Эффективность клеток
Потенциал клетки – это клетки способность дифференцироваться в другие типы клеток. [1] [2] Чем больше типов клеток может дифференцироваться в клетку, тем выше ее эффективность. Потенциал также описывается как потенциал активации гена внутри клетки, который, как и континуум, начинается с тотипотентности , обозначающей клетку с наибольшим потенциалом дифференциации, плюрипотентностью , мультипотентностью , олигопотентностью и, наконец, унипотентностью .

Тотипотентность
[ редактировать ]Тотипотентность (лат. totipotentia , букв. «способность ко всему [вещам]») — это способность отдельной клетки делиться и производить все дифференцированные клетки организма . Споры и зиготы являются примерами тотипотентных клеток. [3] В спектре клеточной активности тотипотентность представляет собой клетку с наибольшим потенциалом дифференцировки , способную дифференцироваться в любую эмбриональную клетку, а также в любую внеэмбриональную тканевую клетку. Напротив, плюрипотентные клетки могут дифференцироваться только в эмбриональные клетки. [4] [5]
Полностью дифференцированная клетка может вернуться в состояние тотипотентности. [6] Преобразование в тотипотентность сложное и до конца не изученное. В 2011 году исследование показало, что клетки могут дифференцироваться не в полностью тотипотентные клетки, а в «сложную клеточную вариацию» тотипотентности. [7]
Модель развития человека можно использовать для описания того, как возникают тотипотентные клетки. [8] Развитие человека начинается, когда сперматозоид оплодотворяет яйцеклетку, и в результате оплодотворенной яйцеклетки образуется единственная тотипотентная клетка — зигота . [9] В первые часы после оплодотворения эта зигота делится на одинаковые тотипотентные клетки, которые в дальнейшем могут развиться в любой из трех зародышевых листков человека ( энтодерму , мезодерму или эктодерму ) или в клетки плаценты ( цитотрофобласт или синцитиотрофобласт ). Достигнув стадии 16 клеток, тотипотентные клетки морулы дифференцируются в клетки, которые в конечном итоге становятся либо , бластоцисты внутренней клеточной массой либо внешними трофобластами . Примерно через четыре дня после оплодотворения и после нескольких циклов клеточного деления эти тотипотентные клетки начинают специализироваться. Внутренняя клеточная масса, источник эмбриональных стволовых клеток , становится плюрипотентной.
Исследования Caenorhabditis elegans показывают, что множество механизмов, включая регуляцию РНК, могут играть роль в поддержании тотипотентности на разных стадиях развития у некоторых видов. [10] Работа с рыбками данио и млекопитающими предполагает дальнейшее взаимодействие между микроРНК и РНК-связывающими белками (RBPs) в определении различий в развитии. [11]
Первичные половые клетки
[ редактировать ]В первичных зародышевых клетках перепрограммирование , мыши полногеномное ведущее к тотипотентности, включает стирание эпигенетических отпечатков. Перепрограммированию способствует активное деметилирование ДНК ДНК с участием ферментативного пути репарации вырезания оснований . [12] Этот путь влечет за собой стирание метилирования CpG (5mC) в первичных зародышевых клетках посредством первоначального превращения 5mC в 5-гидроксиметилцитозин (5hmC), реакции, обусловленной высокими уровнями ферментов десять-одиннадцать диоксигеназ. ТЭТ-1 и ТЭТ-2 . [13]
Плюрипотентность
[ редактировать ]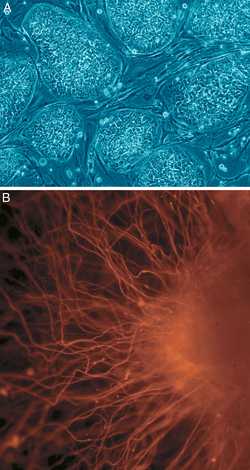
Б: Нервные клетки
В клеточной биологии плюрипотентность (лат. pluripotentia , букв. «способность ко многим [вещам]») [14] относится к стволовым клеткам, которые обладают потенциалом дифференцироваться в любой из трех зародышевых слоев : энтодерму (кишечник, легкие и печень), мезодерму (мышцы, скелет, кровеносные сосуды, мочеполовые органы, дерма) или эктодерму (нервная, сенсорная, эпидермис) . ), но не во внеэмбриональные ткани, такие как плацента или желточный мешок. [15]
