Периферический мембранный белок
Периферические мембранные белки или внешние мембранные белки . [1] представляют собой мембранные белки , которые лишь временно прикрепляются к биологической мембране , с которой они связаны. Эти белки прикрепляются к интегральным мембранным белкам или проникают в периферические области липидного бислоя . регуляторные белковые субъединицы многих ионных каналов и трансмембранных рецепторов Например, можно определить как периферические мембранные белки. В отличие от интегральных мембранных белков, периферические мембранные белки имеют тенденцию накапливаться в водорастворимом компоненте или фракции всех белков, экстрагированных во время процедуры очистки белка . Белки с якорями GPI являются исключением из этого правила и могут иметь свойства очистки, аналогичные свойствам интегральных мембранных белков.
Было показано, что обратимое прикрепление белков к биологическим мембранам регулирует передачу сигналов в клетках и многие другие важные клеточные события посредством множества механизмов. [2] Например, тесная связь между многими ферментами и биологическими мембранами может привести к тому, что они будут находиться в непосредственной близости от своего липидного субстрата (ов). [3] Мембранное связывание может также способствовать перестройке, диссоциации или конформационным изменениям внутри многих структурных доменов белков, что приводит к активации их биологической активности . [4] [5] Кроме того, расположение многих белков локализовано либо на внутренней, либо на внешней поверхности или листках их резидентной мембраны. [6] Это облегчает сборку мультибелковых комплексов за счет увеличения вероятности любых соответствующих белок-белковых взаимодействий .

Связывание с липидным бислоем
[ редактировать ]
Белки периферической мембраны могут взаимодействовать с другими белками или непосредственно с липидным бислоем . В последнем случае их называют амфитропными белками. [4] Некоторые белки, такие как G-белки и некоторые протеинкиназы , одновременно взаимодействуют с трансмембранными белками и липидным бислоем. Некоторые полипептидные гормоны , антимикробные пептиды и нейротоксины накапливаются на поверхности мембраны до того, как локализуются и взаимодействуют с рецепторами клеточной поверхности, которые сами по себе могут быть периферическими мембранными белками.
Фосфолипидный бислой , образующий поверхностную мембрану клетки, состоит из гидрофобной внутренней сердцевинной области, зажатой между двумя областями гидрофильности : одной на внутренней поверхности, а другой на внешней поверхности клеточной мембраны ( липидном бислое) более подробное структурное описание см. в статье о . клеточная мембрана). Было показано, что внутренняя и внешняя поверхности или межфазные области модельных фосфолипидных бислоев имеют толщину от 8 до 10 Å , хотя она может быть шире в биологических мембранах , которые включают большое количество ганглиозидов или липополисахаридов . [7] Гидрофобная внутренняя область ядра типичных биологических мембран может иметь толщину от 27 до 32 Å, по оценкам метода малоуглового рентгеновского рассеяния (SAXS) . [8] Граница между гидрофобным внутренним ядром и гидрофильными межфазными областями очень узкая, около 3 Å ( липидном бислое описание составляющих его химических групп см. в статье о ). При движении наружу от гидрофобной центральной области к межфазной гидрофильной области эффективная концентрация воды быстро меняется в этом пограничном слое от почти нуля до концентрации около М. 2 [9] [10] Фосфатные группы внутри фосфолипидных бислоев полностью гидратированы или насыщены водой и расположены примерно на 5 Å за границей области гидрофобного ядра. [11]
Некоторые водорастворимые белки необратимо связываются с липидными бислоями и могут образовывать трансмембранные альфа-спиральные или бета-бочкообразные каналы. Подобные превращения происходят в порообразующих токсинах, таких как колицин А, альфа-гемолизин и других. Они также могут встречаться в BcL-2-подобном белке , в некоторых амфифильных антимикробных пептидах и в некоторых аннексинах . Эти белки обычно описываются как периферические, поскольку одно из их конформационных состояний водорастворимо или слабо связано с мембраной. [12]
Мембранные механизмы связывания
[ редактировать ]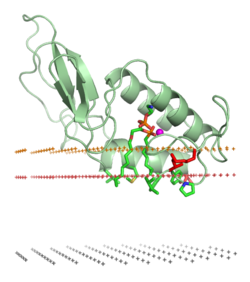
Ассоциация белка с липидным бислоем может включать значительные изменения в третичной структуре белка. Они могут включать в себя сворачивание областей белковой структуры, которые ранее были развернуты, или реорганизацию сворачивания или рефолдинг связанной с мембраной части белков. Он также может включать образование или диссоциацию белковых четвертичных структур или олигомерных комплексов , а также специфическое связывание ионов , лигандов или регуляторных липидов .
Типичные амфитропные белки должны сильно взаимодействовать с липидным бислоем, чтобы выполнять свои биологические функции. К ним относятся ферментативная обработка липидов и других гидрофобных веществ, мембранное закрепление, а также связывание и перенос небольших неполярных соединений между различными клеточными мембранами. Эти белки могут быть прикреплены к бислою в результате гидрофобных взаимодействий между бислоем и открытыми неполярными остатками на поверхности белка. [13] путем специфических нековалентных взаимодействий связывания с регуляторными липидами или посредством их прикрепления к ковалентно связанным липидным якорям .
