ДНК-нанотехнологии
| Часть серии статей о |
| Молекулярная самосборка |
|---|
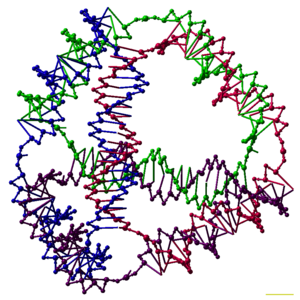
ДНК-нанотехнология — это разработка и производство искусственных структур нуклеиновых кислот для технологических целей. В этой области нуклеиновые кислоты используются как небиологические инженерные материалы для нанотехнологий, а не как носители генетической информации в живых клетках . Исследователи в этой области создали статические структуры, такие как двух- и трехмерные кристаллические решетки , нанотрубки , многогранники и произвольные формы, а также функциональные устройства, такие как молекулярные машины и ДНК-компьютеры . Эта область начинает использоваться в качестве инструмента для решения фундаментальных научных проблем в структурной биологии и биофизике , включая приложения в рентгеновской кристаллографии и спектроскопии ядерного магнитного резонанса белков для определения структур. потенциальные применения в электронике молекулярного масштаба и наномедицине Также исследуются .
Концептуальная основа ДНК-нанотехнологии была впервые заложена Надрианом Симаном в начале 1980-х годов, а широкий интерес к этой области начал привлекать в середине 2000-х годов. Такое использование нуклеиновых кислот возможно благодаря их строгим правилам спаривания оснований только части цепей с комплементарными последовательностями оснований , которые заставляют связываться вместе двойной спирали , образуя прочные, жесткие структуры . Это позволяет рационально разрабатывать базовые последовательности , которые будут избирательно собираться в сложные целевые структуры с точно контролируемыми наноразмерными характеристиками. Для создания этих структур используется несколько методов сборки, в том числе структуры на основе плиток, которые собираются из более мелких структур, складные структуры с использованием метода ДНК-оригами и динамически реконфигурируемые структуры с использованием методов смещения цепей. Название области конкретно отсылает к ДНК , но те же принципы использовались и с другими типами нуклеиновых кислот, что привело к периодическому использованию альтернативного названия « Нанотехнология нуклеиновых кислот». .
История [ править ]
Концептуальная основа нанотехнологии ДНК была впервые заложена Надрианом Зееманом в начале 1980-х годов. [2] Первоначальной мотивацией Симана было создание трехмерной решетки ДНК для ориентации других крупных молекул, что упростило бы их кристаллографическое исследование за счет устранения сложного процесса получения чистых кристаллов. Сообщается, что эта идея пришла к нему в конце 1980 года, после того как он осознал сходство между гравюрой на дереве «Глубина» М. К. Эшера и массивом шестилучевых соединений ДНК. [3] [4] В то время было известно несколько природных разветвленных структур ДНК, в том числе репликационная вилка ДНК и мобильное соединение Холлидея , но идея Симана заключалась в том, что неподвижные соединения нуклеиновых кислот могут быть созданы путем правильного проектирования последовательностей цепей для устранения симметрии в собранной молекуле, и что эти неподвижные соединения в принципе можно было объединить в жесткие кристаллические решетки. Первая теоретическая статья, предлагающая эту схему, была опубликована в 1982 году, а первая экспериментальная демонстрация неподвижного соединения ДНК была опубликована в следующем году. [5] [6]

В 1991 году лаборатория Симана опубликовала отчет о синтезе куба из ДНК, первой синтетической трехмерной наноструктуры нуклеиновой кислоты, за что он получил премию Фейнмана 1995 года в области нанотехнологий . За этим последовал усеченный ДНК октаэдр . Вскоре стало ясно, что эти структуры, представляющие собой многоугольные формы с гибкими соединениями в вершинах , недостаточно жесткие, чтобы образовывать протяженные трехмерные решетки. двойного пересечения (DX) Симан разработал более жесткий структурный мотив , а в 1998 году в сотрудничестве с Эриком Уинфри опубликовал создание двумерных решеток из плиток DX. [3] [2] [7] Преимущество этих структур на основе плиток заключалось в том, что они обеспечивали возможность реализации вычислений на ДНК, что было продемонстрировано Уинфри и Полом Ротемундом в их статье 2004 года об алгоритмической самосборке структуры прокладок Серпинского, и для которой они поделились работой Фейнмана 2006 года. Премия в области нанотехнологий. Ключевая идея Уинфри заключалась в том, что плитки DX можно использовать как плитки Ванга , а это означает, что их сборка может выполнять вычисления. [2] Синтез трехмерной решетки был наконец опубликован Симаном в 2009 году, почти через тридцать лет после того, как он намеревался достичь этого. [8]
На протяжении 2000-х годов продолжали открываться новые способности созданных структур ДНК. Первая наномашина ДНК — мотив, который меняет свою структуру в ответ на входной сигнал — была продемонстрирована в 1999 году Симаном. Усовершенствованная система, которая была первым устройством на основе нуклеиновых кислот, в котором использовалось смещение цепи, опосредованное опорой, было продемонстрировано Бернардом Юрком в 2000 году. [9] Следующим достижением стал перевод этого в механическое движение, и в 2004 и 2005 годах несколько систем передвижения по ДНК были продемонстрированы группами Симана, Найлза Пирса , Эндрю Терберфилда и Чэндэ Мао . [10] Идея использования массивов ДНК для создания шаблонов сборки других молекул, таких как наночастицы и белки, впервые была предложена Брюшем Робинсоном и Симаном в 1987 году. [11] был продемонстрирован в 2002 году Seeman, Kiehl et al. [12] и впоследствии многими другими группами.
В 2006 году Ротемунд впервые продемонстрировал метод ДНК-оригами, позволяющий легко и надежно формировать свернутые структуры ДНК произвольной формы. Ротемунд задумал этот метод как концептуально промежуточный между решетками DX Симана, в которых использовалось множество коротких нитей, и Уильяма Ши октаэдром ДНК , который состоял в основном из одной очень длинной нити. ДНК-оригами Ротемунда содержит длинную нить, сворачиванию которой способствуют несколько коротких нитей. Этот метод позволил сформировать гораздо более крупные структуры, чем это было возможно раньше, и которые менее технически сложны в проектировании и синтезе. [7] ДНК-оригами стало обложкой журнала Nature от 15 марта 2006 года. [13] За исследованием Ротемунда, демонстрирующим двумерные структуры ДНК-оригами, последовала демонстрация твердого трехмерного ДНК-оригами Дугласом и др. в 2009 году, [14] в то время как лаборатории Йоргена Кьемса и Яна продемонстрировали полые трехмерные структуры, состоящие из двухмерных граней. [8]
Нанотехнология ДНК изначально была встречена с некоторым скептицизмом из-за необычного небиологического использования нуклеиновых кислот в качестве материалов для построения структур и выполнения вычислений, а также преобладания экспериментов по доказательству принципов , которые расширяли возможности этой области, но были далеки от реального применения. Статья Симана 1991 года о синтезе куба ДНК была отклонена журналом Science после того, как один рецензент похвалил ее оригинальность, а другой раскритиковал ее за отсутствие биологической значимости. [15] К началу 2010-х годов считалось, что эта область расширила свои возможности до такой степени, что начали реализовываться приложения для фундаментальных научных исследований, а практическое применение в медицине и других областях стало считаться возможным. [8] [16] Эта область выросла с очень небольшого количества действующих лабораторий в 2001 году до как минимум 60 в 2010 году, что увеличило кадровый резерв и, следовательно, количество научных достижений в этой области за это десятилетие. [17]
Фундаментальные понятия [ править ]

Свойства нуклеиновых кислот [ править ]
Нанотехнологию часто определяют как исследование материалов и устройств с характеристиками в масштабе менее 100 нанометров . Нанотехнология ДНК, в частности, является примером снизу вверх молекулярной самосборки , при которой молекулярные компоненты спонтанно организуются в стабильные структуры; Особая форма этих структур обусловлена физическими и химическими свойствами выбранных проектировщиками компонентов. [19] В ДНК-нанотехнологии компонентами материалов являются нити нуклеиновых кислот, таких как ДНК; эти нити часто являются синтетическими и почти всегда используются вне контекста живой клетки. ДНК хорошо подходит для наномасштабного конструирования, поскольку связывание между двумя нитями нуклеиновой кислоты зависит от простых правил спаривания оснований , которые хорошо понятны и образуют специфическую наноразмерную структуру двойной спирали нуклеиновой кислоты . Эти качества позволяют легко контролировать сборку структур нуклеиновой кислоты посредством дизайна нуклеиновой кислоты . Это свойство отсутствует у других материалов, используемых в нанотехнологиях, включая белки , для которых разработка белков очень сложна, и наночастицы , которые не обладают способностью к специфической сборке самостоятельно. [5]
Структура молекулы нуклеиновой кислоты состоит из последовательности нуклеотидов, отличающихся тем, какое азотистое основание они содержат. В ДНК присутствуют четыре основания: аденин (А), цитозин (С), гуанин (G) и тимин (Т). Нуклеиновые кислоты обладают свойством, заключающимся в том, что две молекулы связываются друг с другом с образованием двойной спирали только в том случае, если две последовательности комплементарны , что означает, что они образуют совпадающие последовательности пар оснований, при этом A связывается только с T, а C - только с G. [5] [20] Поскольку образование правильно спаренных пар оснований энергетически выгодно , ожидается, что в большинстве случаев цепи нуклеиновой кислоты будут связываться друг с другом в конформации, которая максимизирует количество правильно спаренных оснований. Таким образом, последовательности оснований в системе нитей легко контролируемым образом определяют характер связывания и общую структуру. В нанотехнологии ДНК последовательности оснований цепей рационально разрабатываются исследователями так, что взаимодействия спаривания оснований заставляют цепи собираться в желаемую конформацию. [3] [5] Хотя ДНК является доминирующим используемым материалом, структуры, включающие другие нуклеиновые кислоты, такие как РНК и пептидно-нуклеиновые кислоты (ПНК). также были созданы [21] [22]
Подполя [ править ]
Нанотехнологию ДНК иногда делят на две пересекающиеся области: структурную нанотехнологию ДНК и динамическую нанотехнологию ДНК. Структурная нанотехнология ДНК, иногда сокращенно называемая SDN, фокусируется на синтезе и характеристике комплексов нуклеиновых кислот и материалов, которые собираются в статическое, равновесное конечное состояние. С другой стороны, нанотехнология динамической ДНК фокусируется на комплексах с полезным неравновесным поведением, например способностью реконфигурироваться на основе химического или физического стимула. Некоторые комплексы, например наномеханические устройства на основе нуклеиновых кислот, сочетают в себе черты как структурного, так и динамического подполей. [23] [24]
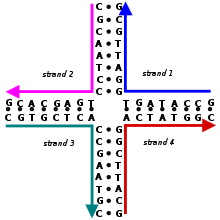
