Клеточная эффективность
Клеточная эффективность - это дифференцироваться способность клеток в другие типы клеток. [ 1 ] [ 2 ] Чем больше типов ячейки могут дифференцироваться, тем больше его активность. Потенция также описывается как потенциал активации гена в ячейке, который, как и континуум, начинается с тотапотенции для обозначения ячейки с наибольшим количеством потенциала дифференцировки, плюрипотентностью , мультипотентностью , олигопотентностью и, наконец, унипотентностью .

Тотипотентность
[ редактировать ]Тотипотентность (латинская: тотапотенция , Lit. «Способность ко всем [вещам]») - это способность одной клетки разделять и продуцировать все дифференцированные клетки в организме . Споры и зиготы являются примерами тотипотентных клеток. [ 3 ] В спектре эффективности клеточной активности тотиппотентность представляет собой клетку с наибольшим потенциалом дифференцировки , способный дифференцироваться в любую эмбриональную клетку, а также любую внеэмбрионусную ткань . Напротив, плюрипотентные клетки могут различаться только на эмбриональные клетки. [ 4 ] [ 5 ]
Полностью дифференцированная ячейка может вернуться в состояние тотапотенции. [ 6 ] Преобразование в Totipatency является сложным и не до конца понятно. В 2011 году исследование показало, что клетки могут дифференцироваться не в полностью тотипотентной клетке, а вместо этого в «сложную клеточную вариацию» тотапотентности. [ 7 ]
Модель развития человека может быть использована для описания того, как возникают тотапотентные клетки. [ 8 ] Человеческое развитие начинается, когда сперма оплодотворяет яйцо, а результирующее оплодотворенное яйцо создает одну тотипотентную клетку, зиготу . [ 9 ] В первые часы после оплодотворения эта зигота делится на идентичные тотипотентные клетки, которые впоследствии могут развиваться в любой из трех зародышевых слоев человека ( эндодерма , мезодерма или эктодерма ) или в клетки плаценты ( цитотрофобласт или синцитотрофласт ). После достижения 16-клеточной стадии, тотипотентные клетки моралы дифференцируются в клетки, которые в конечном итоге станут либо , бластоцисты внутренней клеточной массой либо наружными трофобластами . Приблизительно через четыре дня после оплодотворения и после нескольких циклов деления клеток эти тотипотентные клетки начинают специализироваться. Внутренняя клеточная масса, источник эмбриональных стволовых клеток , становится плюрипотентом.
Исследования по Caenorhabditis elegans предполагают, что множественные механизмы, включая регуляцию РНК, могут играть роль в поддержании тотапотентности на разных стадиях развития у некоторых видов. [ 10 ] Работа с рыбок данио и млекопитающими предполагает дальнейшее взаимодействие между miRNA и РНК-связывающими белками (RBP) при определении различий в развитии. [ 11 ]
Изначальные зародышевые клетки
[ редактировать ]В первичных зародышевых клетках , мыши перепрограммирование генома приводящее к тотипотентности, включает в себя стирание эпигенетических импринтов. Перепрограммирование облегчается активным деметилированием ДНК ДНК с участием ферментативного пути репарации иссечения . [ 12 ] Этот путь влечет за собой стирание метилирования CPG (5MC) в первичных зародышевых клетках посредством начального преобразования от 5 мкм в 5-гидроксиметилцитозин (5HMC), реакция, обусловленная высокими уровнями диоксигеназы с десятинадцатиденазой. Tet-1 и Tet-2 . [ 13 ]
Плюрипотентность
[ редактировать ]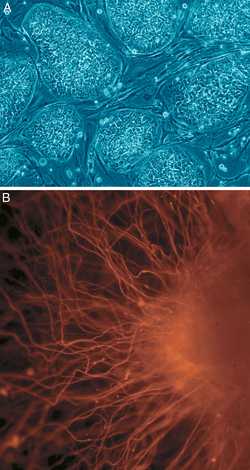
Б: нервные клетки
В клеточной биологии плюрипотентность (латинская: плюрипотенция , горит. «Способность для многих [вещей]») [ 14 ] Относится к стволовой клетке, которая может дифференцироваться в любом из трех слоев зародышей : эндодерма (кишечник, легкие и печень), мезодерму (мышцы, скелет, сосудистый, кровоогенитальный, дерма) или эктодерма (нервный, чувственный, эпидермис ), но не в экстра-эмбриональные ткани, такие как плацента или желток. [ 15 ]