Индуцированная плюрипотентность
[ редактировать ]Индуцированные плюрипотентные стволовые клетки, обычно сокращенно называемые iPS-клетками или iPSC, представляют собой тип плюрипотентных стволовых клеток, искусственно полученных из неплюрипотентной клетки, обычно взрослой соматической клетки , путем индуцирования «принудительной» экспрессии определенных генов и факторов транскрипции . [16] Эти факторы транскрипции играют ключевую роль в определении состояния этих клеток, а также подчеркивают тот факт, что эти соматические клетки сохраняют ту же генетическую информацию, что и ранние эмбриональные клетки. [17] Способность переводить клетки в плюрипотентное состояние была впервые открыта в 2006 году с использованием фибробластов мыши и четырех факторов транскрипции: Oct4 , Sox2 , Klf4 и c- Myc ; [18] эта техника, называемая перепрограммированием , позже принесла Шинья Яманаке и Джону Гердону Нобелевскую премию по физиологии и медицине. [19] Затем в 2007 году последовала успешная индукция ИПСК человека, полученных из фибробластов дермы человека, с использованием методов, аналогичных тем, которые используются для индукции клеток мыши. [20] Эти индуцированные клетки обладают свойствами, сходными с таковыми эмбриональных стволовых клеток (ЭСК), но не требуют использования эмбрионов. Некоторые сходства между ЭСК и ИПСК включают плюрипотентность, морфологию , способность к самообновлению (признак, который подразумевает, что они могут делиться и реплицироваться бесконечно), а также экспрессию генов . [21]
Считается также, что эпигенетические факторы участвуют в фактическом перепрограммировании соматических клеток с целью индукции плюрипотентности. Было высказано предположение, что определенные эпигенетические факторы могут фактически работать на очистку исходных соматических эпигенетических меток с целью приобретения новых эпигенетических меток, которые являются частью достижения плюрипотентного состояния. Хроматин также реорганизуется в ИПСК и становится таким же, как в ЭСК, поскольку он менее конденсирован и, следовательно, более доступен. Модификации эухроматина также распространены, что также соответствует состоянию эухроматина , обнаруженного в ЭСК. [21]
Из-за их большого сходства с ЭСК ИПСК интересуют медицинское и исследовательское сообщество. ИПСК потенциально могут иметь те же терапевтические последствия и применения, что и ЭСК, но без спорного использования эмбрионов в этом процессе, что является темой больших биоэтических дебатов. Индуцированная плюрипотентность соматических клеток в недифференцированные iPS-клетки первоначально рассматривалась как конец спорного использования эмбриональных стволовых клеток . Однако было обнаружено, что ИПСК потенциально канцерогенны , и, несмотря на достижения, [16] до недавнего времени никогда не были одобрены для клинических исследований в США. В настоящее время аутологичные дофаминергические клетки-предшественники, полученные из ИПСК, используются в исследованиях по лечению болезни Паркинсона. [22] При создании ИПСК также сталкивались с такими недостатками, как низкая скорость репликации и раннее старение. [23] препятствуя их использованию в качестве замены ЭСК.
Соматическая экспрессия комбинированных факторов транскрипции может напрямую индуцировать судьбу других определенных соматических клеток ( трансдифференцировка ); Исследователи определили три транскрипционных фактора, специфичных для нейронных линий, которые могут напрямую превращать мышиные фибробласты (клетки соединительной ткани) в полностью функциональные нейроны . [24] Этот результат ставит под сомнение терминальную природу клеточной дифференцировки и целостность клонирования; и подразумевает, что при наличии соответствующих инструментов все клетки тотипотентны и могут образовывать любые виды тканей.
