Сфингомиелин фосфодиэстераза
| Сфингомиелин фосфодиэстераза | |||
|---|---|---|---|
 Кристаллическая структура сфингомиелиназы Bacillus cereus . [ 1 ] | |||
| Идентификаторы | |||
| Номер ЕС. | 3.1.4.12 | ||
| Номер CAS. | 9031-54-3 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
Сфингомиелинфосфодиэстераза (EC 3.1.4.12, также известная как нейтральная сфингомиелиназа , сфингомиелиназа или SMase ; систематическое название сфингомиелинхолинфосфогидролаза ) представляет собой фермент гидролазу , который участвует в реакциях метаболизма сфинголипидов . SMase является членом суперсемейства ферментов ДНКазы I и отвечает за расщепление сфингомиелина (SM) на фосфохолин и церамид . Активация SMase была предложена как основной путь производства церамидов в ответ на клеточный стресс. [ 2 ]
Семейство сфингомиелиназ
[ редактировать ]Идентифицировано пять типов SMase. Они классифицируются в зависимости от катионной зависимости и оптимального значения pH и включают:
- Лизосомальная кислота SMase
- Секретируемая цинкзависимая кислота SMase
- Магнийзависимая нейтральная SMase
- Независимая от магния нейтральная SMase
- Щелочная SMase
Из них лизосомальная кислая SMase и магнийзависимая нейтральная SMase считаются основными кандидатами на выработку церамидов в клеточном ответе на стресс.
Нейтральная сфингомиелиназа
[ редактировать ]Активность нейтральной сфингомиелиназы (N-SMase) была впервые описана в фибробластах пациентов с болезнью Нимана-Пика – лизосомальной болезнью накопления, характеризующейся дефицитом кислой SMase. [ 3 ] Последующее исследование показало, что этот фермент был продуктом отдельного гена, имел оптимальный pH 7,4 и зависел от магния. 2+ ионы для активности и особенно обогащены мозгом. [ 4 ] Однако более недавнее исследование мозга крупного рогатого скота показало существование множества изоформ N-SMase с различными биохимическими и хроматографическими свойствами. [ 5 ]
Крупный прорыв произошел в середине 1980-х годов с клонированием первых N-SMase из Bacillus cereus и Staphylococcus aureus . [ 6 ] [ 7 ] Использование последовательностей этих бактериальных сфингомиелиназ в поиске гомологии в конечном итоге привело к идентификации дрожжевых N-SMase ISC1 в почкующихся дрожжах Saccharomyces cerevisiae . [ 8 ] и ферменты N-SMase млекопитающих, nSMase1 и nSMase2. [ 9 ] [ 10 ] Идентичность между SMase млекопитающих, дрожжей и бактерий очень низкая - она составляет примерно 20% между nSMase2 и SMase B. cereus. Однако выравнивание последовательностей (см. рисунок) указывает на наличие ряда консервативных остатков во всем семействе, особенно в каталитической области ферментов. [ 11 ] Это привело к предположению об общем каталитическом механизме семейства N-SMase.
Третий белок N-SMase, названный nSMase3 , был клонирован и охарактеризован в 2006 году. [ 12 ] nSMase3 имеет небольшое сходство последовательностей с nSMase1 или nSMase2. Однако, по-видимому, существует высокая степень эволюционной консервативности от низших к высшим организмам, что позволяет предположить, что он может содержать уникальную и отличную N-SMase. Высокая экспрессия nSMase3 в сердце и скелетных мышцах также предполагает потенциальную роль в функции сердца. [ 13 ]
Активный сайт
[ редактировать ]
Решение кристаллической структуры нейтральной сфингомиелиназы Listeria ivanovii и Bacillus cereus позволило более полно понять их ферментативный сайт. Активный центр содержит SMазы B. cereus остатки Asn -16, Glu -53, Asp -195, Asn-197 и His -296. Известно, что из них остатки Glu-53, Asp-195 и His-296 необходимы для активности. Относительная каталитическая активность SMase при связывании ионов металлов с активным центром была изучена для ионов двухвалентных металлов Co. 2+ , Мн 2+ , мг 2+ , Как 2+ и сэр 2+ . Из этих пяти ионов металлов Co 2+ , Мн 2+ и мг 2+ связывание с активным центром