Сигнальный путь Wnt
Сигнальные пути Wnt представляют собой группу путей передачи сигнала , которые начинаются с белков , которые передают сигналы в клетку через рецепторы клеточной поверхности . Имя Wnt представляет собой сумку, созданную из названий Wingless и Int-1. [ 1 ] Сигнальные пути Wnt используют либо близлежащую межклеточную связь ( паракринный ), либо межклеточную связь ( аутокринный ). Они высоко эволюционно консервативны у животных, а это означает, что они схожи у всех видов животных, от плодовых мух до людей. [ 2 ] [ 3 ]
Охарактеризованы три сигнальных пути Wnt: канонический путь Wnt , неканонический путь полярности плоских клеток и неканонический путь Wnt/кальций . Все три пути активируются путем связывания лиганда белка Wnt с Frizzled семейства рецептором , который передает биологический сигнал белку Disheveled внутри клетки. Канонический путь Wnt приводит к регуляции генов транскрипции и, как полагают, частично отрицательно регулируется геном SPATS1 . [ 4 ] Неканонический путь планарной клеточной полярности регулирует цитоскелет , который отвечает за форму клетки. Неканонический путь Wnt/кальций регулирует кальций внутри клетки.
Передача сигналов Wnt была сначала идентифицирована из-за ее роли в канцерогенезе , затем из-за ее функции в эмбриональном развитии . Эмбриональные процессы, которые он контролирует, включают формирование паттерна оси тела , клеточной судьбы спецификацию , клеточную пролиферацию и миграцию клеток . Эти процессы необходимы для правильного формирования важных тканей, включая кости, сердце и мышцы. Его роль в эмбриональном развитии была обнаружена, когда генетические мутации в белках пути Wnt привели к образованию аномальных плодовых мух эмбрионов . Более поздние исследования показали, что гены, ответственные за эти аномалии, также влияют на развитие рака молочной железы у мышей. Передача сигналов Wnt также контролирует регенерацию тканей костного мозга, кожи и кишечника взрослых. [ 5 ]
Клиническое значение этого пути было продемонстрировано мутациями , которые приводят к различным заболеваниям, включая рак молочной железы и простаты , глиобластому , диабет II типа и другие. [ 6 ] [ 7 ] В последние годы исследователи сообщили о первом успешном использовании ингибиторов пути Wnt на мышиных моделях заболеваний. [ 8 ]
История и этимология
[ редактировать ]На открытие передачи сигналов Wnt повлияли исследования онкогенных (вызывающих рак) ретровирусов . В 1982 году Роэл Нусс и Гарольд Вармус заразили мышей вирусом опухоли молочной железы , чтобы мутировать мышиные гены и увидеть, какие мутировавшие гены могут вызывать опухоли молочной железы. Они идентифицировали новый мышиный протоонкоген, который назвали int1 (интеграция 1). [ 3 ] [ 9 ]
Int1 высоко консервативен у многих видов, включая человека и дрозофилу . Его присутствие у D. melanogaster привело исследователей к открытию в 1987 году, что ген int1 у дрозофилы на самом деле был уже известным и охарактеризованным геном дрозофилы, известным как Wingless (Wg). [ 3 ] Поскольку предыдущие исследования Кристианы Нюсляйн-Фольхард и Эрика Вишауса (которые принесли им Нобелевскую премию по физиологии и медицине в 1995 году) уже установили функцию Wg как гена полярности сегментов , участвующего в формировании оси тела во время эмбрионального развития , исследователи определили, что int1 млекопитающих, обнаруженный у мышей, также участвует в эмбриональном развитии. [ 10 ]
Продолжение исследований привело к открытию дополнительных генов, связанных с int1; однако, поскольку эти гены не были идентифицированы таким же образом, как int1, номенклатура генов int была неадекватной. Таким образом, семейство int/Wingless стало семейством Wnt, а семейство int1 стало Wnt1. Имя Wnt представляет собой сочетание int и Wg и означает «сайт интеграции, связанный с Wingless». [ 3 ]
Белки
[ редактировать ]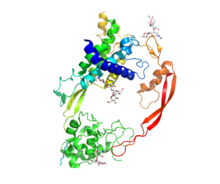
Wnt включает разнообразное семейство секретируемых липид -модифицированных сигнальных гликопротеинов 350–400 аминокислот . длиной [ 11 ] Липидная модификация всех Wnts представляет собой пальмитолеоилирование одного полностью консервативного остатка цистеина. [ 12 ] Пальмитолеоилирование необходимо, поскольку Wnt необходимо для связывания со своим белком-носителем Wntless (WLS), чтобы его можно было транспортировать к плазматической мембране для секреции. [ 13 ] и это позволяет белку Wnt связываться с его рецептором Frizzled [ 14 ] [ 15 ] Белки Wnt также подвергаются гликозилированию , которое присоединяет углеводы для обеспечения правильной секреции. [ 16 ] При передаче сигналов Wnt эти белки действуют как лиганды , активируя различные пути Wnt паракринными и аутокринными путями. [ 2 ] [ 7 ]
Эти белки высококонсервативны у разных видов. [ 3 ] Их можно обнаружить у мышей, людей, Xenopus , рыбок данио , дрозофилы и многих других. [ 17 ]
| Разновидность | Wnt-белки |
|---|---|
| Мудрый человек | WNT1 , WNT2 , WNT2B , WNT3 , WNT3A , WNT4 , WNT5A , WNT5B , WNT6 , WNT7A , WNT7B , WNT8B , WNT9A , WNT9B , WNT10A , WNT10B , WNT11 , WNT8A , WNT16 |
| Mus musculus (идентичны белкам H. sapiens) | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt6, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt9A, Wnt9B, Wnt10A, Wnt10B, Wnt11, Wnt16 |
| Ксеноп | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt10A, Wnt10B, Wnt11, Wnt11R |
| Дания рерио | Wnt1, Wnt2, Wnt2B, Wnt3, Wnt3A, Wnt4, Wnt5A, Wnt5B, Wnt6, Wnt7A, Wnt7B, Wnt8A, Wnt8B, Wnt10A, Wnt10B, Wnt11, Wnt16 |
| Дрозофила | Wg, ДВнт2, ДВнт3/5, ДВнт 4, ДВнт6, ВнтД/ДВнт8, ДВнт10 |
| Гидра | hywnt1, hywnt5a, hywnt8, hywnt7, hywnt9/10a, hywnt9/10b, hywnt9/10c, hywnt11, hywnt16 |
| К. Элеганс | мама-2, лин-44, эгл-20, собака-1, собака-2 [ 18 ] |
Механизм
[ редактировать ]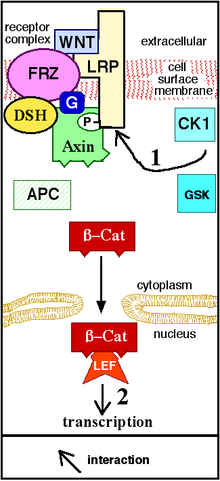

Фундамент
[ редактировать ]Передача сигналов Wnt начинается, когда белок Wnt связывается с N-концевым внеклеточным, богатым цистеином доменом рецептора семейства Frizzled (Fz). [ 19 ] Эти рецепторы охватывают плазматическую мембрану семь раз и составляют отдельное семейство рецепторов, связанных с G-белком (GPCR). [ 20 ] Однако для облегчения передачи сигналов Wnt корецепторы наряду с взаимодействием между белком Wnt и рецептором Fz могут потребоваться . Примеры включают белок, родственный рецептору липопротеина ( LRP )-5/6, рецепторную тирозинкиназу (RTK) и ROR2 . [ 7 ] При активации рецептора сигнал передается фосфопротеину Растрепанный ( Dsh), который находится в цитоплазме . Этот сигнал передается посредством прямого взаимодействия между Fz и Dsh. Белки Dsh присутствуют во всех организмах, и все они имеют следующие высококонсервативные белковые домены : аминоконцевой домен DIX, центральный домен PDZ и карбоксиконцевой домен DEP . Эти разные домены важны, поскольку после Dsh сигнал Wnt может разветвляться на множество путей, и каждый путь взаимодействует с различной комбинацией трех доменов. [ 21 ]
Канонические и неканонические пути
[ редактировать ]Тремя наиболее охарактеризованными сигнальными путями Wnt являются канонический путь Wnt, неканонический путь полярности плоских клеток и неканонический путь Wnt/кальций. Как следует из их названий, эти пути принадлежат к одной из двух категорий: каноническим и неканоническим. Разница между категориями заключается в том, что канонический путь включает белок бета-катенин (β-катенин), тогда как неканонический путь действует независимо от него. [ 19 ]

Канонический путь