Некоторые из возможных медицинских и терапевтических применений ИПСК, полученных от пациентов, включают их использование при трансплантации клеток и тканей без риска отторжения, который обычно встречается. ИПСК потенциально могут заменить непригодные модели на животных, а также модели in vitro, используемые для исследования заболеваний. [25]

Наивные и подготовленные состояния плюрипотентности
[ редактировать ]Результаты, полученные в отношении эпибластов до и после имплантации, позволили классифицировать плюрипотентность на два состояния: «наивное» и «праймированное», представляющее эпибласт до и после имплантации соответственно. [26] Континуум от наивного к праймированному контролируется уменьшением димеризации Sox2/Oct4 на элементах ДНК SoxOct, контролирующих наивную плюрипотентность. [27] Праймированные плюрипотентные стволовые клетки разных видов можно вернуть в наивное состояние с помощью коктейля, содержащего Klf4 и Sox2 или «супер-Sox» — химерного транскрипционного фактора с повышенной способностью димеризоваться с Oct4. [27]
Исходные стволовые клетки, обычно используемые в науке и называемые эмбриональными стволовыми клетками (ЭСК), происходят из предимплантационного эпибласта; такой эпибласт способен генерировать весь плод, а одна клетка эпибласта может вносить вклад во все клеточные линии, если ее инъецировать в другую бластоцисту. С другой стороны, между эпибластами до и после имплантации можно наблюдать несколько заметных различий, таких как разница в морфологии, при которой эпибласт после имплантации меняет свою морфологию на чашеобразную форму, называемую «яйцевым цилиндром», как а также хромосомные изменения, при которых одна из Х-хромосом подвергается случайной инактивации на ранней стадии яйцевого цилиндра, известное как Х-инактивация . [28] Во время этого развития клетки эпибласта яйцевого цилиндра систематически подвергаются воздействию факторов роста фибробластов , передачи сигналов Wnt и других индуктивных факторов через окружающий желточный мешок и ткань трофобласта. [29] так, что они становятся инструктивно конкретными в соответствии с пространственной организацией. [30]
Еще одним важным отличием является то, что постимплантационные стволовые клетки эпибласта не способны вносить вклад в химеры бластоцисты . [31] что отличает их от других известных плюрипотентных стволовых клеток. Клеточные линии, полученные из таких постимплантационных эпибластов, называются стволовыми клетками, полученными из эпибластов , которые впервые были получены в лаборатории в 2007 году. И ESC, и EpiSCs происходят из эпибластов, но находятся на разных фазах развития. Плюрипотентность все еще сохраняется в постимплантационном эпибласте, о чем свидетельствует консервативная экспрессия Nanog , Fut4 и Oct-4 в EpiSCs. [32] до сомитогенеза и может быть обращено вспять в середине индуцированной экспрессии Oct-4 . [33]
Нативная плюрипотентность растений
[ редактировать ]
Неиндуцированная плюрипотентность наблюдалась в культуре ткани корневой меристемы , особенно Каримом и др., 2015 г., Кимом и др., 2018 г. и Росспопоффом и др., 2017 г. Эта плюрипотентность регулируется различными регуляторами, включая PLETHORA 1 и PLETHORA 2 ; и PLETHORA 3 , PLETHORA 5 и PLETHORA 7 , экспрессия которых, как обнаружил Карим, провоцируется ауксином . (Они также известны как PLT1, PLT2, PLT3, PLT5, PLT7 и экспрессируются одноименными генами.) По состоянию на 2019 г. [update]Ожидается, что это откроет возможности для будущих исследований плюрипотентности в тканях корня. [34]
Мультипотентность
[ редактировать ]
Мультипотентность — это когда клетки-предшественники обладают потенциалом активации генов, позволяющим дифференцироваться в отдельные типы клеток. Например, гемопоэтическая стволовая клетка – и этот тип клеток может дифференцироваться в несколько типов клеток крови, таких как лимфоциты , моноциты , нейтрофилы и т. д., но до сих пор неясно, обладают ли ЗСК способностью дифференцироваться в клетки мозга , костные клетки или другие типы клеток, не относящиеся к крови. [ нужна ссылка ]
Исследования, связанные с мультипотентными клетками, показывают, что мультипотентные клетки могут быть способны превращаться в клетки неродственных типов. В другом случае стволовые клетки пуповинной крови человека были преобразованы в человеческие нейроны. [35] Также проводятся исследования по превращению мультипотентных клеток в плюрипотентные клетки. [36]
Мультипотентные клетки обнаружены во многих, но не во всех типах клеток человека. обнаружены мультипотентные клетки В пуповинной крови . [37] жировая ткань, [38] сердечные клетки, [39] костный мозг и мезенхимальные стволовые клетки (МСК), которые обнаруживаются в третьем моляре . [40]
МСК могут оказаться ценным источником стволовых клеток из коренных зубов в возрасте 8–10 лет, до кальцификации зубов у взрослых. МСК могут дифференцироваться в остеобласты, хондроциты и адипоциты. [41]
Олигопотентность
[ редактировать ]В биологии олигопотентность — это способность клеток-предшественников дифференцироваться в несколько типов клеток . Это степень могущества . Примерами олигопотентных стволовых клеток являются лимфоидные или миелоидные стволовые клетки. [2] В частности, лимфоидная клетка может давать начало различным клеткам крови, таким как В- и Т-клетки, но не другому типу клеток крови, например эритроцитам. [42] Примерами клеток-предшественников являются сосудистые стволовые клетки, способные становиться как эндотелиальными , так и гладкомышечными клетками.
