Конформационная корректура
Конформационная корректура или конформационный отбор - это общий механизм систем молекулярного распознавания , предложенный Йонатаном Савиром и Цви Тлусти, в котором введение энергетического барьера - такого как структурное несоответствие между молекулярным распознавателем и его целью - повышает специфичность и качество распознавания. [1] [2] [3] [4] [5] [6] Конформационная корректура не требует затрат энергии и поэтому может использоваться в любой системе молекулярного распознавания. Конформационная корректура особенно полезна в сценариях, когда распознавателю приходится выбирать подходящую цель среди множества аналогичных конкурентов. Белки развивают способность к конформационной корректуре посредством точной настройки своей геометрии, гибкости и химического взаимодействия с мишенью. [7]
Балансировка правильной и неправильной привязки
[ редактировать ]Молекулярное распознавание происходит в шумной, переполненной биологической среде, и распознавателю часто приходится справляться с задачей выбора своей цели среди множества аналогичных конкурентов. Например, рибосома должна выбрать правильную тРНК , соответствующую кодону мРНК, среди множества структурно сходных тРНК. Если распознаватель и его правильная цель идеально совпадают, как замок и ключ , то вероятность привязки будет высокой, поскольку при привязке не требуется никакой деформации. В то же время распознаватель с высокой вероятностью может привязываться и к конкуренту с аналогичной структурой. Введение энергетического барьера , в частности структурного несоответствия между распознавателем (замком) и ключом, снижает вероятность привязки к правильной цели, но еще больше снижает вероятность привязки к аналогичной неправильной цели и, таким образом, повышает специфичность. [1] [2] [3] [7] Тем не менее, введение слишком большой деформации резко снижает вероятность привязки к правильной цели. Следовательно, оптимальный баланс между максимизацией вероятности правильной привязки и минимизацией вероятности неправильной привязки достигается, когда распознаватель немного отклоняется от цели . Это предполагает, что конформационные изменения во время процессов молекулярного узнавания, такие как индуцированное соответствие [8] механизм, выгодны для повышения специфичности распознавания. Такие конформационные изменения могут быть тонко настроены с помощью мутаций, которые влияют на механический ответ узнавателя, в том числе и в положениях, далеких от места связывания. [7]

Использование путем гомологичной рекомбинации для поиска гомологии.
[ редактировать ]Механизм конформационной корректуры используется в системе гомологичной рекомбинации для различения сходных последовательностей ДНК. [3] [4] Гомологичная рекомбинация облегчает обмен генетическим материалом между гомологичными молекулами ДНК. Этот решающий процесс требует обнаружения конкретной гомологичной последовательности ДНК среди огромного разнообразия гетерологичных последовательностей. Обнаружение опосредовано RecA в E. coli или членами его надсемейства в других организмах. RecA сначала полимеризуется на участке одноцепочечной ДНК, а затем эта нить белок-ДНК ищет гомологию вдоль двухцепочечной ДНК. В нити RecA-ДНК расстояние между основаниями значительно увеличивается по сравнению с голыми 3,4 Å в двухцепочечной (в среднем на 50%). [9] ). Это устанавливает значительный энергетический барьер для поиска, поскольку двухцепочечная ДНК должна растягиваться на такую же величину, чтобы проверить гомологию. Сформулировав процесс распознавания ДНК как задачу обнаружения сигнала, было показано, что экспериментально наблюдаемая деформация ДНК, индуцированная RecA, и энергетика связывания точно настроены для обеспечения оптимального обнаружения последовательности. Степень деформации такова, что связывание с гомологичными последовательностями ДНК уменьшается лишь незначительно, тогда как связывание с неправильными последовательностями снижается значительно. Это и есть механизм конформационной корректуры.
Экспериментальные доказательства конформационной корректуры путем гомологичной рекомбинации
[ редактировать ]Группа К. Деккера (Дельфтский университет) непосредственно исследовала взаимодействия, участвующие в поиске гомологии, комбинируя магнитный и оптический пинцеты. [10] Они обнаружили, что поиск и распознавание гомологии требует раскрытия спирали и, следовательно, может быть ускорено за счет раскручивания ДНК. Это именно тот энергетический барьер, который предсказывает модель конформационной корректуры. Данные указывают на физическую картину распознавания гомологии, в которой точность процесса поиска определяется расстоянием между сайтами связывания ДНК. Авторы приходят к выводу, что их интерпретация измерений «сродни схеме конформационной корректуры … где дцДНК, а не нить RecA, является активным, распознающим поисковым объектом. Между связанной с мишенью и несвязанной мишенью существует большое конформационное несоответствие. Доступ к целевому состоянию дцДНК осуществляется через энергетически невыгодные промежуточные состояния, как обсуждалось выше. Конформационное несоответствие улучшает селективность реакции распознавания». Другими словами, они определили энергетический барьер и показали, что двухцепочечная ДНК действительно является активным участником, поскольку ей необходимо преодолеть этот барьер.