Индуцированная плюрипотентность
[ редактировать ]Индуцированные плюрипотентные стволовые клетки, обычно сокращенные в виде IPS-клеток или IPSC, представляют собой тип плюрипотентных стволовых клеток, искусственно полученных из нелурипотентной клетки, как правило, взрослой соматической клетки , путем индуцирования «вынужденной» экспрессии определенных генов и трансписных факторов . [ 16 ] Эти факторы транскрипции играют ключевую роль в определении состояния этих клеток, а также подчеркивают тот факт, что эти соматические клетки сохраняют ту же генетическую информацию, что и ранние эмбриональные клетки. [ 17 ] Способность индуцировать клетки в плюрипотентное состояние была первоначально впервые с использованием фибробластов мышей и четырех факторов транскрипции, OCT4 , SOX2 , KLF4 и C- MYC ; [ 18 ] Эта техника, называемая перепрограммированием , позже заработала Шинья Яманаку и Джон Гурдон Нобелевскую премию по физиологии или медицине. [ 19 ] Затем в 2007 году последовали успешную индукцию человеческих IPSC, полученных из кожных фибробластов человека, используя методы, аналогичные тем, которые использовались для индукции клеток мыши. [ 20 ] Эти индуцированные клетки демонстрируют сходные признаки с эмбриональными стволовыми клетками (ESCS), но не требуют использования эмбрионов. Некоторые из сходств между ESC и IPSC включают плюрипотентность, морфологию , способность к самообновлению, черту, которая подразумевает, что они могут разделить и повторять бесконечно и экспрессию генов . [ 21 ]
Считается, что эпигенетические факторы участвуют в фактическом перепрограммировании соматических клеток, чтобы вызвать плюрипотентность. Было теоретизировано, что некоторые эпигенетические факторы могут действительно работать, чтобы очистить оригинальные соматические эпигенетические метки, чтобы получить новые эпигенетические метки, которые являются частью достижения плюрипотентного состояния. Хроматин также реорганизован в IPSCS и становится таким, что он находится в ESC, поскольку он менее сжатый и, следовательно, более доступный. Модификации эухроматина также распространены, что также согласуется с состоянием эухроматина, обнаруженного в ESCS. [ 21 ]
Из -за их большого сходства с ESCS медицинские и исследовательские сообщества заинтересованы в IPSC. IPSC могут потенциально иметь те же терапевтические последствия и применения, что и ESC, но без спорного использования эмбрионов в процессе, тема великих биоэтических дебатов. Индуцированная плюрипотентность соматических клеток в недифференцированные клетки IPS первоначально приветствовали как конец спорного использования эмбриональных стволовых клеток . Тем не менее, было обнаружено, что IPSC потенциально онкогенные и, несмотря на достижения, [ 16 ] никогда не были одобрены для исследования клинической стадии в Соединенных Штатах до недавнего времени. В настоящее время аутологичные дофаминергические предшественники, полученные из IPSC, используются в исследованиях для лечения болезни Паркинсона. [ 22 ] Такие неудачи, как низкий уровень репликации и раннее старение, также встречались при создании IPSC, [ 23 ] препятствуя их использованию в качестве замены ESCS.
Соматическая экспрессия комбинированных факторов транскрипции может непосредственно индуцировать другие определенные судьбы соматических клеток ( трансдифференцировка ); Исследователи идентифицировали три фактора транскрипции специфических для нейролерости, которые могли бы непосредственно преобразовать фибробласты мыши (клеток соединительной ткани) в полностью функциональные нейроны . [ 24 ] Этот результат бросает вызов терминальной природе клеточной дифференцировки и целостности линии; и подразумевает, что с помощью правильных инструментов все клетки являются тотипентными и могут образовывать все виды ткани.
