нуклеосома

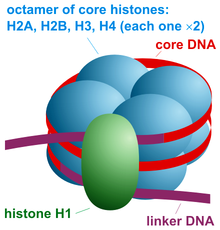
Нуклеосома эукариот — основная структурная единица ДНК у . упаковки Структура нуклеосомы состоит из участка ДНК, намотанного вокруг восьми белков-гистонов. [1] и напоминает нить, намотанную на катушку. Нуклеосома является основной субъединицей хроматина . Каждая нуклеосома состоит из чуть менее двух витков ДНК, обернутых вокруг набора из восьми белков, называемых гистонами, которые известны как гистоновый октамер . Каждый октамер гистонов состоит из двух копий каждого из белков-гистонов H2A , H2B , H3 и H4 .
ДНК должна быть уплотнена в нуклеосомы, чтобы поместиться в ядре клетки . [2] Помимо упаковки нуклеосом, эукариотический хроматин дополнительно уплотняется за счет сворачивания в ряд более сложных структур, в конечном итоге образующих хромосому . Каждая клетка человека содержит около 30 миллионов нуклеосом. [3]
Считается, что нуклеосомы несут эпигенетически наследуемую информацию в форме ковалентных модификаций своих основных гистонов . Положения нуклеосом в геноме не случайны, и важно знать, где расположена каждая нуклеосома, поскольку это определяет доступность ДНК для регуляторных белков . [4]
Нуклеосомы как частицы впервые были обнаружены в электронном микроскопе Доном и Адой Олинс в 1974 году. [5] а их существование и структура (в виде октамеров гистонов, окруженных примерно 200 парами оснований ДНК) были предложены Роджером Корнбергом . [6] [7] Роль нуклеосомы как регулятора транскрипции была продемонстрирована Lorch et al. in vitro [8] в 1987 году и Ханом и Грюнстейном [9] и Кларк-Адамс и др. [10] жив в 1988 году.
Коровая частица нуклеосомы состоит примерно из 146 пар оснований (п.н.) ДНК. [11] обернутый 1,67 левосторонними суперспиральными витками вокруг гистонового октамера, состоящего из 2 копий каждого из основных гистонов H2A , H2B , H3 и H4 . [12] Частицы ядра соединены участками линкерной ДНК , длина которых может достигать примерно 80 пар оснований. Технически нуклеосома определяется как ядро частицы плюс одна из этих линкерных областей; однако это слово часто является синонимом основной частицы. [13] Полногеномные карты расположения нуклеосом теперь доступны для многих модельных организмов и клеток человека. [14]
Линкерные гистоны, такие как H1 и его изоформы, участвуют в уплотнении хроматина и располагаются у основания нуклеосомы рядом с входом и выходом ДНК, связываясь с линкерной областью ДНК. [15] Неконденсированные нуклеосомы без линкерного гистона под электронным микроскопом напоминают «бусинки на нити ДНК» . [16]
В отличие от большинства эукариотических клеток, зрелые сперматозоиды в основном используют протамины для упаковки своей геномной ДНК, скорее всего, для достижения еще более высокого коэффициента упаковки. [17] Эквиваленты гистонов и упрощенная структура хроматина также были обнаружены у архей . [18] предполагая, что эукариоты — не единственные организмы, использующие нуклеосомы.
Структура
[ редактировать ]Структура ядра частицы
[ редактировать ]
Обзор
[ редактировать ]Новаторские структурные исследования, проведенные в 1980-х годах группой Аарона Клуга, предоставили первые доказательства того, что октамер гистоновых белков обертывает ДНК вокруг себя примерно за 1,7 витка левой суперспирали. [19] В 1997 году группа Ричмонда впервые расшифровала кристаллическую структуру нуклеосомы с разрешением, близким к атомному, показав наиболее важные детали частицы. человека, Альфа-сателлитная палиндромная ДНК имеющая решающее значение для достижения кристаллической структуры нуклеосомы 1997 года, была разработана группой Буника в Национальной лаборатории Ок-Ридж в Теннесси. [20] [21] [22] [23] [24] На сегодняшний день решена структура более 20 различных ядерных частиц нуклеосом. [25] включая те, которые содержат варианты гистонов и гистоны разных видов. Структура коровой частицы нуклеосомы удивительно консервативна, и даже замена более чем 100 остатков между гистонами лягушки и дрожжей приводит к появлению карт электронной плотности с общим среднеквадратичным отклонением всего 1,6 Å. [26]
Коровая частица нуклеосомы (NCP)
[ редактировать ]Коровая частица нуклеосомы (показана на рисунке) состоит примерно из 146 пар оснований ДНК. [11] обернутый 1,67 левосторонними суперспиральными витками вокруг гистонового октамера , состоящего из 2 копий каждого из основных гистонов H2A , H2B , H3 и H4 . Соседние нуклеосомы соединены участком свободной ДНК, называемым линкерной ДНК (длина которого варьируется от 10 до 80 пар оснований в зависимости от вида и типа ткани). [18] ).Вся структура образует цилиндр диаметром 11 нм и высотой 5,5 нм.


Частицы ядра нуклеосомы наблюдаются, когда хроматин в интерфазе обрабатывается, чтобы заставить хроматин частично развернуться. Полученное изображение, полученное с помощью электронного микроскопа, представляет собой «бусины на нитке». Нить — это ДНК, а каждая бусинка нуклеосомы — это центральная частица. Коровая частица нуклеосомы состоит из ДНК и белков-гистонов. [29]
Частичное ДНКазой расщепление хроматина раскрывает его нуклеосомную структуру. Поскольку участки ДНК ядерных частиц нуклеосом менее доступны для ДНКазы, чем связывающие участки, ДНК расщепляется на фрагменты длиной, равной кратности расстояния между нуклеосомами (180, 360, 540 пар оснований и т. д.). очень характерный рисунок, похожий на лестницу . виден Следовательно, во время гель-электрофореза этой ДНК [27] Такое переваривание может происходить и в естественных условиях во время апоптоза («самоубийство клетки» или запрограммированная гибель клетки), поскольку его ролью обычно является саморазрушение ДНК . [30]
Белковые взаимодействия внутри нуклеосомы
[ редактировать ]Коровые гистоновые белки содержат характерный структурный мотив, называемый «складкой гистонов», который состоит из трех альфа-спиралей (α1-3), разделенных двумя петлями (L1-2). В растворе гистоны образуют гетеродимеры H2A-H2B и гетеротетрамеры H3-H4. Гистоны димеризуются вокруг своих длинных α2-спиралей в антипараллельной ориентации, а в случае H3 и H4 два таких димера образуют пучок из 4 спиралей, стабилизированный обширным взаимодействием H3-H3'. Димер H2A/H2B связывается с тетрамером H3/H4 за счет взаимодействий между H4 и H2B, которые включают образование гидрофобного кластера. [12] Октамер гистонов образован центральным тетрамером H3/H4, зажатым между двумя димерами H2A/H2B. Из-за сильно основного заряда всех четырех основных гистонов октамер гистонов стабилен только в присутствии ДНК или очень высоких концентраций соли.