Комплексы, построенные с помощью структурной нанотехнологии ДНК, используют топологически разветвленные структуры нуклеиновых кислот, содержащие соединения. (Напротив, большая часть биологической ДНК существует в виде неразветвленной двойной спирали .) Одной из простейших разветвленных структур является четырехветвевое соединение, состоящее из четырех отдельных цепей ДНК, части которых комплементарны по определенному образцу. В отличие от естественных соединений Холлидея , каждое плечо в искусственном неподвижном четырехветвевом соединении имеет различную последовательность оснований , в результате чего точка соединения фиксируется в определенном положении. Множественные соединения могут быть объединены в один и тот же комплекс, например, в широко используемом структурном мотиве двойного кроссовера (DX) , который содержит два параллельных двойных спиральных домена с отдельными цепями, пересекающимися между доменами в двух точках кроссовера. Каждая точка кроссовера топологически представляет собой четырехплечевое соединение, но ограничена одной ориентацией, в отличие от гибкого одиночного четырехплечевого соединения, что обеспечивает жесткость, которая делает мотив DX подходящим в качестве структурного строительного блока для более крупных комплексов ДНК. [3] [5]
Динамическая нанотехнология ДНК использует механизм, называемый смещением цепи, опосредованным фиксацией , чтобы позволить комплексам нуклеиновых кислот реконфигурироваться в ответ на добавление новой цепи нуклеиновой кислоты. В этой реакции входящая цепь связывается с одноцепочечной опорной областью двухцепочечного комплекса, а затем вытесняет одну из цепей, связанных в исходном комплексе, посредством процесса миграции ветвей . Общий эффект заключается в том, что одна из нитей комплекса заменяется другой. [23] Кроме того, реконфигурируемые структуры и устройства могут быть созданы с использованием функциональных нуклеиновых кислот, таких как дезоксирибозимы и рибозимы , которые могут выполнять химические реакции, и аптамеры , которые могут связываться со специфическими белками или небольшими молекулами. [25]
ДНК Структурная нанотехнология
Структурная нанотехнология ДНК, иногда сокращенно называемая SDN, фокусируется на синтезе и характеристике комплексов нуклеиновых кислот и материалов, сборка которых имеет статическую, равновесную конечную точку. имеет Двойная спираль нуклеиновой кислоты надежную, определенную трехмерную геометрию, которая позволяет моделировать, [26] прогнозировать и проектировать структуры более сложных комплексов нуклеиновых кислот. Создано множество таких структур, в том числе двух- и трехмерные, а также периодические, апериодические и дискретные. [24]
Расширенные решетки [ править ]
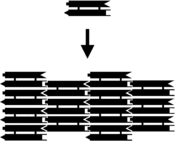




Небольшие комплексы нуклеиновых кислот могут быть снабжены липкими концами и объединены в более крупные двумерные периодические решетки, содержащие определенный мозаичный рисунок отдельных молекулярных плиток. [24] Самый ранний пример этого использовал комплексы двойного кроссовера (DX) в качестве основных плиток, каждый из которых содержал четыре липких конца, созданных с последовательностями, которые заставляли единицы DX объединяться в периодические двумерные плоские листы, которые по сути представляют собой жесткие двумерные кристаллы ДНК. . [30] [31] Двумерные массивы были созданы и из других мотивов, в том числе из решетки Холлидея ромбовидной , [32] и различные массивы на основе DX, использующие схему двойной связи. [33] [34] На двух верхних изображениях справа показаны примеры периодических решеток на основе плиток.
Можно создать двумерные массивы, демонстрирующие апериодические структуры, сборка которых реализует определенный алгоритм, демонстрирующий одну из форм вычислений на ДНК. [17] У плиток DX можно выбрать липкие конечные последовательности, чтобы они действовали как плитки Ванга , что позволяет им выполнять вычисления. массив DX, сборка которого кодирует операцию XOR Был продемонстрирован ; это позволяет массиву ДНК реализовать клеточный автомат , который генерирует фрактал, известный как прокладка Серпинского . Третье изображение справа показывает этот тип массива. [29] Другая система имеет функцию двоичного счетчика , отображающего представление возрастающих двоичных чисел по мере их роста. Эти результаты показывают, что вычисления могут быть включены в сборку массивов ДНК. [35]
Массивы DX были созданы для формирования полых нанотрубок диаметром 4–20 нм , по сути, двумерных решеток, которые изгибаются сами по себе. [36] Эти нанотрубки ДНК по размеру и форме чем-то похожи на углеродные нанотрубки , и хотя им не хватает электропроводности углеродных нанотрубок, нанотрубки ДНК легче модифицируются и соединяются с другими структурами. В одной из многих схем построения нанотрубок ДНК используется решетка из изогнутых плиток DX, которые закручиваются вокруг себя и замыкаются в трубку. [37] В альтернативном методе, который позволяет задавать длину окружности простым модульным способом с использованием однониточных плиток, жесткость трубки является новым свойством . [38]
Формирование трехмерных решеток ДНК было самой ранней целью ДНК-нанотехнологии, но ее оказалось одной из самых трудных для реализации. В 2009 году наконец было сообщено об успехе использования мотива, основанного на концепции тенсегрити , баланса между силами напряжения и сжатия. [17] [39]
Дискретные структуры [ править ]
Исследователи синтезировали множество трехмерных комплексов ДНК, каждый из которых имеет связность многогранника , например куба или октаэдра . Это означает, что дуплексы ДНК прослеживают края многогранника с соединением ДНК в каждой вершине. [6] Самые ранние демонстрации многогранников ДНК были очень трудоемкими: требовалось многократное лигирование и твердофазного синтеза этапы для создания составных многогранников. [40] Последующие работы привели к созданию многогранников, синтез которых оказался намного проще. К ним относятся октаэдр ДНК, состоящий из длинной одиночной цепи, предназначенной для сворачивания в правильную конформацию. [41] и тетраэдр, который можно получить из четырех нитей ДНК за один этап, изображенный в верхней части этой статьи. [1]
Наноструктуры произвольной, нерегулярной формы обычно изготавливаются методом ДНК-оригами . Эти структуры состоят из длинной природной вирусной цепи в качестве «каркаса», который складывается в желаемую форму с помощью компьютерно спроектированных коротких «штапельных» цепей. Преимущество этого метода заключается в простоте разработки, поскольку базовая последовательность заранее определяется последовательностью цепи каркаса, и он не требует высокой чистоты цепи и точной стехиометрии , как это делают большинство других методов нанотехнологии ДНК. ДНК-оригами было впервые продемонстрировано для двумерных фигур, таких как смайлик , грубая карта Западного полушария и картина Моны Лизы. [6] [13] [42] Твердые трехмерные структуры можно создавать, используя параллельные спирали ДНК, расположенные в виде сот. [14] а конструкции с двумерными гранями можно складывать в полую трехмерную форму, похожую на картонную коробку. Их можно запрограммировать на открытие и высвобождение молекулярного груза в ответ на раздражитель, что делает их потенциально полезными в качестве программируемых молекулярных клеток . [43] [44]
Шаблонная сборка [ править ]
Структуры нуклеиновых кислот могут быть созданы с включением молекул, отличных от нуклеиновых кислот, иногда называемых гетероэлементами, включая белки, металлические наночастицы, квантовые точки , амины , [45] и фуллерены . Это позволяет создавать материалы и устройства с гораздо большим набором функциональных возможностей, чем это возможно при использовании только нуклеиновых кислот. Цель состоит в том, чтобы использовать самосборку структур нуклеиновых кислот для создания шаблона сборки размещенных на них наночастиц, контролируя их положение, а в некоторых случаях и ориентацию. [6] [46] Многие из этих схем используют схему ковалентного присоединения, используя олигонуклеотиды с амидными или тиоловыми функциональными группами в качестве химического средства для связывания гетероэлементов. Эта схема ковалентного связывания использовалась для размещения наночастиц золота на матрице на основе DX. [47] и расположить молекулы белка стрептавидина в определенные структуры на массиве DX. [48] Схема нековалентного хостинга с использованием полиамидов Дервана на матрице DX использовалась для расположения белков стрептавидина по определенному шаблону на матрице DX. [49] Углеродные нанотрубки размещены на матрицах ДНК по образцу, позволяющему сборке действовать как молекулярное электронное устройство, полевой транзистор из углеродных нанотрубок . [50] Кроме того, существуют методы металлизации нуклеиновой кислоты, при которых нуклеиновая кислота заменяется металлом, который принимает общую форму исходной структуры нуклеиновой кислоты. [51] и схемы использования наноструктур нуклеиновых кислот в качестве литографических масок с переносом их рисунка на твердую поверхность. [52]
ДНК Динамическая нанотехнология

Нанотехнология динамической ДНК направлена на формирование систем нуклеиновых кислот с разработанными динамическими функциональными возможностями, связанными с их общими структурами, такими как вычисления и механическое движение. Существует некоторое совпадение между структурной и динамической нанотехнологией ДНК, поскольку структуры могут быть сформированы путем отжига, а затем динамически реконфигурированы, или их можно вообще заставить формироваться динамически. [6] [10]
Наномеханические устройства [ править ]
Были созданы комплексы ДНК, которые меняют свою конформацию под воздействием некоторых стимулов, что делает их одной из форм наноробототехники . Эти структуры изначально формируются так же, как статические структуры, созданные с помощью структурной нанотехнологии ДНК, но сконструированы так, что после первоначальной сборки возможна динамическая реконфигурация. [23] [10] Самое раннее такое устройство использовало переход между формами B-ДНК и Z-ДНК , чтобы реагировать на изменение буферных условий, совершая скручивающее движение. [53] Эта зависимость от условий буфера привела к одновременному изменению состояния всех устройств. Последующие системы могли менять состояния в зависимости от наличия цепей управления, что позволяло нескольким устройствам работать независимо друг от друга. Некоторыми примерами таких систем являются конструкция «молекулярного пинцета», имеющая открытое и закрытое состояние. [54] устройство, которое могло бы переключаться с конформации паранемического кроссинговера (PX) на конформацию (JX2) с двумя непересекающимися соединениями основной цепи ДНК, совершающими при этом вращательное движение, [55] и двумерный массив, который может динамически расширяться и сжиматься в ответ на управляющие нити. [56] Также были созданы структуры, которые динамически открываются и закрываются, потенциально действуя как молекулярная клетка, высвобождающая или раскрывающая функциональный груз при открытии. [43] [57] [58] В другом примере наноструктура ДНК-оригами была соединена с РНК-полимеразой Т7 и, таким образом, могла работать как двигатель, управляемый химической энергией, который можно соединить с пассивным последователем, которым он затем управляет. [59]