Показано, что сродство многих периферических белков к мембраносвязыванию зависит от конкретного липидного состава мембраны, с которой они связаны. [14]

Неспецифическая гидрофобная ассоциация
[ редактировать ]Амфитропные белки связываются с липидными бислоями посредством различных гидрофобных якорных структур. Такие как амфифильные α-спирали , открытые неполярные петли, посттрансляционно ацилированные или липидированные аминокислотные остатки или ацильные цепи специфически связанных регуляторных липидов, таких как фосфатидилинозитолфосфаты . Было показано, что гидрофобные взаимодействия важны даже для высококатионных пептидов и белков, таких как многоосновный домен белка MARCKS или гистактофилин, когда присутствуют их естественные гидрофобные якоря. [15]
Ковалентно связанные липидные якоря
[ редактировать ]Белки, заякоренные в липидах, ковалентно присоединяются к различным цепям жирных кислот ацильным на цитоплазматической стороне клеточной мембраны посредством пальмитоилирования , миристоилирования или пренилирования . На экзоплазматической стороне клеточной мембраны белки, заякоренные в липидах, ковалентно прикреплены к липидам гликозилфосфатидилинозитолу (ГФИ) и холестерину . [16] [17] Ассоциация белка с мембраной за счет использования ацилированных остатков является обратимым процессом , поскольку ацильная цепь может быть похоронена в гидрофобном связывающем кармане белка после диссоциации от мембраны. Этот процесс происходит внутри бета-субъединиц G-белков . Возможно, из-за этой дополнительной потребности в структурной гибкости липидные якоря обычно связаны с очень гибкими сегментами третичной структуры белков, которые плохо определяются кристаллографическими исследованиями белков .
Специфическое белок-липидное связывание
[ редактировать ]
Некоторые цитозольные белки рекрутируются в различные клеточные мембраны, распознавая определенные типы липидов, находящихся внутри данной мембраны. [18] Связывание белка со специфическим липидом происходит через специфические мембраноориентированные структурные домены, которые встречаются внутри белка и имеют специфические карманы связывания для липидных головных групп липидов, с которыми они связываются. Это типичное биохимическое взаимодействие белок- лиганд , которое стабилизируется за счет образования межмолекулярных водородных связей , ван-дер-ваальсовых взаимодействий и гидрофобных взаимодействий между белком и липидным лигандом . Такие комплексы также стабилизируются за счет образования ионных мостиков между остатками аспартата или глутамата белка и липидофосфатами через промежуточные ионы кальция (Ca 2+ ). Такие ионные мостики могут возникать и стабильны, когда ионы (например, Ca 2+ ) уже связаны с белком в растворе до связывания с липидом. Образование ионных мостиков наблюдается при белок-липидном взаимодействии как между доменами типа белка С2, так и с аннексинами .
Белко-липидные электростатические взаимодействия
[ редактировать ]Любой положительно заряженный белок будет притягиваться к отрицательно заряженной мембране за счет неспецифических электростатических взаимодействий. Однако не все периферические пептиды и белки являются катионными, и только определенные стороны мембраны заряжены отрицательно. К ним относятся цитоплазматическая сторона плазматических мембран , внешний листок внешних мембран бактерий и митохондриальные мембраны. Следовательно, электростатические взаимодействия играют важную роль в нацеливании на мембраны переносчиков электронов , таких как цитохром c , катионные токсины, такие как харибдотоксин , и специфические мембранно-нацеленные домены, такие как некоторые домены PH , домены C1 и домены C2 .
Электростатические взаимодействия сильно зависят от ионной силы раствора. Эти взаимодействия относительно слабы при физиологической ионной силе ( 0,14 М NaCl ): от ~3 до 4 ккал/моль для небольших катионных белков, таких как цитохром с , харибдотоксин или гисактофилин . [15] [19] [20]
Пространственное положение в мембране
[ редактировать ]Ориентации и глубины проникновения многих амфитропных белков и пептидов в мембраны изучаются с помощью сайт-направленного спинового мечения . [21] химическая маркировка, измерение аффинности связывания мембран белковых мутантов , [22] флуоресцентная спектроскопия, [23] раствора или твердого тела ЯМР-спектроскопия , [24] ATR FTIR-спектроскопия , [25] Рентгеновская или нейтронная дифракция, [26] и вычислительные методы. [27] [28] [29] [30]
Были идентифицированы два различных способа мембранной ассоциации белков. Типичные водорастворимые белки не имеют открытых неполярных остатков или каких-либо других гидрофобных якорей. Поэтому они полностью остаются в водном растворе и не проникают в липидный бислой, что было бы энергетически затратно. Такие белки взаимодействуют с бислоями только электростатически, например, рибонуклеаза и полилизин таким образом с мембранами взаимодействуют . Однако типичные амфитропные белки имеют различные гидрофобные якоря, которые проникают в межфазную область и достигают углеводородной внутренней части мембраны. Такие белки «деформируют» липидный бислой, снижая температуру перехода липид-гель. [31] Связывание обычно представляет собой сильно экзотермическую реакцию. [32] Аналогично происходит ассоциация амфифильных α-спиралей с мембранами. [26] [33] Неструктурированные или развернутые пептиды с неполярными остатками или липидными якорями также могут проникать через межфазную область мембраны и достигать углеводородного ядра, особенно когда такие пептиды являются катионными и взаимодействуют с отрицательно заряженными мембранами. [34] [35] [36]
Категории
[ редактировать ]Ферменты
[ редактировать ]Периферические ферменты участвуют в метаболизме различных компонентов мембран, таких как липиды ( фосфолипазы и холестериноксидазы ), клеточной стенки олигосахариды ( гликозилтрансфераза и трансгликозидазы ) или белки ( сигнальная пептидаза и пальмитоилпротеинтиоэстераза ). Липазы также могут переваривать липиды, образующие в воде мицеллы или неполярные капли.