Некоторые из возможных медицинских и терапевтических применений для IPSC, полученных от пациентов, включают их применение в трансплантации клеток и тканей без риска отторжения, который обычно встречается. IPSC могут потенциально заменить модели на животных как не подходящие, так и модели in vitro, используемые для исследований заболеваний. [ 25 ]

Наив против первичных плюрипотентности
[ редактировать ]Результаты в отношении эпибластов до и после имплантации дали предложения для классификации плюрипотентности на два состояния: «наивные» и «заполненные», представляющие эпибласт до и постмплантации соответственно. [ 26 ] Наивный-насокий континуум контролируется уменьшением димеризации SOX2/OCT4 на элементах ДНК SOXOCT, контролирующих наивную плюрипотентность. [ 27 ] Приправленные плюрипотентные стволовые клетки из разных видов могут быть сброшены в наивное состояние, используя коктейль, содержащий KLF4 и SOX2 или «супер-сокс»-химерный фактор транскрипции с повышенной способностью димеризировать с OCT4. [ 27 ]
Базовые стволовые клетки, обычно используемые в науке, которые называются эмбриональными стволовыми клетками (ESC), получены из предкмплантационной эпибласта; Такой эпибласт способен генерировать весь плод, и одна клетка эпибластов способна вносить вклад во все клеточные линии, если он вводится в другую бластоцисту. С другой стороны, можно наблюдать несколько заметных различий между эпибластами до и после имплантации, такими как их разница в морфологии, в которых эпибласт после имплантации превращает свою морфологию в чашеподобную форму, называемую «яичным цилиндром» как а также хромосомное изменение, при котором одна из X-хромосомов при случайной инактивации на ранней стадии яичного цилиндра, известного как X-инактивация . [ 28 ] Во время этого развития клетки эпибластов яичного цилиндра систематически нацелены на факторы роста фибробластов , передачу сигналов Wnt и другие индуктивные факторы через окружающий желток и ткань трофобласта, [ 29 ] Таковы, что они становятся инструктивными конкретными в соответствии с пространственной организацией. [ 30 ]
Другое важное отличие состоит в том, что стволовые клетки после имплантации не могут внести свой вклад в бластоцисты, химеры , [ 31 ] что отличает их от других известных плюрипотентных стволовых клеток. Клеточные линии, полученные из таких постмплантационных эпибластов, называются стволовыми клетками, полученными из эпибластов , которые впервые были получены в лаборатории в 2007 году. И ESC, и EPISC получены из эпибластов, но на различных фазах развития. Плюрипотентность все еще не повреждена в эпибласте после имплантации, о чем свидетельствуют консервативная экспрессия Nanog , FUT4 и Oct-4 в Episcs, [ 32 ] до сомитогенеза и может быть изменен на полпути через индуцированную экспрессию OCT-4 . [ 33 ]
Местная плюрипотентность в растениях
[ редактировать ]
Неиндуцированная плюрипотентность наблюдалась в культуре тканей корневой меристемы , особенно в Кариме и др. 2015, Ким и др. 2018, и Роспопофф и др. 2017. Эта плюрипотентность регулируется различными регуляторами, включая PLETHORA 1 и PLETHORA 2 ; и PLETHORA 3 , PLETHORA 5 , и PLTHORA 7 , выражение которого Карим был обнаружено для ауксина . (Они также известны как PLT1, PLT2, PLT3, PLT5, PLT7 и экспрессируются генами одинаковых имен.) По состоянию на 2019 год. [update]Ожидается, что это откроет будущие исследования плюрипотентности в корневых тканях. [ 34 ]
Мультипотентность
[ редактировать ]
Мультипотентность - это когда клетки -предшественники имеют потенциал активации гена, чтобы дифференцироваться в дискретные типы клеток. Например, гематопоэтическая стволовая клетка - и этот тип клеток может дифференцироваться в несколько типов клеток крови, таких как лимфоциты , моноциты , нейтрофилы и т. Д., Но все еще неоднозначно, обладает ли HSC способность дифференцироваться в клетки мозга , костные клетки или Другие типы неровных клеток. [ Цитация необходима ]