Унипотентность
[ редактировать ]В клеточной биологии унипотентная клетка — это концепция, согласно которой одна стволовая клетка способна дифференцироваться только в один тип клеток. [43] В настоящее время неясно, существуют ли настоящие унипотентные стволовые клетки. Гепатобласты, которые дифференцируются в гепатоциты (составляющие большую часть печени ) или холангиоциты (эпителиальные клетки желчных протоков), являются бипотентными. [44] Близким синонимом унипотентной клетки является клетка-предшественник .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Тотипотентность клеток была открыта Хабертландом, а этот термин был придуман Томасом Хандом Морганом. Махла РС (2016). «Применение стволовых клеток в регенеративной медицине и терапии заболеваний» . Международный журнал клеточной биологии . 2016 (7): 6940283. doi : 10.1155/2016/6940283 . ПМЦ 4969512 . ПМИД 27516776 .
- ^ Jump up to: а б Шёлер Х.Р. (2007). «Потенциал стволовых клеток: инвентарь». В Кнопффлер М., Шипански Д., Зоргнер С.Л. (ред.). Биотехнология человека как социальный вызов . Ашгейт Паблишинг, ООО с. 28. ISBN 978-0-7546-5755-2 .
- ^ Миталипов С , Вольф Д (2009). «Тотипотентность, плюрипотентность и ядерное перепрограммирование». Инженерия стволовых клеток . Достижения в области биохимической инженерии/биотехнологии. Том. 114. стр. 185–199. Бибкод : 2009esc..book..185M . дои : 10.1007/10_2008_45 . ISBN 978-3-540-88805-5 . ПМЦ 2752493 . ПМИД 19343304 .
- ^ Лодиш Х (2016). Молекулярно-клеточная биология (8-е изд.). У. Х. Фриман. стр. 975–977. ISBN 978-1319067748 .
- ^ «В чем разница между тотипотентным, плюрипотентным и мультипотентным?» .
- ^ Вестерн П (2009). «Фетальные половые клетки: достижение баланса между плюрипотентностью и дифференцировкой» . Международный журнал биологии развития . 53 (2–3): 393–409. doi : 10.1387/ijdb.082671pw . ПМИД 19412894 .
- ^ Сугимото К., Гордон С.П., Мейеровиц Э.М. (апрель 2011 г.). «Регенерация у растений и животных: дедифференцировка, трансдифференцировка или просто дифференцировка?». Тенденции в клеточной биологии . 21 (4): 212–218. дои : 10.1016/j.tcb.2010.12.004 . ПМИД 21236679 .
- ^ Сейду Дж., Браун Р.Э. (декабрь 2006 г.). «Путь к тотипотентности: уроки половых клеток» . Клетка . 127 (5): 891–904. дои : 10.1016/j.cell.2006.11.016 . ПМИД 17129777 . S2CID 16988032 .