| Сорт | Функция | Физиология | Структура |
|---|---|---|---|
| Альфа/бета-гидролазная складка | Катализирует гидролиз химических связей. [37] | Включает бактериальные , грибковые , желудочные и панкреатические липазы , пальмитоилпротеинтиоэстеразы холинэстеразу , кутиназу и . | центральный бета-лист вставлен между двумя слоями альфа-спиралей [38] |
| Фосфолипаза А2 (секреторная и цитозольная) | Гидролиз жирнокислотной связи sn-2 фосфолипидов . [39] | Переваривание липидов, разрушение мембран и передача сигналов липидов . | содержит каталитическую триаду аминокислот: аспарагиновую кислоту , серин и гистидин. [40] |
| Фосфолипаза С | Гидролизует PIP2, фосфатидилинозитол , на два вторых информатора, инозитолтрифосфат и диацилглицерин . [41] | Липидная сигнализация | Основная структура состоит из разделенного ствола триозофосфат-изомеразы (TIM), который имеет активный центр, каталитические остатки и Ca 2+ сайт связывания [42] |
| Холестериноксидазы | Окисляет и изомеризует холестерин в холест-4-ен-3-он. [43] | Истощает клеточные мембраны холестерина, используемого в бактериальном патогенезе . | две петли остатка, которые действуют как крышка на активном сайте [44] |
| Каротиноидоксигеназа | Расщепляет каротиноиды . [45] | Каротиноиды функционируют как у растений, так и у животных в качестве гормонов (включая витамин А у человека), пигментов , ароматизаторов , цветочных ароматов и защитных соединений. | состоит из множества ферментов, соединенных вместе, образующих разветвленные структуры [46] |
| Липоксигеназы | Железосодержащие ферменты, диоксигенацию полиненасыщенных катализирующие жирных кислот . [47] | У животных липоксигеназы участвуют в синтезе медиаторов воспаления , известных как лейкотриены . | сотни аминокислот , составляющих белок, организованы в два домена: бета-лист N-концевой и спиральный C-концевой. [48] |
| Альфа-токсины | Расщепляет фосфолипиды в клеточной мембране, подобно фосфолипазе С. [49] | Бактериальный патогенез, особенно Clostridium perfringens . | растворимый мономер с олигомерными предпоровыми комплексами [50] |
| Сфингомиелиназа С | Фосфодиэстераза . расщепляет фосфодиэфирные связи [51] | Переработка липидов, таких как сфингомиелин . | Домен сапозина и соединительные области с металлофосфатным каталитическим доменом [52] |
| Гликозилтрансферазы : MurG и трансгликозидазы. | Катализирует перенос сахарных фрагментов от активированных молекул-доноров к специфическим молекулам-акцепторам, образуя гликозидные связи. [53] | Биосинтез дисахаридов , олигосахаридов и полисахаридов (гликоконъюгатов), MurG участвует в биосинтезе бактериальных пептидогликанов . | три петли, богатые глицином: одна на С-конце и две на N-конце. [54] |
| Феррохелатаза | Превращает протопорфирин IX в гем . [55] | Участвуя в порфиринов обмене , протопорфирины используются для укрепления яичной скорлупы . | полипептид , свернутый в два домена, каждый из которых имеет четырехцепочечный параллельный бета-лист, окруженный альфа- спиралями. [56] |
| Семейство белков, родственных миотубулярину | Липидфосфатаза , дефосфорилирующая PtdIns3P и PtdIns(3,5)P2 . [57] | Необходим для дифференцировки мышечных клеток. | содержит домен GRAM , взаимодействующий домен SET и PDZ. домен привязки [58] |
| Дигидрооротатдегидрогеназы | Окисление дигидрооротата (DHO) до оротата. [59] | Биосинтез пиримидиновых нуклеотидов в прокариотических и эукариотических клетках. | состоит из двух доменов: альфа-/бета-бочкового домена, который содержит активный сайт, и альфа-спирального домена, который образует открывающийся туннель к активному сайту. [60] |
| Гликолатоксидаза | Катализирует окисление α -гидроксикарбоновых кислот до соответствующих α- кетокислот . [61] | У зеленых растений фермент участвует в фотодыхании . У животных фермент участвует в производстве оксалата . | Складка β8/α8, содержащая альфа-спирали, бета-цепи, петли и повороты. [62] |
Мембранно-нацеленные домены («липидные зажимы»)
[ редактировать ]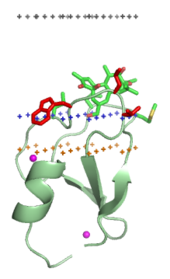
Домены, нацеленные на мембрану, специфически связываются с головными группами своих липидных лигандов, встроенных в мембрану. Эти липидные лиганды присутствуют в разных концентрациях в различных типах биологических мембран (например, PtdIns3P можно обнаружить преимущественно в мембранах ранних эндосом , PtdIns(3,5)P2 — в поздних эндосомах , а PtdIns4P — в мембранах Гольджи ). [18] Следовательно, каждый домен нацелен на определенную мембрану.
- Домены C1 и сложные эфиры форболов .
- Домены C2 связывают фосфатидилсерин , фосфатидилхолин или PtdIns(3,4)P2 или PtdIns(4,5)P2 .
- Домены гомологии плекстрина , домены PX и домены Табби связывают различные фосфоинозитиды.
- Домены FYVE более специфичны для PtdIns3P.
- Домены ENTH связывают PtdIns(3,4)P2 или PtdIns(4,5)P2 .
- ANTH связывает PtdIns(4,5)P2. Домен
- Белки семейства ERM (эзрин/радиксин/моэзин) связывают PtdIns(4,5)P2.
- Другие фосфоинозитид -связывающие белки включают фосфотирозин -связывающий домен и некоторые PDZ-домены . Они связывают PtdIns(4,5)P2.