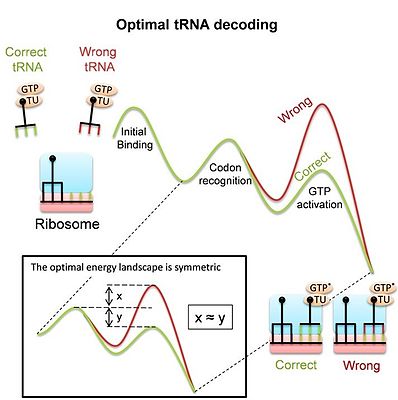
Использование рибосомой для декодирования тРНК
[ редактировать ]Рибосома путем представляет собой сложную молекулярную машину, которая для синтеза белков в процессе трансляции должна декодировать мРНК спаривания их кодонов с соответствующими тРНК . Декодирование является основным фактором, определяющим приспособленность, и требует точного и быстрого выбора правильных тРНК среди множества аналогичных конкурентов. Следует иметь в виду, что большинство событий связывания происходит за счет несовпадающих («неродственных») тРНК, и рибосоме необходимо отвергнуть их как можно быстрее, чтобы освободить сайт связывания. В то же время рибосома должна удерживать соответствующие тРНК связанными достаточно долго, чтобы начался процесс синтеза белка. Несмотря на важность декодирования тРНК, до недавнего времени было неясно, являются ли современные рибосомы и, в частности, ее большие конформационные изменения во время декодирования результатом адаптации к своей задаче в качестве декодера или результатом других ограничений. Недавнее исследование [5] получили энергетический ландшафт, который обеспечивает оптимальное распознавание конкурирующих субстратов тРНК и, следовательно, оптимальное декодирование тРНК. Оптимальный ландшафт — симметричный (см. изображение). Исследование показывает, что измеренный ландшафт прокариотической рибосомы действительно симметричен . Эта модель предполагает, что конформационные изменения рибосомы и тРНК при декодировании являются средством получения такого оптимального декодера тРНК. Тот факт, что и гомологичная рекомбинация, и декодирование тРНК используют конформационную корректировку, предполагает, что это общий механизм, который может широко использоваться системами молекулярного распознавания.
В других биологических системах
[ редактировать ]Ремонт повреждений, вызванных ультрафиолетом человека
[ редактировать ]Недавнее исследование показывает, что конформационная корректура используется механизмами репарации ДНК человека. [11] Исследование было сосредоточено на вопросе о том, как белки репарации ДНК сканируют геном человека на предмет повреждений, вызванных ультрафиолетом, на начальном этапе эксцизионного восстановления нуклеотидов (NER). Детальные измерения одиночных молекул показали, как поврежденный УФ-излучением ДНК-связывающий белок человека (UV-DDB) выполняет трехмерный поиск. Авторы обнаружили, что «UV-DDB исследует сайты на ДНК дискретными шагами, прежде чем сформировать долгоживущие неподвижные димеры UV-DDB ( DDB1 - DDB2 ) 2 в местах повреждения. Анализ скоростей диссоциации для временных связывающих молекул на обоих неповрежденная и поврежденная ДНК демонстрирует многократное время пребывания, превышающее три порядка... Считается, что эти промежуточные состояния представляют собой дискретные конформеры UV-DDB на траектории стабильного обнаружения повреждений». На основании детальных кинетических измерений авторы пришли к выводу, что UV-DDB распознает повреждения, используя механизм конформационной корректировки через множество промежуточных соединений.
Другие схемы признания
[ редактировать ]Связь с кинетической корректурой
[ редактировать ]В кинетической корректуре [12] [13] В схеме вводится временная задержка (т. е. необратимый промежуточный этап) при образовании правильных или неправильных комплексов. Эта временная задержка снижает производительность обоих комплексов, но повышает точность за пределами равновесного предела. Необратимость схемы требует источника энергии. Временная задержка при кинетической корректуре аналогична пространственной разнице при конформационной корректуре. Однако конформационная корректура может представлять собой равновесную схему, не потребляющую энергии.
Ссылки
[ редактировать ]- ^ Jump up to: а б Савир Ю., Тлусти Т. (май 2007 г.). Скалас Э (ред.). «Конформационная корректура: влияние конформационных изменений на специфику молекулярного распознавания» . ПЛОС ОДИН . 2 (5): е468. Бибкод : 2007PLoSO...2..468S . дои : 10.1371/journal.pone.0000468 . ПМЦ 1868595 . ПМИД 17520027 .