Взаимодействие гистона и ДНК
[ редактировать ]Нуклеосома содержит более 120 прямых взаимодействий белок-ДНК и несколько сотен водоопосредованных. [31] Прямые белок-ДНК-взаимодействия не распределены равномерно по поверхности октамера, а расположены в дискретных участках. Это связано с образованием двух типов сайтов связывания ДНК внутри октамера; сайт α1α1, который использует спираль α1 двух соседних гистонов, и сайт L1L2, образованный петлями L1 и L2. Солевые связи и водородные связи между основными и гидроксильными группами боковой цепи, а также амидами основной цепи с фосфатами основной цепи ДНК составляют основную часть взаимодействий с ДНК. Это важно, учитывая, что повсеместное распространение нуклеосом по геномам требует, чтобы они были неспецифическим для последовательности ДНК-связывающим фактором. Хотя нуклеосомы имеют тенденцию отдавать предпочтение одним последовательностям ДНК другим, [32] они способны связываться практически с любой последовательностью, что, как полагают, обусловлено гибкостью формирования этих опосредованных водой взаимодействий. Кроме того, между боковыми цепями белка и группами дезоксирибозы возникают неполярные взаимодействия, а боковая цепь аргинина вставляется в малую бороздку ДНК во всех 14 сайтах, где она обращена к поверхности октамера.Распределение и сила сайтов связывания ДНК на поверхности октамера искажают ДНК внутри ядра нуклеосомы. ДНК неравномерно изогнута, а также содержит дефекты скручивания. Скрутка свободной В-формы ДНК в растворе составляет 10,5 п.н. на виток. Однако общий поворот нуклеосомной ДНК составляет всего 10,2 п.н. на оборот, варьируясь от значения от 9,4 до 10,9 п.н. на оборот.
Хвостовые домены гистонов
[ редактировать ]Отростки гистоновых хвостов составляют до 30% массы гистонов, но не видны в кристаллических структурах нуклеосом из-за их высокой внутренней гибкости и считаются в значительной степени неструктурированными. [33] N-концевые хвосты гистонов H3 и H2B проходят через канал, образованный малыми бороздками двух цепей ДНК, выступая из ДНК каждые 20 п.н. N -концевой хвост гистона Н4, напротив, имеет участок высокоосновных аминокислот (16–25), который в кристаллической структуре образует взаимодействие с высококислотной поверхностной областью димера H2A-H2B. другой нуклеосомы, что потенциально имеет отношение к структуре нуклеосом более высокого порядка. Считается, что это взаимодействие происходит и в физиологических условиях и предполагает, что ацетилирование хвоста H4 искажает структуру хроматина высшего порядка. [ нужна ссылка ]
Структура высшего порядка
[ редактировать ]
Организация ДНК, достигаемая нуклеосомой, не может полностью объяснить упаковку ДНК, наблюдаемую в ядре клетки. Дальнейшее уплотнение хроматина в клеточное ядро необходимо, но оно еще недостаточно изучено. Текущее понимание [25] заключается в том, что повторяющиеся нуклеосомы с промежуточной «линкерной» ДНК образуют волокно длиной 10 нм , описываемое как «бусинки на нитке», и имеют коэффициент упаковки примерно от пяти до десяти. [18] Цепочка нуклеосом может быть организована в волокно длиной 30 нм — компактную структуру с коэффициентом упаковки ~50. [18] и чье образование зависит от присутствия гистона H1 .
Кристаллическая структура тетрануклеосомы была представлена и использована для создания предлагаемой структуры волокна диаметром 30 нм в виде двухзачатковой спирали. [34] По поводу этой модели до сих пор существуют определенные разногласия, поскольку она несовместима с недавними данными электронной микроскопии . [35] Помимо этого, структура хроматина плохо изучена, но классически предполагается, что волокна длиной 30 нм расположены в виде петель вдоль центрального белкового каркаса с образованием транскрипционно активного эухроматина . Дальнейшее уплотнение приводит к транскрипционно неактивному гетерохроматину .
Динамика
[ редактировать ]Хотя нуклеосома представляет собой очень стабильный комплекс белок-ДНК, она не статична и, как было показано, претерпевает ряд различных структурных перестроек, включая скольжение нуклеосомы и обнажение участков ДНК. В зависимости от контекста нуклеосомы могут ингибировать или облегчать связывание факторов транскрипции. Положения нуклеосом контролируются тремя основными факторами: во-первых, внутренняя аффинность связывания октамера гистонов зависит от последовательности ДНК. Во-вторых, нуклеосома может быть вытеснена или рекрутирована за счет конкурентного или кооперативного связывания др. белковых факторов. В-третьих, нуклеосома может активно транслоцироваться с помощью АТФ-зависимых комплексов ремоделирования. [36]
Нуклеосомное скольжение
[ редактировать ]Работа, проведенная в лаборатории Брэдбери, показала, что нуклеосомы, восстановленные в позиционирующей последовательности 5S ДНК, способны трансляционно перемещаться на соседние последовательности при термической инкубации. [37] Более поздние работы показали, что это изменение положения не требует разрушения октамера гистонов, но соответствует способности нуклеосом «скользить» вдоль ДНК в цис . В 2008 году было дополнительно обнаружено, что сайты связывания CTCF действуют как якоря позиционирования нуклеосом, так что при использовании для выравнивания различных геномных сигналов можно легко идентифицировать множественные фланкирующие нуклеосомы. [38] Хотя нуклеосомы по своей природе подвижны, у эукариот развилось большое семейство АТФ-зависимых ферментов ремоделирования хроматина, изменяющих структуру хроматина, многие из которых делают это посредством скольжения нуклеосом. В 2012 году лаборатория Бины Пиллаи продемонстрировала, что скольжение нуклеосом является одним из возможных механизмов крупномасштабной тканеспецифичной экспрессии генов. Работа показывает, что сайт начала транскрипции генов, экспрессируемых в определенной ткани, истощен нуклеосомами, в то время как тот же набор генов в другой ткани, где они не экспрессируются, связан с нуклеосомами. [39]
Воздействие участка ДНК
[ редактировать ]Работа лаборатории Видома показала, что нуклеосомная ДНК находится в равновесии между завернутым и развернутым состоянием. Измерения этих скоростей с использованием FRET с временным разрешением показали, что ДНК внутри нуклеосомы остается полностью завернутой только в течение 250 мс, прежде чем она разворачивается в течение 10-50 мс, а затем быстро переворачивается. [40] Это означает, что ДНК не обязательно должна активно отделяться от нуклеосомы, но существует значительный промежуток времени, в течение которого она полностью доступна. Действительно, это можно распространить на наблюдение, что введение ДНК-связывающей последовательности внутри нуклеосомы увеличивает доступность соседних областей ДНК при связывании. [41] Эта склонность ДНК внутри нуклеосомы «дышать» имеет важные функциональные последствия для всех ДНК-связывающих белков, которые действуют в среде хроматина. [40] В частности, динамическое дыхание нуклеосом играет важную роль в ограничении продвижения РНК-полимеразы II во время элонгации транскрипции. [42]
Нуклеосомная свободная область
[ редактировать ]Промоторы активных генов имеют свободные от нуклеосом области (NFR). Это обеспечивает доступность промоторной ДНК для различных белков, таких как факторы транскрипции. обычно имеет длину 200 нуклеотидов. Свободная от нуклеосом область у S. cerevisiae [43] Хорошо расположенные нуклеосомы образуют границы NFR. Эти нуклеосомы называются +1-нуклеосомой и -1-нуклеосомой и расположены на канонических расстояниях ниже и выше соответственно от места начала транскрипции. [44] +1-нуклеосома и несколько последующих нуклеосом также имеют тенденцию включать вариант гистона H2A.Z. [44]
Модулирующая структура нуклеосомы
[ редактировать ]Геномы эукариот повсеместно связаны с хроматином; однако клетки должны пространственно и временно регулировать специфические локусы независимо от объема хроматина. Чтобы достичь высокого уровня контроля, необходимого для координации ядерных процессов, таких как репликация, репарация и транскрипция ДНК, клетки разработали множество средств для локальной и специфической модуляции структуры и функции хроматина. Это может включать ковалентную модификацию гистонов, включение вариантов гистонов и нековалентное ремоделирование с помощью АТФ-зависимых ферментов ремоделирования.