- Дискоидиновые домены свертывания крови факторов
- ENTH , VHS и ANTH Домены
Структурные домены
[ редактировать ]Структурные домены опосредуют прикрепление других белков к мембранам. Их связывание с мембранами может осуществляться ионами кальция (Ca 2+ ), которые образуют мостики между кислыми остатками белка и фосфатными группами липидов, как в аннексинах или доменах GLA.
| Сорт | Функция | Физиология | Структура |
|---|---|---|---|
| Аннексины | Кальций мембраны и фосфолипидов . -зависимое внутриклеточное связывание [63] | Функции включают транспортировку пузырьков , слияние мембран и ионных каналов . образование | |
| Синапсин I | Покрывает синаптические пузырьки и связывается с несколькими элементами цитоскелета . [64] | Функции регуляции высвобождения нейромедиаторов . | |
| Синуклеин | Неизвестная клеточная функция. [65] | Считается, что он играет роль в регулировании стабильности и/или оборота плазматической мембраны . Связан как с болезнью Паркинсона , так и с болезнью Альцгеймера . | |
| GLA-домены свертывающей системы | Домены гамма-карбоксиглутамата (GLA) отвечают за высокоаффинное связывание ионов кальция. [66] | Участвует в функции факторов свертывания крови в каскаде свертывания крови. | |
| Спектрин и α- актинин -2 | Обнаружен в некоторых цитоскелета и микрофиламентов . белках [67] | Поддержание целостности плазматической мембраны и структуры цитоскелета. |
Транспортеры малых гидрофобных молекул
[ редактировать ]Эти периферические белки функционируют как переносчики неполярных соединений между различными типами клеточных мембран или между мембранами и цитозольными белковыми комплексами. Транспортируемыми веществами являются фосфатидилинозитол, токоферол, ганглиозиды, гликолипиды, производные стерола, ретинол, жирные кислоты, вода, макромолекулы, эритроциты, фосфолипиды и нуклеотиды.
- Белки-переносчики гликолипидов
- Липокалины, включая белки, связывающие ретинол и жирные кислоты . белки, связывающие
- Полиизопреноидсвязывающий белок, такой как белковый домен YceI
- Белки-активаторы ганглиозидов GM2
- Домен CRAL-TRIO (белки-переносчики альфа- токоферола и фосфатидилинозитола sec14p)
- Белки-переносчики стерола
- Белки-переносчики фосфатидилинозитола и домены STAR
- Оксистеролсвязывающий белок
Переносчики электронов
[ редактировать ]Эти белки участвуют в цепях переноса электронов . К ним относятся цитохром с , купредоксины , белок железа с высоким потенциалом , адренодоксинредуктаза, некоторые флавопротеины и другие.
Полипептидные гормоны, токсины и антимикробные пептиды
[ редактировать ]Многие гормоны, токсины , ингибиторы или антимикробные пептиды специфически взаимодействуют с трансмембранными белковыми комплексами. Они также могут накапливаться на поверхности липидного бислоя до связывания с белками-мишенями. Такие полипептидные лиганды часто заряжены положительно и электростатически взаимодействуют с анионными мембранами.
Некоторые водорастворимые белки и пептиды также могут образовывать трансмембранные каналы . Обычно они подвергаются олигомеризации , значительным конформационным изменениям и необратимо связываются с мембранами. трехмерная структура одного из таких трансмембранных каналов — α-гемолизина Определена . В других случаях экспериментальная структура представляет собой водорастворимую конформацию, периферически взаимодействующую с липидным бислоем, хотя некоторые каналообразующие пептиды достаточно гидрофобны и поэтому изучались методом ЯМР-спектроскопии в органических растворителях или в присутствии мицелл .
| Сорт | Белки | Физиология |
|---|---|---|
| Ядовитые токсины |
| Хорошо известные типы биотоксинов включают нейротоксины , цитотоксины , гемотоксины и некротоксины . Биотоксины имеют две основные функции: хищничество ( токсины змей , скорпионов и конусных улиток ) и защита ( токсины медоносных пчел и муравьев ). [68] |
| морских анемонов Токсины |
| Ингибирование натриевых и калиевых каналов и образование мембранных пор являются основными действиями более 40 известных пептидных токсинов актиний. Морские анемоны — плотоядные животные и используют токсины для хищничества и защиты; Токсин анемона по токсичности аналогичен наиболее токсичным фосфорорганическим отравляющим веществам. [69] |
| Бактериальные токсины |
| Микробные токсины являются основными факторами вирулентности для различных патогенных бактерий. Некоторые токсины представляют собой порообразующие токсины , которые лизируют клеточные мембраны. Другие токсины ингибируют синтез белка или активируют пути вторичных мессенджеров, вызывая резкие изменения в путях передачи сигнала , имеющих решающее значение для поддержания различных клеточных функций. Некоторые бактериальные токсины могут действовать непосредственно на иммунную систему , действуя как суперантигены и вызывая массивную Т-клеток пролиферацию , что приводит к чрезмерному растяжению иммунной системы. Ботулинический токсин представляет собой нейротоксин, который предотвращает стыковку/слияние нейросекреторных везикул с плазматической мембраной нервного синапса , ингибируя нейромедиатора . высвобождение [70] |
| Грибковые токсины |
| Эти пептиды характеризуются наличием необычной аминокислоты, α-аминоизомасляной кислоты , и проявляют антибиотические и противогрибковые свойства благодаря своей активности по формированию мембранных каналов. [71] |
| Антимикробные пептиды |
| Способы действия антимикробных пептидов, убивающие бактерии, разнообразны и включают разрушение мембран, нарушение метаболизма и воздействие на цитоплазматические компоненты. В отличие от многих обычных антибиотиков эти пептиды оказывают бактерицидное, а не бактериостатическое действие . |
| Дефензины |
| Дефенсины представляют собой разновидность антимикробного пептида; и являются важным компонентом практически всех врожденных защитных механизмов хозяина против микробной инвазии. Дефенсины проникают через мембраны микробных клеток посредством электрического притяжения и образуют поры в мембране, обеспечивающие отток, что в конечном итоге приводит к лизису микроорганизмов. [72] |
| Нейрональные пептиды |
| Эти белки возбуждают нейроны, вызывают поведенческие реакции, являются мощными вазодилататорами и отвечают за сокращение многих типов гладких мышц . [73] |
| апоптоза Регуляторы | Члены семейства Bcl-2 регулируют проницаемость внешней мембраны митохондрий . Bcl-2 сам по себе подавляет апоптоз в различных типах клеток, включая лимфоциты и нейрональные клетки . |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Внешний белок | биология | Британника» . www.britanica.com . Проверено 4 июля 2022 г.