- ^ Аш Р., Симерли С., Орд Т., Орд В.А., Шаттен Г. (июль 1995 г.). «Стадии, на которых прекращается оплодотворение человека: конфигурации микротрубочек и хромосом в оплодотворенных ооцитах, которые не смогли завершить оплодотворение и развитие у человека». Репродукция человека . 10 (7): 1897–1906. doi : 10.1093/oxfordjournals.humrep.a136204 . ПМИД 8583008 .
- ^ Чиоск Р., ДеПальма М., Присс-младший (февраль 2006 г.). «Регуляторы трансляции поддерживают тотипотентность зародышевой линии Caenorhabditis elegans». Наука . 311 (5762): 851–853. Бибкод : 2006Sci...311..851C . дои : 10.1126/science.1122491 . ПМИД 16469927 . S2CID 130017 .
- ^ Кедде М., Агами Р. (апрель 2008 г.). «Взаимодействие между микроРНК и РНК-связывающими белками определяет процессы развития» . Клеточный цикл . 7 (7): 899–903. дои : 10.4161/cc.7.7.5644 . ПМИД 18414021 .
- ^ Хайкова П., Джеффрис С.Дж., Ли С., Миллер Н., Джексон С.П., Сурани М.А. (июль 2010 г.). «Полногеномное перепрограммирование в зародышевой линии мыши влечет за собой путь эксцизионной репарации основания» . Наука . 329 (5987): 78–82. Бибкод : 2010Sci...329...78H . дои : 10.1126/science.1187945 . ПМЦ 3863715 . ПМИД 20595612 .
- ^ Хакетт Дж.А., Сенгупта Р., Зилич Дж.Дж., Мураками К., Ли С., Даун Т.А., Сурани М.А. (январь 2013 г.). «Динамика деметилирования зародышевой ДНК и стирание отпечатка с помощью 5-гидроксиметилцитозина» . Наука . 339 (6118): 448–452. Бибкод : 2013Sci...339..448H . дои : 10.1126/science.1229277 . ПМЦ 3847602 . ПМИД 23223451 .
- ^ «Биология онлайн» . Биология-Online.org . Проверено 25 апреля 2013 г.
- ^ Биндер, М.Д., Хирокава Н., Нобутака, Виндхорст У., ред. (2009). Энциклопедия нейробиологии . Берлин: Шпрингер. ISBN 978-3540237358 .
- ^ Jump up to: а б Бейкер М. (06 декабря 2007 г.). «Взрослые клетки, перепрограммированные на плюрипотентность, без опухолей» . Стволовые клетки Nature Reports : 1. doi : 10.1038/stemcells.2007.124 .
- ^ Штадтфельд М., Хохдлингер К. (октябрь 2010 г.). «Индуцированная плюрипотентность: история, механизмы и приложения» . Гены и развитие . 24 (20): 2239–2263. дои : 10.1101/gad.1963910 . ПМК 2956203 . ПМИД 20952534 .
- ^ Такахаши К., Яманака С. (август 2006 г.). «Индукция плюрипотентных стволовых клеток из культур эмбриональных и взрослых фибробластов мышей с помощью определенных факторов». Клетка . 126 (4): 663–676. дои : 10.1016/j.cell.2006.07.024 . hdl : 2433/159777 . ПМИД 16904174 . S2CID 1565219 .
- ^ « Нобелевская премия по физиологии и медицине 2012 г. ». Нобелевская премия.org. Nobel Media AB 2013. Интернет. 28 ноября 2013 г.
- ^ Такахаши К., Танабэ К., Онуки М., Нарита М., Ичисака Т., Томода К., Яманака С. (ноябрь 2007 г.). «Индукция плюрипотентных стволовых клеток из фибробластов взрослого человека определенными факторами». Клетка . 131 (5): 861–872. дои : 10.1016/j.cell.2007.11.019 . hdl : 2433/49782 . ПМИД 18035408 . S2CID 8531539 .
- ^ Jump up to: а б Лян Г, Чжан И (январь 2013 г.). «Эмбриональные стволовые клетки и индуцированные плюрипотентные стволовые клетки: эпигенетическая перспектива» . Клеточные исследования . 23 (1): 49–69. дои : 10.1038/cr.2012.175 . ПМЦ 3541668 . ПМИД 23247625 .