[ редактировать ]Канонический путь Wnt (или путь Wnt/ β-катенин ) — это путь Wnt, который вызывает накопление β-катенина в цитоплазме и его возможную транслокацию в ядро , чтобы действовать как транскрипционный коактиватор транскрипционных факторов , принадлежащих TCF/ Семья ЛЕФ . Без Wnt β-катенин не накапливался бы в цитоплазме, поскольку комплекс деструкции обычно разрушает его. В этот комплекс разрушения входят следующие белки: аксин , аденоматозная полипозная палочка (APC), протеинфосфатаза 2А (PP2A), киназа гликогенсинтазы 3 (GSK3) и казеинкиназа 1α (CK1α). [ 22 ] [ 23 ] Он разрушает β-катенин, направляя его на убиквитинирование , которое впоследствии отправляет его в протеасому для переваривания. [ 19 ] [ 24 ] Однако как только Wnt связывает Fz и LRP5 / 6 , функция комплекса деструкции нарушается. Это связано с тем, что Wnt вызывает транслокацию негативного регулятора Wnt, аксина, и деструктивного комплекса на плазматическую мембрану. Фосфорилирование другими белками комплекса разрушения впоследствии связывает аксин с цитоплазматическим хвостом LRP5/6. Аксин дефосфорилируется, его стабильность и уровень снижаются. Затем Dsh активируется посредством фосфорилирования, и его домены DIX и PDZ ингибируют активность GSK3 деструктивного комплекса. Это позволяет β-катенину накапливаться и локализоваться в ядре, а затем вызывать клеточный ответ посредством трансдукции гена наряду с TCF/LEF (фактор усиления Т-клеток/фактор усиления лимфоида). [ 25 ] транскрипционные факторы. [ 24 ] β-катенин рекрутирует другие коактиваторы транскрипции, такие как BCL9 , Pygopus [ 26 ] и Парафибромин/Хиракс. [ 27 ] Сложность транскрипционного комплекса, собранного β-катенином, начинает проявляться благодаря новым высокопроизводительным протеомным исследованиям. [ 28 ] Однако единая теория того, как β-катенин управляет экспрессией целевых генов, до сих пор отсутствует, и тканеспецифичные игроки могут помочь β-катенину определить его гены-мишени. [ 29 ] Обширность белков, взаимодействующих с β-катенином, усложняет наше понимание: β-катенин может напрямую фосфорилироваться по Ser552 с помощью Akt, что вызывает его диссоциацию от межклеточных контактов и накопление в цитозоле, после чего 14-3-3ζ взаимодействует с β-катенином. (pSer552) и усиливает его ядерную транслокацию. [ 30 ] BCL9 и Pygopus Сообщалось, что фактически обладают несколькими β-catenin -независимыми функциями (следовательно, вероятно, независимыми от передачи сигналов Wnt). [ 31 ] [ 32 ] [ 33 ]
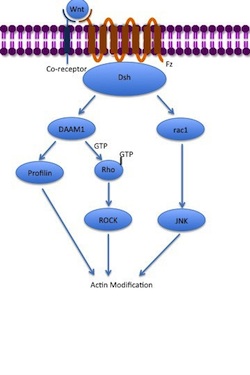
Неканонические пути
[ редактировать ]Путь неканонической планарной клеточной полярности (PCP) не включает β-катенин. Он не использует LRP-5/6 в качестве корецептора и, как полагают, использует NRH1 , Ryk , PTK7 или ROR2 . Путь PCP активируется посредством связывания Wnt с Fz и его корецептором. Затем рецептор рекрутирует Dsh , который использует свои домены PDZ и DIX для формирования комплекса с Dishavelled-ассоциированным активатором морфогенеза 1 ( DAAM1 ). Затем Daam1 активирует небольшой G-белок Rho посредством фактора обмена гуанина . Rho активирует Rho-ассоциированную киназу (ROCK), которая является одним из основных регуляторов цитоскелета . Dsh также образует комплекс с rac1 и опосредует профилина связывание с актином . Rac1 активирует JNK и также может приводить к актина полимеризации . Связывание профилина с актином может привести к реструктуризации цитоскелета и гаструляции . [ 7 ] [ 34 ]

Неканонический путь Wnt/кальций также не включает β-катенин . Его роль заключается в том, чтобы помочь регулировать высвобождение кальция из эндоплазматического ретикулума (ЭР), чтобы контролировать внутриклеточные уровни кальция. Как и другие пути Wnt, при связывании лиганда активированный рецептор Fz напрямую взаимодействует с Dsh и активирует специфические домены белка Dsh. Домены, участвующие в передаче сигналов Wnt/кальций, представляют собой домены PDZ и DEP. [ 7 ] Однако, в отличие от других путей Wnt, рецептор Fz напрямую взаимодействует с тримерным G-белком. Эта совместная стимуляция Dsh и G-белка может привести к активации либо PLC , либо цГМФ-специфической ФДЭ . Если PLC активирован, компонент плазматической мембраны PIP2 расщепляется на DAG и IP3 . Когда IP3 связывает свой рецептор на ЭР, высвобождается кальций. Повышенные концентрации кальция и DAG могут активировать Cdc42 через PKC . Cdc42 является важным регулятором формирования вентрального паттерна. Повышенное содержание кальция также активирует кальциневрин и CaMKII . CaMKII индуцирует активацию транскрипционного фактора NFAT , который регулирует клеточную адгезию, миграцию и разделение тканей. [ 7 ] Кальцинеурин активирует киназу TAK1 и NLK , которая может мешать передаче сигналов TCF/β-катенина в каноническом пути Wnt. [ 35 ] Однако если ФДЭ активирована, высвобождение кальция из ЭР ингибируется. ФДЭ опосредует это посредством ингибирования ПКГ, что впоследствии вызывает ингибирование высвобождения кальция. [ 7 ]
Интегрированный путь Wnt
[ редактировать ]Бинарное различие канонических и неканонических сигнальных путей Wnt подверглось тщательному изучению, и был предложен интегрированный, конвергентный путь Wnt. [ 36 ] Некоторые доказательства этого были найдены для одного лиганда Wnt (Wnt5A). [ 37 ] Доказательства конвергентного сигнального пути Wnt, который демонстрирует интегрированную активацию передачи сигналов Wnt/Ca2+ и Wnt/ β-катенина для множественных лигандов Wnt, были описаны в клеточных линиях млекопитающих. [ 38 ]
Другие пути
[ редактировать ]Передача сигналов Wnt также регулирует ряд других сигнальных путей, которые еще не так подробно изучены. Один из таких путей включает взаимодействие между Wnt и GSK3 . Во время роста клеток Wnt может ингибировать GSK3, чтобы активировать mTOR в отсутствие β-катенина. Однако Wnt также может служить негативным регулятором mTOR посредством активации опухолевого супрессора TSC2 , который активируется посредством взаимодействия Dsh и GSK3. [ 39 ] Во время миогенеза Wnt использует PA и CREB для активации MyoD и Myf5 . генов [ 40 ] Wnt также действует совместно с Ryk и Src, обеспечивая регуляцию отталкивания нейронов во время наведения аксонов . Wnt регулирует гаструляцию , когда CK1 служит ингибитором Rap1-АТФазы для модуляции цитоскелета во время гаструляции. Дальнейшая регуляция гаструляции достигается, когда Wnt использует ROR2 вместе с путем CDC42 и JNK для регулирования экспрессии PAPC . Dsh может также взаимодействовать с aPKC, Pa3 , Par6 и LGl , чтобы контролировать полярность клеток и развитие цитоскелета микротрубочек . Хотя эти пути перекрываются с компонентами, связанными с передачей сигналов PCP и Wnt/кальция, они считаются отдельными путями, поскольку вызывают разные ответы. [ 7 ]
Регулирование
[ редактировать ]Чтобы обеспечить правильное функционирование, передача сигналов Wnt постоянно регулируется в нескольких точках сигнальных путей. [ 41 ] Например, белки Wnt пальмитоилированы . Белок дикобраза опосредует этот процесс, а это означает, что он помогает регулировать момент секреции лиганда Wnt, определяя, когда он полностью сформирован. Секреция дополнительно контролируется с помощью белков, таких как GPR177 (wntless), и равномерности прерывания , а также комплексов, таких как ретромерный комплекс. [ 7 ] [ 24 ]
При секреции лиганду можно предотвратить достижение своего рецептора за счет связывания белков, таких как стабилизаторы Dally и глипикан 3 (GPC3), которые ингибируют диффузию. В раковых клетках обе цепи гепарансульфата [ 42 ] [ 43 ] и основной белок [ 44 ] [ 45 ] GPC3 участвуют в регуляции связывания Wnt и активации пролиферации клеток. [ 46 ] [ 47 ] Wnt распознает структуру гепарансульфата на GPC3, которая содержит IdoA2S и GlcNS6S, а 3-O-сульфатирование в GlcNS6S3S усиливает связывание Wnt с гепарансульфатглипиканом. [ 48 ] Было идентифицировано, что богатый цистеином домен в N-доле GPC3 образует гидрофобную бороздку, связывающую Wnt, включающую фенилаланин-41, который взаимодействует с Wnt. [ 45 ] [ 49 ] Блокирование связывающего домена Wnt с помощью нанотела под названием HN3 может ингибировать активацию Wnt. [ 45 ]
Связывание с рецептором Fz белков, отличных от Wnt, может противодействовать передаче сигналов. Специфические антагонисты включают Диккопфа (Dkk), фактор, ингибирующий Wnt 1 (WIF-1), [ 50 ] [ 51 ] секретируемые белки, связанные с Frizzled (SFRP), Cerberus , Frzb , Wise , SOST и Naked Cuticle . Они представляют собой ингибиторы передачи сигналов Wnt. Однако другие молекулы также действуют как активаторы. Норрин и R-спондин2 активируют передачу сигналов Wnt в отсутствие лиганда Wnt.