Исследования, связанные с мультипотентными клетками, позволяют предположить, что мультипотентные клетки могут быть способны преобразовать в не связанные типы клеток. В другом случае стволовые клетки пуповинной крови человека были преобразованы в нейроны человека. [ 35 ] Существует также исследование по преобразованию мультипотентных клеток в плюрипотентные клетки. [ 36 ]
Мультипотентные клетки встречаются во многих, но не во всех типах клеток человека. Мультипотентные клетки были обнаружены в пуповинной крови , [ 37 ] жировая ткань, [ 38 ] сердечные клетки, [ 39 ] костный мозг и мезенхимальные стволовые клетки (MSC), которые обнаруживаются в третьем моляре . [ 40 ]
MSC могут оказаться ценным источником для стволовых клеток из моляров в возрасте 8–10 лет, до кальцификации зубов взрослых. MSC могут дифференцироваться в остеобласты, хондроциты и адипоциты. [ 41 ]
Олигопотентность
[ редактировать ]В биологии олигопотентность - это способность клеток -предшественников дифференцироваться в несколько типов клеток . Это степень потенции . Примерами олигопотентных стволовых клеток являются лимфоидные или миелоидные стволовые клетки. [ 2 ] В частности, лимфоидные клетки могут вызывать различные клетки крови, такие как B и T -клетки, однако, не к другому типу крови, как эритроциты. [ 42 ] Примерами клеток -предшественников являются сосудистые стволовые клетки, которые способны стать как эндотелиальными , так и гладких мышечных клетками.
Unipatency
[ редактировать ]В клеточной биологии однопотентная клетка - это концепция, что одна стволовая клетка способна дифференцироваться только на один тип клеток. [ 43 ] В настоящее время неясно, существуют ли истинные унипотентные стволовые клетки. Гепатобласты, которые дифференцируются в гепатоциты (которые составляют большую часть печени ) или холангиоциты (эпителиальные клетки желчного протока), являются бипотентными. [ 44 ] Беспокойным синоним однопотентной ячейки является ячейка -предшественника .
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Клеточная тотазотенция была обнаружена Хабертлендом, и этот термин был придуман Томасом Хандом Морганом. Махла Р.С. (2016). «Применение стволовых клеток в регенеративной медицине и терапии заболевания» . Международный журнал клеточной биологии . 2016 (7): 6940283. DOI : 10.1155/2016/6940283 . PMC 4969512 . PMID 27516776 .
- ^ Jump up to: а беременный Шёлер HR (2007). «Потенциал стволовых клеток: инвентарь». В Knoepffler M, Schipanski D, Sorgner SL (Eds.). Человеческая биотехнология как социальная проблема . Ashgate Publishing, Ltd. p. 28. ISBN 978-0-7546-5755-2 .
- ^ Mitalipov S , Wolf D (2009). «Тотипотентность, плюрипотентность и ядерное перепрограммирование». Инженерия стволовых клеток . Достижения в области биохимической инженерии/биотехнологии. Тол. 114. С. 185–199. Bibcode : 2009esc..book..185m . doi : 10.1007/10_2008_45 . ISBN 978-3-540-88805-5 Полем PMC 2752493 . PMID 19343304 .
- ^ Lodish H (2016). Молекулярная клеточная биология (8 -е изд.). WH Freeman. С. 975–977. ISBN 978-1319067748 .
- ^ «В чем разница между тотипотентом, плюрипотентом и мультипотентом?» Полем
- ^ Western P (2009). «Зародовые клетки плода: удаление баланса между плюрипотентностью и дифференцировкой» . Международный журнал биологии развития . 53 (2–3): 393–409. doi : 10.1387/ijdb.082671pw . PMID 19412894 .
- ^ Sugimoto K, Gordon SP, Meyerowitz EM (апрель 2011 г.). «Регенерация у растений и животных: дедифференциация, трансдифференциация или просто дифференциация?». Тенденции в клеточной биологии . 21 (4): 212–218. doi : 10.1016/j.tcb.2010.12.004 . PMID 21236679 .