- ^ Швейцер Дж.С., Сонг Б., Херрингтон Т.М., Парк Т.И., Ли Н., Ко С. и др. (май 2020 г.). «Персонализированные клетки-предшественники дофамина, полученные из ИПСК, для лечения болезни Паркинсона» . Медицинский журнал Новой Англии . 382 (20): 1926–1932. дои : 10.1056/NEJMoa1915872 . ПМЦ 7288982 . ПМИД 32402162 .
- ^ Чой, Чарльз. «Отключение клеток: индуцированные плюрипотентные стволовые клетки не соответствуют потенциалу, обнаруженному в эмбриональной версии» . Научный американец . Проверено 25 апреля 2013 г.
- ^ Вирбухен Т., Остермайер А., Панг З.П., Кокубу Й., Зюдхоф Т.К., Верниг М. (февраль 2010 г.). «Прямое преобразование фибробластов в функциональные нейроны с помощью определенных факторов» . Природа . 463 (7284): 1035–1041. Бибкод : 2010Natur.463.1035V . дои : 10.1038/nature08797 . ПМЦ 2829121 . ПМИД 20107439 .
- ^ Парк И.Х., Леру П.Х., Чжао Р., Хо Х., Дейли GQ (2008). «Получение индуцированных человеком плюрипотентных стволовых клеток». Протоколы природы . 3 (7): 1180–1186. дои : 10.1038/nprot.2008.92 . ПМИД 18600223 . S2CID 13321484 .
- ^ Николс Дж., Смит А. (июнь 2009 г.). «Наивные и подготовленные плюрипотентные государства» . Клеточная стволовая клетка . 4 (6): 487–492. дои : 10.1016/j.stem.2009.05.015 . ПМИД 19497275 .
- ^ Jump up to: а б Маккарти, Кейтлин М.; Ву, Гуанмин; Малик, Викас; Менухин-Ласовский, Йотам; Величко, Тарас; Кешет, Гал; Фан, Руи; Беджов, Иван; Черч, Джордж М.; Яух, Ральф; Кожокару, Влад; Шёлер, Ганс Р.; Величко, Сергей (декабрь 2023 г.). «Химерный супер-SOX с высокой степенью взаимодействия вызывает наивную плюрипотентность у разных видов» . Клеточная стволовая клетка . дои : 10.1016/j.stem.2023.11.010 .
- ^ Слышал E (июнь 2004 г.). «Последние достижения в инактивации Х-хромосомы». Современное мнение в области клеточной биологии . 16 (3): 247–255. дои : 10.1016/j.ceb.2004.03.005 . ПМИД 15145348 .
- ^ Беддингтон Р.С., Робертсон Э.Дж. (январь 1999 г.). «Развитие оси и ранняя асимметрия у млекопитающих» . Клетка . 96 (2): 195–209. дои : 10.1016/s0092-8674(00)80560-7 . ПМИД 9988215 . S2CID 16264083 .
- ^ Лоусон К.А., Менесес Дж.Дж., Педерсен Р.А. (ноябрь 1991 г.). «Клональный анализ судьбы эпибластов во время формирования зародышевого слоя у эмбриона мыши». Разработка . 113 (3): 891–911. дои : 10.1242/dev.113.3.891 . ПМИД 1821858 . S2CID 17685207 .
- ^ Россант Дж. (февраль 2008 г.). «Стволовые клетки и раннее развитие линий» . Клетка . 132 (4): 527–531. дои : 10.1016/j.cell.2008.01.039 . ПМИД 18295568 . S2CID 14128314 .
- ^ Бронс И.Г., Смитерс Л.Е., Троттер М.В., Рагг-Ганн П., Сан Б., Чува де Соуза Лопес С.М. и др. (июль 2007 г.). «Получение плюрипотентных стволовых клеток эпибласта из эмбрионов млекопитающих». Природа . 448 (7150): 191–195. Бибкод : 2007Natur.448..191B . дои : 10.1038/nature05950 . ПМИД 17597762 . S2CID 4365390 .