Взаимодействия между сигнальными путями Wnt также регулируют передачу сигналов Wnt. Как упоминалось ранее, путь Wnt/кальций может ингибировать TCF/β-катенин, предотвращая передачу сигналов канонического пути Wnt. [ 7 ] [ 24 ] Простагландин E2 (PGE2) является важным активатором канонического сигнального пути Wnt. Взаимодействие PGE2 с его рецепторами E2/E4 стабилизирует β-катенин посредством фосфорилирования, опосредованного цАМФ/ПКА. Синтез PGE2 необходим для процессов, опосредованных передачей сигналов Wnt, таких как регенерация тканей и контроль популяции стволовых клеток у рыбок данио и мышей. [ 5 ] Интересно, что неструктурированные области нескольких крупных внутренне неупорядоченных белков играют решающую роль в регуляции передачи сигналов Wnt. [ 52 ]
Индуцированные клеточные ответы
[ редактировать ]Эмбриональное развитие
[ редактировать ]Передача сигналов Wnt играет решающую роль в эмбриональном развитии. Он действует как на позвоночных , так и на беспозвоночных , включая человека, лягушек, рыбок данио, C. elegans , дрозофилу и других. Впервые он был обнаружен в полярности сегментов дрозофилы, где он помогает установить переднюю и заднюю полярности. Он участвует в других процессах развития . Как предполагает его функция у дрозофилы , он играет ключевую роль в формировании осей тела , особенно в формировании переднезадней и дорсовентральной осей. Он участвует в индукции дифференцировки клеток , что приводит к образованию важных органов, таких как легкие и яичники . Wnt также обеспечивает развитие этих тканей посредством правильной регуляции клеток пролиферации и миграции . Функции передачи сигналов Wnt можно разделить на формирование паттерна оси, спецификацию клеточной судьбы, клеточную пролиферацию и клеточную миграцию. [ 53 ]
Осевое моделирование
[ редактировать ]На раннем этапе развития эмбриона формирование основных осей тела является решающим шагом в установлении общего плана тела организма. Оси включают переднезаднюю ось, дорсовентральную ось и право-левую ось. Передача сигналов Wnt участвует в формировании переднезадней и дорсовентральной (DV) осей. Активность передачи сигналов Wnt в передне-заднем развитии можно наблюдать у млекопитающих, рыб и лягушек. У млекопитающих примитивная полоска и другие окружающие ткани продуцируют морфогенные соединения Wnts, BMPs , FGFs , Nodal и ретиноевую кислоту для образования задней области во время поздней гаструлы . Эти белки образуют градиенты концентрации. Области с наибольшей концентрацией определяют заднюю область, а области с наименьшей концентрацией указывают на переднюю область. У рыб и лягушек β-catenin, продуцируемый канонической передачей сигналов Wnt, вызывает образование организующих центров, которые, наряду с BMPs, вызывают формирование задних частей. Участие Wnt в формировании оси DV можно увидеть по активности формирования Организатор Спемана , устанавливающий спинную область. Каноническая передача сигналов Wnt вырабатывает β-катенин, индуцируя образование этого организатора посредством активации генов Twin и Siamois. [ 36 ] [ 53 ] Сходным образом, при гаструляции птиц клетки серпа Коллера экспрессируют различные мезодермальные маркерные гены, которые обеспечивают дифференциальное движение клеток во время формирования первичной полоски. За это движение отвечает передача сигналов Wnt, активируемая FGF. [ 54 ] [ 55 ]
Передача сигналов Wnt также участвует в формировании осей определенных частей тела и систем органов на более поздних стадиях развития. У позвоночных морфогенетические сигнальные градиенты sonic hedgehog (Shh) и Wnt устанавливают дорсовентральную ось центральной нервной системы во время формирования осевого паттерна нервной трубки . Высокая передача сигналов Wnt устанавливает дорсальную область, тогда как высокая передача сигналов Shh указывает на вентральную область. [ 56 ] Wnt участвует в формировании DV центральной нервной системы посредством участия в наведении аксонов . Белки Wnt направляют аксоны спинного мозга в передне-заднем направлении. [ 57 ] Wnt также участвует в формировании оси DV конечностей. В частности, Wnt7a помогает создавать дорсальный паттерн развивающейся конечности. [ 36 ] [ 53 ]
В модели развития волн эмбриональной дифференцировки Wnt играет решающую роль как часть сигнального комплекса в компетентных клетках, готовых к дифференцировке. Wnt реагирует на активность цитоскелета, стабилизируя первоначальные изменения, вызванные проходящей волной сокращения или расширения, и одновременно сигнализирует ядру, используя различные сигнальные пути, о том, в какой волне участвовала отдельная клетка. Таким образом, активность Wnt усиливается. механическая передача сигналов, возникающая во время развития. [ 58 ] [ 59 ]
Спецификация судьбы клеток
[ редактировать ]Спецификация клеточной судьбы или дифференцировка клеток — это процесс, при котором недифференцированные клетки могут стать более специализированным типом клеток. Передача сигналов Wnt индуцирует дифференцировку плюрипотентных стволовых клеток в мезодермы и энтодермы клетки-предшественники . [ 60 ] Эти клетки-предшественники далее дифференцируются в типы клеток, такие как эндотелиальные, сердечные и сосудистые гладкомышечные линии. [ 61 ] Передача сигналов Wnt индуцирует образование крови из стволовых клеток. В частности, Wnt3 приводит к образованию мезодермальных коммитированных клеток с гемопоэтическим потенциалом. [ 62 ] Wnt1 противодействует нейрональной дифференцировке и является основным фактором самообновления нервных стволовых клеток. Это обеспечивает регенерацию клеток нервной системы, что является еще одним доказательством его роли в пролиферации нервных стволовых клеток. [ 60 ] Передача сигналов Wnt участвует в зародышевых клеток детерминации кишечника , спецификации тканей , развитии волосяных фолликулов , развитии легочной ткани, клеток ствола нервного гребня дифференцировке , развитии нефронов , развитии яичников и определении пола . [ 53 ] Передача сигналов Wnt также препятствует формированию сердца, и было показано, что ингибирование Wnt является критическим индуктором сердечной ткани во время развития. [ 63 ] [ 64 ] [ 65 ] и низкомолекулярные ингибиторы Wnt обычно используются для получения кардиомиоцитов из плюрипотентных стволовых клеток. [ 66 ] [ 67 ]
Пролиферация клеток
[ редактировать ]Для того чтобы произошла массовая дифференциация клеток, необходимая для формирования определенных клеточных тканей разных организмов, пролиферация и рост эмбриональных стволовых клеток должны иметь место . Этот процесс опосредован канонической передачей сигналов Wnt, которая увеличивает ядерный и цитоплазматический β-катенин. Увеличение β-катенина может инициировать транскрипционную активацию белков, таких как циклин D1 и c-myc , которые контролируют G1 в S фазовый переход в клеточном цикле . Переход в фазу S вызывает репликацию ДНК и, в конечном итоге, митоз , которые отвечают за пролиферацию клеток. [ 68 ] Это увеличение пролиферации напрямую связано с дифференцировкой клеток, поскольку по мере пролиферации стволовые клетки также дифференцируются. Это обеспечивает общий рост и развитие определенных тканевых систем во время эмбрионального развития. Это очевидно в таких системах, как система кровообращения, где Wnt3a приводит к пролиферации и расширению гемопоэтических стволовых клеток, необходимых для образования эритроцитов. [ 69 ]