приводит к высокой каталитической активности SMase. Калифорния 2+ и старший 2+ связанные с активным центром, проявляют гораздо более низкую каталитическую активность SMase. Когда один мг 2+ ион или два Co 2+ ионы связываются с активным центром, в результате получается двойная гексакоординированная геометрия с двумя октаэдрическими бипирамидами для Co 2+ и одна октаэдрическая бипирамида для Mg 2+ . Когда один Ка 2+ Ион связывается с активным центром, в результате чего образуется гепта-координированная геометрия. Таким образом, предполагается, что разница в каталитической активности ионов металлов обусловлена геометрическими различиями. Ко 2+ и мг 2+ , SMase имеет лучшую реакционную способность, когда два Co 2+ ионы связаны с SMase; когда эти Ко 2+ Ионы связываются, Glu-53 и His-296 связывают по одному катиону двухвалентного металла. Эти катионы окружены мостиковыми молекулами воды и действуют как кислоты Льюиса . [ 1 ]
Механизм
[ редактировать ]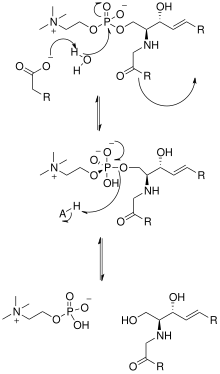
Решение кристаллической структуры нейтральной сфингомиелиназы Listeria ivanovii и Bacillus cereus также пролило свет на их каталитические механизмы. Активный центр SMase содержит остатки Glu и His, каждый из которых связан с одним или двумя катионами двухвалентных металлов, обычно Co. 2+ , мг 2+ , или Ка 2+ для оптимальной производительности. Эти два катиона участвуют в катализе, привлекая SM к активному центру SMase. Двухвалентный катион, связанный с остатком Glu, взаимодействует с амидокислородом и сложноэфирным кислородом между С1 и фосфатной группой СМ; Остаток Asn и катион двухвалентного металла, связанный с остатком His, связываются с атомами кислорода фосфатной группы СМ. Это стабилизирует отрицательный заряд фосфатной группы. Катион металла, связанный с остатком His и боковыми цепями Asp и Asn, снижает значение pKa одной из мостиковых молекул воды, тем самым активируя молекулу воды. Эта молекула воды затем действует как нуклеофил и атакует фосфатную группу SM, создавая пятивалентный атом фосфора, отрицательный заряд которого стабилизируется катионами двухвалентных металлов. Затем фосфат преобразует свою тетраэдрическую конформацию и приводит к образованию продуктов церамида и фосфохолина . [ 1 ] В 2016 году была предложена модель, основанная на исследовании кристаллической структуры кислой сфингомиелиназы млекопитающих, согласно которой ASMase существует в равновесии между открытой и закрытой формами домена сапозина . В отсутствие мембран закрытый ASMasesap, отделенный от ASMasecat, будет преобладать и делать фермент неактивным. В присутствии анионных мембран открытый ASMasesap становится преобладающим, прикрепляется к поверхности мембраны и одновременно образует интерфейс с каталитическим доменом, активируя его для сфингомиелина гидролиза . [ 14 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с ВВП : 2ддт ; Аго Х., Ода М., Такахаси М., Цугэ Х., Очи С., Катунума Н., Мияно М., Сакураи Дж. (июнь 2006 г.). «Структурные основы активности сфингомиелинфосфодиэстеразы нейтральной сфингомиелиназы Bacillus cereus» . Ж. Биол. Хим . 281 (23): 16157–67. дои : 10.1074/jbc.M601089200 . ПМИД 16595670 .
- ^ Ханнун Ю.А., Обейд Л.М. (июль 2002 г.). «Церамидоцентричная вселенная липид-опосредованной клеточной регуляции: стрессовые ситуации липидного типа» . Ж. Биол. Хим . 277 (29): 25847–50. дои : 10.1074/jbc.R200008200 . ПМИД 12011103 .
- ^ Шнайдер П.Б., Кеннеди Е.П. (май 1967 г.). «Сфингомиелиназа в селезенке нормального человека и в селезенке пациентов с болезнью Нимана-Пика» . Дж. Липид Рес . 8 (3): 202–9. дои : 10.1016/S0022-2275(20)40138-5 . ПМИД 4962590 .
- ^ Рао Б.Г., Спенс М.В. (сентябрь 1976 г.). «Активность сфингомиелиназы при pH 7,4 в мозге человека и сравнение с активностью при pH 5,0» . Дж. Липид Рес . 17 (5): 506–15. дои : 10.1016/S0022-2275(20)41749-3 . ПМИД 9463 .