ДНК-ходоки — это класс наномашин из нуклеиновых кислот, которые демонстрируют направленное движение по линейному пути. Было продемонстрировано большое количество схем. [10] Одна из стратегий заключается в управлении движением ходунка по дорожке с помощью управляющих нитей, которые необходимо последовательно добавлять вручную. [60] [61] Также возможно контролировать отдельные шаги ДНК-ходока путем облучения светом разной длины волны. [62] Другой подход заключается в использовании ферментов рестрикции или дезоксирибозимов, чтобы расщепить нити и заставить ходунка двигаться вперед, что имеет преимущество автономного бега. [63] [64] Более поздняя система могла ходить по двумерной поверхности, а не по линейному пути, и продемонстрировала способность выборочно подбирать и перемещать молекулярный груз. [65] В 2018 году было показано, что цепная ДНК, использующая транскрипцию по катящемуся кругу с помощью прикрепленной РНК-полимеразы Т7 , движется по пути ДНК, направляемый сгенерированной цепью РНК. [66] Кроме того, был продемонстрирован линейный ходунок, который выполняет синтез по шаблону ДНК по мере продвижения ходунка по рельсам, что обеспечивает автономный многоэтапный химический синтез под руководством ходунка. [67] Функция синтетических ДНК-ходоков аналогична функции белков динеина и кинезина. [68]
смещения прядей Каскады
Каскады реакций смещения цепей можно использовать как для вычислительных, так и для структурных целей. Реакция смещения отдельной цепи включает выявление новой последовательности в ответ на присутствие некоторой цепи-инициатора. Многие такие реакции могут быть связаны в каскад , где вновь обнаруженная выходная последовательность одной реакции может инициировать другую реакцию замещения цепи в другом месте. Это, в свою очередь, позволяет создавать сети химических реакций со многими компонентами, демонстрируя сложные вычислительные возможности и возможности обработки информации. Эти каскады становятся энергетически выгодными за счет образования новых пар оснований и увеличения энтропии в результате реакций разборки. Каскады смещения цепей позволяют выполнять изотермическую операцию сборки или вычислительный процесс, в отличие от традиционного требования сборки нуклеиновой кислоты к этапу термического отжига, на котором температура повышается, а затем медленно снижается, чтобы обеспечить правильное формирование желаемой структуры. Они также могут поддержать каталитическая функция инициатора, при которой менее одного эквивалента инициатора может привести к завершению реакции. [23] [69]
Комплексы смещения нитей можно использовать для создания молекулярных логических элементов, способных выполнять сложные вычисления. [70] В отличие от традиционных электронных компьютеров, которые используют электрический ток в качестве входных и выходных данных, молекулярные компьютеры используют концентрации определенных химических веществ в качестве сигналов. В случае схем смещения цепи нуклеиновой кислоты сигналом является наличие цепей нуклеиновой кислоты, которые высвобождаются или потребляются в результате событий связывания и разъединения с другими цепями в комплексах замещения. Этот подход использовался для создания логических элементов, таких как логические элементы И, ИЛИ и НЕ. [71] Совсем недавно была продемонстрирована четырехбитная схема, которая может вычислять квадратный корень из целых чисел 0–15, используя систему вентилей, содержащую 130 нитей ДНК. [72]
Другое применение каскадов смещения нитей — создание динамически собираемых структур. В них используется шпилечная структура реагентов, так что при связывании входной цепи вновь обнаруженная последовательность оказывается в той же молекуле, а не разбирается. Это позволяет добавлять в растущий комплекс новые открытые шпильки. Этот подход использовался для создания простых структур, таких как трех- и четырехветвевые соединения и дендримеры . [69]
Приложения [ править ]
Нанотехнология ДНК обеспечивает один из немногих способов формирования сложных структур с точным контролем над наноразмерными характеристиками. Эта область начинает находить применение для решения фундаментальных научных задач в области структурной биологии и биофизики . Самое раннее такое применение, предусмотренное в этой области, и еще находящееся в разработке, относится к кристаллографии , где молекулы, которые трудно кристаллизовать изолированно, можно расположить внутри трехмерной решетки нуклеиновой кислоты, что позволяет определить их структуру. Другое применение - использование стержней ДНК-оригами для замены жидких кристаллов в экспериментах по остаточному диполярному соединению в ЯМР-спектроскопии белков ; использование ДНК-оригами выгодно, поскольку, в отличие от жидких кристаллов, они устойчивы к детергентам, необходимым для суспендирования мембранных белков в растворе. ДНК-ходоходы использовались в качестве наноразмерных сборочных линий для перемещения наночастиц и прямого химического синтеза . Кроме того, структуры ДНК-оригами помогли в биофизических исследованиях Функция фермента и сворачивание белка . [24] [8]
Нанотехнологии ДНК приближаются к потенциальным практическим приложениям. Способность массивов нуклеиновых кислот упорядочивать другие молекулы указывает на их потенциальное применение в электронике молекулярного масштаба. Сборка структуры нуклеиновой кислоты может быть использована для создания шаблона сборки молекулярных электронных элементов, таких как молекулярные провода , обеспечивая метод нанометрового контроля размещения и общей архитектуры устройства, аналогичного молекулярному макету . [24] [6] Нанотехнологию ДНК сравнивают с концепцией программируемой материи из-за связи вычислений со свойствами ее материала. [73]
В ходе исследования, проведенного группой ученых из центров iNANO и CDNA Орхусского университета , исследователям удалось сконструировать небольшую мультипереключаемую трехмерную коробку ДНК-оригами. Предложенная наночастица была охарактеризована методами атомно-силовой микроскопии (АСМ), просвечивающей электронной микроскопии (ПЭМ) и резонансного переноса энергии Фёрстера (FRET). Было показано, что построенный ящик имеет уникальный механизм повторного закрытия, который позволяет ему неоднократно открываться и закрываться в ответ на уникальный набор ключей ДНК или РНК. Авторы предположили, что это «устройство ДНК потенциально может быть использовано для широкого спектра применений, таких как контроль функций отдельных молекул, контролируемая доставка лекарств и молекулярные вычисления». [74]
Существуют потенциальные применения ДНК-нанотехнологии в наномедицине, позволяющей использовать ее способность выполнять вычисления в биосовместимом формате для создания «умных лекарств» для адресной доставки лекарств , а также для диагностических приложений. Одна из таких исследуемых систем использует полый ящик ДНК, содержащий белки, вызывающие апоптоз или гибель клеток, который открывается только в непосредственной близости от раковой клетки . [8] [75] Кроме того, существует интерес к экспрессии этих искусственных структур в сконструированных живых бактериальных клетках, скорее всего, с использованием транскрибируемой РНК для сборки, хотя неизвестно, способны ли эти сложные структуры эффективно сворачиваться или собираться в цитоплазме клетки . В случае успеха это могло бы обеспечить направленную эволюцию наноструктур нуклеиновых кислот. [6] Ученые из Оксфордского университета сообщили о самосборке четырех коротких нитей синтетической ДНК в клетку, которая может проникать в клетки и сохраняться в течение как минимум 48 часов. Было обнаружено, что флуоресцентно-меченные тетраэдры ДНК остаются интактными в лабораторных культивируемых клетках почек человека, несмотря на атаку клеточных ферментов через два дня. Этот эксперимент показал возможность доставки лекарств внутрь живых клеток с использованием «клетки» ДНК. [76] [77] ДНК- тетраэдр использовался для доставки РНК-интерференции (РНКи) на мышиной модели, сообщила группа исследователей из Массачусетского технологического института . Доставка интерферирующей РНК для лечения показала некоторый успех с использованием полимера или липида , но существуют ограничения безопасности и неточное нацеливание, а также короткий срок хранения в кровотоке. Наноструктура ДНК, созданная командой, состоит из шести нитей ДНК, образующих тетраэдр, к каждому из шести краев которого прикреплена одна нить РНК. Тетраэдр дополнительно оснащен нацеливающим белком, тремя молекулами фолата , которые приводят наночастицы ДНК к обильным рецепторам фолата, обнаруженным в некоторых опухолях. Результат показал, что экспрессия гена, на который воздействует РНКи, люцифераза , снизилась более чем вдвое. Это исследование демонстрирует перспективность использования нанотехнологии ДНК в качестве эффективного инструмента для лечения с использованием новой технологии РНК-интерференции. [78] [79] Тетраэдр ДНК также использовался в попытке преодолеть феномен множественной лекарственной устойчивости . Доксорубицин (DOX) конъюгировали с тетраэдром и загружали в клетки рака молочной железы MCF-7, которые содержали насос оттока лекарственного средства P-гликопротеина . Результаты эксперимента показали, что DOX не откачивался и был достигнут апоптоз раковых клеток. Тетраэдр без DOX был загружен в клетки для проверки его биосовместимости, и сама структура не проявила цитотоксичности. [80] Тетраэдр ДНК также использовался в качестве штрих-кода для профилирования субклеточной экспрессии и распределения белков в клетках в диагностических целях. Тетраэдрическая наноструктура продемонстрировала усиленный сигнал благодаря более высокой эффективности и стабильности мечения. [81]
Применение нанотехнологий ДНК в наномедицине также направлено на имитацию структуры и функций природных мембранных белков с помощью разработанных наноструктур ДНК. В 2012 году Лангекер и др. [82] представили структуру ДНК-оригами в форме пор, которая может самостоятельно внедряться в липидные мембраны посредством гидрофобных модификаций холестерина и индуцировать ионные токи через мембрану. За этой первой демонстрацией синтетического ионного канала ДНК последовало множество конструкций, индуцирующих поры, начиная от одного дуплекса ДНК и заканчивая созданием пор . [83] к небольшим конструкциям на основе плитки, [84] [85] [86] [87] [88] и большие трансмембранные порины ДНК-оригами . [89] Подобно естественным белковым ионным каналам , этот ансамбль синтетических аналогов, созданных из ДНК, таким образом, охватывает несколько порядков проводимости. Исследование встраивающего в мембрану одиночного дуплекса ДНК показало, что ток также должен течь по границе раздела ДНК-липид, поскольку в конструкции отсутствует просвет центрального канала, который позволяет ионам проходить через липидный бислой . Это указывало на то, что липидная пора, индуцированная ДНК, имеет тороидальную форму, а не цилиндрическую, поскольку головные группы липидов переориентируются в сторону встроенной в мембрану части ДНК. [83] Затем исследователи из Кембриджского университета и Университета Иллинойса в Урбана-Шампейн продемонстрировали, что такая тороидальная пора, индуцированная ДНК, может способствовать быстрому переворачиванию липидов между листочками липидного бислоя. построенный на ДНК Используя этот эффект, они разработали синтетический фермент, , который переворачивает липиды в биологических мембранах на несколько порядков быстрее, чем встречающиеся в природе белки, называемые скрамблазами . [90] Эта разработка подчеркивает потенциал синтетических наноструктур ДНК для персонализированных лекарств и терапевтических средств.