- ^ Кафизо Д.С. (2005). «Структура и взаимодействие доменов C2 на поверхности мембран». В Тамме Л.К. (ред.). Белково-липидные взаимодействия: от мембранных доменов к клеточным сетям . Чичестер: Джон Уайли и сыновья. стр. 403–22. ISBN 3-527-31151-3 .
- ^ Гош М., Такер Д.Е., Берчетт С.А., Лесли CC (ноябрь 2006 г.). «Свойства семейства фосфолипаз А2 группы IV». Прогресс в исследованиях липидов . 45 (6): 487–510. дои : 10.1016/j.plipres.2006.05.003 . ПМИД 16814865 .
- ^ Jump up to: Перейти обратно: а б Джонсон Дж. Э., Корнелл Р.Б. (2002). «Амфитропные белки: регуляция посредством обратимых мембранных взаимодействий (обзор)» . Молекулярная мембранная биология . 16 (3): 217–235. дои : 10.1080/096876899294544 . ПМИД 10503244 .
- ^ Тудуппати Г.Р., Крейг Дж.В., Холоденко В., Шон А., Хилл Р.Б. (июнь 2006 г.). «Доказательства того, что вставка цитозольного домена Bcl-xL в мембрану регулируется электростатическим механизмом» . Журнал молекулярной биологии . 359 (4): 1045–1058. дои : 10.1016/j.jmb.2006.03.052 . ПМЦ 1785297 . ПМИД 16650855 .
- ^ Такида С., Ведегартнер П.Б. (июнь 2004 г.). «Независимое от экзоцитов воздействие на плазматическую мембрану гетеротримерных G-белков» . Письма ФЭБС . 567 (2–3): 209–213. doi : 10.1016/j.febslet.2004.04.062 . ПМИД 15178324 . S2CID 36940600 .
- ^ Макинтош Т.Дж., Видал А., Саймон С.А. (2003). «Энергетика пептидно-липидных взаимодействий: модификация межфазными диполями и холестерином». Актуальные темы мембран . Том. 52. Академическая пресса. стр. 205–253. ISBN 978-0-12-643871-0 .
- ^ Митра К., Убарретсена-Беландия И., Тагучи Т., Уоррен Г., Энгельман Д.М. (март 2004 г.). «Модуляция толщины бислоя мембран экзоцитарных путей мембранными белками, а не холестерином» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (12): 4083–4088. Бибкод : 2004PNAS..101.4083M . дои : 10.1073/pnas.0307332101 . ПМЦ 384699 . ПМИД 15016920 .
- ^ Марш Д. (июль 2001 г.). «Полярность и профили проникновения в липидных мембранах» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (14): 7777–7782. Бибкод : 2001PNAS...98.7777M . дои : 10.1073/pnas.131023798 . ПМК 35418 . ПМИД 11438731 .
- ^ Марш Д. (декабрь 2002 г.). «Профили мембранной водопроницаемости на спин-этикетках». Европейский биофизический журнал . 31 (7): 559–562. дои : 10.1007/s00249-002-0245-z . ПМИД 12602343 . S2CID 36212541 .
- ^ Нэгл Дж. Ф., Тристрам-Нэгл С. (ноябрь 2000 г.). «Строение липидных бислоев» . Biochimica et Biophysical Acta (BBA) — Обзоры биомембран . 1469 (3): 159–195. дои : 10.1016/S0304-4157(00)00016-2 . ПМЦ 2747654 . ПМИД 11063882 .
- ^ Гони FM (2002). «Непостоянные белки в мембранах: когда белки приходят в гости (обзор)». Молекулярная мембранная биология . 19 (4): 237–245. дои : 10.1080/0968768021000035078 . ПМИД 12512770 . S2CID 20892603 .
- ^ Гофорт Р.Л., Чи А.К., Грейтхаус Д.В., Провиденс Л.Л., Кеппе Р.Э., Андерсен О.С. (май 2003 г.). «Гидрофобная связь энергетики липидного бислоя с функцией каналов» . Журнал общей физиологии . 121 (5): 477–493. дои : 10.1085/jgp.200308797 . ПМК 2217378 . ПМИД 12719487 .
- ^ Макинтош Т.Дж., Саймон С.А. (2006). «Роль свойств бислойного материала в функции и распределении мембранных белков». Ежегодный обзор биофизики и биомолекулярной структуры . 35 (1): 177–198. doi : 10.1146/annurev.biophys.35.040405.102022 . ПМИД 16689633 .
- ^ Jump up to: Перейти обратно: а б Ханакам Ф., Гериш Г., Лотц С., Альт Т., Силиг А. (август 1996 г.). «Связывание гисактофилина I и II с липидными мембранами контролируется рН-зависимым миристоил-гистидиновым переключателем». Биохимия . 35 (34): 11036–11044. дои : 10.1021/bi960789j . ПМИД 8780505 .
- ^ Сильвиус-младший (2003). «Липидированные пептиды как инструменты для понимания мембранных взаимодействий липид-модифицированных белков». Актуальные темы мембран . Том. 52. Академическая пресса. стр. 371–395. ISBN 978-0-12-643871-0 .