- ^ Jump up to: а б Савир Ю., Тлусти Т. (2008). «Оптимальная конструкция молекулярного распознавателя: молекулярное распознавание как проблема обнаружения байесовского сигнала». IEEE J Sel Темы Сигнальный процесс . 2 (3): 390–399. arXiv : 1007.4527 . Бибкод : 2008ИССП...2..390С . дои : 10.1109/JSTSP.2008.923859 . S2CID 7210763 .
- ^ Jump up to: а б с д Савир Ю., Тлусти Т. (ноябрь 2010 г.). «Поиск гомологии, опосредованный RecA, как почти оптимальная система обнаружения сигналов». Молекулярная клетка . 40 (3): 388–396. arXiv : 1011.4382 . doi : 10.1016/j.molcel.2010.10.020 . ПМИД 21070965 . S2CID 1682936 .
- ^ Jump up to: а б с Рэмбо Р.П., Уильямс Дж.Дж., Тайнер Дж.А. (ноябрь 2010 г.). «Достижение точности гомологичной рекомбинации, несмотря на чрезвычайную сложность: обоснованные решения с помощью молекулярного профилирования» . Молекулярная клетка . 40 (3): 347–348. doi : 10.1016/j.molcel.2010.10.032 . ПМК 3003302 . ПМИД 21070960 .
- ^ Jump up to: а б с Савир Ю., Тлусти Т. (апрель 2013 г.). «Рибосома как оптимальный декодер: урок молекулярного распознавания» . Клетка . 153 (2): 471–479. Бибкод : 2013APS..MARY46006T . дои : 10.1016/j.cell.2013.03.032 . ПМИД 23582332 .
- ^ Алон У (2008). «Журнал-клуб» . Природа . 453 (7196): 701. Бибкод : 2008Natur.453..701A . дои : 10.1038/453701e . S2CID 29639642 .
- ^ Jump up to: а б с Макбрайд Дж. М., Экманн Дж. П., Тласти Т. (ноябрь 2022 г.). Эчаве Дж. (ред.). «Общая теория специфического связывания: данные генетико-механо-химической модели белка» . Молекулярная биология и эволюция . 39 (11): msac217. дои : 10.1093/molbev/msac217 . ПМК 9641994 . ПМИД 36208205 .
- ^ Кошланд Д.Э. (февраль 1958 г.). «Применение теории специфичности ферментов к синтезу белка» . Труды Национальной академии наук Соединенных Штатов Америки . 44 (2): 98–104. Бибкод : 1958ПНАС...44...98К . дои : 10.1073/pnas.44.2.98 . ПМЦ 335371 . ПМИД 16590179 .
- ^ Чен З, Ян Х, Павлетич Н.П. (май 2008 г.). «Механизм гомологичной рекомбинации структур RecA-ssDNA/dsDNA». Природа . 453 (7194): 489–484. Бибкод : 2008Natur.453..489C . дои : 10.1038/nature06971 . ПМИД 18497818 . S2CID 4416531 .
- ^ Де Вламинк И., ван Лёнхаут М.Т., Цвайфель Л., ден Бланкен Дж., Хунинг К., Хаге С. и др. (июнь 2012 г.). «Механизм распознавания гомологии при рекомбинации ДНК в экспериментах с двумя молекулами» . Молекулярная клетка . 46 (5): 616–624. doi : 10.1016/j.molcel.2012.03.029 . ПМИД 22560720 .
- ^ Годке Х., Ван Х., Се К.Л., Вольдемескель С., Уоткинс С.К., Рапич-Отрин В., Ван Хаутен Б. (май 2014 г.). «Анализ одиночных молекул показывает, что поврежденный УФ-излучением ДНК-связывающий белок человека (UV-DDB) димеризуется на ДНК посредством множества кинетических промежуточных продуктов» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (18): E1862–E1871. Бибкод : 2014PNAS..111E1862G . дои : 10.1073/pnas.1323856111 . ПМК 4020048 . ПМИД 24760829 .
- ^ Хопфилд Джей-Джей (октябрь 1974 г.). «Кинетическая корректура: новый механизм уменьшения ошибок в процессах биосинтеза, требующий высокой специфичности» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (10): 4135–4139. Бибкод : 1974PNAS...71.4135H . дои : 10.1073/pnas.71.10.4135 . ПМК 434344 . ПМИД 4530290 .
- ^ Нинио Дж (1975). «Кинетическая амплификация дискриминации ферментов». Биохимия . 57 (5): 587–595. дои : 10.1016/S0300-9084(75)80139-8 . ПМИД 1182215 .