- ^ Сейду Г., Браун Р.Е. (декабрь 2006 г.). «Путь к тотипотентности: уроки из зародышевых клеток» . Клетка . 127 (5): 891–904. doi : 10.1016/j.cell.2006.11.016 . PMID 17129777 . S2CID 16988032 .
- ^ Asch R, Simerly C, Ord T, Ord Va, Schatten G (июль 1995). «Стадии, на которых арестовывает человеческое оплодотворение: конфигурации микротрубочек и хромосом в осемененных ооцитах, которые не смогли завершить оплодотворение и развитие у людей». Человеческое воспроизведение . 10 (7): 1897–1906. doi : 10.1093/oxfordjournals.humrep.a136204 . PMID 8583008 .
- ^ Ciosk R, Depalma M, Priess Jr (февраль 2006 г.). «Трансляционные регуляторы поддерживают тотипотентность в зародышевой линии Caenorhabditis elegans». Наука . 311 (5762): 851–853. Bibcode : 2006sci ... 311..851c . doi : 10.1126/science.1122491 . PMID 16469927 . S2CID 130017 .
- ^ Kedde M, Agami R (апрель 2008 г.). «Взаимодействие между микроРНК и РНК-связывающими белками определяет процессы развития» . Клеточный цикл . 7 (7): 899–903. doi : 10.4161/cc.7.7.5644 . PMID 18414021 .
- ^ Хаджкова П., Джеффрис С.Дж., Ли С., Миллер Н., Джексон С.П., Сурани Ма (июль 2010 г.). «Репрограммирование по всему геному в линии зародышевой мыши влечет за собой базовый путь восстановления иссечения» . Наука . 329 (5987): 78–82. Bibcode : 2010sci ... 329 ... 78H . doi : 10.1126/science.1187945 . PMC 3863715 . PMID 20595612 .
- ^ Хакетт Дж.А., Сенгупта Р., Зилиши Дж.Дж., Мураками К., Ли С., Даун Т.А., Сурани Ма (январь 2013 г.). «Динамика деметилирования ДНК зародышевой линии и стирание отпечатка через 5-гидроксиметилцитозин» . Наука . 339 (6118): 448–452. Bibcode : 2013sci ... 339..448H . doi : 10.1126/science.1229277 . PMC 3847602 . PMID 23223451 .
- ^ «Биология онлайн» . Biology-online.org . Получено 25 апреля 2013 года .
- ^ Binder MD, Hirokawa N, Nobutaka, Windhorst U, Eds. (2009). Энциклопедия нейробиологии . Берлин: Спрингер. ISBN 978-3540237358 .
- ^ Jump up to: а беременный Бейкер М. (2007-12-06). «Взрослые клетки перепрограммированы в плюрипотентность без опухолей» . Nature Reports Стволовые клетки : 1. doi : 10.1038/stemcells.2007.124 .
- ^ Stadtfeld M, Hochedlinger K (октябрь 2010 г.). «Индуцированная плюрипотентность: история, механизмы и приложения» . Гены и развитие . 24 (20): 2239–2263. doi : 10.1101/gad.1963910 . PMC 2956203 . PMID 20952534 .
- ^ Такахаши К, Яманака С (август 2006 г.). «Индукция плюрипотентных стволовых клеток из культур эмбриональных и фибробластов мыши с помощью определенных факторов». Клетка . 126 (4): 663–676. doi : 10.1016/j.cell.2006.07.024 . HDL : 2433/159777 . PMID 16904174 . S2CID 1565219 .
- ^ « Нобелевская премия по физиологии или медицине 2012 ». Nobelprize.org. Nobel Media AB 2013. Веб. 28 ноября 2013 года.
- ^ Такахаши К., Танабе К., Онуки М., Нарита М., Ичисака Т., Томода К, Яманака С. (ноябрь 2007 г.). «Индукция плюрипотентных стволовых клеток из фибробластов взрослого человека с помощью определенных факторов». Клетка . 131 (5): 861–872. doi : 10.1016/j.cell.2007.11.019 . HDL : 2433/49782 . PMID 18035408 . S2CID 8531539 .