Биохимия раковых стволовых клеток немного отличается от биохимии других опухолевых клеток. Эти так называемые Wnt-зависимые клетки захватывают и зависят от постоянной стимуляции пути Wnt, способствуя их неконтролируемому росту, выживанию и миграции. При раке передача сигналов Wnt может стать независимой от обычных стимулов за счет мутаций в нижестоящих онкогенах и генах-супрессорах опухолей, которые становятся постоянно активированными, даже если нормальный рецептор не получил сигнала. β-катенин связывается с факторами транскрипции, такими как белок TCF4 , и в сочетании молекулы активируют необходимые гены. LF3 сильно ингибирует это связывание in vitro, в клеточных линиях и снижает рост опухоли на мышиных моделях. Это предотвратило репликацию и снизило их способность к миграции, не затрагивая при этом здоровые клетки. После лечения раковых стволовых клеток не осталось. Открытие стало результатом « рациональной разработки лекарств » с использованием технологий AlphaScreens и ELISA. [ 70 ]
Миграция клеток
[ редактировать ]Миграция клеток во время эмбрионального развития позволяет устанавливать оси тела, формировать ткани, индукцию конечностей и некоторые другие процессы. Передача сигналов Wnt помогает опосредовать этот процесс, особенно во время конвергентного расширения. Передача сигналов как от пути Wnt PCP, так и от канонического пути Wnt необходима для правильного конвергентного расширения во время гаструляции. Конвергентное расширение дополнительно регулируется путем Wnt/кальций, который блокирует конвергентное расширение при активации. Передача сигналов Wnt также индуцирует миграцию клеток на более поздних стадиях развития посредством контроля миграционного поведения нейробластов , нервного гребня клеток , миоцитов и клеток трахеи. [ 71 ]
Передача сигналов Wnt участвует в другом ключевом процессе миграции, известном как эпителиально-мезенхимальный переход (EMT). Этот процесс позволяет эпителиальным клеткам трансформироваться в мезенхимальные клетки, так что они больше не удерживаются на месте в ламинине . Он включает в себя подавление кадгерина, благодаря чему клетки могут отделяться от ламинина и мигрировать. Передача сигналов Wnt является индуктором ЕМТ, особенно при развитии молочных желез. [ 72 ]
Чувствительность к инсулину
[ редактировать ]
Инсулин — это пептидный гормон, участвующий в глюкозы гомеостазе в некоторых организмах. В частности, это приводит к активации транспортеров глюкозы в клеточной мембране с целью увеличения поглощения глюкозы из кровотока . Этот процесс частично опосредован активацией передачи сигналов Wnt/β-катенина, которая может повышать чувствительность клеток к инсулину. В частности, Wnt10b представляет собой белок Wnt, который повышает чувствительность клеток скелетных мышц. [ 73 ]
Клинические последствия
[ редактировать ]Рак
[ редактировать ]С момента своего первоначального открытия передача сигналов Wnt была связана с раком . Когда был открыт Wnt1, он был впервые идентифицирован как протоонкоген на мышиной модели рака молочной железы. Тот факт, что Wnt1 является гомологом Wg, показывает, что он участвует в эмбриональном развитии, которое часто требует быстрого деления и миграции клеток. Неправильная регуляция этих процессов может привести к развитию опухоли за счет избыточной пролиферации клеток. [ 3 ]
Активность канонического пути Wnt участвует в развитии доброкачественных и злокачественных опухолей молочной железы. Роль пути Wnt в химиорезистентности опухоли также хорошо документирована, а также его роль в поддержании отдельной субпопуляции клеток, инициирующих рак. [ 74 ] Его присутствие выявляют по повышенным уровням β-катенина в ядре и/или цитоплазме, что можно обнаружить с помощью иммуногистохимического окрашивания и вестерн-блоттинга . Повышенная экспрессия β-катенина коррелирует с плохим прогнозом у пациентов с раком молочной железы. Это накопление может быть связано с такими факторами, как мутации в β-катенине , дефицит комплекса разрушения β-катенина, чаще всего из-за мутаций в структурно нарушенных участках APC , сверхэкспрессия лигандов Wnt, потеря ингибиторов и/или снижение активности регуляторных пути (такие как путь Wnt/кальций). [ 52 ] [ 75 ] [ 76 ] Опухоли молочной железы могут метастазировать из-за участия Wnt в ЕМТ. Исследования метастазирования базальноподобного рака молочной железы в легкие показали, что репрессия передачи сигналов Wnt/β-катенина может предотвратить ЕМТ, которая может ингибировать метастазирование. [ 77 ]
Передача сигналов Wnt вовлечена в развитие других видов рака, а также в десмоидный фиброматоз . [ 78 ] Изменения в экспрессии CTNNB1 , гена, кодирующего β-катенин, можно измерить при раке молочной железы, колоректальном раке , меланоме , предстательной железе , раке легких и других видах рака. Повышенная экспрессия белков-лигандов Wnt, таких как Wnt1, Wnt2 и Wnt7A, наблюдалась при развитии глиобластомы , рака пищевода и рака яичников соответственно. Другие белки, которые вызывают несколько типов рака при отсутствии правильного функционирования, включают ROR1, ROR2, SFRP4 , Wnt5A, WIF1 и белки семейства TCF/LEF. [ 79 ] Передача сигналов Wnt также участвует в патогенезе костных метастазов рака молочной железы и простаты, при этом исследования предполагают дискретные состояния включения и выключения. Wnt подавляется на стадии покоя аутокринным DKK1 , чтобы избежать иммунного надзора. [ 80 ] а также на стадиях диссеминации внутриклеточным Dact1. [ 81 ] Между тем, Wnt активируется во время ранней фазы роста с помощью E-селектина . [ 82 ]
Связь между PGE2 и Wnt предполагает, что связанное с хроническим воспалением увеличение PGE2 может привести к активации пути Wnt в различных тканях, что приводит к канцерогенезу . [ 5 ]
Диабет второго типа
[ редактировать ]Сахарный диабет 2 типа — распространенное заболевание, вызывающее снижение секреции инсулина и повышение инсулинорезистентности на периферии. Это приводит к повышению уровня глюкозы в крови или гипергликемии , которая при отсутствии лечения может привести к летальному исходу. Поскольку передача сигналов Wnt участвует в чувствительности к инсулину, это может быть связано с нарушением работы его пути. Например, сверхэкспрессия Wnt5b может повысить восприимчивость из-за его роли в адипогенезе , поскольку ожирение и диабет II типа имеют высокую коморбидность . [ 83 ] Передача сигналов Wnt является сильным активатором биогенеза митохондрий . Это приводит к увеличению производства активных форм кислорода (АФК), которые, как известно, вызывают повреждение ДНК и клеток. [ 84 ] Это повреждение, вызванное АФК, является значительным, поскольку оно может вызвать острую печеночную резистентность к инсулину или резистентность к инсулину, вызванную повреждением. [ 85 ] Мутации в транскрипционных факторах, связанных с передачей сигналов Wnt, таких как TCF7L2 , связаны с повышенной восприимчивостью. [ 86 ]
См. также
[ редактировать ]- АКСИН1
- ГСК-3
- Управление выпадением волос
- Бескрылый элемент локализации 3 (WLE3)
- Белок 1 индуцируемого сигнального пути WNT1 (WISP1)
- Белок 2 индуцируемого сигнального пути WNT1 (WISP2)
- Белок 3 индуцируемого сигнального пути WNT1 (WISP3)
Ссылки
[ редактировать ]- ^ Нусс Р., Браун А., Папкофф Дж., Скамблер П., Шеклфорд Г., МакМахон А. и др. (январь 1991 г.). «Новая номенклатура int-1 и родственных генов: семейство генов Wnt» . Клетка . 64 (2): 231. doi : 10.1016/0092-8674(91)90633-a . ПМИД 1846319 . S2CID 3189574 .