- ^ Осорно Р., Цакиридис А., Вонг Ф., Камбрей Н., Эконому С., Уилки Р. и др. (июль 2012 г.). «Разрушение плюрипотентности в процессе развития обращается вспять за счет эктопической экспрессии Oct4» . Разработка . 139 (13): 2288–2298. дои : 10.1242/dev.078071 . ПМК 3367440 . ПМИД 22669820 .
- ^ Икеучи М., Фаверо Д.С., Сакамото Ю., Ивасе А., Коулман Д., Раймен Б., Сугимото К. (апрель 2019 г.). «Молекулярные механизмы регенерации растений» . Ежегодный обзор биологии растений . 70 (1). Годовые обзоры : 377–406. doi : 10.1146/annurev-arplant-050718-100434 . ПМИД 30786238 . S2CID 73498853 .
- ^ Джорджетти А., Маркетто М.С., Ли М., Ю Д., Фаццина Р., Му Ю. и др. (июль 2012 г.). «Нейрональные клетки, полученные из пуповинной крови, путем эктопической экспрессии Sox2 и c-Myc» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (31): 12556–12561. Бибкод : 2012PNAS..10912556G . дои : 10.1073/pnas.1209523109 . ПМК 3412010 . ПМИД 22814375 .
- ^ Гуан К., Найерния К., Майер Л.С., Вагнер С., Дрессел Р., Ли Дж.Х. и др. (апрель 2006 г.). «Плюрипотентность сперматогониальных стволовых клеток из семенников взрослой мыши». Природа . 440 (7088): 1199–1203. Бибкод : 2006Natur.440.1199G . дои : 10.1038/nature04697 . ПМИД 16565704 . S2CID 4350560 .
- ^ Чжао Ю, Маццоне Т (декабрь 2010 г.). «Стволовые клетки пуповинной крови человека и путь к лекарству от диабета 1 типа». Обзоры аутоиммунитета . 10 (2): 103–107. дои : 10.1016/j.autrev.2010.08.011 . ПМИД 20728583 .
- ^ Таллоне Т., Реалини С., Бёмлер А., Корнфельд С., Вассалли Г., Моччетти Т. и др. (апрель 2011 г.). «Жировая ткань взрослого человека содержит несколько типов мультипотентных клеток». Журнал сердечно-сосудистых трансляционных исследований . 4 (2): 200–210. дои : 10.1007/s12265-011-9257-3 . ПМИД 21327755 . S2CID 36604144 .
- ^ Бельтрами А.П., Барлучки Л., Торелла Д., Бейкер М., Лимана Ф., Чименти С. и др. (сентябрь 2003 г.). «Взрослые сердечные стволовые клетки мультипотентны и поддерживают регенерацию миокарда» . Клетка . 114 (6): 763–776. дои : 10.1016/S0092-8674(03)00687-1 . ПМИД 14505575 . S2CID 15588806 .
- ^ Огуши Х., Арима Н., Такетани Т. (декабрь 2011 г.). «[Регенеративная терапия с использованием аллогенных мезенхимальных стволовых клеток]». Нихон Ринсё. Японский журнал клинической медицины (на японском языке). 69 (12): 2121–2127. ПМИД 22242308 .
- ^ Уччелли А., Моретта Л., Пистойя В. (сентябрь 2008 г.). «Мезенхимальные стволовые клетки в здоровье и болезни». Обзоры природы. Иммунология . 8 (9): 726–736. дои : 10.1038/nri2395 . ПМИД 19172693 . S2CID 3347616 .
- ^ Ибельгауфтс, Хорст. «Интернет-энциклопедия Pathfinder «Цитокины и клетки»» . Проверено 25 апреля 2013 г.
- ^ Беттс, Дж. Гордон; Дезе, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Круз, Дин; По, Брэндон; Мудро, Джеймс; Уомбл, Марк Д; Янг, Келли А. (8 июня 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 3.5 Рост и деление клеток. ISBN 978-1-947172-04-3 .
- ^ «дифференцировка гепатобластов» . ГОНУТЫ . Архивировано из оригинала 3 марта 2016 г. Проверено 31 августа 2012 г.
Внешние ссылки
[ редактировать ]| Источники/типы | |
|---|---|
| Эффективность клеток |
|
| Похожие статьи | |