Дизайн [ править ]
Наноструктуры ДНК должны быть рационально спроектированы так, чтобы отдельные цепи нуклеиновых кислот могли собираться в нужные структуры. Этот процесс обычно начинается с определения желаемой целевой структуры или функции. Затем определяется общая вторичная структура целевого комплекса с указанием расположения цепей нуклеиновой кислоты внутри структуры и того, какие части этих цепей должны быть связаны друг с другом. Последним шагом является проектирование первичной структуры , которое представляет собой спецификацию реальных последовательностей оснований каждой цепи нуклеиновой кислоты. [36] [91]
Структурный проект [ править ]
Первым шагом в разработке наноструктуры нуклеиновой кислоты является решение о том, как данная структура должна быть представлена определенным расположением нитей нуклеиновой кислоты. На этом этапе проектирования определяется вторичная структура или положение пар оснований, которые удерживают отдельные нити вместе в желаемой форме. [36] Было продемонстрировано несколько подходов:
- Конструкции на основе плитки. Этот подход разбивает целевую структуру на более мелкие единицы с сильной связью между нитями, содержащимися в каждой единице, и более слабыми взаимодействиями между единицами. Его часто используют для создания периодических решеток, но также можно использовать для реализации алгоритмической самосборки, что делает их платформой для вычислений на ДНК . Это была доминирующая стратегия дизайна, использовавшаяся с середины 1990-х до середины 2000-х годов, когда была разработана методология ДНК-оригами. [36] [92]
- Складные конструкции. В качестве альтернативы подходу, основанному на плитке, подходы со складыванием создают наноструктуру из одной длинной нити, которая может либо иметь заданную последовательность, которая складывается из-за взаимодействия с самой собой, либо ее можно сложить в желаемую форму, используя более короткие, «штапельные» нити. "пряди. Этот последний метод называется ДНК-оригами , который позволяет формировать наноразмерные двух- и трехмерные формы (см. Дискретные структуры выше). [6] [13]
- Динамическая сборка. Этот подход напрямую контролирует кинетику самосборки ДНК, определяя все промежуточные этапы механизма реакции помимо конечного продукта. Это делается с использованием исходных материалов, имеющих структуру шпильки ; затем они собираются в окончательную конформацию в каскадной реакции в определенном порядке (см. Каскады смещения цепей ниже). Преимущество этого подхода состоит в том, что он протекает изотермически при постоянной температуре. Это контрастирует с термодинамическими подходами, которые требуют стадии термического отжига , где изменение температуры требуется для запуска сборки и содействия правильному формированию желаемой структуры. [6] [69]
Дизайн последовательности [ править ]
После того, как любой из вышеперечисленных подходов будет использован для создания вторичной структуры целевого комплекса, необходимо разработать реальную последовательность нуклеотидов, которая сформируется в желаемую структуру. Дизайн нуклеиновой кислоты — это процесс присвоения конкретной последовательности оснований нуклеиновой кислоты каждой из составляющих цепей структуры так, чтобы они ассоциировались в желаемой конформации. Целью большинства методов является разработка последовательностей так, чтобы целевая структура имела наименьшую энергию и, следовательно, была наиболее термодинамически выгодной, в то время как неправильно собранные структуры имели более высокие энергии и, следовательно, оказывались нежелательными. Это делается либо с помощью простых и быстрых эвристических методов, таких как минимизация симметрии последовательности , либо с использованием полной термодинамической модели ближайших соседей , которая более точна, но медленнее и требует больше вычислительных затрат. Геометрические модели используются для изучения третичной структуры наноструктур и обеспечения того, чтобы комплексы не были чрезмерно напряжены . [91] [93]
Дизайн нуклеиновых кислот преследует те же цели, что и дизайн белков . В обоих случаях последовательность мономеров разработана так, чтобы отдавать предпочтение желаемой целевой структуре и препятствовать другим структурам. Преимущество дизайна нуклеиновых кислот заключается в том, что он намного проще в вычислительном отношении, чем дизайн белка, поскольку простых правил спаривания оснований достаточно, чтобы предсказать энергетическую выгодность структуры, и подробная информация об общем трехмерном сворачивании структуры не требуется. Это позволяет использовать простые эвристические методы, которые позволяют получить экспериментально надежные конструкции. Структуры нуклеиновых кислот менее универсальны, чем белки, в своих функциях из-за повышенной способности белков сворачиваться в сложные структуры и ограниченного химического разнообразия четырех нуклеотидов по сравнению с двадцатью протеиногенными аминокислотами . [93]
Материалы и методы [ править ]

Последовательности цепей ДНК, составляющие целевую структуру, разрабатываются вычислительным путем с использованием программного обеспечения для молекулярного и термодинамического моделирования . [91] [93] Сами нуклеиновые кислоты затем синтезируются с использованием стандартных методов синтеза олигонуклеотидов , обычно автоматизированных в синтезаторе олигонуклеотидов , а цепи заказных последовательностей коммерчески доступны. [94] нити можно очистить с помощью денатурирующего гель-электрофореза . При необходимости [95] и точные концентрации, определяемые с помощью любого из нескольких количественного определения нуклеиновых кислот методов с использованием спектроскопии ультрафиолетового поглощения . [96]
Полностью сформировавшиеся целевые структуры можно проверить с помощью нативного гель-электрофореза, который дает информацию о размере и форме комплексов нуклеиновых кислот. Анализ электрофоретического изменения подвижности позволяет оценить, включает ли структура все желаемые цепи. [97] Флуоресцентная маркировка и резонансный перенос энергии Фёрстера (FRET) иногда используются для характеристики структуры комплексов. [98]
Структуры нуклеиновых кислот можно непосредственно визуализировать с помощью атомно-силовой микроскопии , которая хорошо подходит для расширенных двумерных структур, но менее полезна для дискретных трехмерных структур из-за взаимодействия кончика микроскопа с хрупкой структурой нуклеиновой кислоты; просвечивающую электронную микроскопию и криоэлектронную микроскопию В этом случае часто используют . Расширенные трехмерные решетки анализируются методом рентгеновской кристаллографии . [99] [100]
См. также [ править ]
- Международное общество наномасштабной науки, вычислений и инженерии
- Сравнение программного обеспечения для моделирования нуклеиновых кислот
- Молекулярные модели ДНК
- Нанобиотехнологии
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б Полиэдры ДНК: Гудман Р.П., Шаап И.А., Тардин К.Ф., Эрбен К.М., Берри Р.М., Шмидт К.Ф., Терберфилд А.Дж. (декабрь 2005 г.). «Быстрая хиральная сборка жестких строительных блоков ДНК для молекулярного нанопроизводства». Наука . 310 (5754): 1661–1665. Бибкод : 2005Sci...310.1661G . дои : 10.1126/science.1120367 . ПМИД 16339440 . S2CID 13678773 .
- ^ Jump up to: Перейти обратно: а б с История: Пелеско Ю.А. (2007). Самосборка: наука о вещах, которые соединяются воедино . Нью-Йорк: Чепмен и Холл/CRC. стр. 201, 242, 259. ISBN. 978-1-58488-687-7 .
- ^ Jump up to: Перейти обратно: а б с д и Обзор: Симан, Северная Каролина (июнь 2004 г.). «Нанотехнологии и двойная спираль». Научный американец . 290 (6): 64–75. Бибкод : 2004SciAm.290f..64S . doi : 10.1038/scientificamerican0604-64 . ПМИД 15195395 .
- ^ История: См. «Текущий протокол кристаллизации» . Лаборатория Надриана Зеемана. для постановки задачи и «Клетки ДНК, содержащие ориентированных гостей» . Лаборатория Надриана Зеемана. за предлагаемое решение.