- ^ Перейти обратно: а б Нусс Р., Вармус Х.Э. (июнь 1992 г.). «Wnt-гены». Клетка . 69 (7): 1073–87. дои : 10.1016/0092-8674(92)90630-У . ПМИД 1617723 . S2CID 10422968 .
- ^ Перейти обратно: а б с д и ж Нусс Р. (январь 2005 г.). «Передача сигналов Wnt при заболевании и развитии» . Клеточные исследования . 15 (1): 28–32. дои : 10.1038/sj.cr.7290260 . ПМИД 15686623 .
- ^ Чжан Х, Чжан Х, Чжан Ю, Нг СС, Жэнь Ф, Ван Ю, Дуань Ю, Чен Л, Чжай Ю, Го Ц, Чанг Цзы (ноябрь 2010 г.). «Белок, взаимодействующий с доменом Dishavelled-DEP (DDIP), ингибирует передачу сигналов Wnt, способствуя деградации TCF4 и разрушая комплекс TCF4/бета-катенин». Сотовая сигнализация . 22 (11): 1753–60. doi : 10.1016/j.cellsig.2010.06.016 . ПМИД 20603214 .
- ^ Перейти обратно: а б с Гесслинг В., Норт Т.Э., Лоуэр С., Лорд А.М., Ли С., Стоик-Купер С.Л., Вайдингер Г., Пудер М., Дейли Г.К., Мун Р.Т., Зон Л.И. (март 2009 г.). «Генетическое взаимодействие передачи сигналов PGE2 и Wnt регулирует спецификацию развития стволовых клеток и регенерацию» . Клетка . 136 (6): 1136–47. дои : 10.1016/j.cell.2009.01.015 . ПМК 2692708 . ПМИД 19303855 .
- ^ Логан С.А., Нусс Р. (2004). «Сигнальный путь Wnt в развитии и заболеваниях». Ежегодный обзор клеточной биологии и биологии развития . 20 : 781–810. CiteSeerX 10.1.1.322.311 . doi : 10.1146/annurev.cellbio.20.010403.113126 . ПМИД 15473860 .
- ^ Перейти обратно: а б с д и ж г час я дж Комия Ю., Бинс Р. (апрель 2008 г.). «Пути передачи сигнала Wnt» . Органогенез . 4 (2): 68–75. дои : 10.4161/org.4.2.5851 . ПМК 2634250 . ПМИД 19279717 .
- ^ Циммерли Д., Хаусманн Г., Канту К., Баслер К. (декабрь 2017 г.). «Фармакологические вмешательства в путь Wnt: ингибирование секреции Wnt по сравнению с нарушением межбелковых интерфейсов ядерных факторов» . Британский журнал фармакологии . 174 (24): 4600–4610. дои : 10.1111/bph.13864 . ПМЦ 5727313 . ПМИД 28521071 .
- ^ Нусс Р., ван Оойен А., Кокс Д., Фунг Ю.К., Вармус Х. (1984). «Режим провирусной активации предполагаемого онкогена молочной железы (int-1) на 15 хромосоме мыши». Природа . 307 (5947): 131–6. Бибкод : 1984Natur.307..131N . дои : 10.1038/307131a0 . ПМИД 6318122 . S2CID 4261052 .
- ^ Клаус А., Бирчмайер В. (май 2008 г.). «Передача сигналов Wnt и ее влияние на развитие и рак». Обзоры природы. Рак . 8 (5): 387–98. дои : 10.1038/nrc2389 . ПМИД 18432252 . S2CID 31382024 .
- ^ Кэдиган К.М., Нусс Р. (декабрь 1997 г.). «Передача сигналов Wnt: общая тема в развитии животных» . Гены и развитие . 11 (24): 3286–305. дои : 10.1101/gad.11.24.3286 . ПМИД 9407023 .
- ^ Ханнуш Р.Н. (октябрь 2015 г.). «Синтетическое белковое липидирование». Современное мнение в области химической биологии . 28 : 39–46. дои : 10.1016/j.cbpa.2015.05.025 . ПМИД 26080277 .
- ^ Ю Дж., Чиа Дж., Каннинг К.А., Джонс К.М., Бард Ф.А., Виршуп Д.М. (май 2014 г.). «Ретроградный транспорт WLS в эндоплазматический ретикулум во время секреции Wnt» . Развивающая клетка . 29 (3): 277–91. дои : 10.1016/j.devcel.2014.03.016 . ПМИД 24768165 .
- ^ Янда С.А., Вагрей Д., Левин А.М., Томас С., Гарсия К.К. (июль 2012 г.). «Структурная основа распознавания Wnt Frizzled» . Наука . 337 (6090): 59–64. Бибкод : 2012Sci...337...59J . дои : 10.1126/science.1222879 . ПМЦ 3577348 . ПМИД 22653731 .
- ^ Хоссейни В., Дани С., Геранмайе М.Х., Мохаммадзаде Ф., Назари Солтан Ахмад С., Дараби М. (июнь 2019 г.). «Липидирование Wnt: роль в транспортировке, модуляции и функции». Журнал клеточной физиологии . 234 (6): 8040–8054. дои : 10.1002/jcp.27570 . ПМИД 30341908 . S2CID 53009014 .
- ^ Кураёси М., Ямамото Х., Изуми С., Кикучи А. (март 2007 г.). «Посттрансляционное пальмитоилирование и гликозилирование Wnt-5a необходимы для передачи сигналов» . Биохимический журнал . 402 (3): 515–23. дои : 10.1042/BJ20061476 . ПМЦ 1863570 . ПМИД 17117926 .
- ^ Нусс, Рул. «Домашняя страница WNT» . Проверено 15 апреля 2013 г.
- ^ Сава Х, Korswagen HC (март 2013 г.). «Сигнализация WNT в C. Elegans» . Червячная книга : 1–30. дои : 10.1895/wormbook.1.7.2 . ПМК 5402212 . ПМИД 25263666 .
- ^ Перейти обратно: а б с Рао Т.П., Кюль М. (июнь 2010 г.). «Обновленный обзор сигнальных путей Wnt: прелюдия к большему» . Исследование кровообращения . 106 (12): 1798–806. дои : 10.1161/CIRCRESAHA.110.219840 . ПМИД 20576942 .
- ^ Шульте Г., Брия В. (октябрь 2007 г.). «Семейство Frizzled нетрадиционных рецепторов, связанных с G-белком». Тенденции в фармакологических науках . 28 (10): 518–25. дои : 10.1016/j.tips.2007.09.001 . ПМИД 17884187 .
- ^ Хабас Р., Давид И.Б. (февраль 2005 г.). «Растрепанная и Wnt-сигнализация: является ли ядро последней границей?» . Журнал биологии . 4 (1): 2. doi : 10.1186/jbiol22 . ПМК 551522 . ПМИД 15720723 .
- ^ Минде Д.П., Анвариан З., Рюдигер С.Г., Морис М.М. (август 2011 г.). «Беспорядок: как миссенс-мутации в белке-супрессоре опухолей APC приводят к раку?» . Молекулярный рак . 10 :101. дои : 10.1186/1476-4598-10-101 . ПМК 3170638 . ПМИД 21859464 .
- ^ Минде Д.П., Радли М., Форнерис Ф., Морис М.М., Рюдигер С.Г. (2013). Пряжка AM (ред.). «Большая степень нарушений при аденоматозном полипозе Coli предлагает стратегию защиты передачи сигналов Wnt от точечных мутаций» . ПЛОС ОДИН . 8 (10): е77257. Бибкод : 2013PLoSO...877257M . дои : 10.1371/journal.pone.0077257 . ПМЦ 3793970 . ПМИД 24130866 .
- ^ Перейти обратно: а б с д Макдональд Б.Т., Тамай К., Хе Х (июль 2009 г.). «Передача сигналов Wnt/бета-катенина: компоненты, механизмы и заболевания» . Развивающая клетка . 17 (1): 9–26. дои : 10.1016/j.devcel.2009.06.016 . ПМЦ 2861485 . ПМИД 19619488 .
