массимо (2005). «Линейный сигнальный каскад RET/PTC-RAS-BRAF опосредует подвижный и митогенный фенотип клеток рака щитовидной железы» . Журнал клинических исследований . 115 (4): 1068–81. дои : 10.1172/JCI22758 . ПМЦ 1062891 . ПМИД 15761501 . (Отозвано, см. два : 10.1172/JCI87345 , PMID 27035814 , Часы втягивания )
- ^ Колх, Уолтер (2000). «Значимые отношения: регуляция пути Ras/Raf/MEK/ERK посредством белковых взаимодействий» . Биохимический журнал . 351 (2): 289–305. дои : 10.1042/0264-6021:3510289 . ПМЦ 1221363 . ПМИД 11023813 .
- ^ Jump up to: Перейти обратно: а б Ааронсон, Дэвид С.; Хорват, Курт М. (2002). «Дорожная карта для тех, кто не знает JAK-STAT». Наука . 296 (5573): 1653–5. Бибкод : 2002Sci...296.1653A . дои : 10.1126/science.1071545 . ПМИД 12040185 . S2CID 20857536 .
- ^ Роулингс, Джейсон С.; Рослер, Кристин М.; Харрисон, Дуглас А. (2004). «Сигнальный путь JAK/STAT» . Журнал клеточной науки . 117 (8): 1281–3. дои : 10.1242/jcs.00963 . ПМИД 15020666 .
- ^ О'Ши, Джон Дж; Гадина, Массимо; Шрайбер, Роберт Д. (2002). «Передача сигналов цитокинов в 2002 году: новые сюрпризы на пути Jak/Stat» . Клетка . 109 (2): S121–31. дои : 10.1016/S0092-8674(02)00701-8 . ПМИД 11983158 .
- ^ Шианг, Рита; Томпсон, Лесли М.; Чжу, Я-Чжэнь; Черч, Дина М.; Филдер, Томас Дж.; Боциан, Морин; Винокур, Сара Т.; Васмут, Джон Дж. (1994). «Мутации в трансмембранном домене FGFR3 вызывают наиболее распространенную генетическую форму карликовости — ахондроплазию». Клетка . 78 (2): 335–42. дои : 10.1016/0092-8674(94)90302-6 . ПМИД 7913883 . S2CID 20325070 .
- ^ Каллури, Рагху; Вайнберг, Роберт А. (2009). «Основы эпителиально-мезенхимального перехода» . Журнал клинических исследований . 119 (6): 1420–8. дои : 10.1172/JCI39104 . ПМК 2689101 . ПМИД 19487818 .
- ^ Сильвер, Дебра Л.; Монтелл, Дениз Дж. (2001). «Паракринная передача сигналов через путь JAK/STAT активирует инвазивное поведение эпителиальных клеток яичников у дрозофилы» . Клетка . 107 (7): 831–41. дои : 10.1016/S0092-8674(01)00607-9 . ПМИД 11779460 .
- ^ Ингэм, PW; МакМахон, AP (2001). «Передача сигналов ежа в развитии животных: парадигмы и принципы» . Гены и развитие . 15 (23): 3059–87. дои : 10.1101/gad.938601 . ПМИД 11731473 .
- ^ Битгуд, Марк Дж.; МакМахон, Эндрю П. (1995). « Гены Hedgehog и Bmp совместно экспрессируются во многих различных местах межклеточного взаимодействия в мышином эмбрионе» . Биология развития . 172 (1): 126–38. дои : 10.1006/dbio.1995.0010 . ПМИД 7589793 .
- ^ Jump up to: Перейти обратно: а б Джейкоб, Л.; Лам, Л. (2007). «Сигнальный путь ежа». СТКЭ науки . 2007 (407): см6. doi : 10.1126/stke.4072007cm6 . ПМИД 17925577 . S2CID 35653781 .
- ^ Джонсон, Рональд Л.; Скотт, Мэтью П. (1998). «Новые игроки и загадки сигнального пути Hedgehog» . Текущее мнение в области генетики и развития . 8 (4): 450–6. дои : 10.1016/S0959-437X(98)80117-2 . ПМИД 9729722 .
- ^ Нибаккен, К; Перримон, Н. (2002). «Передача сигнала ежа: недавние открытия». Текущее мнение в области генетики и развития . 12 (5): 503–11. дои : 10.1016/S0959-437X(02)00333-7 . ПМИД 12200154 .