- ^ Jump up to: Перейти обратно: а б с д и Обзор: Симан, Северная Каролина (2010). «Наноматериалы на основе ДНК» . Ежегодный обзор биохимии . 79 : 65–87. doi : 10.1146/annurev-biochem-060308-102244 . ПМЦ 3454582 . ПМИД 20222824 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я Обзор: Пинейру А.В., Хан Д., Ши В.М., Ян Х. (ноябрь 2011 г.). «Проблемы и возможности структурной ДНК-нанотехнологии» . Природные нанотехнологии . 6 (12): 763–772. Бибкод : 2011НатНа...6..763П . дои : 10.1038/nnano.2011.187 . ПМЦ 3334823 . ПМИД 22056726 .
- ^ Jump up to: Перейти обратно: а б ДНК-оригами: Ротемунд П.В. (2006). «Оригами из каркасной ДНК: от обобщенных мультикроссоверов к полигональным сетям». В Чен Дж., Йоноска Н., Розенберг Г. (ред.). Нанотехнологии: наука и вычисления . Серия естественных вычислений. Нью-Йорк: Спрингер. стр. 3–21. CiteSeerX 10.1.1.144.1380 . дои : 10.1007/3-540-30296-4_1 . ISBN 978-3-540-30295-7 .
- ^ Jump up to: Перейти обратно: а б с д и История/приложения: Служба РФ (июнь 2011 г.). «ДНК-нанотехнология. ДНК-нанотехнология растет». Наука . 332 (6034): 1140–1, 1143. Бибкод : 2011Sci...332.1140S . дои : 10.1126/science.332.6034.1140 . ПМИД 21636754 .
- ^ Юрке, Бернард; Терберфилд, Эндрю Дж.; Миллс, Аллен П.; Зиммель, Фридрих К.; Нойманн, Дженнифер Л. (август 2000 г.). «Молекулярная машина, работающая на ДНК и сделанная из ДНК» . Природа . 406 (6796): 605–608. Бибкод : 2000Natur.406..605Y . дои : 10.1038/35020524 . ISSN 1476-4687 . S2CID 2064216 .
- ^ Jump up to: Перейти обратно: а б с д ДНК-машины: Бат Дж., Терберфилд Эй.Дж. (май 2007 г.). «ДНК-наномашины». Природные нанотехнологии . 2 (5): 275–284. Бибкод : 2007NatNa...2..275B . дои : 10.1038/nnano.2007.104 . ПМИД 18654284 .
- ^ Наноархитектура: Робинсон Б.Х., Симан, Северная Каролина (август 1987 г.). «Дизайн биочипа: самособирающееся устройство памяти молекулярного масштаба». Белковая инженерия . 1 (4): 295–300. дои : 10.1093/белок/1.4.295 . ПМИД 3508280 .
- ^ Наноархитектура: Сяо С., Лю Ф., Розен А.Е., Хайнфельд Дж.Ф., Симан Н.К., Мюзье-Форсайт К., Киль Р.А. (август 2002 г.). «Самосборка массивов металлических наночастиц с помощью каркасов ДНК». Журнал исследований наночастиц . 4 (4): 313–317. Бибкод : 2002JNR.....4..313X . дои : 10.1023/А:1021145208328 . S2CID 2257083 .
- ^ Jump up to: Перейти обратно: а б с ДНК-оригами: Ротемунд П.В. (март 2006 г.). «Складывание ДНК для создания наноразмерных форм и узоров» (PDF) . Природа . 440 (7082): 297–302. Бибкод : 2006Natur.440..297R . дои : 10.1038/nature04586 . ПМИД 16541064 . S2CID 4316391 .
- ^ Jump up to: Перейти обратно: а б ДНК-оригами: Дуглас С.М., Дитц Х., Лидл Т., Хёгберг Б., Граф Ф., Ши В.М. (май 2009 г.). «Самосборка ДНК в наноразмерные трехмерные формы» . Природа . 459 (7245): 414–418. Бибкод : 2009Natur.459..414D . дои : 10.1038/nature08016 . ПМЦ 2688462 . ПМИД 19458720 .
- ^ Служба РФ (июнь 2011 г.). «ДНК-нанотехнология. ДНК-нанотехнология растет». Наука . 332 (6034): 1140–1, 1143. Бибкод : 2011Sci...332.1140S . дои : 10.1126/science.332.6034.1140 . ПМИД 21636754 .
- ^ История: Хопкин К. (август 2011 г.). «Профиль: 3-D провидец» . Ученый . Архивировано из оригинала 10 октября 2011 года . Проверено 8 августа 2011 г.
- ^ Jump up to: Перейти обратно: а б с История: Симан, Северная Каролина (июнь 2010 г.). «Структурная нанотехнология ДНК: растет вместе с Nano Letters» . Нано-буквы . 10 (6): 1971–1978. Бибкод : 2010NanoL..10.1971S . дои : 10.1021/nl101262u . ПМК 2901229 . ПМИД 20486672 .
- ^ Jump up to: Перейти обратно: а б с Обзор: Мао С. (декабрь 2004 г.). «Возникновение сложности: уроки ДНК» . ПЛОС Биология . 2 (12): е431. дои : 10.1371/journal.pbio.0020431 . ПМЦ 535573 . ПМИД 15597116 .
- ^ Предыстория: Пелеско Ю.А. (2007). Самосборка: наука о вещах, которые соединяются воедино . Нью-Йорк: Чепмен и Холл/CRC. стр. 5, 7. ISBN 978-1-58488-687-7 .
- ^ Предыстория: Длинный ЕС (1996). «Основы нуклеиновых кислот». В Хехте С.М. (ред.). Биоорганическая химия: нуклеиновые кислоты . Нью-Йорк: Издательство Оксфордского университета. стр. 4–10. ISBN 978-0-19-508467-2 .
- ^ Нанотехнология РНК: Хворос А., Северчан И., Койфман А.Ю., Вайнкам П., Оруджев Э., Хансма Х.Г., Джагер Л. (декабрь 2004 г.). «Создание программируемых пазлов с помощью РНК». Наука . 306 (5704): 2068–2072. Бибкод : 2004Sci...306.2068C . дои : 10.1126/science.1104686 . ПМИД 15604402 . S2CID 9296608 .
- ^ Нанотехнология РНК: Го П (декабрь 2010 г.). «Новая область нанотехнологий РНК» . Природные нанотехнологии . 5 (12): 833–842. Бибкод : 2010НатНа...5..833Г . дои : 10.1038/nnano.2010.231 . ПМК 3149862 . ПМИД 21102465 .
- ^ Jump up to: Перейти обратно: а б с д Динамическая ДНК-нанотехнология: Чжан Д.Ю., Силиг Г. (февраль 2011 г.). «Динамическая нанотехнология ДНК с использованием реакций смещения цепи». Природная химия . 3 (2): 103–113. Бибкод : 2011НатЧ...3..103Z . дои : 10.1038/nchem.957 . ПМИД 21258382 .
- ^ Jump up to: Перейти обратно: а б с д и Структурная ДНК-нанотехнология: Симан, Северная Каролина (ноябрь 2007 г.). «Обзор структурной ДНК-нанотехнологии» . Молекулярная биотехнология . 37 (3): 246–257. дои : 10.1007/s12033-007-0059-4 . ПМЦ 3479651 . ПМИД 17952671 .
- ^ Динамическая нанотехнология ДНК: Лу Ю, Лю Дж (декабрь 2006 г.). «Функциональная нанотехнология ДНК: новые применения ДНКзимов и аптамеров». Современное мнение в области биотехнологии . 17 (6): 580–588. дои : 10.1016/j.copbio.2006.10.004 . ПМИД 17056247 .
- ^ Моделирование структур ДНК: Дой Дж.П., Оулдридж Т.Э., Луис А.А., Романо Ф., Шулк П., Матек С. и др. (декабрь 2013 г.). «Крупнозернистая ДНК для моделирования нанотехнологий ДНК». Физическая химия Химическая физика . 15 (47): 20395–20414. arXiv : 1308.3843 . Бибкод : 2013PCCP...1520395D . дои : 10.1039/C3CP53545B . ПМИД 24121860 . S2CID 15324396 .
- ^ Другие массивы: Стронг М (март 2004 г.). «Белковые наномашины» . ПЛОС Биология . 2 (3): Е73. дои : 10.1371/journal.pbio.0020073 . ПМК 368168 . ПМИД 15024422 .
- ^ Ян Х., Пак С.Х., Финкельштейн Г., Рейф Дж.Х., ЛаБин Т.Х. (сентябрь 2003 г.). «Самосборка белковых массивов и высокопроводящих нанопроводов по шаблону ДНК». Наука . 301 (5641): 1882–1884. Бибкод : 2003Sci...301.1882Y . дои : 10.1126/science.1089389 . ПМИД 14512621 . S2CID 137635908 .
- ^ Jump up to: Перейти обратно: а б Алгоритмическая самосборка: Ротемунд П.В., Пападакис Н., Уинфри Э. (декабрь 2004 г.). «Алгоритмическая самосборка треугольников ДНК Серпинского» . ПЛОС Биология . 2 (12): е424. doi : 10.1371/journal.pbio.0020424 . ПМК 534809 . ПМИД 15583715 .
- ^ DX-массивы: Уинфри Э., Лю Ф., Венцлер Л.А., Симан Н.К. (август 1998 г.). «Дизайн и самосборка двумерных кристаллов ДНК». Природа . 394 (6693): 539–544. Бибкод : 1998Natur.394..539W . дои : 10.1038/28998 . ПМИД 9707114 . S2CID 4385579 .
- ^ DX-массивы: Лю Ф., Ша Р., Симан, Северная Каролина (10 февраля 1999 г.). «Модификация свойств поверхности двумерных кристаллов ДНК». Журнал Американского химического общества . 121 (5): 917–922. дои : 10.1021/ja982824a .
- ^ Другие массивы: Мао С., Сунь В., Симан, Северная Каролина (16 июня 1999 г.). «Спроектированные двумерные массивы соединений ДНК Холлидея, визуализированные с помощью атомно-силовой микроскопии». Журнал Американского химического общества . 121 (23): 5437–5443. дои : 10.1021/ja9900398 .
- ^ Другие массивы: Константину П.Е., Ван Т., Копач Дж., Исраэль Л.Б., Чжан Х., Дин Б. и др. (сентябрь 2006 г.). «Двойная когезия в структурных нанотехнологиях ДНК» . Органическая и биомолекулярная химия . 4 (18): 3414–3419. дои : 10.1039/b605212f . ПМК 3491902 . ПМИД 17036134 .