- ^ Jump up to: а беременный Liang G, Zhang Y (январь 2013 г.). «Эмбриональная стволовая клетка и индуцированные плюрипотентные стволовые клетки: эпигенетическая перспектива» . Клеточные исследования . 23 (1): 49–69. doi : 10.1038/cr.2012.175 . PMC 3541668 . PMID 23247625 .
- ^ Schweitzer JS, Song B, Herrington TM, Park TY, Lee N, Ko S, et al. (Май 2020). «Персонализированные клетки дофамина, полученные из IPSC, при болезни Паркинсона» . Новая Англия Журнал медицины . 382 (20): 1926–1932. doi : 10.1056/nejmoa1915872 . PMC 7288982 . PMID 32402162 .
- ^ Чой, Чарльз. «Cell-Off: индуцированные плюрипотентные стволовые клетки не достигают потенциала, обнаруженного в эмбриональной версии» . Scientific American . Получено 25 апреля 2013 года .
- ^ Vierbuchen T, Ostermeier A, Pang ZP, Kokubu Y, Südhof TC, Wernig M (февраль 2010 г.). «Прямое преобразование фибробластов в функциональные нейроны с помощью определенных факторов» . Природа . 463 (7284): 1035–1041. Bibcode : 2010natur.463.1035V . doi : 10.1038/nature08797 . PMC 2829121 . PMID 20107439 .
- ^ Park IH, Lerou PH, Zhao R, Huo H, Daley GQ (2008). «Генерация индуцированных человеком плюрипотентных стволовых клеток». Природные протоколы . 3 (7): 1180–1186. doi : 10.1038/nprot.2008.92 . PMID 18600223 . S2CID 13321484 .
- ^ Николс Дж., Смит А (июнь 2009 г.). «Наивные и придуманные плюрипотентные состояния» . Клеточная стволовая клетка . 4 (6): 487–492. doi : 10.1016/j.stem.2009.05.015 . PMID 19497275 .
- ^ Jump up to: а беременный MacCarthy, Caitlin M.; Ву, Гуанминг; Малик, Викас; Menuchin-Lasowski, Yotam; Элечко, Тарас; Кеше, Гал; Фанат, Руи; Бедцхов, Иван; Церковь, Джордж М.; Джаух, Ральф; Cojocaru, VLAD; Шёлер, Ганс Р.; Вельчко, Сергия (декабрь 2023 г.). «Высокий кооперативный химерный супер-SOX вызывает наивную плюрипотентность по видам» . Клеточная стволовая клетка . doi : 10.1016/j.stem.2023.11.010 .
- ^ Слышал E (июнь 2004 г.). «Последние достижения в области инактивации X-Chromosome». Современное мнение в клеточной биологии . 16 (3): 247–255. doi : 10.1016/j.ceb.2004.03.005 . PMID 15145348 .
- ^ Беддингтон Р.С., Робертсон Э.Дж. (январь 1999 г.). «Развитие оси и ранняя асимметрия у млекопитающих» . Клетка . 96 (2): 195–209. doi : 10.1016/s0092-8674 (00) 80560-7 . PMID 9988215 . S2CID 16264083 .
- ^ Лоусон К.А., Менесес Дж.Дж., Педерсен Р.А. (ноябрь 1991 г.). «Клональный анализ судьбы эпибластов во время образования зародышевых слоев у эмбриона мыши». Разработка . 113 (3): 891–911. doi : 10.1242/dev.113.3.891 . PMID 1821858 . S2CID 17685207 .
- ^ Россант Дж (февраль 2008 г.). «Стволовые клетки и раннее развитие линии» . Клетка . 132 (4): 527–531. doi : 10.1016/j.cell.2008.01.039 . PMID 18295568 . S2CID 14128314 .
- ^ Brons IG, Smithers LE, Totter MW, Rugg-Gunn P, Sun B, Chuva de Sousa Lopes SM и др. (Июль 2007 г.). «Вывод плюрипотентных стволовых клеток эпибластов от эмбрионов млекопитающих». Природа . 448 (7150): 191–195. Bibcode : 2007natur.448..191b . doi : 10.1038/nature05950 . PMID 17597762 . S2CID 4365390 .