Посттрансляционные модификации гистонов
[ редактировать ]
С момента их открытия в середине 1960-х годов было предсказано, что модификации гистонов будут влиять на транскрипцию. [45] Тот факт, что большинство ранних обнаруженных посттрансляционных модификаций были сосредоточены в отростках хвостов, выступающих из ядра нуклеосомы, приводит к двум основным теориям относительно механизма модификации гистонов. Первая из теорий предполагала, что они могут влиять на электростатические взаимодействия между хвостами гистонов и ДНК, «расслабляя» структуру хроматина. Позже было высказано предположение, что комбинации этих модификаций могут создавать связывающие эпитопы, с помощью которых можно рекрутировать другие белки. [46] Недавно, учитывая, что в структурированных областях гистонов было обнаружено больше модификаций, было высказано предположение, что эти модификации могут влиять на ДНК гистонов. [47] и гистон-гистон [48] взаимодействия внутри ядра нуклеосомы. Предполагается, что модификации (такие как ацетилирование или фосфорилирование), которые снижают заряд ядра глобулярного гистона, «ослабляют» ассоциацию ядро-ДНК; сила эффекта зависит от местоположения модификации внутри ядра. [49] Было показано, что некоторые модификации коррелируют с молчанием генов; другие, по-видимому, коррелируют с активацией генов. Общие модификации ацетилирование , метилирование или убиквитинирование лизина ; включают метилирование аргинина ; и фосфорилирование серина . Информация, хранящаяся таким образом, считается эпигенетической , поскольку она не кодируется в ДНК, но по-прежнему передается по наследству дочерним клеткам. Поддержание подавленного или активированного статуса гена часто необходимо для клеточной дифференцировки . [18]
Варианты гистонов
[ редактировать ]Хотя гистоны удивительно консервативны на протяжении эволюции, было идентифицировано несколько их вариантов форм. Это разнообразие функций гистонов ограничивается H2A и H3, при этом H2B и H4 в основном инвариантны. H2A может быть заменен на H2AZ (что приводит к снижению стабильности нуклеосом) или H2AX (который связан с репарацией ДНК и дифференцировкой Т-клеток ), тогда как неактивные Х-хромосомы у млекопитающих обогащены макроH2A. H3 может быть заменен на H3.3 (который коррелирует с активирующими генами и регуляторными элементами), а в центромерах H3 заменяется на CENPA . [18]
АТФ-зависимое ремоделирование нуклеосом
[ редактировать ]Ряд различных реакций связан с термином АТФ-зависимое ремоделирование хроматина . Было показано, что ферменты ремоделирования скользят нуклеосомы вдоль ДНК. [50] нарушают контакты гистона с ДНК вплоть до дестабилизации димера H2A/H2B [51] [52] и генерировать отрицательное сверхспиральное скручивание ДНК и хроматина. [53] Недавно было показано, что фермент ремоделирования Swr1 вводит вариант гистона H2A.Z в нуклеосомы. [54] В настоящее время неясно, представляют ли все это отдельные реакции или просто альтернативные результаты общего механизма. Что общего между всеми и действительно является отличительной чертой АТФ-зависимого ремоделирования хроматина, так это то, что все они приводят к изменению доступности ДНК.
Исследования, изучающие активацию генов in vivo [55] и, что еще более удивительно, ремоделирование in vitro. [56] обнаружили, что события ремоделирования хроматина и связывание транскрипционных факторов носят циклический и периодический характер. Хотя последствия этого для механизма реакции ремоделирования хроматина неизвестны, динамическая природа системы может позволить ей быстрее реагировать на внешние стимулы. Недавнее исследование показывает, что положения нуклеосом значительно изменяются во время развития эмбриональных стволовых клеток мыши, и эти изменения связаны со связыванием факторов транскрипции развития. [57]
Динамическое ремоделирование нуклеосом в геноме дрожжей
[ редактировать ]Исследования 2007 года каталогизировали положения нуклеосом у дрожжей и показали, что нуклеосомы истощены в промоторных регионах и местах начала репликации . [58] [59] [60] Около 80% генома дрожжей покрыто нуклеосомами. [61] и характер позиционирования нуклеосом явно связан с областями ДНК, которые регулируют транскрипцию , областями, которые транскрибируются, и областями, которые инициируют репликацию ДНК. [62] Совсем недавно новое исследование изучило динамические изменения в репозиции нуклеосом во время глобального события транскрипционного перепрограммирования, чтобы выяснить влияние на смещение нуклеосом во время общегеномных транскрипционных изменений у дрожжей ( Saccharomyces cerevisiae ). [63] Результаты показали, что нуклеосомы, локализованные в промоторных областях, смещаются в ответ на стресс (например, тепловой шок ). Кроме того, удаление нуклеосом обычно соответствовало активации транскрипции, а замена нуклеосом обычно соответствовала репрессии транскрипции, предположительно потому, что сайты связывания транскрипционных факторов становились более или менее доступными соответственно. В общем, только одна или две нуклеосомы были перемещены в промоторе, чтобы вызвать эти транскрипционные изменения. Однако даже в хромосомных регионах, которые не были связаны с транскрипционными изменениями, наблюдалось репозиционирование нуклеосом, что указывает на то, что покрытие и раскрытие транскрипционной ДНК не обязательно приводит к событию транскрипции. После транскрипции область рДНК должна быть защищена от любых повреждений. Это предполагает, что белки HMGB играют важную роль в защите свободной от нуклеосомы области. [64] [65]
Дефекты скручивания ДНК