- ^ Любик А.А., Нури М., Труонг С., Гаффари М., Адомат Х.Х., Кори Э., Кокс М.Э., Ли Н., Ганс Э.С., Йенки П., Фам С., Буттян Р. (2016). «Паракринная передача сигналов Sonic Hedgehog вносит значительный вклад в приобретенный стероидогенез в микроокружении опухоли простаты» . Международный журнал рака . 140 (2): 358–369. дои : 10.1002/ijc.30450 . ПМИД 27672740 . S2CID 2354209 .
- ^ Коллинз, RT; Коэн, С.М. (2005). «Генетический экран у дрозофилы для выявления новых компонентов сигнального пути ежа» . Генетика . 170 (1): 173–84. дои : 10.1534/genetics.104.039420 . ПМЦ 1449730 . ПМИД 15744048 .
- ^ Евангелиста, М.; Тиан, Х.; Де Соваж, Ф.Дж. (2006). «Сигнальный путь ежа при раке» . Клинические исследования рака . 12 (20): 5924–8. дои : 10.1158/1078-0432.CCR-06-1736 . ПМИД 17062662 .
- ^ Тайпале, Юсси; Бичи, Филип А. (2001). «Сигнальные пути Hedgehog и Wnt при раке». Природа . 411 (6835): 349–54. Бибкод : 2001Natur.411..349T . дои : 10.1038/35077219 . ПМИД 11357142 . S2CID 4414768 .
- ^ Кадиган, КМ; Нусс, Р. (1997). «Передача сигналов Wnt: общая тема развития животных» . Гены и развитие . 11 (24): 3286–305. дои : 10.1101/gad.11.24.3286 . ПМИД 9407023 .
- ^ Jump up to: Перейти обратно: а б с Дейл, Тревор К. (1998). «Передача сигнала лигандами семейства Wnt» . Биохимический журнал . 329 (Часть 2): 209–23. дои : 10.1042/bj3290209 . ПМК 1219034 . ПМИД 9425102 .
- ^ Jump up to: Перейти обратно: а б с Чен, Си; Ян, Цзюнь; Эванс, Пол М; Лю, Чуньмин (2008). «Сигнализация Wnt: хорошие и плохие» . Acta Biochimica et Biophysica Sinica . 40 (7): 577–94. дои : 10.1111/j.1745-7270.2008.00440.x . ПМК 2532600 . ПМИД 18604449 .
- ^ Jump up to: Перейти обратно: а б с Комия, Юко; Хабас, Раймонд (2008). «Пути передачи сигнала Wnt» . Органогенез . 4 (2): 68–75. дои : 10.4161/org.4.2.5851 . ПМК 2634250 . ПМИД 19279717 .
- ^ Логан, Катриона Ю.; Нуссе, Роэл (2004). «Сигнальный путь Wnt в развитии и заболеваниях». Ежегодный обзор клеточной биологии и биологии развития . 20 : 781–810. CiteSeerX 10.1.1.322.311 . doi : 10.1146/annurev.cellbio.20.010403.113126 . ПМИД 15473860 .
- ^ Люстиг, Б; Беренс, Дж (2003). «Сигнальный путь Wnt и его роль в развитии опухоли». Журнал исследований рака и клинической онкологии . 129 (4): 199–221. дои : 10.1007/s00432-003-0431-0 . ПМИД 12707770 . S2CID 28959851 .
- ^ Нет, Питер; Райс, Кристиан; Кароу, Мариса; Эгея, Вирджиния; Ильмер, Матиас; Йохум, Марианна (2007). «Путь передачи сигнала Wnt в стволовых клетках и раковых клетках: влияние на клеточную инвазию». Обзоры стволовых клеток . 3 (1): 18–29. дои : 10.1007/s12015-007-0001-y . ПМИД 17873378 . S2CID 25793825 .
- ^ Jump up to: Перейти обратно: а б с Бандйопадхьяй, Амитабха; Цудзи, Куниказу; Кокс, Карен; Харф, Брайан Д.; Розен, Вики; Табин, Клиффорд Дж. (2006). «Генетический анализ роли BMP2, BMP4 и BMP7 в формировании паттерна конечностей и скелетогенезе» . ПЛОС Генетика . 2 (12): е216. дои : 10.1371/journal.pgen.0020216 . ПМЦ 1713256 . ПМИД 17194222 .
- ^ Аттисано, Лилиана; Врана, Джеффри Л. (2002). «Передача сигнала суперсемейством TGF-β». Наука . 296 (5573): 1646–7. Бибкод : 2002Sci...296.1646A . дои : 10.1126/science.1071809 . ПМИД 12040180 . S2CID 84138159 .
