Расхождение хромосом
Сегрегация хромосом — процесс у эукариот, при котором две сестринские хроматиды, образовавшиеся в результате репликации ДНК , или парные гомологичные хромосомы , отделяются друг от друга и мигрируют к противоположным полюсам ядра . Этот процесс сегрегации происходит как во время митоза , так и во время мейоза . Расщепление хромосом происходит и у прокариот . Однако, в отличие от сегрегации хромосом эукариот, репликация и сегрегация не разделены во времени. Вместо этого сегрегация происходит постепенно после репликации. [1]
Митотическая сегрегация хроматид
[ редактировать ]
Во время митоза сегрегация хромосом происходит обычно как этап деления клетки (см. диаграмму митоза). Как показано на диаграмме митоза, митозу предшествует цикл репликации ДНК, так что каждая хромосома образует две копии, называемые хроматидами . Эти хроматиды расходятся к противоположным полюсам, этому процессу способствует белковый комплекс, называемый когезином . При правильном разделении полный набор хроматид оказывается в каждом из двух ядер, а когда деление клетки завершается, каждая копия ДНК, ранее называвшаяся хроматидой, теперь называется хромосомой.
Мейотическая хромосома и сегрегация хроматид
[ редактировать ]Сегрегация хромосом происходит на двух отдельных стадиях мейоза, называемых анафазой I и анафазой II (см. диаграмму мейоза). В диплоидной клетке имеется два набора гомологичных хромосом различного родительского происхождения (например, отцовский и материнский набор). Во время фазы мейоза, обозначенной на диаграмме мейоза как «интерфаза s», происходит цикл репликации ДНК, так что каждая из первоначально присутствующих хромосом теперь состоит из двух копий, называемых хроматидами . Эти хромосомы (парные хроматиды) затем спариваются с гомологичной хромосомой (также парными хроматидами), присутствующими в том же ядре (см. профазу I на диаграмме мейоза). Процесс выравнивания парных гомологичных хромосом называется синапсисом (см. Синапсис ). Во время синапса обычно происходит генетическая рекомбинация. Некоторые из событий рекомбинации происходят путем кроссинговера (включающего физический обмен между двумя хроматидами), но большинство событий рекомбинации включают обмен информацией, а не физический обмен между двумя хроматидами (см. Синтез-зависимый отжиг цепи (SDSA) ). После рекомбинации происходит сегрегация хромосом, на что указывают стадии метафазы I и анафазы I на диаграмме мейоза.
Различные пары хромосом разделяются независимо друг от друга, этот процесс получил название «независимый набор негомологичных хромосом» . В результате этого процесса каждая гамета обычно содержит смесь хромосом от обоих исходных родителей.
Неправильная сегрегация хромосом (см. нерасхождение , дисомия ) может привести к тому, что анеуплоидные гаметы будут иметь либо слишком мало, либо слишком много хромосом.
Вторая стадия, на которой происходит сегрегация во время мейоза, - это профаза II (см. схему мейоза). На этой стадии сегрегация происходит по тому же принципу, что и при митозе, за исключением того, что в этом случае профазе II не предшествует цикл репликации ДНК. Таким образом, две хроматиды, составляющие каждую хромосому, разделяются на разные ядра , так что каждое ядро получает один набор хроматид (теперь называемых хромосомами), и каждое ядро включается в гаплоидную гамету (см. стадии после профазы II на диаграмме мейоза). Этот процесс сегрегации также облегчается когезином . Нарушение правильной сегрегации во время профазы II также может привести к образованию анеуплоидных гамет. Анеуплоидные гаметы могут подвергнуться оплодотворению с образованием анеуплоидных зигот, что может привести к серьезным неблагоприятным последствиям для потомства.
Кроссоверы облегчают сегрегацию, но не являются обязательными.
[ редактировать ]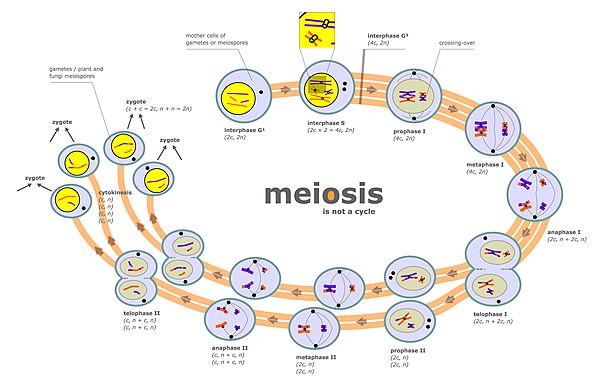

Мейотическая хромосомная рекомбинация (СО) облегчает правильное разделение гомологичных хромосом . Это связано с тем, что в конце профазы I мейоза рекомбинация CO обеспечивает физическую связь, которая удерживает вместе гомологичные пары хромосом. Эти связи устанавливаются хиазмами , которые являются цитологическими проявлениями рекомбинации CO. Вместе с когезионной связью между сестринскими хроматидами рекомбинация CO может помочь обеспечить упорядоченное разделение парных гомологичных хромосом к противоположным полюсам. В подтверждение этого исследование анеуплоидии отдельных сперматозоидов с помощью полногеномного секвенирования показало, что в среднем человеческие сперматозоиды с анеуплоидными аутосомами демонстрируют значительно меньшее количество кроссинговеров, чем нормальные клетки. [2] После завершения первого разделения хромосом в мейозе I происходит дальнейшее разделение хромосом во время второго эквационного деления мейоза II . Как правильная начальная сегрегация хромосом в профазе I, так и следующая сегрегация хромосом во время эквационного деления в мейозе II необходимы для образования гамет с правильным количеством хромосом.