[ редактировать ]Дефекты скручивания ДНК — это когда добавление одной или нескольких пар оснований из одного сегмента ДНК переносится на следующий сегмент, что приводит к изменению скручивания ДНК. Это не только изменит скручивание ДНК, но и изменит длину. [66] Этот дефект скручивания в конечном итоге перемещается вокруг нуклеосомы посредством переноса пары оснований, это означает, что скручивания ДНК могут вызвать скольжение нуклеосомы. [67] Кристаллические структуры нуклеосом показали, что в местах суперспирали 2 и 5 нуклеосомы обычно возникают дефекты скручивания ДНК, поскольку это общие сайты связывания ремоделеров. [68] Существует множество ремоделаторов хроматина, но все они имеют общий двигатель АТФазы, который облегчает скольжение хроматина по ДНК посредством связывания и гидролиза АТФ. [69] АТФаза имеет открытое и закрытое состояние. Когда двигатель АТФазы переходит из открытого в закрытое состояние, дуплекс ДНК меняет геометрию и демонстрирует наклон пары оснований. [68] Инициирование дефектов скручивания с помощью мотора АТФазы вызывает накопление напряжения вокруг участка ремоделирования. Напряжение снимается, когда скольжение ДНК завершено по нуклеосоме за счет распространения двух дефектов скручивания (по одному на каждой цепи) в противоположных направлениях. [69]
Сборка нуклеосом in vitro
[ редактировать ]
Нуклеосомы можно собирать in vitro с использованием очищенных нативных или рекомбинантных гистонов. [70] [71] Один стандартный метод загрузки ДНК вокруг гистонов включает использование солевого диализа . Реакцию, состоящую из октамеров гистонов и обнаженной матрицы ДНК, можно инкубировать вместе при концентрации соли 2 М. При постепенном уменьшении концентрации соли ДНК придет в равновесие до положения, в котором она обертывается вокруг октамеров гистонов, образуя нуклеосомы. В соответствующих условиях этот процесс восстановления позволяет экспериментально картировать аффинность позиционирования нуклеосом данной последовательности. [72]
Частицы ядра нуклеосомы, сшитые дисульфидом
[ редактировать ]Недавний прогресс в производстве ядерных частиц с повышенной стабильностью включает в себя сайт-специфические дисульфидные сшивки. [73] В ядро нуклеосомной частицы можно ввести две разные поперечные связи. Первый связывает две копии H2A посредством введенного цистеина (N38C), в результате чего образуется октамер гистонов , который устойчив к потере димера H2A/H2B во время восстановления нуклеосомы. Вторая сшивка может быть введена между N-концевым хвостом гистона H3 и концами нуклеосомной ДНК посредством встроенного конвертируемого нуклеотида. [74] Сшивка октамера ДНК-гистон стабилизирует ядро нуклеосомной частицы против диссоциации ДНК при очень низких концентрациях частиц и при повышенных концентрациях соли.
Сборка нуклеосом in vivo
[ редактировать ]
Нуклеосомы — это основная единица упаковки геномной ДНК, построенная из белков-гистонов, вокруг которых свернута ДНК. Они служат основой для формирования структуры хроматина более высокого порядка, а также для уровня регуляторного контроля экспрессии генов. Нуклеосомы быстро собираются на вновь синтезированной ДНК за репликационной вилкой.
Н3 и Н4
[ редактировать ]Гистоны H3 и H4 из разобранных старых нуклеосом остаются поблизости и случайным образом распределяются по вновь синтезированной ДНК. [75] Они собираются комплексом фактора сборки хроматина 1 (CAF-1), который состоит из трех субъединиц (p150, p60 и p48). [76] Вновь синтезированные H3 и H4 собираются с помощью фактора сборки репликационной связи (RCAF). RCAF содержит субъединицу Asf1, которая связывается с вновь синтезированными белками H3 и H4. [77] Старые белки H3 и H4 сохраняют свои химические модификации, что способствует передаче эпигенетической подписи. Вновь синтезированные белки H3 и H4 постепенно ацетилируются по различным остаткам лизина в рамках процесса созревания хроматина. [78] Также считается, что старые белки H3 и H4 в новых нуклеосомах привлекают ферменты, модифицирующие гистоны, которые маркируют новые гистоны, способствуя эпигенетической памяти.
H2A и H2B
[ редактировать ]В отличие от старых H3 и H4, старые белки-гистоны H2A и H2B высвобождаются и разрушаются; следовательно, вновь собранные белки H2A и H2B включаются в новые нуклеосомы. [79] H2A и H2B собираются в димеры, которые затем загружаются на нуклеосомы с помощью белка сборки нуклеосомы-1 (NAP-1), который также способствует скольжению нуклеосом. [80] Нуклеосомы также разделены АТФ-зависимыми комплексами ремоделирования нуклеосом, содержащими ферменты, такие как Isw1 Ino80 и Chd1, и впоследствии собираются в структуру более высокого порядка. [81] [82]
Галерея
[ редактировать ]

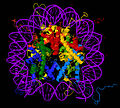
Кристаллическая структура ядра нуклеосомной частицы ( PDB : 1EQZ [28] ) — разные виды, показывающие детали сворачивания и организации гистонов. Гистоны Н2А , H2B , Н3 , H4 и ДНК окрашена.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Рис Дж., Кэмпбелл Н. (2006). Биология . Сан-Франциско: Бенджамин Каммингс. ISBN 978-0-8053-6624-2 .
- ^ Альбертс Б (2002). «Хромосомная ДНК и ее упаковка в хроматиновом волокне». Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. п. 207. ИСБН 978-0-8153-4072-0 .
- ^ Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). Хромосомная ДНК и ее упаковка в хроматиновом волокне . Гирляндная наука.
- ^ Тейф В.Б., Кларксон, Коннектикут (2019). «Позиционирование нуклеосом». Энциклопедия биоинформатики и вычислительной биологии . 2 : 308–317. дои : 10.1016/B978-0-12-809633-8.20242-2 . ISBN 9780128114322 . S2CID 43929234 .
- ^ Олинс А.Л., Олинс Д.Э. (январь 1974 г.). «Единицы сфероидного хроматина (v тельца)». Наука . 183 (4122): 330–332. Бибкод : 1974Sci...183..330O . дои : 10.1126/science.183.4122.330 . ПМИД 4128918 . S2CID 83480762 .
- ^ Макдональд Д. (декабрь 2005 г.). «Веха 9 (1973–1974) Нуклеосомная гипотеза: альтернативная теория струн» . Вехи природы: экспрессия генов . дои : 10.1038/nrm1798 .
- ^ Корнберг Р.Д. (май 1974 г.). «Структура хроматина: повторяющаяся единица гистонов и ДНК». Наука . 184 (4139): 868–871. Бибкод : 1974Sci...184..868K . дои : 10.1126/science.184.4139.868 . ПМИД 4825889 .
- ^ Лорх Ю., ЛаПойнт Дж.В., Корнберг Р.Д. (апрель 1987 г.). «Нуклеосомы ингибируют инициацию транскрипции, но допускают удлинение цепи со смещением гистонов». Клетка . 49 (2): 203–210. дои : 10.1016/0092-8674(87)90561-7 . ПМИД 3568125 . S2CID 21270171 .
- ^ Хан М., Грунштейн М. (декабрь 1988 г.). «Потеря нуклеосомы активирует нижние промоторы дрожжей in vivo». Клетка . 55 (6): 1137–1145. дои : 10.1016/0092-8674(88)90258-9 . ПМИД 2849508 . S2CID 41520634 .