- ^ Стаал Ф.Дж., Клеверс Х. (февраль 2000 г.). «Факторы транскрипции Tcf/Lef во время развития Т-клеток: уникальные и перекрывающиеся функции». Гематологический журнал . 1 (1): 3–6. дои : 10.1038/sj.thj.6200001 . ПМИД 11920163 .
- ^ Крампс Т., Питер О., Бруннер Э., Неллен Д., Фрош Б., Чаттерджи С., Муроне М., Цюллиг С., Баслер К. (апрель 2002 г.). «Передача сигналов Wnt/wingless требует BCL9/безногого рекрутирования пигопуса в ядерный комплекс бета-катенин-TCF» (PDF) . Клетка . 109 (1): 47–60. дои : 10.1016/s0092-8674(02)00679-7 . ПМИД 11955446 . S2CID 16720801 . Архивировано из оригинала (PDF) 26 сентября 2021 г. Проверено 3 июня 2020 г.
- ^ Мосиманн С., Хаусманн Г., Баслер К. (апрель 2006 г.). «Парафибромин/Hyrax активирует транскрипцию целевого гена Wnt/Wg путем прямой ассоциации с бета-катенином/Armadillo» . Клетка . 125 (2): 327–41. дои : 10.1016/j.cell.2006.01.053 . ПМИД 16630820 .
- ^ ван Тинен Л.М., Мещанек Дж., Фидлер М., Резерфорд Т.Дж., Биенц М. (март 2017 г.). «Конститутивный каркас из нескольких компонентов энхансосом Wnt с помощью Legless/BCL9» . электронная жизнь . 6 : е20882. дои : 10.7554/elife.20882 . ПМЦ 5352222 . ПМИД 28296634 .
- ^ Седерхольм, Саймон; Канту, Клаудио (21 октября 2020 г.). «Зависимая транскрипция WNT/β-катенин: тканеспецифический процесс» . WIREs Системная биология и медицина . 13 (3): e1511. дои : 10.1002/wsbm.1511 . ПМЦ 9285942 . ПМИД 33085215 .
- ^ Фанг Д., Хоук Д., Чжэн Ю., Ся Ю., Мейзенхельдер Дж., Ника Х., Миллс ГБ, Кобаяши Р., Хантер Т., Лу З. (апрель 2007 г.). «Фосфорилирование бета-катенина с помощью АКТ способствует транскрипционной активности бета-катенина» . Журнал биологической химии . 282 (15): 11221–9. дои : 10.1074/jbc.M611871200 . ПМК 1850976 . ПМИД 17287208 .
- ^ Канту С, Валента Т, Хаусманн Г, Вилен Н, Аге М, Баслер К (июнь 2013 г.). «Взаимодействие Pygo2-H3K4me2/3 необходимо для развития мышей и транскрипции, зависимой от передачи сигналов Wnt» . Разработка . 140 (11): 2377–86. дои : 10.1242/dev.093591 . ПМИД 23637336 .
- ^ Канту С, Циммерли Д, Хаусманн Г, Валента Т, Мур А, Аге М, Баслер К (сентябрь 2014 г.). «Pax6-зависимая, но β-катенин-независимая функция белков Bcl9 в развитии хрусталика мыши» . Гены и развитие . 28 (17): 1879–84. дои : 10.1101/gad.246140.114 . ПМК 4197948 . ПМИД 25184676 .
- ^ Канту С, Пагелла П, Шаджии ТД, Циммерли Д, Валента Т, Хаусманн Г, Баслер К, Мициадис Т.А. (февраль 2017 г.). «Цитоплазматическая роль транскрипционных кофакторов Wnt/β-катенина Bcl9, Bcl9l и Pygopus в формировании зубной эмали» . Научная сигнализация . 10 (465): дааа4 дои : 10.1126/scisignal.aah4598 . ПМИД 28174279 . S2CID 6845295 .
- ^ Гордон, доктор медицинских наук, Нусс Р. (август 2006 г.). «Передача сигналов Wnt: множественные пути, множественные рецепторы и множественные факторы транскрипции» . Журнал биологической химии . 281 (32): 22429–33. дои : 10.1074/jbc.R600015200 . ПМИД 16793760 .
- ^ Сугимура Р., Ли Л (декабрь 2010 г.). «Неканоническая передача сигналов Wnt в развитии позвоночных, стволовых клетках и заболеваниях». Исследование врожденных дефектов. Часть C, Эмбрион сегодня . 90 (4): 243–56. дои : 10.1002/bdrc.20195 . ПМИД 21181886 .
- ^ Перейти обратно: а б с ван Амеронген Р., Нусс Р. (октябрь 2009 г.). «На пути к комплексному взгляду на передачу сигналов Wnt в разработке». Разработка . 136 (19): 3205–14. дои : 10.1242/dev.033910 . ПМИД 19736321 . S2CID 16120512 .
- ^ ван Амеронген Р., Фюрер С., Мизутани М., Нусс Р. (сентябрь 2012 г.). «Wnt5a может как активировать, так и подавлять передачу сигналов Wnt/β-катенина во время эмбрионального развития мыши» . Биология развития . 369 (1): 101–14. дои : 10.1016/j.ydbio.2012.06.020 . ПМЦ 3435145 . ПМИД 22771246 .
- ^ Трасивулу С., Миллар М., Ахмед А. (декабрь 2013 г.). «Активация внутриклеточного кальция множественными лигандами Wnt и транслокация β-катенина в ядро: конвергентная модель путей Wnt/Ca2+ и Wnt/β-катенин» . Журнал биологической химии . 288 (50): 35651–9. дои : 10.1074/jbc.M112.437913 . ПМЦ 3861617 . ПМИД 24158438 .
- ^ Иноки К., Оуян Х., Чжу Т., Линдвалл С., Ван Й., Чжан Х., Ян К., Беннетт С., Харада Й., Станкунас К., Ван С.И., Хе Х, Макдугалд О.А., Ю М., Уильямс БО, Гуань К.Л. (сентябрь 2006 г.) ). «TSC2 объединяет Wnt и энергетические сигналы посредством скоординированного фосфорилирования с помощью AMPK и GSK3 для регулирования роста клеток» . Клетка . 126 (5): 955–68. дои : 10.1016/j.cell.2006.06.055 . ПМИД 16959574 . S2CID 16047397 .
- ^ Курода К., Куанг С., Такето М.М., Рудницки М.А. (март 2013 г.). «Каноническая передача сигналов Wnt индуцирует BMP-4, определяющий медленный миофиброгенез миобластов плода» . Скелетная мышца . 3 (1): 5. дои : 10.1186/2044-5040-3-5 . ПМК 3602004 . ПМИД 23497616 .
- ^ Малинаускас Т., Джонс Э.Ю. (декабрь 2014 г.). «Внеклеточные модуляторы передачи сигналов Wnt» . Современное мнение в области структурной биологии . 29 : 77–84. дои : 10.1016/j.sbi.2014.10.003 . ПМИД 25460271 .
- ^ Гао В., Ким Х., Фэн М., Фунг Й., Ксавье С.П., Рубин Дж.С., Хо М. (август 2014 г.). «Инактивация передачи сигналов Wnt человеческим антителом, которое распознает гепарансульфатные цепи глипикана-3, для терапии рака печени» . Гепатология . 60 (2): 576–87. дои : 10.1002/hep.26996 . ПМК 4083010 . ПМИД 24492943 .
- ^ Гао В, Сюй Ю, Лю Дж, Хо М (май 2016 г.). «Картирование эпитопов с помощью Wnt-блокирующего антитела: свидетельства наличия Wnt-связывающего домена в гепарансульфате» . Научные отчеты . 6 : 26245. Бибкод : 2016NatSR...626245G . дои : 10.1038/srep26245 . ПМЦ 4869111 . ПМИД 27185050 .
- ^ Гао В., Тан З., Чжан Ю.Ф., Фэн М., Цянь М., Димитров Д.С., Хо М. (март 2015 г.). «Иммунотоксин, нацеленный на глипикан-3, регрессирует рак печени посредством двойного ингибирования передачи сигналов Wnt и синтеза белка» . Природные коммуникации . 6 : 6536. Бибкод : 2015NatCo...6.6536G . дои : 10.1038/ncomms7536 . ПМЦ 4357278 . ПМИД 25758784 .