- ^ Бауманн Н.А., Меннон АК (2002). «Липидные модификации белков». В Вэнсе Д.Э., Вэнсе Дж.Э. (ред.). Биохимия липидов, липопротеинов и мембран (4-е изд.). Эльзевир Наука. стр. 37–54. ISBN 978-0-444-51139-3 .
- ^ Jump up to: Перейти обратно: а б Чо В., Стахелин Р.В. (июнь 2005 г.). «Мембранно-белковые взаимодействия в клеточной передаче сигналов и мембранном транспорте». Ежегодный обзор биофизики и биомолекулярной структуры . 34 : 119–151. doi : 10.1146/annurev.biophys.33.110502.133337 . ПМИД 15869386 .
- ^ Бен-Тал Н., Хониг Б., Миллер С., Маклафлин С. (октябрь 1997 г.). «Электростатическое связывание белков с мембранами. Теоретические предсказания и экспериментальные результаты с харибдотоксином и фосфолипидными везикулами» . Биофизический журнал . 73 (4): 1717–1727. Бибкод : 1997BpJ....73.1717B . дои : 10.1016/S0006-3495(97)78203-1 . ПМК 1181073 . ПМИД 9336168 .
- ^ Шанкарам М.Б., Марш Д. (1993). «Белко-липидные взаимодействия с белками периферических мембран». В Уоттсе А. (ред.). Белково-липидные взаимодействия . Эльзевир. стр. 127–162. ISBN 0-444-81575-9 .
- ^ Мальмберг, штат Нью-Джерси, Фальке Дж. Дж. (2005). «Использование насыщения мощности ЭПР для анализа геометрии стыковки мембран периферических белков: применение к доменам C2» . Ежегодный обзор биофизики и биомолекулярной структуры . 34 (1): 71–90. doi : 10.1146/annurev.biophys.34.040204.144534 . ПМЦ 3637887 . ПМИД 15869384 .
- ^ Спенсер А.Г., Турессон Э., Отто Дж.К., Сонг И., Смит Т., ДеВитт Д.Л. и др. (ноябрь 1999 г.). «Мембраносвязывающие домены простагландин-эндопероксид-H-синтаз 1 и 2. Пептидное картирование и мутационный анализ» . Журнал биологической химии . 274 (46): 32936–32942. дои : 10.1074/jbc.274.46.32936 . ПМИД 10551860 .
- ^ Латроп Б., Гэдд М., Билтонен Р.Л., Правило GS (март 2001 г.). «Изменения сродства к Ca2+ при активации фосфолипазы A2 Agkistrodon piscivorus piscivorus». Биохимия . 40 (11): 3264–3272. дои : 10.1021/bi001901n . ПМИД 11258945 .
- ^ Кутателадзе Т., Овердуин М. (март 2001 г.). «Структурный механизм стыковки эндосом доменом FYVE». Наука . 291 (5509): 1793–1796. Бибкод : 2001Sci...291.1793K . дои : 10.1126/science.291.5509.1793 . ПМИД 11230696 .
- ^ Татулиан С.А., Цинь С., Панде А.Х., Хе Х (сентябрь 2005 г.). «Позиционирование мембранных белков с помощью новых белковой инженерии и биофизических подходов» . Журнал молекулярной биологии . 351 (5): 939–947. дои : 10.1016/j.jmb.2005.06.080 . ПМИД 16055150 .
- ^ Jump up to: Перейти обратно: а б Христова К., Уимли В.К., Мишра В.К., Анантарамия ГМ, Сегрест Дж.П., Уайт С.Х. (июль 1999 г.). «Амфипатическая альфа-спираль на границе раздела мембран: структурное исследование с использованием нового метода дифракции рентгеновских лучей». Журнал молекулярной биологии . 290 (1): 99–117. дои : 10.1006/jmbi.1999.2840 . ПМИД 10388560 .
- ^ Мюррей Д., Хониг Б. (январь 2002 г.). «Электростатический контроль мембранного нацеливания доменов C2» . Молекулярная клетка . 9 (1): 145–154. дои : 10.1016/S1097-2765(01)00426-9 . ПМИД 11804593 .
- ^ Ефремов Р.Г., Нольде Д.Е., Коншина А.Г., Сырцев Н.П., Арсеньев А.С. (сентябрь 2004 г.). «Пептиды и белки в мембранах: чему мы можем научиться с помощью компьютерного моделирования?». Современная медицинская химия . 11 (18): 2421–2442. дои : 10.2174/0929867043364496 . ПМИД 15379706 .
- ^ Ломизе А.Л., Погожева И.Д., Ломизе М.А., Мосберг Х.И. (июнь 2006 г.). «Позиционирование белков в мембранах: вычислительный подход» . Белковая наука . 15 (6): 1318–1333. дои : 10.1110/ps.062126106 . ПМК 2242528 . ПМИД 16731967 .
- ^ Ломизе А., Ломизе М., Погожева И. «Сравнение с экспериментальными данными» . Ориентации белков в мембранах . Мичиганский университет . Проверено 8 февраля 2007 г.
- ^ Папахаджопулос Д., Москарелло М., Эйлар Э.Х., Исак Т. (сентябрь 1975 г.). «Влияние белков на термотропные фазовые переходы фосфолипидных мембран». Biochimica et Biophysical Acta (BBA) – Биомембраны . 401 (3): 317–335. дои : 10.1016/0005-2736(75)90233-3 . ПМИД 52374 .
- ^ Силиг Дж. (ноябрь 2004 г.). «Термодинамика липид-пептидных взаимодействий» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1666 (1–2): 40–50. дои : 10.1016/j.bbamem.2004.08.004 . ПМИД 15519307 .