- ^ Кларк-Адамс CD, Норрис Д., Осли М.А., Фасслер Дж.С., Уинстон Ф. (февраль 1988 г.). «Изменения в дозировке генов гистонов изменяют транскрипцию у дрожжей» . Гены и развитие . 2 (2): 150–159. дои : 10.1101/gad.2.2.150 . ПМИД 2834270 .
- ^ Перейти обратно: а б В разных кристаллах наблюдались значения 146 и 147 пар оснований.
- ^ Перейти обратно: а б Люгер К., Мэдер А.В., Ричмонд Р.К., Сарджент Д.Ф., Ричмонд Т.Дж. (сентябрь 1997 г.). «Кристаллическая структура ядра нуклеосомы при разрешении 2,8 А». Природа . 389 (6648): 251–260. Бибкод : 1997Natur.389..251L . дои : 10.1038/38444 . ПМИД 9305837 . S2CID 4328827 .
- ^ Альбертс Б. (2007). Молекулярная биология клетки (5-е изд.). Нью-Йорк: Garland Science. п. 211. ИСБН 978-0-8153-4106-2 .
- ^ Штумпф М., Пироева К.В., Агравал С.П., Якоб Д.Р., Тейф В.Б. (июнь 2022 г.). «NucPosDB: база данных позиционирования нуклеосом in vivo и нуклеосомики бесклеточной ДНК» . Хромосома . 131 (1–2): 19–28. дои : 10.1007/s00412-021-00766-9 . ПМЦ 8776978 . ПМИД 35061087 .
- ^ Чжоу Ю.Б., Герхман С.Е., Рамакришнан В., Трэверс А., Мюлдерманс С. (сентябрь 1998 г.). «Положение и ориентация глобулярного домена линкерного гистона H5 на нуклеосоме» . Природа . 395 (6700): 402–405. Бибкод : 1998Natur.395..402Z . дои : 10.1038/26521 . ПМИД 9759733 . S2CID 204997317 .
- ^ Тома Ф., Коллер Т., Клуг А. (ноябрь 1979 г.). «Участие гистона H1 в организации нуклеосомы и солевых суперструктур хроматина» . Журнал клеточной биологии . 83 (2 ч. 1): 403–427. дои : 10.1083/jcb.83.2.403 . ПМК 2111545 . ПМИД 387806 .
- ^ Кларк Х.Дж. (1992). «Ядерный и хроматиновый состав гамет млекопитающих и ранних эмбрионов». Биохимия и клеточная биология . 70 (10–11): 856–866. дои : 10.1139/o92-134 . ПМИД 1297351 .
- ^ Перейти обратно: а б с д и ж Фельзенфельд Г., Грудин М. (январь 2003 г.). «Управление двойной спиралью» . Природа . 421 (6921): 448–453. Бибкод : 2003Natur.421..448F . дои : 10.1038/nature01411 . ПМИД 12540921 .
- ^ Ричмонд Т.Дж., Финч Дж.Т., Раштон Б., Роудс Д., Клуг А. (1984). «Структура ядра нуклеосомы при разрешении 7 А». Природа . 311 (5986): 532–7. Бибкод : 1984Natur.311..532R . дои : 10.1038/311532a0 . ПМИД 6482966 . S2CID 4355982 .
- ^ Харп Дж.М., Палмер Э.Л., Йорк М.Х., Гевисс А., Дэвис М., Буник Г.Дж. (октябрь 1995 г.). «Препаративное разделение ядерных частиц нуклеосомы, содержащих ДНК с определенной последовательностью, в нескольких фазах трансляции». Электрофорез . 16 (10): 1861–1864. дои : 10.1002/elps.11501601305 . ПМИД 8586054 . S2CID 20178479 .
- ^ Палмер Э.Л., Гевисс А., Харп Дж.М., Йорк М.Х., Буник Г.Дж. (октябрь 1995 г.). «Массовое производство фрагментов палиндромной ДНК». Аналитическая биохимия . 231 (1): 109–114. дои : 10.1006/abio.1995.1509 . ПМИД 8678288 .
- ^ Харп Дж.М., Убербахер Э.К., Роберсон А.Е., Палмер Э.Л., Гевисс А., Буник Г.Дж. (март 1996 г.). «Рентгеноструктурный анализ кристаллов, содержащих частицы ядра двусимметричных нуклеосом» . Акта Кристаллографика. Раздел D. Биологическая кристаллография . 52 (Часть 2): 283–288. Бибкод : 1996AcCrD..52..283H . дои : 10.1107/S0907444995009139 . ПМИД 15299701 .
- ^ Харп Дж.М., Хэнсон Б.Л., Тимм Д.Е., Буник Г.Дж. (декабрь 2000 г.). «Асимметрия ядра нуклеосомной частицы при разрешении 2,5 А». Акта Кристаллографика. Раздел D. Биологическая кристаллография . 56 (Часть 12): 1513–1534. дои : 10.1107/s0907444900011847 . ПМИД 11092917 .
- ^ Хэнсон Б.Л., Александр С., Харп Дж.М., Буник Г.Дж. (2004). «Получение и кристаллизация ядерной частицы нуклеосомы». Хроматин и ферменты ремоделирования хроматина, часть А. Методы энзимологии. Том. 375. стр. 44–62. дои : 10.1016/s0076-6879(03)75003-4 . ISBN 9780121827793 . ПМИД 14870658 .
- ^ Перейти обратно: а б Чакраварти С., Пак Ю.Дж., Чодапарамбил Дж., Эдайатхумангалам Р.С., Люгер К. (февраль 2005 г.). «Структура и динамические свойства частиц ядра нуклеосомы» . Письма ФЭБС . 579 (4): 895–898. Бибкод : 2005FEBSL.579..895C . дои : 10.1016/j.febslet.2004.11.030 . ПМИД 15680970 . S2CID 41706403 .
- ^ Уайт CL, Суто Р.К., Люгер К. (сентябрь 2001 г.). «Структура ядра нуклеосомной частицы дрожжей обнаруживает фундаментальные изменения в межнуклеосомных взаимодействиях» . Журнал ЭМБО . 20 (18): 5207–5218. дои : 10.1093/emboj/20.18.5207 . ПМЦ 125637 . PMID 11566884 .
- ^ Перейти обратно: а б Страйер Л. (1995). Биохимия (четвертое изд.). Нью-Йорк - Бейзингсток: WH Freeman and Company. ISBN 978-0716720096 .
- ^ Перейти обратно: а б Харп Дж.М., Хэнсон Б.Л., Тимм Д.Е., Буник Г.Дж. (декабрь 2000 г.). «Асимметрия ядра нуклеосомной частицы при разрешении 2,5 А». Акта Кристаллографика. Раздел D. Биологическая кристаллография . 56 (Часть 12): 1513–1534. дои : 10.1107/S0907444900011847 . ПМИД 11092917 . Идентификатор PDB: 1EQZ.
- ^ Альбертс Б (2009). Основная клеточная биология (2-е изд.). Нью-Йорк: Garland Science.