Рекомбинанты CO производятся в результате процесса, включающего образование и разделение промежуточных продуктов соединения Холлидея . Как показано на рисунке под названием «Современная модель мейотической рекомбинации», образование мейотических кроссинговеров может быть инициировано двухцепочечным разрывом (DSB). Для введения DSB в ДНК часто используется топоизомеразоподобный белок SPO11. [3] Рекомбинация CO также может быть инициирована внешними источниками повреждения ДНК, такими как рентгеновское облучение, [4] или внутренние источники. [5] [6]
Есть доказательства того, что рекомбинация CO облегчает мейотическую сегрегацию хромосом. [2] Другие исследования, однако, показывают, что хиазма , хотя и поддерживает, но не важна для мейотической сегрегации хромосом. Почкующиеся дрожжи Saccharomyces cerevisiae являются модельным организмом, используемым для изучения мейотической рекомбинации. Было обнаружено, что мутанты S. cerevisiae , дефектные в рекомбинации CO на уровне разрешения соединения Холлидея, эффективно подвергаются правильной сегрегации хромосом. Путь, который производит большую часть CO у S. cerevisiae и, возможно, у млекопитающих, включает комплекс белков, включая MLH1 - MLH3 гетеродимер (называемый MutL gamma). [7] MLH1-MLH3 преимущественно связывается с соединениями Холлидея. [8] Это эндонуклеаза , которая делает одноцепочечные разрывы в суперскрученной двухцепочечной ДНК. [8] [9] и способствует образованию рекомбинантов CO. [10] Двойные мутанты, удаленные как для MLH3 (основной путь), так и для MMS4 (который необходим для второстепенного пути разрешения соединения Холлидея), показали резкое снижение кроссинговера по сравнению с диким типом (снижение в 6-17 раз); однако жизнеспособность спор была достаточно высокой (62%), а расхождение хромосом оказалось в основном функциональным. [10]
Белки MSH4 и MSH5 образуют гетероолигомерную структуру ( гетеродимер ) у S. cerevisiae и человека. [11] [12] [13] У S. cerevisiae MSH4 и MSH5 действуют специфически, облегчая кроссинговеры между гомологичными хромосомами во время мейоза. [11] Комплекс MSH4/MSH5 связывает и стабилизирует двойные соединения Холлидея и способствует их расщеплению в продукты кроссовера. Гипоморфный (частично функциональный) мутант MSH4 S. cerevisiae показал 30%-ное снижение числа кроссинговеров по всему геному и большое количество мейозов с необменными хромосомами. [14] Тем не менее, этот мутант дал образцы жизнеспособности спор, что позволяет предположить, что сегрегация необменных хромосом происходит эффективно. [14] Таким образом, оказывается, что рекомбинация CO облегчает правильную сегрегацию хромосом во время мейоза у S. cerevisiae , но это не существенно.
Делящиеся дрожжи Schizosaccharomyces pombe обладают способностью сегрегировать гомологичные хромосомы в отсутствие мейотической рекомбинации (ахиазматная сегрегация). [15] микротрубочек Эта способность зависит от моторного динеина , регулирующего движение хромосом к полюсам мейотического веретена .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Нильсен, Х.Дж.; Янгрен, Б.; Хансен, ФГ; Остин, С. (1 декабря 2007 г.). «Динамика разделения хромосом Escherichia coli во время мультивилочной репликации» . Журнал бактериологии . 189 (23): 8660–8666. дои : 10.1128/JB.01212-07 . ISSN 0021-9193 . ПМК 2168957 . ПМИД 17905986 .
- ^ Jump up to: а б Лу С, Цзун С, Фань В, Ян М, Ли Дж, Чепмен А.Р., Чжу П, Ху Х, Сюй Л, Ян Л, Бай Ф, Цяо Дж, Тан Ф, Ли Р, Се XS (2012). «Изучение мейотической рекомбинации и анеуплоидии отдельных сперматозоидов с помощью полногеномного секвенирования» . Наука . 338 (6114): 1627–30. Бибкод : 2012Sci...338.1627L . дои : 10.1126/science.1229112 . ПМК 3590491 . ПМИД 23258895 .
- ^ Сансам КЛ, Пецца Р.Дж. (2015). «Соединение путем разрыва и восстановления: механизмы обмена цепей ДНК при мейотической рекомбинации» . ФЕБС Дж . 282 (13): 2444–57. дои : 10.1111/февраль 13317 . ПМЦ 4573575 . ПМИД 25953379 .