- ^ Даркс М.Дж., Дэвис С.М., Брэдшоу Дж.П. (1997). «Взаимодействие тахикининов с фосфолипидными мембранами: нейтронографическое исследование». Физика Б. 241 : 1144–1147. Бибкод : 1997PhyB..241.1144D . дои : 10.1016/S0921-4526(97)00811-9 .
- ^ Эллена Дж. Ф., Моултроп Дж., Ву Дж., Раух М., Джейсингхне С., Касл Дж. Д., Кафизо Д. С. (ноябрь 2004 г.). «Мембранное положение основного ароматического пептида, который изолирует фосфатидилинозитол-4,5-бисфосфат, определенное с помощью сайт-направленного спинового мечения и ЯМР высокого разрешения» . Биофизический журнал . 87 (5): 3221–3233. Бибкод : 2004BpJ....87.3221E . дои : 10.1529/biophysj.104.046748 . ПМЦ 1304792 . ПМИД 15315949 .
- ^ Маркотт I, Дюфур Э.Дж., Уэлле М., Оже М. (июль 2003 г.). «Взаимодействие нейропептида мет-энкефалина с цвиттер-ионными и отрицательно заряженными бицеллами, как видно с помощью твердотельного ЯМР 31P и 2H» . Биофизический журнал . 85 (1): 328–339. Бибкод : 2003BpJ....85..328M . дои : 10.1016/S0006-3495(03)74477-4 . ПМК 1303088 . ПМИД 12829487 .
- ^ Чжан В., Крокер Э., Маклафлин С., Смит С.О. (июнь 2003 г.). «Связывание пептидов с основными и ароматическими остатками с бислойными мембранами: фенилаланин в эффекторном домене миристоилированного, богатого аланином субстрата C-киназы, проникает в гидрофобное ядро бислоя» . Журнал биологической химии . 278 (24): 21459–21466. дои : 10.1074/jbc.M301652200 . ПМИД 12670959 .
- ^ «Pfam-запись Абгидролаза 1» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ Бауэр, Табеа Л.; Бухгольц, Патрик К.Ф.; Плейсс, Юрген (март 2020 г.). «Модульная структура α/β-гидролаз» . Журнал ФЭБС . 287 (5): 1035–1053. дои : 10.1111/февраль 15071 . ISSN 1742-464X .
- ^ «Запись Pfam: фосфолипаза А2» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ Казале, Джаретт; Качими, Салах Эддин О.; Варакалло, Мэтью (2023), «Биохимия, фосфолипаза A2» , StatPearls , Остров сокровищ (Флорида): StatPearls Publishing, PMID 30521272 , получено 29 ноября 2023 г.
- ^ «Запись Pfam: фосфатидилинозитол-специфическая фосфолипаза C, домен X» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ «Фосфолипаза C» , Arc.Ask3.Ru , 16 августа 2023 г. , получено 29 ноября 2023 г.
- ^ «Запись Pfam: холестериноксидаза» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ Юэ, К. Кимберли; Касс, Игнатиус Дж.; Сэмпсон, Николь С.; Врилинк, Алиса (1 апреля 1999 г.). «Определение кристаллической структуры холестериноксидазы из Streptomyces и структурная характеристика мутантов ключевых активных сайтов» . Биохимия . 38 (14): 4277–4286. дои : 10.1021/bi982497j . ISSN 0006-2960 .
- ^ «Запись Pfam: белок мембраны пигментного эпителия сетчатки» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ «Каротиноидоксигеназа» , Arc.Ask3.Ru , 29 ноября 2023 г. , получено 29 ноября 2023 г.
- ^ «Запись Pfam: липоксигеназа» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ Пригге, Северная Каролина; Бойингтон, Джей Си; Фаиг, М.; Доктор, КС; Гаффни, Би Джей; Амзель, Л.М. (1 ноября 1997 г.). «Строение и механизм липоксигеназ» . Биохимия . 79 (11): 629–636. дои : 10.1016/S0300-9084(97)83495-5 . ISSN 0300-9084 .
- ^ Запись PDBsum: Альфа-токсин
- ^ «Альфа-токсин — обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 29 ноября 2023 г.
- ^ «Запись Pfam: фосфодиэстераза типа I» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ Сюн, Цзы-Цзянь; Хуан, Цзинцзин; Пода, Геннадий; Помес, Режис; Приве, Гилберт Г. (31 июля 2016 г.). «Структура кислой сфингомиелиназы человека раскрывает роль домена сапозина в активации гидролиза субстрата» . Журнал молекулярной биологии . 428 (15): 3026–3042. дои : 10.1016/j.jmb.2016.06.012 . ISSN 0022-2836 .
- ^ «Запись Pfam: группа гликозилтрансфераз 1» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ Юнлигил, Улуг М; Рини, Джеймс М. (01 октября 2000 г.). «Структура и механизм гликозилтрансферазы» . Современное мнение в области структурной биологии . 10 (5): 510–517. дои : 10.1016/S0959-440X(00)00124-X . ISSN 0959-440X .
- ^ «Запись Pfam: Феррохелатаза» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ Аль-Карадаги, Салам; Ханссон, Матс; Никонов Станислав; Йонссон, Бодил; Хедерстедт, Ларс (ноябрь 1997 г.). «Кристаллическая структура феррохелатазы: терминальный фермент биосинтеза гема» . Структура . 5 (11): 1501–1510. дои : 10.1016/s0969-2126(97)00299-2 . ISSN 0969-2126 .
- ^ «Запись Pfam: Связанный с миотубулярином» . Архивировано из оригинала 26 сентября 2007 г. Проверено 25 января 2007 г.
- ^ «Принципы и практика медицинской генетики Эмери и Римоэна» . НаукаДирект . Проверено 29 ноября 2023 г.
- ^ «Запись Pfam: Дигидрооротатдегидрогеназа» . Архивировано из оригинала 26 сентября 2007 г. Проверено 25 января 2007 г.