- ^ Аллен, Пол Д.; Ньюленд, Адриан К. (1 июня 1998 г.). «Электрофоретический анализ ДНК для выявления апоптоза» . Молекулярная биотехнология . 9 (3): 247–251. дои : 10.1007/BF02915798 . ISSN 1559-0305 . ПМИД 9718585 .
- ^ Дэйви К.А., Сарджент Д.Ф., Люгер К., Мэдер А.В., Ричмонд Т.Дж. (июнь 2002 г.). «Взаимодействия, опосредованные растворителем, в структуре ядерной частицы нуклеосомы с разрешением 1,9». Журнал молекулярной биологии . 319 (5): 1097–1113. дои : 10.1016/S0022-2836(02)00386-8 . ПМИД 12079350 .
- ^ Сигал Э., Фондюфе-Миттендорф Ю., Чен Л., Тострем А., Филд Ю., Мур И.К. и др. (август 2006 г.). «Геномный код позиционирования нуклеосом» . Природа . 442 (7104): 772–778. Бибкод : 2006Natur.442..772S . дои : 10.1038/nature04979 . ПМЦ 2623244 . ПМИД 16862119 .
- ^ Чжэн С., Хейс Джей-Джей (апрель 2003 г.). «Структуры и взаимодействия основных хвостовых доменов гистонов». Биополимеры . 68 (4): 539–546. дои : 10.1002/bip.10303 . ПМИД 12666178 .
- ^ Шальх Т., Дуда С., Сарджент Д.Ф., Ричмонд Т.Дж. (июль 2005 г.). «Рентгеновская структура тетрануклеосомы и ее значение для хроматинового волокна». Природа . 436 (7047): 138–141. Бибкод : 2005Natur.436..138S . дои : 10.1038/nature03686 . ПМИД 16001076 . S2CID 4387396 .
- ^ Робинсон П.Дж., Фэйролл Л., Хюинь В.А., Роудс Д. (апрель 2006 г.). «ЭМ-измерения определяют размеры «30-нм» хроматинового волокна: свидетельство компактной, встречно-пальцевой структуры» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (17): 6506–6511. Бибкод : 2006PNAS..103.6506R . дои : 10.1073/pnas.0601212103 . ПМЦ 1436021 . ПМИД 16617109 .
- ^ Тейф В.Б., Риппе К. (сентябрь 2009 г.). «Предсказание положения нуклеосом в ДНК: сочетание предпочтений внутренней последовательности и активности ремоделера» . Исследования нуклеиновых кислот . 37 (17): 5641–5655. дои : 10.1093/нар/gkp610 . ПМК 2761276 . ПМИД 19625488 .
- ^ Пеннингс С., Мюлдерманс С., Меерсеман Г., Винс Л. (май 1989 г.). «Формирование, стабильность и расположение основных гистонов нуклеосом, повторно собранных на изогнутой ДНК и другой ДНК, полученной из нуклеосом». Журнал молекулярной биологии . 207 (1): 183–192. дои : 10.1016/0022-2836(89)90449-X . ПМИД 2738923 .
- ^ Фу Ю, Синха М, Петерсон С.Л., Венг З. (июль 2008 г.). Ван Стинсел Б. (ред.). «Инсулятор-связывающий белок CTCF размещает 20 нуклеосом вокруг своих сайтов связывания в геноме человека» . ПЛОС Генетика . 4 (7): e1000138. дои : 10.1371/journal.pgen.1000138 . ПМЦ 2453330 . ПМИД 18654629 .
- ^ Баргадже Р., Алам М.П., Патовари А., Саркар М., Али Т., Гупта С. и др. (октябрь 2012 г.). «Близость нуклеосомы, содержащей H2A.Z, к месту начала транскрипции влияет на уровни экспрессии генов в печени и мозге млекопитающих» . Исследования нуклеиновых кислот . 40 (18): 8965–8978. дои : 10.1093/нар/gks665 . ПМК 3467062 . ПМИД 22821566 .
- ^ Перейти обратно: а б Ли Г, Левитус М, Бустаманте С, Видом Дж (январь 2005 г.). «Быстрая спонтанная доступность нуклеосомной ДНК». Структурная и молекулярная биология природы . 12 (1): 46–53. дои : 10.1038/nsmb869 . ПМИД 15580276 . S2CID 14540078 .
- ^ Ли Дж., Видом Дж. (август 2004 г.). «Нуклеосомы способствуют собственному вторжению». Структурная и молекулярная биология природы . 11 (8): 763–769. дои : 10.1038/nsmb801 . ПМИД 15258568 . S2CID 11299024 .
- ^ Ходжес С., Бинту Л., Лубковска Л., Кашлев М., Бустаманте С. (июль 2009 г.). «Нуклеосомные флуктуации управляют динамикой транскрипции РНК-полимеразы II» . Наука . 325 (5940): 626–628. Бибкод : 2009Sci...325..626H . дои : 10.1126/science.1172926 . ПМЦ 2775800 . ПМИД 19644123 .
- ^ Юань Г.К., Лю Ю.Дж., Дион М.Ф., Слэк М.Д., Ву Л.Ф., Альтшулер С.Дж., Рандо О.Дж. (июль 2005 г.). «Идентификация положений нуклеосом в масштабе генома у S. cerevisiae» . Наука . 309 (5734): 626–630. Бибкод : 2005Sci...309..626Y . дои : 10.1126/science.1112178 . ПМИД 15961632 . S2CID 43625066 .
- ^ Перейти обратно: а б Лай В.К., Пью Б.Ф. (сентябрь 2017 г.). «Понимание динамики нуклеосом и их связи с экспрессией генов и репликацией ДНК» . Обзоры природы. Молекулярно-клеточная биология . 18 (9): 548–562. дои : 10.1038/номер.2017.47 . ПМЦ 5831138 . ПМИД 28537572 .
- ^ Олфри В.Г., Фолкнер Р., Мирский А.Е. (май 1964 г.). «Ацетилирование и метилирование гистонов и их возможная роль в регуляции синтеза РНК» . Труды Национальной академии наук Соединенных Штатов Америки . 51 (5): 786–794. Бибкод : 1964PNAS...51..786A . дои : 10.1073/pnas.51.5.786 . ПМК 300163 . ПМИД 14172992 .
- ^ Strahl BD, Allis CD (январь 2000 г.). «Язык ковалентных модификаций гистонов». Природа . 403 (6765): 41–45. Бибкод : 2000Natur.403...41S . дои : 10.1038/47412 . ПМИД 10638745 . S2CID 4418993 .
- ^ Косгроув М.С., Буке Дж.Д., Вольбергер С. (ноябрь 2004 г.). «Регулируемая подвижность нуклеосом и код гистонов». Структурная и молекулярная биология природы . 11 (11): 1037–1043. дои : 10.1038/nsmb851 . ПМИД 15523479 . S2CID 34704745 .
- ^ Йе Дж., Ай Х, Евгений Э.Э., Чжан Л., Карпентер Л.Р., Елинек М.А. и др. (апрель 2005 г.). «Ацетилирование лизина 91 гистона H4 - модификация основного домена, связанная со сборкой хроматина» . Молекулярная клетка . 18 (1): 123–130. doi : 10.1016/j.molcel.2005.02.031 . ПМЦ 2855496 . ПМИД 15808514 .