- ^ Другие массивы: Матье Ф., Ляо С., Копач Дж., Ван Т., Мао С., Симан Н.К. (апрель 2005 г.). «Шестиспиральные пучки, созданные из ДНК» . Нано-буквы . 5 (4): 661–665. Бибкод : 2005NanoL...5..661M . дои : 10.1021/nl050084f . ПМЦ 3464188 . ПМИД 15826105 .
- ^ Алгоритмическая самосборка: Бариш Р.Д., Ротемунд П.В., Уинфри Э. (декабрь 2005 г.). «Два вычислительных примитива для алгоритмической самосборки: копирование и счет». Нано-буквы . 5 (12): 2586–2592. Бибкод : 2005NanoL...5.2586B . CiteSeerX 10.1.1.155.676 . дои : 10.1021/nl052038l . ПМИД 16351220 .
- ^ Jump up to: Перейти обратно: а б с д Дизайн: Фельдкамп У, Нимейер К.М. (март 2006 г.). «Рациональный дизайн наноархитектур ДНК». Ангеванде Хеми . 45 (12): 1856–1876. дои : 10.1002/anie.200502358 . ПМИД 16470892 .
- ^ ДНК-нанотрубки: Ротемунд П.В., Экани-Нкодо А., Пападакис Н., Кумар А., Файгенсон Д.К., Уинфри Э. (декабрь 2004 г.). «Дизайн и характеристика программируемых ДНК-нанотрубок» . Журнал Американского химического общества . 126 (50): 16344–16352. дои : 10.1021/ja044319l . ПМИД 15600335 .
- ^ ДНК-нанотрубки: Инь П., Хариади Р.Ф., Саху С., Чой Х.М., Пак Ш., Лабин Т.Х., Рейф Дж.Х. (август 2008 г.). «Программирование окружностей пробирок ДНК» . Наука . 321 (5890): 824–826. Бибкод : 2008Sci...321..824Y . дои : 10.1126/science.1157312 . ПМИД 18687961 . S2CID 12100380 .
- ^ Трехмерные массивы: Чжэн Дж., Бирктофт Дж.Дж., Чен Ю., Ван Т., Ша Р., Константину П.Е. и др. (сентябрь 2009 г.). «От молекулярного к макроскопическому через рациональный дизайн самособирающегося трехмерного кристалла ДНК» . Природа . 461 (7260): 74–77. Бибкод : 2009Natur.461...74Z . дои : 10.1038/nature08274 . ПМК 2764300 . ПМИД 19727196 .
- ^ Полиэдры ДНК: Чжан Ю, Симан, Северная Каролина (1 марта 1994 г.). «Построение усеченного ДНК октаэдра». Журнал Американского химического общества . 116 (5): 1661–1669. дои : 10.1021/ja00084a006 .
- ^ Полиэдры ДНК: Ши В.М., Киспе Дж.Д., Джойс Г.Ф. (февраль 2004 г.). «Одноцепочечная ДНК длиной 1,7 тыс. оснований, которая сворачивается в наноразмерный октаэдр». Природа . 427 (6975): 618–621. Бибкод : 2004Natur.427..618S . дои : 10.1038/nature02307 . ПМИД 14961116 . S2CID 4419579 .
- ^ Тихомиров Г., Петерсен П., Цянь Л. (декабрь 2017 г.). «Фрактальная сборка массивов ДНК-оригами микрометрового масштаба с произвольными узорами» . Природа . 552 (7683): 67–71. Бибкод : 2017Natur.552...67T . дои : 10.1038/nature24655 . ПМИД 29219965 . S2CID 4455780 .
- ^ Jump up to: Перейти обратно: а б Коробки ДНК: Андерсен Э.С., Донг М., Нильсен М.М., Ян К., Субрамани Р., Мамду В. и др. (май 2009 г.). «Самостоятельная сборка наноразмерного ящика ДНК с управляемой крышкой». Природа . 459 (7243): 73–76. Бибкод : 2009Natur.459...73A . дои : 10.1038/nature07971 . hdl : 11858/00-001M-0000-0010-9363-9 . ПМИД 19424153 . S2CID 4430815 .
- ^ Коробки ДНК: Ке Ю, Шарма Дж, Лю М, Ян К, Лю Ю, Ян Х (июнь 2009 г.). «Каркас ДНК-оригами из молекулярного контейнера тетраэдра ДНК». Нано-буквы . 9 (6): 2445–2447. Бибкод : 2009NanoL...9.2445K . дои : 10.1021/nl901165f . ПМИД 19419184 .
- ^ Заборова О.В.; Войнова, А.Д.; Шмыков, Б.Д.; Сергеев, В.Г. (2021). «Твердые липидные наночастицы для инкапсуляции нуклеиновых кислот» . Обзоры и достижения в области химии . 11 (3–4): 178–188. дои : 10.1134/S2079978021030055 . ISSN 2634-8276 . S2CID 246946068 .
- ^ Обзор: Эндо М., Сугияма Х. (октябрь 2009 г.). «Химические подходы к нанотехнологии ДНК». ХимБиоХим . 10 (15): 2420–2443. дои : 10.1002/cbic.200900286 . ПМИД 19714700 . S2CID 205554125 .
- ^ Наноархитектура: Чжэн Дж., Константину П.Е., Майкл С., Аливисатос А.П., Киль Р.А., Симан Н.К. (июль 2006 г.). «Двумерные массивы наночастиц демонстрируют организационную силу надежных мотивов ДНК» . Нано-буквы . 6 (7): 1502–1504. Бибкод : 2006NanoL...6.1502Z . дои : 10.1021/nl060994c . ПМЦ 3465979 . ПМИД 16834438 .
- ^ Наноархитектура: Парк С.Х., Пистол С., Ан С.Дж., Рейф Дж.Х., Лебек А.Р., Дуайер С., ЛаБин Т.Д. (январь 2006 г.). «Полностью адресуемые решетки ДНК конечного размера, сформированные с помощью иерархических процедур сборки» . Ангеванде Хеми . 45 (5): 735–739. Бибкод : 2006АнгЧ.118.6759П . дои : 10.1002/ange.200690141 . ПМИД 16374784 .
- ^ Наноархитектура: Коэн Дж.Д., Садовски Дж.П., Дерван П.Б. (22 октября 2007 г.). «Адресация одиночных молекул на наноструктурах ДНК» . Ангеванде Хеми . 46 (42): 7956–7959. дои : 10.1002/anie.200702767 . ПМИД 17763481 .
- ^ Наноархитектура: Мауне Х.Т., Хан С.П., Бариш Р.Д., Бократ М., Годдард В.А., Ротемунд П.В., Уинфри Э. (январь 2010 г.). «Самосборка углеродных нанотрубок в двумерную геометрию с использованием шаблонов ДНК-оригами» . Природные нанотехнологии . 5 (1): 61–66. Бибкод : 2010НатНа...5...61М . дои : 10.1038/nnano.2009.311 . ПМИД 19898497 .
- ^ Наноархитектура: Лю Дж., Гэн Ю., Паунд Э., Гьявали С., Эштон Дж.Р., Хики Дж. и др. (март 2011 г.). «Металлизация разветвленной ДНК-оригами для изготовления наноэлектронных схем». АСУ Нано . 5 (3): 2240–2247. дои : 10.1021/nn1035075 . ПМИД 21323323 .
- ^ Наноархитектура: Дэн Цз, Мао С (август 2004 г.). «Молекулярная литография с наноструктурами ДНК». Ангеванде Хеми . 43 (31): 4068–4070. дои : 10.1002/anie.200460257 . ПМИД 15300697 .
- ^ ДНК-машины: Мао С., Сунь В., Шэнь З., Симан, Северная Каролина (январь 1999 г.). «Наномеханическое устройство на основе BZ-перехода ДНК». Природа . 397 (6715): 144–146. Бибкод : 1999Natur.397..144M . дои : 10.1038/16437 . ПМИД 9923675 . S2CID 4406177 .
- ^ ДНК-машины: Юрк Б., Терберфилд А.Дж., Миллс А.П., Зиммель ФК, Нойманн Дж.Л. (август 2000 г.). «Молекулярная машина, работающая на ДНК и сделанная из ДНК». Природа . 406 (6796): 605–608. Бибкод : 2000Natur.406..605Y . дои : 10.1038/35020524 . ПМИД 10949296 . S2CID 2064216 .
- ^ ДНК-машины: Ян Х, Чжан Х, Шэнь З, Симан, Северная Каролина (январь 2002 г.). «Надежное механическое устройство ДНК, управляемое топологией гибридизации». Природа . 415 (6867): 62–65. Бибкод : 2002Natur.415...62Y . дои : 10.1038/415062а . ПМИД 11780115 . S2CID 52801697 .
- ^ ДНК-машины: Фэн Л., Пак Ш.Х., Рейф Дж.Х., Ян Х. (сентябрь 2003 г.). «Решетка ДНК с двумя состояниями, переключаемая наноактюатором ДНК». Ангеванде Хеми . 42 (36): 4342–4346. Бибкод : 2003АнгЧ.115.4478F . дои : 10.1002/ange.200351818 . ПМИД 14502706 .
- ^ ДНК-машины: Гудман Р.П., Хайлеманн М., Дуз С., Эрбен К.М., Капанидис А.Н., Терберфилд А.Дж. (февраль 2008 г.). «Реконфигурируемые, скрепленные, трехмерные наноструктуры ДНК». Природные нанотехнологии . 3 (2): 93–96. Бибкод : 2008НатНа...3...93Г . дои : 10.1038/nnano.2008.3 . ПМИД 18654468 .
- ^ Приложения: Дуглас С.М., Бачелет I, генеральный директор Черча (февраль 2012 г.). «Наноробот с логическим управлением для целевой транспортировки молекулярных грузов». Наука . 335 (6070): 831–834. Бибкод : 2012Sci...335..831D . дои : 10.1126/science.1214081 . ПМИД 22344439 . S2CID 9866509 .