- ^ Перейти обратно: а б с Ли Н, Вэй Л, Лю Х, Бай Х, Йе Ю, Ли Д и др. (апрель 2019 г.). «Вьющийся, богатый цистеином домен в глипикане-3 опосредует связывание Wnt и регулирует рост опухоли гепатоцеллюлярной карциномы у мышей» . Гепатология . 70 (4): 1231–1245. дои : 10.1002/hep.30646 . ПМК 6783318 . ПМИД 30963603 .
- ^ Хо М, Ким Х (февраль 2011 г.). «Глипикан-3: новая мишень для иммунотерапии рака» . Европейский журнал рака . 47 (3): 333–8. дои : 10.1016/j.ejca.2010.10.024 . ПМК 3031711 . ПМИД 21112773 .
- ^ Ли Н, Гао В, Чжан Ю. Ф., Хо М (ноябрь 2018 г.). «Глипиканы как мишени для лечения рака» . Тенденции рака . 4 (11): 741–754. дои : 10.1016/j.trecan.2018.09.004 . ПМК 6209326 . ПМИД 30352677 .
- ^ Гао, Вэй; Сюй, Юнмей; Лю, Цзянь; Хо, Митчелл (17 мая 2016 г.). «Картирование эпитопов с помощью Wnt-блокирующего антитела: свидетельства наличия Wnt-связывающего домена в гепарансульфате» . Научные отчеты . 6 : 26245. Бибкод : 2016NatSR...626245G . дои : 10.1038/srep26245 . ISSN 2045-2322 . ПМЦ 4869111 . ПМИД 27185050 .
- ^ Коллури А, Хо М (2 августа 2019 г.). «Роль глипикана-3 в регуляции Wnt, YAP и Hedgehog при раке печени» . Границы онкологии . 9 : 708. doi : 10.3389/fonc.2019.00708 . ПМК 6688162 . ПМИД 31428581 .
- ^ Малинаускас Т., Арическу А.Р., Лу В., Зибольд С., Джонс Э.Ю. (июль 2011 г.). «Модульный механизм ингибирования передачи сигналов Wnt ингибирующим фактором 1 Wnt» . Структурная и молекулярная биология природы . 18 (8): 886–93. дои : 10.1038/nsmb.2081 . ПМК 3430870 . ПМИД 21743455 .
- ^ Малинаускас Т. (март 2008 г.). «Стыковка жирных кислот с доменом WIF человеческого фактора-1, ингибирующего Wnt». Липиды . 43 (3): 227–30. дои : 10.1007/s11745-007-3144-3 . ПМИД 18256869 . S2CID 31357937 .
- ^ Перейти обратно: а б Минде Д.П., Радли М., Форнерис Ф., Морис М.М., Рюдигер С.Г. (2013). «Большая степень нарушений при аденоматозном полипозе Coli предлагает стратегию защиты передачи сигналов Wnt от точечных мутаций» . ПЛОС ОДИН . 8 (10): е77257. Бибкод : 2013PLoSO...877257M . дои : 10.1371/journal.pone.0077257 . ПМЦ 3793970 . ПМИД 24130866 .
- ^ Перейти обратно: а б с д Гилберт С.Ф. (2010). Биология развития (9-е изд.). Сандерленд, Массачусетс: Sinauer Associates. ISBN 9780878933846 .
- ^ Васиев Б., Балтер А., Капеллан М., Глейзер Дж.А., Вейер С.Дж. (май 2010 г.). «Моделирование гаструляции у куриного эмбриона: формирование первичной полоски» . ПЛОС ОДИН . 5 (5): е10571. Бибкод : 2010PLoSO...510571V . дои : 10.1371/journal.pone.0010571 . ПМК 2868022 . ПМИД 20485500 .
- ^ Гилберт С.Ф. (2014). «Раннее развитие птиц». Биология развития (10-е изд.). Сандерленд (Массачусетс): Sinauer Associates.
- ^ Уллоа Ф, Марти Э (январь 2010 г.). «Wnt выиграл войну: антагонистическая роль Wnt над Shh контролирует дорсо-вентральное паттернирование нервной трубки позвоночных» . Динамика развития . 239 (1): 69–76. дои : 10.1002/dvdy.22058 . ПМИД 19681160 .
- ^ Цзоу Ю (сентябрь 2004 г.). «Передача сигналов Wnt в наведении аксонов». Тенденции в нейронауках . 27 (9): 528–32. doi : 10.1016/j.tins.2004.06.015 . ПМИД 15331234 . S2CID 15635026 .
- ^ Гордон Н.К., Гордон Р. (март 2016 г.). «Органелла дифференцировки эмбрионов: расщепитель клеточного состояния» . Теоретическая биология и медицинское моделирование . 13:11 . дои : 10.1186/s12976-016-0037-2 . ПМЦ 4785624 . ПМИД 26965444 .
- ^ Гордон Н., Гордон Р. (2016). Объяснение эмбриогенеза . Сингапур: Всемирное научное издательство . стр. 580–591. дои : 10.1142/8152 . ISBN 978-981-4740-69-2 .
- ^ Перейти обратно: а б Нусс Р. (май 2008 г.). «Передача сигналов Wnt и контроль стволовых клеток» . Клеточные исследования . 18 (5): 523–7. дои : 10.1038/cr.2008.47 . ПМИД 18392048 .
- ^ Бакре М.М., Хой А., Монг Дж.К., Ко Й.И., Вонг К.Ю., Стэнтон Л.В. (октябрь 2007 г.). «Поколение мультипотентных мезендодермальных предшественников из эмбриональных стволовых клеток мыши посредством устойчивой активации пути Wnt» . Журнал биологической химии . 282 (43): 31703–12. дои : 10.1074/jbc.M704287200 . ПМИД 17711862 .
- ^ Уолл П.С., Моррис Дж.К., Пейншаб М.С., Маркус Р.К., Кон А.Д., Бихеле Т.Л., Мун Р.Т., Кауфман Д.С. (январь 2008 г.). «Передача сигналов Wnt способствует развитию гематоэндотелиальных клеток из эмбриональных стволовых клеток человека» . Кровь . 111 (1): 122–31. дои : 10.1182/blood-2007-04-084186 . ПМК 2200802 . ПМИД 17875805 .
- ^ Шнайдер В.А., Меркола М. (февраль 2001 г.). «Антагонизм Wnt инициирует кардиогенез у Xenopus laevis» . Гены и развитие . 15 (3): 304–15. дои : 10.1101/gad.855601 . ПМК 312618 . ПМИД 11159911 .
- ^ Марвин М.Дж., Ди Рокко Дж., Гардинер А., Буш С.М., Лассар А.Б. (февраль 2001 г.). «Ингибирование активности Wnt индуцирует образование сердца из задней мезодермы» . Гены и развитие . 15 (3): 316–27. дои : 10.1101/gad.855501 . ПМК 312622 . ПМИД 11159912 .
- ^ Уэно С., Вайдингер Г., Осуги Т., Кон А.Д., Голоб Дж.Л., Пабон Л., Райнеке Х., Мун Р.Т., Мерри CE (июнь 2007 г.). «Двухфазная роль передачи сигналов Wnt/бета-катенина в спецификации сердца у рыбок данио и эмбриональных стволовых клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (23): 9685–90. Бибкод : 2007PNAS..104.9685U . дои : 10.1073/pnas.0702859104 . ПМК 1876428 . ПМИД 17522258 .
- ^ Виллемс Э., Спиринг С., Давидович Х., Ланье М., Ся З., Доусон М., Кэшман Дж., Меркола М. (август 2011 г.). «Низкомолекулярные ингибиторы пути Wnt мощно стимулируют кардиомиоциты из мезодермы, полученной из эмбриональных стволовых клеток человека» . Исследование кровообращения . 109 (4): 360–4. дои : 10.1161/CIRCRESAHA.111.249540 . ПМЦ 3327303 . ПМИД 21737789 .
- ^ Берридж П.В., Матса Э., Шукла П., Лин З.К., Чурко Дж.М., Эберт А.Д., Лан Ф., Дике С., Хубер Б., Мордвинкин Н.М., Плевс Дж.Р., Абилез О.Дж., Цуй Б., Голд Дж.Д., Ву Дж.К. (август 2014 г.). «Химически определяемая генерация кардиомиоцитов человека» . Природные методы . 11 (8): 855–60. дои : 10.1038/nmeth.2999 . ПМК 4169698 . ПМИД 24930130 .