- ^ Дернбург А.Ф., Макдональд К., Молдер Г., Барстед Р., Дрессер М., Вильнёв А.М. (1998). «Мейотическая рекомбинация у C. elegans инициируется консервативным механизмом и не является обязательной для синапса гомологичных хромосом» . Клетка . 94 (3): 387–98. дои : 10.1016/s0092-8674(00)81481-6 . ПМИД 9708740 . S2CID 10198891 .
- ^ Фара Дж.А., Кроми Дж., Дэвис Л., Штайнер В.В., Смит Г.Р. (2005). «Активация альтернативного, независимого от Rec12 (spo11) пути мейотической рекомбинации делящихся дрожжей в отсутствие эндонуклеазы лоскута ДНК» . Генетика . 171 (4): 1499–511. doi : 10.1534/genetics.105.046821 . ПМК 1456079 . ПМИД 16118186 .
- ^ Пауклин С., Буркерт Дж.С., Мартин Дж., Осман Ф., Веллер С., Бултон С.Дж., Уитби М.К., Петерсен-Март С.К. (2009). «Альтернативная индукция мейотической рекомбинации от одноосновных повреждений ДНК-дезаминаз» . Генетика . 182 (1): 41–54. дои : 10.1534/genetics.109.101683 . ПМЦ 2674839 . ПМИД 19237686 .
- ^ Захарьевич К, Тан С, Ма Ю, Хантер Н (2012). «Определение путей разрешения совместных молекул в мейозе идентифицирует резольвазу, специфичную для кроссовера» . Клетка . 149 (2): 334–47. дои : 10.1016/j.cell.2012.03.023 . ПМЦ 3377385 . ПМИД 22500800 .
- ^ Jump up to: а б Ранджа Л., Ананд Р., Чейка П. (2014). «Гетеродимер Saccharomyces cerevisiae Mlh1-Mlh3 представляет собой эндонуклеазу, которая преимущественно связывается с соединениями Холлидея» . Ж. Биол. Хим . 289 (9): 5674–86. дои : 10.1074/jbc.M113.533810 . ПМЦ 3937642 . ПМИД 24443562 .
- ^ Рогачева М.В., Манхарт С.М., Чен С., Гуарн А., Сёртиз Дж., Алани Э. (2014). «Mlh1-Mlh3, фактор мейотического кроссинговера и репарации несоответствия ДНК, представляет собой эндонуклеазу, стимулируемую Msh2-Msh3» . Ж. Биол. Хим . 289 (9): 5664–73. дои : 10.1074/jbc.M113.534644 . ПМЦ 3937641 . ПМИД 24403070 .
- ^ Jump up to: а б Зоннтаг Браун М., Лим Э., Чен С., Нишант К.Т., Алани Э. (2013). «Генетический анализ мутаций mlh3 выявляет взаимодействие между факторами, способствующими кроссоверу во время мейоза у пекарских дрожжей» . G3: Гены, геномы, генетика . 3 (1): 9–22. дои : 10.1534/g3.112.004622 . ПМК 3538346 . ПМИД 23316435 .
- ^ Jump up to: а б Почарт П., Уолтеринг Д., Холлингсворт Н.М. (1997). «Консервативные свойства функционально различных гомологов MutS у дрожжей» . Ж. Биол. Хим . 272 (48): 30345–9. дои : 10.1074/jbc.272.48.30345 . ПМИД 9374523 .
- ^ Винанд, Нью-Джерси, Panzer JA, Колоднер Р.Д. (1998). «Клонирование и характеристика гомологов человека и Caenorhabditis elegans гена Saccharomyces cerevisiae MSH5» . Геномика . 53 (1): 69–80. дои : 10.1006/geno.1998.5447 . ПМИД 9787078 .
- ^ Бокер Т., Барусевичус А., Сноуден Т., Расио Д., Герретт С., Роббинс Д., Шмидт С., Бурчак Дж., Кроче С.М., Коупленд Т., Коватич А.Дж., Фишел Р. (1999). «hMSH5: гомолог MutS человека, который образует новый гетеродимер с hMSH4 и экспрессируется во время сперматогенеза». Рак Рез . 59 (4): 816–22. ПМИД 10029069 .
- ^ Jump up to: а б Кришнапрасад Г.Н., Ананд М.Т., Лин Г., Теккедил М.М., Стейнмец Л.М., Нишант К.Т. (2015). «Изменение частоты кроссовера нарушает гарантию кроссовера, не влияя на мейотическую сегрегацию хромосом у Saccharomyces cerevisiae» . Генетика . 199 (2): 399–412. дои : 10.1534/genetics.114.172320 . ПМК 4317650 . ПМИД 25467183 .
- ^ Дэвис Л., Смит Г.Р. (2005). «Динеин способствует сегрегации ахиазматов у Schizosaccharomyces pombe» . Генетика . 170 (2): 581–90. doi : 10.1534/genetics.104.040253 . ПМЦ 1450395 . ПМИД 15802518 .