- ^ Фенли А.Т., Адамс Д.А., Онуфриев А.В. (сентябрь 2010 г.). «Зарядовое состояние ядра глобулярного гистона контролирует стабильность нуклеосомы» . Биофизический журнал . 99 (5): 1577–1585. Бибкод : 2010BpJ....99.1577F . дои : 10.1016/j.bpj.2010.06.046 . ПМЦ 2931741 . ПМИД 20816070 .
- ^ Уайтхаус I, Флаус А., Кэрнс Б.Р., Уайт М.Ф., Уоркман Дж.Л., Оуэн-Хьюз Т. (август 1999 г.). «Мобилизация нуклеосом, катализируемая дрожжевым комплексом SWI/SNF». Природа . 400 (6746): 784–787. Бибкод : 1999Natur.400..784W . дои : 10.1038/23506 . PMID 10466730 . S2CID 2841873 .
- ^ Касабов С.Р., Чжан Б., Персингер Дж., Бартоломью Б. (февраль 2003 г.). «SWI/SNF разворачивает, сдвигает и перезаворачивает нуклеосому» . Молекулярная клетка . 11 (2): 391–403. дои : 10.1016/S1097-2765(03)00039-X . ПМИД 12620227 .
- ^ Бруно М., Флаус А., Стокдейл С., Ренкюрел С., Феррейра Х., Оуэн-Хьюз Т. (декабрь 2003 г.). «Обмен димеров гистонов H2A/H2B посредством АТФ-зависимой активности ремоделирования хроматина» . Молекулярная клетка . 12 (6): 1599–1606. дои : 10.1016/S1097-2765(03)00499-4 . ПМЦ 3428624 . ПМИД 14690611 .
- ^ Хавас К., Флаус А., Фелан М., Кингстон Р., Уэйд П.А., Лилли Д.М., Оуэн-Хьюз Т. (декабрь 2000 г.). «Поколение суперспирального кручения за счет АТФ-зависимой активности ремоделирования хроматина» . Клетка . 103 (7): 1133–1142. дои : 10.1016/S0092-8674(00)00215-4 . ПМИД 11163188 . S2CID 7911590 .
- ^ Мизугути Г, Шен Х, Лэндри Дж, Ву ВХ, Сен С, Ву С (январь 2004 г.). «АТФ-обмен варианта гистона H2AZ, катализируемый комплексом ремоделирования хроматина SWR1» . Наука . 303 (5656): 343–348. Бибкод : 2004Sci...303..343M . дои : 10.1126/science.1090701 . PMID 14645854 . S2CID 9881829 .
- ^ Метивье Р., Пено Г., Хюбнер М.Р., Рид Г., Бранд Х., Кос М., Ганнон Ф. (декабрь 2003 г.). «Альфа-рецептор эстрогена управляет упорядоченным, циклическим и комбинаторным набором кофакторов на естественном целевом промоторе» . Клетка . 115 (6): 751–763. дои : 10.1016/S0092-8674(03)00934-6 . ПМИД 14675539 . S2CID 145525 .
- ^ Нагаич А.К., Уокер Д.А., Вулфорд Р., Хагер Г.Л. (апрель 2004 г.). «Быстрое периодическое связывание и смещение глюкокортикоидного рецептора во время ремоделирования хроматина» . Молекулярная клетка . 14 (2): 163–174. дои : 10.1016/S1097-2765(04)00178-9 . ПМИД 15099516 .
- ^ Тейф В.Б., Вайнштейн Й., Кодрон-Хергер М., Мальм Дж.П., Март С., Хефер Т., Риппе К. (ноябрь 2012 г.). «Полногеномное расположение нуклеосом во время развития эмбриональных стволовых клеток». Структурная и молекулярная биология природы . 19 (11): 1185–1192. дои : 10.1038/nsmb.2419 . ПМИД 23085715 . S2CID 34509771 .
- ^ Альберт I, Маврич Т.Н., Томшо Л.П., Ци Дж, Зантон С.Дж., Шустер С.К., Пью Б.Ф. (март 2007 г.). «Настройки трансляции и вращения нуклеосом H2A.Z в геноме Saccharomyces cerevisiae». Природа . 446 (7135): 572–576. Бибкод : 2007Natur.446..572A . дои : 10.1038/nature05632 . ПМИД 17392789 . S2CID 4416890 .
- ^ Ли Б., Кэри М., Уоркман Дж.Л. (февраль 2007 г.). «Роль хроматина во время транскрипции» . Клетка . 128 (4): 707–719. дои : 10.1016/j.cell.2007.01.015 . ПМИД 17320508 . S2CID 1773333 .
- ^ Уайтхаус И., Рандо О.Дж., Делроу Дж., Цукияма Т. (декабрь 2007 г.). «Ремоделирование хроматина на промоторах подавляет антисмысловую транскрипцию». Природа . 450 (7172): 1031–1035. Бибкод : 2007Natur.450.1031W . дои : 10.1038/nature06391 . ПМИД 18075583 . S2CID 4305576 .
- ^ Ли В., Тилло Д., Брэй Н., Морс Р.Х., Дэвис Р.В., Хьюз Т.Р., Нислоу С. (октябрь 2007 г.). «Атлас занятости нуклеосом в дрожжах в высоком разрешении». Природная генетика . 39 (10): 1235–1244. дои : 10.1038/ng2117 . ПМИД 17873876 . S2CID 12816925 .
- ^ Итон М.Л., Галани К., Канг С., Белл С.П., Макэлпайн Д.М. (апрель 2010 г.). «Консервативное расположение нуклеосом определяет начало репликации» . Гены и развитие . 24 (8): 748–753. дои : 10.1101/gad.1913210 . ПМЦ 2854390 . ПМИД 20351051 .
- ^ Шивасвами С., Бхинге А., Чжао Ю., Джонс С., Херст М., Айер В.Р. (март 2008 г.). «Динамическое ремоделирование отдельных нуклеосом эукариотического генома в ответ на транскрипционные нарушения» . ПЛОС Биология . 6 (3): е65. doi : 10.1371/journal.pbio.0060065 . ПМК 2267817 . ПМИД 18351804 .
- ^ Муругесапиллай Д., МакКоли М.Дж., Хуо Р., Нельсон Холте М.Х., Степаньянц А., Махер Л.Дж. и др. (август 2014 г.). «Соединение ДНК и образование петель с помощью HMO1 обеспечивает механизм стабилизации безнуклеосомного хроматина» . Исследования нуклеиновых кислот . 42 (14): 8996–9004. дои : 10.1093/nar/gku635 . ПМЦ 4132745 . ПМИД 25063301 .
- ^ Муругесапиллай Д., Макколи М.Дж., Махер Л.Д., Уильямс М.К. (февраль 2017 г.). «Одномолекулярные исследования высокомобильных белков, изгибающих архитектурную ДНК группы В» . Биофизические обзоры . 9 (1): 17–40. дои : 10.1007/s12551-016-0236-4 . ПМЦ 5331113 . ПМИД 28303166 .