- ^ Чентола, Матиас; Попплтон, Эрик; Рэй, Суджей; Чентола, Мартин; Велти, Робб; Валеро, Хулиан; Вальтер, Нильс Г.; Шульц, Петр; Фамулок, Майкл (19 октября 2023 г.). «Ритмически пульсирующий нанодвигатель ДНК-оригами с листовой пружиной, приводящий в движение пассивного ведомого» . Природные нанотехнологии : 1–11. дои : 10.1038/s41565-023-01516-x . ISSN 1748-3395 . ПМЦ 10873200 . ПМИД 37857824 .
- ^ ДНК-ходоки: Шин Дж.С., Пирс Н.А. (сентябрь 2004 г.). «Синтетический ходок ДНК для молекулярного транспорта» . Журнал Американского химического общества . 126 (35): 10834–10835. дои : 10.1021/ja047543j . ПМИД 15339155 .
- ^ ДНК-ходоки: Шерман В.Б., Симан, Северная Каролина (июль 2004 г.). «Двуногое шагающее устройство с точным управлением ДНК». Нано-буквы . 4 (7): 1203–1207. Бибкод : 2004NanoL...4.1203S . дои : 10.1021/nl049527q .
- ^ ДНК-ходоки: Шкугор М., Валеро Дж., Мураяма К., Чентола М., Асанума Х., Фамулок М. (май 2019 г.). «Ортогонально фотоуправляемый неавтономный ДНК-ходок». Ангеванде Хеми . 58 (21): 6948–6951. дои : 10.1002/anie.201901272 . ПМИД 30897257 . S2CID 85446523 .
- ^ ДНК-ходоки: Тянь Ю, Хэ Ю, Чэнь Ю, Инь П, Мао С (июль 2005 г.). «ДНКзим, который движется процессивно и автономно по одномерному пути». Ангеванде Хеми . 44 (28): 4355–4358. Бибкод : 2005АнгЧ.117.4429Т . дои : 10.1002/ange.200500703 . ПМИД 15945114 .
- ^ ДНК-ходоки: Бат Дж., Грин С.Дж., Терберфилд Эй.Дж. (июль 2005 г.). «Свободно работающий ДНК-мотор, приводимый в действие разрывающим ферментом». Ангеванде Хеми . 44 (28): 4358–4361. дои : 10.1002/anie.200501262 . ПМИД 15959864 .
- ^ Функциональные ходоки ДНК: Лунд К., Манзо А.Дж., Дабби Н., Микелотти Н., Джонсон-Бак А., Нангрив Дж. и др. (май 2010 г.). «Молекулярные роботы, управляемые предписывающими ландшафтами» . Природа . 465 (7295): 206–210. Бибкод : 2010Natur.465..206L . дои : 10.1038/nature09012 . ПМК 2907518 . ПМИД 20463735 .
- ^ Функциональные ходоки ДНК: Валеро Дж., Пал Н., Дакал С., Уолтер Н.Г., Фамулок М. (июнь 2018 г.). «Биогибридный ДНК-роторно-статорный нанодвигатель, который движется по заданным траекториям» . Природные нанотехнологии . 13 (6): 496–503. Бибкод : 2018НатНа..13..496В . дои : 10.1038/s41565-018-0109-z . ПМК 5994166 . ПМИД 29632399 .
- ^ Функциональные ходоки ДНК: Хэ Ю, Лю Д. Р. (ноябрь 2010 г.). «Автономный многостадийный органический синтез в одном изотермическом растворе, опосредованный ходоком ДНК» . Природные нанотехнологии . 5 (11): 778–782. Бибкод : 2010NatNa...5..778H . дои : 10.1038/nnano.2010.190 . ПМК 2974042 . ПМИД 20935654 .
- ^ Пан Дж., Ли Ф., Ча Т.Г., Чен Х., Чой Дж.Х. (август 2015 г.). «Последний прогресс в создании ходунков на основе ДНК». Современное мнение в области биотехнологии . 34 : 56–64. дои : 10.1016/j.copbio.2014.11.017 . ПМИД 25498478 .
- ^ Jump up to: Перейти обратно: а б с Кинетическая сборка: Инь П., Чой Х.М., Калверт Ч.Р., Пирс Н.А. (январь 2008 г.). «Программирование путей биомолекулярной самосборки» . Природа . 451 (7176): 318–322. Бибкод : 2008Natur.451..318Y . дои : 10.1038/nature06451 . ПМИД 18202654 . S2CID 4354536 .
- ^ Нечеткие и логические логические элементы на основе ДНК: Задеган Р.М., Джепсен, доктор медицины, Хильдебрандт Л.Л., Биркедал В., Кьемс Дж. (апрель 2015 г.). «Построение нечетких и булевых логических элементов на основе ДНК». Маленький . 11 (15): 1811–1817. дои : 10.1002/smll.201402755 . ПМИД 25565140 .
- ^ Каскады смещения прядей: Силиг Г., Соловейчик Д., Чжан Д.Ю., Уинфри Э. (декабрь 2006 г.). «Безферментные логические схемы нуклеиновых кислот» . Наука . 314 (5805): 1585–1588. Бибкод : 2006Sci...314.1585S . дои : 10.1126/science.1132493 . ПМИД 17158324 . S2CID 10966324 .
- ^ Каскады смещения прядей: Цянь Л., Уинфри Э. (июнь 2011 г.). «Масштабирование вычислений цифровых схем с помощью каскадов смещения нитей ДНК». Наука . 332 (6034): 1196–1201. Бибкод : 2011Sci...332.1196Q . дои : 10.1126/science.1200520 . ПМИД 21636773 . S2CID 10053541 .
- ^ Приложения: Ритман Э.А. (2001). Молекулярная инженерия наносистем . Спрингер. стр. 209–212. ISBN 978-0-387-98988-4 . Проверено 17 апреля 2011 г.
- ^ Задеган Р.М., Джепсен М.Д., Томсен К.Е., Охольм А.Х., Шафферт Д.Х., Андерсен Э.С. и др. (ноябрь 2012 г.). «Создание переключаемого 3D-ящика ДНК-оригами емкостью 4 зептолитра». АСУ Нано . 6 (11): 10050–10053. дои : 10.1021/nn303767b . ПМИД 23030709 .
- ^ Приложения: Юнгманн Р., Реннер С., Зиммель (апрель 2008 г.). «От ДНК-нанотехнологии к синтетической биологии» . Журнал HFSP . 2 (2): 99–109. дои : 10.2976/1.2896331 . ПМЦ 2645571 . ПМИД 19404476 .
- ^ Лови, Ховард (5 июля 2011 г.). «Клетки ДНК могут высвобождать лекарства внутри клеток» . www.fightedrugdelivery.com . Проверено 22 сентября 2013 г. [ постоянная мертвая ссылка ]
- ^ Уолш А.С., Инь Х., Эрбен К.М., Вуд М.Дж., Терберфилд А.Дж. (июль 2011 г.). «Доставка клеток ДНК в клетки млекопитающих». АСУ Нано . 5 (7): 5427–5432. дои : 10.1021/nn2005574 . ПМИД 21696187 .
- ^ Трафтон, Энн (4 июня 2012 г.). «Исследователи достигли РНК-интерференции в более легкой упаковке» . Новости МТИ . Проверено 22 сентября 2013 г.
- ^ Ли Х., Литтон-Джин А.К., Чен Ю., Лав К.Т., Парк А.И., Карагианнис Э.Д. и др. (июнь 2012 г.). «Молекулярно самособирающиеся наночастицы нуклеиновой кислоты для адресной доставки миРНК in vivo» . Природные нанотехнологии . 7 (6): 389–393. Бибкод : 2012НатНа...7..389Л . дои : 10.1038/NNANO.2012.73 . ПМЦ 3898745 . ПМИД 22659608 .
- ^ Ким КР, Ким Д.Р., Ли Т., Йи Дж.И., Ким Б.С., Квон И.С., Ан Д.Р. (март 2013 г.). «Доставка лекарств с помощью самособирающегося тетраэдра ДНК для преодоления лекарственной устойчивости в клетках рака молочной железы». Химические коммуникации . 49 (20): 2010–2012. дои : 10.1039/c3cc38693g . ПМИД 23380739 .
- ^ Сунда Н.Р., Хо Н.Р., Лим Г.С., Наталья А., Дин Х., Лю Ю. и др. (сентябрь 2019 г.). «Наноструктуры ДНК со штрих-кодом для мультиплексного профилирования субклеточного распределения белков». Природная биомедицинская инженерия . 3 (9): 684–694. дои : 10.1038/s41551-019-0417-0 . ПМИД 31285580 . S2CID 195825879 .
- ^ Ионные каналы ДНК: Лангекер М., Арнаут В., Мартин Т.Г., Лист Дж., Реннер С., Майер М. и др. (ноябрь 2012 г.). «Синтетические липидные мембранные каналы, образованные сконструированными наноструктурами ДНК» . Наука . 338 (6109): 932–936. Бибкод : 2012Sci...338..932L . дои : 10.1126/science.1225624 . ПМЦ 3716461 . ПМИД 23161995 .
- ^ Jump up to: Перейти обратно: а б Ионные каналы ДНК: Гёпфрих К., Ли С.И., Мамес И., Бхамидимарри С.П., Риччи М., Ю Дж. и др. (июль 2016 г.). «Ионные каналы, созданные из одного перекрывающего мембрану дуплекса ДНК» . Нано-буквы . 16 (7): 4665–4669. Бибкод : 2016NanoL..16.4665G . дои : 10.1021/acs.nanolett.6b02039 . ПМЦ 4948918 . ПМИД 27324157 .
- ^ Ионные каналы ДНК: Бернс-младший, Стульц Э., Ховорка С. (июнь 2013 г.). «Самособирающиеся нанопоры ДНК, охватывающие липидные бислои». Нано-буквы . 13 (6): 2351–2356. Бибкод : 2013NanoL..13.2351B . CiteSeerX 10.1.1.659.7660 . дои : 10.1021/nl304147f . ПМИД 23611515 .