- ^ Лю, Шэньпин; Нейдхардт, Эди А; Гроссман, Труди Х; Окаин, Тим; Кларди, Джон (январь 2000 г.). «Структуры дигидрооротатдегидрогеназы человека в комплексе с антипролиферативными средствами» . Структура . 8 (1): 25–33. дои : 10.1016/s0969-2126(00)00077-0 . ISSN 0969-2126 .
- ^ «Запись Pfam: FMN-зависимая дегидрогеназа» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ «Гликолатоксидаза — Протеопедия, жизнь в 3D» . сайт proteopedia.org . Проверено 28 ноября 2023 г.
- ^ «Запись Pfam: Аннексин» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ «Запись Pfam Синапсин Н» . Архивировано из оригинала 26 сентября 2007 г. Проверено 25 января 2007 г.
- ^ «Пфам-вход Синуклеин» . Архивировано из оригинала 26 сентября 2007 г. Проверено 25 января 2007 г.
- ^ «Запись в Pfam: Gla» . Архивировано из оригинала 29 сентября 2007 г. Проверено 25 января 2007 г.
- ^ «Вход Pfam Спектрин» . Архивировано из оригинала 26 сентября 2007 г. Проверено 25 января 2007 г.
- ^ Роша Х, Мартин-Оклер М.Ф., ред. (2000). Животные токсины: факты и протоколы . Базель: Биркхузер Верлаг. ISBN 3-7643-6020-8 .
- ^ Паточка Дж., Струнечка А. (1999). «Токсины морских анемонов» . Информационный бюллетень АСА . Архивировано из оригинала 15 июня 2013 года.
- ^ Шмитт К.К., Мейсик К.К., О'Брайен А.Д. (1999). «Бактериальные токсины: друзья или враги?» . Новые инфекционные заболевания . 5 (2): 224–234. дои : 10.3201/eid0502.990206 . ПМК 2640701 . ПМИД 10221874 .
- ^ Чу Дж. К., Уоллес Б. А. (август 2001 г.). «Пептайболы: модели ионных каналов». Труды Биохимического общества . 29 (Часть 4): 565–570. дои : 10.1042/BST0290565 . ПМИД 11498029 .
- ^ Оппенгейм Дж. Дж., Бирагин А., Квак Л.В., Ян Д. (ноябрь 2003 г.). «Роль противомикробных пептидов, таких как дефензины, во врожденном и адаптивном иммунитете» . Анналы ревматических болезней . 62 (Приложение 2): ii17–ii21. дои : 10.1136/ard.62.suppl_2.ii17 . ПМК 1766745 . ПМИД 14532141 .
- ^ «Пфам вход Тахикинин» . Архивировано из оригинала 26 сентября 2007 г. Проверено 25 января 2007 г.
Дальнейшее чтение
[ редактировать ]- Тамм Л.К., изд. (2005). Белково-липидные взаимодействия: от мембранных доменов к клеточным сетям . Чичестер: Джон Уайли и сыновья. ISBN 3-527-31151-3 .
- Чо В., Стахелин Р.В. (июнь 2005 г.). «Мембранно-белковые взаимодействия в клеточной передаче сигналов и мембранном транспорте». Ежегодный обзор биофизики и биомолекулярной структуры . 34 (1): 119–151. doi : 10.1146/annurev.biophys.33.110502.133337 . ПМИД 15869386 .
- Гони FM (2002). «Непостоянные белки в мембранах: когда белки приходят в гости (обзор)». Молекулярная мембранная биология . 19 (4): 237–245. дои : 10.1080/0968768021000035078 . ПМИД 12512770 . S2CID 20892603 .
- Джонсон Дж. Э., Корнелл Р.Б. (1999). «Амфитропные белки: регуляция посредством обратимых мембранных взаимодействий (обзор)» . Молекулярная мембранная биология . 16 (3): 217–235. дои : 10.1080/096876899294544 . ПМИД 10503244 .
- Ситон Б.А., Робертс М.Ф. (1996). «Периферические мембранные белки». В Мерц К., Ру Б. (ред.). Биологические мембраны . Бостон, Массачусетс: Биркхаузер. стр. 355–403.
- Бенга Дж. (1985). «Белко-липидные взаимодействия в биологических мембранах». В Бенге Дж. (ред.). Строение и свойства биологических мембран . Том. 1. Бока-Ратон, Флорида: CRC Press. стр. 159–188.
- Кессель А., Бен-Тал Н. (январь 2002 г.). «Детерминанты свободной энергии ассоциации пептидов с липидными бислоями». Актуальные темы мембран . 52 : 205–253. дои : 10.1016/S1063-5823(02)52010-X . ISBN 9780121533526 .
- Мальмберг, штат Нью-Джерси, Фальке Дж. Дж. (2005). «Использование насыщения мощности ЭПР для анализа геометрии стыковки мембран периферических белков: применение к доменам C2» . Ежегодный обзор биофизики и биомолекулярной структуры . 34 (1): 71–90. doi : 10.1146/annurev.biophys.34.040204.144534 . ПМЦ 3637887 . ПМИД 15869384 .
- Макинтош Т.Дж., Саймон С.А. (2006). «Роль свойств бислойного материала в функции и распределении мембранных белков». Ежегодный обзор биофизики и биомолекулярной структуры . 35 (1): 177–198. doi : 10.1146/annurev.biophys.35.040405.102022 . ПМИД 16689633 .
Внешние ссылки
[ редактировать ]- Белки периферической мембраны в базе данных OPM
- DOLOP Genomics. База данных бактериальных липопротеинов, ориентированная на
- База данных пептайбола
- База данных по антимикробным пептидам, заархивированная 20 июля 2011 г. в Wayback Machine.