- ^ Вингер Дж., Нодельман И.М., Левендоски Р.Ф., Боуман Г.Д. (май 2018 г.). «Механизм твист-дефекта АТФ-зависимой транслокации нуклеосомной ДНК» . электронная жизнь . 7 : е34100. doi : 10.7554/eLife.34100 . ПМК 6031429 . ПМИД 29809147 .
- ^ Боуман Г.Д. (август 2019 г.). «Открытие нового шага в скользящих нуклеосомах» . Тенденции биохимических наук . 44 (8): 643–645. дои : 10.1016/j.tibs.2019.05.001 . ПМК 7092708 . ПМИД 31171402 .
- ^ Перейти обратно: а б Нодельман И.М., Боуман Г.Д. (май 2021 г.). «Биофизика ремоделирования хроматина» . Ежегодный обзор биофизики . 50 (1): 73–93. doi : 10.1146/annurev-biophys-082520-080201 . ПМЦ 8428145 . ПМИД 33395550 .
- ^ Перейти обратно: а б Брандани ГБ, Такада С (ноябрь 2018 г.). Онуфриев А (ред.). «Ремоделеры хроматина сочетают движение дюймового червя с образованием искривленных дефектов, чтобы сдвинуть нуклеосомную ДНК» . PLOS Вычислительная биология . 14 (11): e1006512. Бибкод : 2018PLSCB..14E6512B . дои : 10.1371/journal.pcbi.1006512 . ПМК 6237416 . ПМИД 30395604 .
- ^ Хейс Джей-Джей, Ли К.М. (май 1997 г.). «Восстановление и анализ мононуклеосом in vitro, содержащих определенные ДНК и белки» . Методы . 12 (1): 2–9. дои : 10.1006/meth.1997.0441 . ПМИД 9169189 .
- ^ Дайер П.Н., Эдайятхумангалам Р.С., Уайт С.Л., Бао Й., Чакраварти С., Мутураджан У.М., Люгер К. (2004). «Восстановление ядерных частиц нуклеосомы из рекомбинантных гистонов и ДНК». Хроматин и ферменты ремоделирования хроматина, часть А. Методы энзимологии. Том. 375. стр. 23–44. дои : 10.1016/s0076-6879(03)75002-2 . ISBN 9780121827793 . ПМИД 14870657 .
- ^ Енидунья А., Дэйви С., Кларк Д., Фельзенфельд Г., Аллан Дж. (апрель 1994 г.). «Позиционирование нуклеосом на промоторах генов глобина курицы и человека in vitro. Новые методы картирования». Журнал молекулярной биологии . 237 (4): 401–414. дои : 10.1006/jmbi.1994.1243 . ПМИД 8151701 .
- ^ Фроуз Т.Д., Барт П.Д., Ричмонд Т.Дж. (январь 2018 г.). «Сайт-специфические дисульфидно-сшитые нуклеосомы с повышенной стабильностью» . Журнал молекулярной биологии . 430 (1): 45–57. дои : 10.1016/j.jmb.2017.10.029 . ПМЦ 5757783 . ПМИД 29113904 .
- ^ Ференц А.Е., Вердин Г.Л. (1994). «Подход с конвертируемыми нуклеозидами: структурная инженерия нуклеиновых кислот путем дисульфидной сшивки». В Экстайне Ф., Лилли Д.М. (ред.). Нуклеиновые кислоты и молекулярная биология . Том. 8. стр. 14–40. дои : 10.1007/978-3-642-78666-2_2 . ISBN 978-3-642-78668-6 .
- ^ Ямасу К., Сэнсю Т. (январь 1990 г.). «Консервативное разделение тетрамерных единиц гистонов H3 и H4 во время репликации нуклеосомы». Журнал биохимии . 107 (1): 15–20. doi : 10.1093/oxfordjournals.jbchem.a122999 . ПМИД 2332416 .
- ^ Кауфман П.Д., Кобаяши Р., Кесслер Н., Стиллман Б. (июнь 1995 г.). «Субъединицы p150 и p60 фактора сборки хроматина I: молекулярная связь между вновь синтезированными гистонами и репликацией ДНК» . Клетка . 81 (7): 1105–1114. дои : 10.1016/S0092-8674(05)80015-7 . ПМИД 7600578 . S2CID 13502921 .
- ^ Тайлер Дж.К., Адамс С.Р., Чен С.Р., Кобаяши Р., Камакака Р.Т., Кадонага Дж.Т. (декабрь 1999 г.). «Комплекс RCAF опосредует сборку хроматина во время репликации и репарации ДНК». Природа . 402 (6761): 555–560. Бибкод : 1999Natur.402..555T . дои : 10.1038/990147 . ПМИД 10591219 . S2CID 205097512 .
- ^ Бенсон Л.Дж., Гу Ю., Яковлева Т., Тонг К., Барроуз С., Страк К.Л. и др. (апрель 2006 г.). «Модификации H3 и H4 во время репликации хроматина, сборки нуклеосом и обмена гистонов» . Журнал биологической химии . 281 (14): 9287–9296. дои : 10.1074/jbc.M512956200 . ПМИД 16464854 .
- ^ Лутерс Л., Чалкли Р. (июнь 1985 г.). «Обмен гистонов H1, H2A и H2B in vivo». Биохимия . 24 (13): 3080–3085. дои : 10.1021/bi00334a002 . ПМИД 4027229 .
- ^ Пак Й.Дж., Чодапарамбил Й.В., Бао Й., МакБрайант С.Дж., Люгер К. (январь 2005 г.). «Белок сборки нуклеосомы 1 обменивает димеры гистонов H2A-H2B и способствует скольжению нуклеосомы» . Журнал биологической химии . 280 (3): 1817–1825. дои : 10.1074/jbc.M411347200 . ПМИД 15516689 .
- ^ Винсент Дж.А., Квонг Т.Дж., Цукияма Т. (май 2008 г.). «АТФ-зависимое ремоделирование хроматина формирует ландшафт репликации ДНК» . Структурная и молекулярная биология природы . 15 (5): 477–484. дои : 10.1038/nsmb.1419 . ПМЦ 2678716 . ПМИД 18408730 .
- ^ Ядав Т., Белый дом I (апрель 2016 г.). «Сборка и позиционирование нуклеосом, связанных с репликацией, с помощью АТФ-зависимых ферментов, ремоделирующих хроматин» . Отчеты по ячейкам . 15 (4): 715–723. doi : 10.1016/J.CELREP.2016.03.059 . ПМК 5063657 . ПМИД 27149855 .
Внешние ссылки
[ редактировать ]- MBInfo - Что такое нуклеосомы
- Нуклеосомы на сайте Richmond Lab
- Протеопедия Нуклеосомы
- Нуклеосома в PDB
- Динамическое ремоделирование отдельных нуклеосом в геноме эукариот в ответ на транскрипционные нарушения
- Данные и инструменты позиционирования нуклеосом онлайн (аннотированный список, постоянно обновляемый)
- Структура белка гистонов
- HistoneDB 2.0 - База данных гистонов и вариантов в NCBI