- ^ Ионные каналы ДНК: Бернс-младший, Гёпфрих К., Вуд Дж.В., Такер В.В., Стульц Э., Кейзер У.Ф., Ховорка С. (ноябрь 2013 г.). «Нанопоры ДНК, охватывающие липидный бислой, с бифункциональным порфириновым якорем» . Ангеванде Хеми . 52 (46): 12069–12072. дои : 10.1002/anie.201305765 . ПМК 4016739 . ПМИД 24014236 .
- ^ Ионные каналы ДНК: Зайферт А., Гёпфрих К., Бернс-младший, Фертиг Н., Кейзер У.Ф., Ховорка С. (февраль 2015 г.). «Двуслойные нанопоры ДНК с переключением напряжения между открытым и закрытым состоянием» . АСУ Нано . 9 (2): 1117–1126. дои : 10.1021/nn5039433 . ПМК 4508203 . ПМИД 25338165 .
- ^ Ионные каналы ДНК: Гёпфрих К., Зеттл Т., Мейеринг А.Е., Эрнандес-Аинса С., Кокабей С., Лидл Т., Кейзер У.Ф. (май 2015 г.). «Структуры ДНК-плитки индуцируют ионные токи через липидные мембраны» . Нано-буквы . 15 (5): 3134–3138. Бибкод : 2015NanoL..15.3134G . дои : 10.1021/acs.nanolett.5b00189 . ПМИД 25816075 .
- ^ Ионные каналы ДНК: Бернс-младший, Зайферт А., Фертиг Н., Ховорка С. (февраль 2016 г.). «Биомиметический канал на основе ДНК для лиганд-контролируемого транспорта заряженных молекулярных грузов через биологическую мембрану» . Природные нанотехнологии . 11 (2): 152–156. Бибкод : 2016НатНа..11..152Б . дои : 10.1038/nnano.2015.279 . ПМИД 26751170 .
- ^ Ионные каналы ДНК: Гёпфрих К., Ли С.И., Риччи М., Бхамидимарри С.П., Ю Дж., Гинес Б. и др. (сентябрь 2016 г.). «Трансмембранный порин с большой проводимостью, сделанный из ДНК-оригами» . АСУ Нано . 10 (9): 8207–8214. дои : 10.1021/acsnano.6b03759 . ПМК 5043419 . ПМИД 27504755 .
- ^ Скрамблаза ДНК: Оманн А., Ли С.И., Маффео С., Аль Нахас К., Бауманн К.Н., Гёпфрих К. и др. (июнь 2018 г.). «Синтетический фермент, построенный из ДНК, переворачивает 10 7 липидов в секунду в биологических мембранах» . Nature Communications . 9 (1): 2426. Bibcode : ...9.2426O . doi : 10.1038/s41467-018-04821-5 . PMC 6013447. 2018NatCo PMID 29930243 .
- ^ Jump up to: Перейти обратно: а б с Дизайн: Бреннеман А., Кондон А. (25 сентября 2002 г.). «Дизайн нитей для биомолекулярных вычислений» . Теоретическая информатика . 287 : 39–58. дои : 10.1016/S0304-3975(02)00135-4 .
- ^ Обзор: Линь С., Лю Ю., Ринкер С., Ян Х. (август 2006 г.). «Самосборка на основе плитки ДНК: построение сложных наноархитектур». ХимияФизХим . 7 (8): 1641–1647. дои : 10.1002/cphc.200600260 . ПМИД 16832805 .
- ^ Jump up to: Перейти обратно: а б с Дизайн: Диркс Р.М., Лин М., Уинфри Э., Пирс Н.А. (15 февраля 2004 г.). «Парадигмы вычислительного дизайна нуклеиновых кислот» . Исследования нуклеиновых кислот . 32 (4): 1392–1403. дои : 10.1093/nar/gkh291 . ПМК 390280 . ПМИД 14990744 .
- ^ Методы: Эллингтон А., Поллард Дж.Д. (1 мая 2001 г.). «Синтез и очистка олигонуклеотидов». Современные протоколы молекулярной биологии . 42 : 2.11.1–2.11.25. дои : 10.1002/0471142727.mb0211s42 . ISBN 978-0471142720 . ПМИД 18265179 . S2CID 205152989 .
- ^ Методы: Эллингтон А., Поллард Дж.Д. (1 мая 2001 г.). «Очистка олигонуклеотидов с использованием электрофореза в денатурирующем полиакриламидном геле». Современные протоколы молекулярной биологии . 42 : Блок 2.12. дои : 10.1002/0471142727.mb0212s42 . ISBN 978-0471142720 . ПМИД 18265180 . S2CID 27187583 .
- ^ Методы: Галлахер С.Р., Дежарден П. (1 июля 2011 г.). «Количественное определение нуклеиновых кислот и белков». Текущие протоколы Основные лабораторные методы . Том. 5. дои : 10.1002/9780470089941.et0202s5 . ISBN 978-0470089934 . S2CID 94329398 .
- ^ Методы: Чори Дж., Поллард Дж.Д. (1 мая 2001 г.). «Разделение небольших фрагментов ДНК с помощью обычного гель-электрофореза». Современные протоколы молекулярной биологии . 47 : Блок 2.7. дои : 10.1002/0471142727.mb0207s47 . ISBN 978-0471142720 . ПМИД 18265187 . S2CID 43406338 .
- ^ Методы: Вальтер Н.Г. (1 февраля 2003 г.). «Изучение структурной динамики и функции РНК с помощью резонансной передачи энергии флуоресценции (FRET)». Современные протоколы химии нуклеиновых кислот . 11 : 11.10.1–11.10.23. дои : 10.1002/0471142700.nc1110s11 . ISBN 978-0471142706 . ПМИД 18428904 . S2CID 9978415 .
- ^ Методы: Лин С, Ке Ю, Чабра Р, Шарма Дж, Лю Ю, Ян Х (2011). «Синтез и характеристика самособирающихся наноструктур ДНК». В Зуккери Г., Самори Б. (ред.). ДНК-нанотехнологии . Методы молекулярной биологии. Том. 749. стр. 1–11. дои : 10.1007/978-1-61779-142-0_1 . ISBN 978-1-61779-141-3 . ПМИД 21674361 .
- ^ Методы: Блумфилд В.А., Кротерс Д.М., Тиноко-младший I (2000). Нуклеиновые кислоты: структура, свойства и функции . Саусалито, Калифорния: Университетские научные книги. стр. 84–86, 396–407. ISBN 978-0-935702-49-1 .
Дальнейшее чтение [ править ]
Общий:
- Симан, Северная Каролина (июнь 2004 г.). «Нанотехнологии и двойная спираль». Научный американец . 290 (6): 64–75. Бибкод : 2004SciAm.290f..64S . doi : 10.1038/scientificamerican0604-64 . ПМИД 15195395 . — Статья, написанная основателем направления для непрофессионалов.
- Симан, Северная Каролина (июнь 2010 г.). «Структурная нанотехнология ДНК: растет вместе с Nano Letters» . Нано-буквы . 10 (6): 1971–1978. Бибкод : 2010NanoL..10.1971S . дои : 10.1021/nl101262u . ПМК 2901229 . ПМИД 20486672 . —Обзор результатов за период 2001–2010 гг.
- Симан, Северная Каролина (2010). «Наноматериалы на основе ДНК» . Ежегодный обзор биохимии . 79 : 65–87. doi : 10.1146/annurev-biochem-060308-102244 . ПМЦ 3454582 . ПМИД 20222824 . — Более полный обзор, включающий как старые, так и новые результаты в этой области.
- Служба РФ (июнь 2011 г.). «ДНК-нанотехнология. ДНК-нанотехнология растет». Наука . 332 (6034): 1140–1, 1143. Бибкод : 2011Sci...332.1140S . дои : 10.1126/science.332.6034.1140 . ПМИД 21636754 . и Служба РФ (июнь 2011 г.). «ДНК-нанотехнологии. Следующий шаг: ДНК-роботы?». Наука . 332 (6034): 1142. doi : 10.1126/science.332.6034.1142 . ПМИД 21636755 . .—Новостная статья, посвященная истории этой области и разработке новых приложений.
- Задеган Р.М., Нортон М.Л. (июнь 2012 г.). «Структурная ДНК-нанотехнология: от дизайна к применению» . Международный журнал молекулярных наук . 13 (6): 7149–7162. дои : 10.3390/ijms13067149 . ПМК 3397516 . ПМИД 22837684 . — Совсем недавний и всесторонний обзор в этой области.
Конкретные подполя:
- Бат Дж., Терберфилд Эй.Дж. (май 2007 г.). «ДНК-наномашины». Природные нанотехнологии . 2 (5): 275–284. Бибкод : 2007NatNa...2..275B . дои : 10.1038/nnano.2007.104 . ПМИД 18654284 . — Обзор наномеханических устройств на основе нуклеиновых кислот.
- Фельдкамп У, Нимейер К.М. (март 2006 г.). «Рациональный дизайн наноархитектур ДНК». Ангеванде Хеми . 45 (12): 1856–1876. дои : 10.1002/anie.200502358 . ПМИД 16470892 . — Обзор с точки зрения проектирования вторичной конструкции.
- Линь С., Лю Ю., Ринкер С., Ян Х. (август 2006 г.). «Самосборка на основе плитки ДНК: построение сложных наноархитектур». ХимияФизХим . 7 (8): 1641–1647. дои : 10.1002/cphc.200600260 . ПМИД 16832805 . —Мини-обзор, посвященный тайловой сборке.
- Чжан Д.Ю., Силиг Г. (февраль 2011 г.). «Динамическая нанотехнология ДНК с использованием реакций смещения цепи». Природная химия . 3 (2): 103–113. Бибкод : 2011НатЧ...3..103Z . дои : 10.1038/nchem.957 . ПМИД 21258382 . — Обзор систем ДНК, использующих механизмы смещения цепей.
Внешние ссылки [ править ]
- Что такое бионанотехнология? — видео-знакомство с нанотехнологиями ДНК
| Обзор | |
|---|---|
| Влияние и применение | |
| Наноматериалы |
|
| Молекулярная самосборка | |
| Наноэлектроника | |
| Сканирующая зондовая микроскопия | |
| Молекулярная нанотехнология | |
| Базы данных органов управления : Национальные |
|---|