- ^ Osorno R, Tsakiridis A, Wong F, Cambray N, Ecoverou C, Wilkie R, et al. (Июль 2012 г.). «Развитие демонтажа плюрипотентности изменяется в результате эктопической экспрессии OCT4» . Разработка . 139 (13): 2288–2298. doi : 10.1242/dev.078071 . PMC 3367440 . PMID 22669820 .
- ^ Икеучи М., Фаверо Д.С., Сакамото Ю., Ивасе А., Коулман Д., Раймен Б., Сугимото К (апрель 2019 г.). «Молекулярные механизмы регенерации растений» . Ежегодный обзор биологии растений . 70 (1). Ежегодные обзоры : 377–406. doi : 10.1146/annurev-arplant-050718-100434 . PMID 30786238 . S2CID 73498853 .
- ^ Giorgetti A, Marchetto MC, Li M, Yu D, Fazzina R, Mu Y, et al. (Июль 2012 г.). «Нейрональные клетки, полученные из пуповины, путем эктопической экспрессии SOX2 и C-Myc» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (31): 12556–12561. BIBCODE : 2012PNAS..10912556G . doi : 10.1073/pnas.1209523109 . PMC 3412010 . PMID 22814375 .
- ^ Guan K, Nayernia K, Maier LS, Wagner S, Dressel R, Lee JH, et al. (Апрель 2006 г.). «Плюрипотентность сперматогониальных стволовых клеток от яичка для взрослых мышей». Природа . 440 (7088): 1199–1203. Bibcode : 2006natur.440.1199G . doi : 10.1038/nature04697 . PMID 16565704 . S2CID 4350560 .
- ^ Чжао Y, Mazzone T (декабрь 2010 г.). «Стволовые клетки пуповинной крови человека и путешествие к лечению от диабета 1 типа». Обзоры аутоиммунитета . 10 (2): 103–107. doi : 10.1016/j.autrev.2010.08.011 . PMID 20728583 .
- ^ Tallone T, Realini C, Böhmler A, Kornfeld C, Vassalli G, Moccetti T, et al. (Апрель 2011). «Жировая ткань для взрослых содержит несколько типов мультипотентных клеток». Журнал сердечно -сосудистых трансляционных исследований . 4 (2): 200–210. doi : 10.1007/s12265-011-9257-3 . PMID 21327755 . S2CID 36604144 .
- ^ Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S, et al. (Сентябрь 2003 г.). «Взрослые сердечные стволовые клетки являются мультипотентными и поддерживают регенерацию миокарда» . Клетка . 114 (6): 763–776. doi : 10.1016/s0092-8674 (03) 00687-1 . PMID 14505575 . S2CID 15588806 .
- ^ Ohgushi H, Arima N, Taketani T (декабрь 2011 г.). «[Регенеративная терапия с использованием аллогенных мезенхимальных стволовых клеток]». Нихон Риншо. Японский журнал клинической медицины (на японском языке). 69 (12): 2121–2127. PMID 22242308 .
- ^ Uccelli A, Moretta L, Pistoia V (сентябрь 2008 г.). «Мезенхимальные стволовые клетки со здоровьем и заболеванием». Природные обзоры. Иммунология . 8 (9): 726–736. doi : 10.1038/nri2395 . PMID 19172693 . S2CID 3347616 .
- ^ Ibelgaufts, Horst. «Цитокины и клетки онлайн -энциклопедия Pathfinder» . Получено 25 апреля 2013 года .
- ^ Беттс, Дж. Гордон; Десакс, Питер; Джонсон, Эдди; Джонсон, Джоди Э; Король, Оксана; Крузе, Дин; По, Брэндон; Мудрый, Джеймс; Womble, Mark D; Янг, Келли А (8 июня 2023 г.). Анатомия и физиология . Хьюстон: OpenStax CNX. 3.5 Рост и деление клеток. ISBN 978-1-947172-04-3 .
- ^ «Дифференциация гепатобластов» . Гон . Архивировано с оригинала 2016-03-03 . Получено 2012-08-31 .
Внешние ссылки
[ редактировать ]| Источники/типы | |
|---|---|
| Клеточная эффективность |
|
| Связанные статьи | |