- ^ Юнг С.Ю., Су Дж.Х., Пак Х.Дж., Юнг К.М., Ким М.Ю., На Д.С., Ким Д.К. (сентябрь 2000 г.). «Идентификация множественных форм мембранассоциированной нейтральной сфингомиелиназы в мозге крупного рогатого скота» . Дж. Нейрохем . 75 (3): 1004–14. дои : 10.1046/j.1471-4159.2000.0751004.x . ПМИД 10936181 . S2CID 46397368 .
- ^ Коулман, округ Колумбия, Арбутнотт Дж.П., Помрой Х.М., Биркбек Т.Х. (декабрь 1986 г.). «Клонирование и экспрессия в Escherichia coli и Staphylococcus aureus детерминанты бета-лизина из Staphylococcus aureus: доказательства того, что преобразование бактериофагом активности бета-лизина вызвано инсерционной инактивацией детерминанты бета-лизина». Микроб. Патог . 1 (6): 549–64. дои : 10.1016/0882-4010(86)90040-9 . ПМИД 3334158 .
- ^ Ямада А., Цукагоси Н., Удака С., Сасаки Т., Макино С., Накамура С., Литтл С., Томита М., Икезава Х. (август 1988 г.). «Нуклеотидная последовательность и экспрессия в Escherichia coli гена, кодирующего сфингомиелиназу Bacillus cereus». Евро. Дж. Биохим . 175 (2): 213–20. дои : 10.1111/j.1432-1033.1988.tb14186.x . ПМИД 2841128 .
- ^ Савай Х., Окамото Ю., Луберто С., Мао С., Белявска А., Домае Н., Ханнун Ю.А. (декабрь 2000 г.). «Идентификация ISC1 (YER019w) как инозитолфосфинголипидфосфолипазы C в Saccharomyces cerevisiae» . Ж. Биол. Хим . 275 (50): 39793–8. дои : 10.1074/jbc.M007721200 . ПМИД 11006294 .
- ^ Томюк С., Хофманн К., Никс М., Зумбансен М., Стоффель В. (март 1998 г.). «Клонированная нейтральная сфингомиелиназа млекопитающих: функции в передаче сигналов сфинголипидов?» . Учеб. Натл. акад. наук. США . 95 (7): 3638–43. Бибкод : 1998PNAS...95.3638T . дои : 10.1073/pnas.95.7.3638 . ЧВК 19888 . ПМИД 9520418 .
- ^ Томюк С., Зумбансен М., Стоффель В. (февраль 2000 г.). «Характеристика и субклеточная локализация магнийзависимой нейтральной сфингомиелиназы мыши и человека» . Ж. Биол. Хим . 275 (8): 5710–7. дои : 10.1074/jbc.275.8.5710 . ПМИД 10681556 .
- ^ Кларк С.Дж., Снук С.Ф., Тани М., Матмати Н., Маркезини Н., Ханнун Ю.А. (сентябрь 2006 г.). «Расширенное семейство нейтральных сфингомиелиназ». Биохимия . 45 (38): 11247–56. дои : 10.1021/bi061307z . ПМИД 16981685 .
- ^ Круть, Олег; Вигманн, Катя; Кашкар, Хамид; Язданпана, Бенджамин; Крёнке, Мартин (12 мая 2006 г.). «Новая нейтральная сфингомиелиназа-3 млекопитающих, реагирующая на фактор некроза опухоли, представляет собой белок, заякоренный в С-хвосте» . Журнал биологической химии . 281 (19): 13784–13793. дои : 10.1074/jbc.M511306200 . ISSN 0021-9258 . ПМИД 16517606 .
- ^ Крут О, Вигманн К, Кашкар Х, Язданпана Б, Крёнке М (май 2006 г.). «Новая нейтральная сфингомиелиназа-3 млекопитающих, реагирующая на фактор некроза опухоли, представляет собой белок, заякоренный в С-хвосте» . Ж. Биол. Хим . 281 (19): 13784–93. дои : 10.1074/jbc.M511306200 . ПМИД 16517606 .
- ^ Горелик, Алексей; Иллес, Каталин; Хайнц, Леонард X.; Суперти-Фурга, Джулио; Нагар, Бхушан (20 июля 2016 г.). «Кристаллическая структура кислой сфингомиелиназы млекопитающих» . Природные коммуникации . 7 (1): 12196. doi : 10.1038/ncomms12196 . ISSN 2041-1723 . ПМЦ 4961792 . ПМИД 27435900 . S2CID 32841772 .
Дальнейшее чтение
[ редактировать ]Внешние ссылки
[ редактировать ]- Сфингомиелин + фосфодиэстераза в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)