- ^ Калдис П., Пагано М. (декабрь 2009 г.). «Передача сигналов Wnt в митозе» . Развивающая клетка . 17 (6): 749–50. дои : 10.1016/j.devcel.2009.12.001 . ПМИД 20059944 .
- ^ Виллерт К., Джонс К.А. (июнь 2006 г.). «Сигнализация Wnt: партия в ядре?» . Гены и развитие . 20 (11): 1394–404. дои : 10.1101/gad.1424006 . ПМИД 16751178 .
- ^ Ходж, Расс (25 января 2016 г.). «Взлом программ раковых стволовых клеток» . www.medicalxpress.com . Медицинский экспресс . Проверено 12 февраля 2016 г.
- ^ Шамбони А., Ведлих Д. (2013). Передача сигналов Wnt и миграция клеток . Ландес Бионауки . Проверено 7 мая 2013 г.
{{cite book}}:|website=игнорируется ( помогите ) - ^ Микалицци Д.С., Фарабо С.М., Форд Х.Л. (июнь 2010 г.). «Эпителиально-мезенхимальный переход при раке: параллели между нормальным развитием и прогрессированием опухоли» . Журнал биологии молочной железы и неоплазии . 15 (2): 117–34. дои : 10.1007/s10911-010-9178-9 . ПМК 2886089 . ПМИД 20490631 .
- ^ Абиола М., Фавье М., Христодулу-Вафейаду Э., Пишар А.Л., Мартелли И., Гийе-Деньо I (декабрь 2009 г.). «Активация передачи сигналов Wnt/бета-катенин повышает чувствительность к инсулину посредством взаимной регуляции Wnt10b и SREBP-1c в клетках скелетных мышц» . ПЛОС ОДИН . 4 (12): е8509. Бибкод : 2009PLoSO...4.8509A . дои : 10.1371/journal.pone.0008509 . ПМК 2794543 . ПМИД 20041157 .
- ^ Милошевич, В. и др. Аутокринные схемы Wnt/IL-1β/IL-8 контролируют химиорезистентность в клетках, инициирующих мезотелиому, путем индукции ABCB5.Int. Дж. Рак, https://doi.org/10.1002/ijc.32419
- ^ Хоу Л.Р., Браун А.М. (январь 2004 г.). «Передача сигналов Wnt и рак молочной железы» . Биология и терапия рака . 3 (1): 36–41. дои : 10.4161/cbt.3.1.561 . ПМИД 14739782 .
- ^ Такето М.М. (апрель 2004 г.). «Отключение рака, активируемого сигналом Wnt» . Природная генетика . 36 (4): 320–2. дои : 10.1038/ng0404-320 . ПМИД 15054482 .
- ^ ДиМео Т.А., Андерсон К., Фадке П., Фан С., Фэн С., Перу С.М., Набер С., Купервассер С. (июль 2009 г.). «Новая характеристика метастазирования в легкие связывает передачу сигналов Wnt с самообновлением раковых клеток и эпителиально-мезенхимальным переходом при базальноподобном раке молочной железы» . Исследования рака . 69 (13): 5364–73. дои : 10.1158/0008-5472.CAN-08-4135 . ПМЦ 2782448 . ПМИД 19549913 .
- ^ Ховард, Дж. Харрисон; Поллок, Рафаэль Э. (июнь 2016 г.). «Интраабдоминальный и десмоидный фиброматоз брюшной стенки» . Онкология и терапия . 4 (1): 57–72. дои : 10.1007/s40487-016-0017-z . ISSN 2366-1070 . ПМК 5315078 . ПМИД 28261640 .
- ^ Анастас Дж.Н., Moon RT (январь 2013 г.). «Сигнальные пути WNT как терапевтические мишени при раке». Обзоры природы. Рак . 13 (1): 11–26. дои : 10.1038/nrc3419 . ПМИД 23258168 . S2CID 35599667 .
- ^ Маллади, Шринивас; Макалинао, Данило Г.; Цзинь, Синь; Он, Лан; Баснет, Харихар; Цзоу, Илун; де Станчина, Элиза; Массаге, Джоан (24 марта 2016 г.). «Метастатическая латентность и уклонение от иммунитета посредством аутокринного ингибирования WNT» . Клетка . 165 (1): 45–60. дои : 10.1016/j.cell.2016.02.025 . ISSN 1097-4172 . ПМК 4808520 . ПМИД 27015306 .
- ^ Эспозито, Марк; Фанг, Цао; Кук, Кейтлин С.; Пак, Нана; Вэй, Юн; Спадацци, Кьяра; Брача, Дэн; Гунаратна, Рамеш Т.; Лаевский, Гэри; ДеКосте, Кристина Дж.; Слабодкин, Ханна (март 2021 г.). «Биомолекулярные конденсаты DACT1, индуцированные TGF-β, подавляют передачу сигналов Wnt, способствуя метастазированию в кости» . Природная клеточная биология . 23 (3): 257–267. дои : 10.1038/s41556-021-00641-w . ISSN 1476-4679 . ПМЦ 7970447 . ПМИД 33723425 .
- ^ Эспозито, Марк; Мондал, Нандини; Греко, Тодд М.; Вэй, Юн; Спадацци, Кьяра; Линь, Сон-Чанг; Чжэн, Ханьцю; Чунг, Кори; Маньяни, Джон Л.; Лин, Сью-Хва; Кристя, Илеана М. (май 2019 г.). «Е-селектин костно-сосудистой ниши индуцирует мезенхимально-эпителиальный переход и активацию Wnt в раковых клетках, способствуя метастазированию в кости» . Природная клеточная биология . 21 (5): 627–639. дои : 10.1038/s41556-019-0309-2 . ISSN 1476-4679 . ПМК 6556210 . ПМИД 30988423 .
- ^ Welters HJ, Кулкарни Р.Н. (декабрь 2008 г.). «Передача сигналов Wnt: значение для биологии бета-клеток и диабета». Тенденции в эндокринологии и обмене веществ . 19 (10): 349–55. дои : 10.1016/j.tem.2008.08.004 . ПМИД 18926717 . S2CID 19299033 .
- ^ Юн Дж.К., Нг А., Ким Б.Х., Бьянко А., Ксавье Р.Дж., Элледж С.Дж. (июль 2010 г.). «Передача сигналов Wnt регулирует физиологию митохондрий и чувствительность к инсулину» . Гены и развитие . 24 (14): 1507–18. дои : 10.1101/gad.1924910 . ПМК 2904941 . ПМИД 20634317 .
- ^ Чжай Л., Баллинджер С.В., Мессина Дж.Л. (март 2011 г.). «Роль активных форм кислорода в резистентности к инсулину, вызванной травмой» . Молекулярная эндокринология . 25 (3): 492–502. дои : 10.1210/me.2010-0224 . ПМК 3045736 . ПМИД 21239612 .
- ^ Грант С.Ф., Торлейфссон Г., Рейнисдоттир И., Бенедиктссон Р., Манолеску А., Сайнс Дж. и др. (март 2006 г.). «Вариант гена транскрипционного фактора 7, подобного 2 (TCF7L2), повышает риск развития диабета 2 типа». Природная генетика . 38 (3): 320–3. дои : 10.1038/ng1732 . ПМИД 16415884 . S2CID 28825825 .
Дальнейшее чтение
[ редактировать ]- Милошевич В. и др. (январь 2020 г.). «Аутокринные схемы Wnt/IL-1β/IL-8 контролируют химиорезистентность в клетках, инициирующих мезотелиому, путем индукции ABCB5» . Межд. Дж. Рак . 146 (1): 192–207. дои : 10.1002/ijc.32419 . hdl : 2318/1711962 . ПМИД 31107974 . S2CID 160014053 .
- Динасарапу А.Р., Сондерс Б., Озерлат И., Азам К., Субраманиам С. (июнь 2011 г.). «Страницы молекул сигнального шлюза — взгляд на модель данных» . Биоинформатика . 27 (12): 1736–8. doi : 10.1093/биоинформатика/btr190 . ПМК 3106186 . ПМИД 21505029 .
Внешние ссылки
[ редактировать ]- Wnt + Proteins Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
| Базы данных органов управления : Национальные |
|---|