- ^ Jump up to: Перейти обратно: а б с Врана, Джеффри Л.; Оздамар, Бариш; Ле Рой, Кристина; Бенчабане, Хасина (2008). «Сигнальные рецепторы семейства TGF-β» . В Деринке, Рик; Миядзоно, Кохей (ред.). Семейство TGF-β . ЦШЛ Пресс. стр. 151–77. ISBN 978-0-87969-752-5 .
- ^ тен Дейке, Питер; Хелдин, Карл-Хенрик (2006). «Семья Смад» . В тен Дейке, Питер; Хелдин, Карл-Хенрик (ред.). Передача сигнала Smad: Smads в пролиферации, дифференцировке и заболевании . Белки и регуляция клеток. Том. 5. Дордрехт: Спрингер. стр. 1–13. ISBN 978-1-4020-4709-1 .
- ^ Мустакас, Аристидис (1 сентября 2002 г.). «Смадная сеть сигнализации» . Журнал клеточной науки . 115 (17): 3355–6. дои : 10.1242/jcs.115.17.3355 . ПМИД 12154066 .
- ^ Ву, Цзя-Вэй; Ху, Мин; Чай, Цзицзе; Соан, Джоан; Хьюз, Морган; Ли, Кэри; Риготти, Дэниел Дж.; Кин, Пила; Мьюир, Том В.; Фэрман, Роберт; Массаге, Джоан; Ши, Игун (2001). «Кристаллическая структура фосфорилированного Smad2» . Молекулярная клетка . 8 (6): 1277–89. дои : 10.1016/S1097-2765(01)00421-X . ПМИД 11779503 .
- ^ Павлетич Никола П.; Хата, Йигун; Ло, Акико; Массаге, Роджер С.; Павлетич, Джоан (1997). «Структурная основа мутационной инактивации супрессора опухоли Smad4» . Природа . 388 (6637): 87–93. Бибкод : 1997Natur.388R..87S . дои : 10.1038/40431 . ПМИД 9214508 .
- ^ Ито, Фумико; Асао, Хиронобу; Сугамура, Кадзуо; Хелдин, Карл-Хенрик; Тен Дейке, Питер; Ито, Сусуму (2001). «Стимулирование передачи сигналов костных морфогенетических белков посредством негативной регуляции ингибирующих Smads» . Журнал ЭМБО . 20 (15): 4132–42. дои : 10.1093/emboj/20.15.4132 . ПМК 149146 . ПМИД 11483516 .
- ^ Шмирер, Бернхард; Хилл, Кэролайн С. (2007). «Передача сигнала TGFβ –SMAD: молекулярная специфичность и функциональная гибкость». Nature Reviews Молекулярно-клеточная биология . 8 (12): 970–82. дои : 10.1038/nrm2297 . ПМИД 18000526 . S2CID 131895 .
- ^ Мустакас, Аристидис; Хелдин, Карл-Хенрик (2005). «Не-Smad сигналы TGF-β» . Журнал клеточной науки . 118 (16): 3573–84. дои : 10.1242/jcs.02554 . ПМИД 16105881 .
- ^ Окавара, Бисей; Иэмура, Сюн-Ичиро; Тен Дейке, Питер; Уэно, Наото (2002). «Диапазон действия BMP определяется его N-концевым ядром основной аминокислоты» . Современная биология . 12 (3): 205–9. дои : 10.1016/S0960-9822(01)00684-4 . ПМИД 11839272 .
- ^ Мунир, Садия; Сюй, Госюн; Ву, Яоцзюн; Ян, Бертон; Лала, Пиюш К.; Пэн, Чун (2004). «Nodal и ALK7 ингибируют пролиферацию и индуцируют апоптоз в клетках трофобласта человека» . Журнал биологической химии . 279 (30): 31277–86. дои : 10.1074/jbc.M400641200 . ПМИД 15150278 .
- ^ Дустер, Грегг (сентябрь 2008 г.). «Синтез ретиноевой кислоты и передача сигналов на раннем органогенезе» . Клетка . 134 (6): 921–31. дои : 10.1016/j.cell.2008.09.002 . ПМК 2632951 . ПМИД 18805086 .
- ^
 В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (24 июля 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 17.1 Обзор эндокринной системы. ISBN 978-1-947172-04-3 .
В эту статью включен текст , доступный по лицензии CC BY 4.0 . Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (24 июля 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 17.1 Обзор эндокринной системы. ISBN 978-1-947172-04-3 .
Внешние ссылки
[ редактировать ]- Паракринная + сигнализация в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- «паракринный» в Медицинском словаре Дорланда