Молекулярные модели ДНК


Молекулярные модели структур ДНК представляют собой представления молекулярной геометрии и топологии молекул дезоксирибонуклеиновой кислоты ( ДНК ) с использованием одного из нескольких способов с целью упрощения и представления основных физических и химических свойств молекулярных структур ДНК как in vivo , так и in vivo. пробирка . Эти изображения включают плотно упакованные сферы ( модели CPK ), изготовленные из пластика, металлические проволоки для скелетных моделей , графические вычисления и анимацию на компьютерах, художественную визуализацию. Компьютерные молекулярные модели также позволяют создавать анимацию и моделировать молекулярную динамику, что очень важно для понимания того, как функционирует ДНК in vivo .
Более продвинутые компьютерные молекулярные модели ДНК включают молекулярно-динамическое моделирование и квантово-механические расчеты вибровращения, делокализованных молекулярных орбиталей (МО), электрических дипольных моментов , водородных связей и так далее. Моделирование молекулярной динамики ДНК включает моделирование молекул дезоксирибонуклеиновой кислоты ( ДНК ), геометрии и топологии изменяющихся со временем в результате как внутри-, так и межмолекулярных взаимодействий ДНК. В то время как молекулярные модели молекул ДНК, такие как плотно упакованные сферы (модели CPK), сделанные из пластика или металлических проволок для моделей скелета, являются полезными представлениями статических структур ДНК, их полезность очень ограничена для представления сложной динамики ДНК. Компьютерное молекулярное моделирование позволяет использовать как анимацию, так и моделирование молекулярной динамики, что очень важно для понимания того, как ДНК функционирует in vivo .
История
[ редактировать ]
С самых ранних этапов структурных исследований ДНК с помощью дифракции рентгеновских лучей и биохимических методов молекулярные модели, такие как модель двойной спирали нуклеиновой кислоты Уотсона-Крика, успешно использовались для решения «загадки» структуры ДНК, а также для выяснения того, как последнее связано с его ключевыми функциями в живых клетках. Первые высококачественные рентгенограммы.Об А-ДНК сообщили Розалинд Франклин и Рэймонд Гослинг в 1953 году. [1] Розалинда Франклин сделала критическое наблюдение, что ДНК существует в двух различных формах, А и Б, и создала наиболее четкие изображения обеих форм с помощью метода дифракции рентгеновских лучей. [2] О первых расчетах преобразования Фурье атомной спирали годом ранее сообщили Кохран, Крик и Ванд. [3] За ними в 1953 году последовало вычисление Криком преобразования Фурье спиральной катушки. [4]
Структурная информация получается в результате рентгеноструктурных исследований ориентированных волокон ДНК с помощью молекулярных моделей ДНК в сочетании с кристаллографическим и математическим анализом рентгеновских картин.
Первые сообщения о молекулярной модели структуры B-ДНК с двойной спиралью были сделаны Джеймсом Уотсоном и Фрэнсисом Криком в 1953 году. [5] [6] В том же году Морис Ф. УилкинсА. Стоукс и Х. Р. Уилсон сообщили о первых рентгеновских картинах.in vivo B-ДНК в частично ориентированных головках сперматозоидов лосося. [7]
Разработка Криком и Уотсоном первой правильной молекулярной модели двойной спирали ДНК, возможно, была бы невозможна без биохимических доказательств спаривания нуклеотидных оснований ([A---T]; [C---G]), или Правила Чаргаффа . [8] [9] [10] [11] [12] [13] Хотя такие первоначальные исследования структур ДНК с помощью молекулярных моделей были по существу статичными, их последствия для объяснения in vivo функций ДНК были значительными в области биосинтеза белка и квазиуниверсальности генетического кода. Однако исследования эпигенетической трансформации ДНК in vivo развивались гораздо медленнее, несмотря на их важность для эмбриологии, морфогенеза и исследований рака. Такая химическая динамика и биохимические реакции ДНК гораздо сложнее, чем молекулярная динамика физических взаимодействий ДНК с водой, ионами и белками/ферментами в живых клетках.
Важность
[ редактировать ]Старая динамическая проблема заключается в том, как происходит «самовоспроизведение» ДНК в живых клетках, что должно включать временное раскручивание сверхспиральных волокон ДНК. Хотя ДНК состоит из относительно жестких, очень больших удлиненных молекул биополимера, называемых волокнами или цепями (которые состоят из повторяющихся нуклеотидных единиц четырех основных типов, прикрепленных к дезоксирибозе и фосфатным группам), ее молекулярная структура in vivo претерпевает динамические конфигурационные изменения, которые включают динамически присоединенные Молекулы и ионы воды. Суперспирализация, упаковка гистонов в структурах хромосом и другие подобные супрамолекулярные аспекты также включают in vivo топологию ДНК , которая даже более сложна, чем геометрия молекулы ДНК, что превращает молекулярное моделирование ДНК в особенно сложную проблему как для молекулярных биологов, так и для биотехнологов. Как и другие большие молекулы и биополимеры, ДНК часто существует в нескольких стабильных геометриях (то есть демонстрирует конформационную изомерию ) и конфигурационных квантовых состояниях, которые близки друг к другу по энергии на потенциальной энергетической поверхности молекулы ДНК.
Такие изменяющиеся молекулярные геометрии также могут быть вычислены, по крайней мере в принципе, с использованием ab initio методов квантовой химии , которые могут обеспечить высокую точность для малых молекул, хотя недавно были сделаны заявления о том, что приемлемая точность может быть достигнута также для полинуклеотидов и конформаций ДНК. на основе спектральных данных колебательного кругового дихроизма (VCD). Такая квантовая геометрия определяет важный класс ab initio молекулярных моделей ДНК, исследование которых едва началось, особенно в связи с результатами, полученными методом VCD в растворах. Более подробные сравнения с такими квантовыми расчетами ab initio в принципе можно получить с помощью 2D-FT ЯМР-спектроскопии и исследований релаксации растворов полинуклеотидов или специально меченной ДНК, например, с дейтериевыми метками.
В интересном повороте ролей было предложено использовать молекулу ДНК для квантовых вычислений с помощью ДНК. как наноструктуры ДНК, так и вычислительные биочипы ДНК Были созданы .
Фундаментальные понятия
[ редактировать ]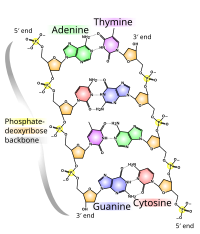
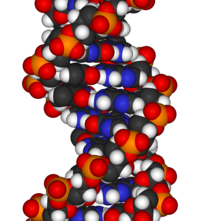
Химической структуры ДНК недостаточно, чтобы понять сложность трехмерных структур ДНК. Напротив, анимированные молекулярные модели позволяют визуально исследовать трехмерную (3D) структуру ДНК. Показанная модель ДНК (крайняя справа) представляет собой заполняющую пространство ( CPK ) модель двойной спирали ДНК. Анимированные молекулярные модели, такие как проволочные или скелетные модели, показанные вверху этой статьи, позволяют визуально исследовать трехмерную (3D) структуру ДНК. Другой тип модели ДНК — это модель заполнения пространства, или CPK.
Динамика водородных связей и протонный обмен сильно различаются на многие порядки между двумя системами полностью гидратированной ДНК и молекул воды во льду. Таким образом, динамика ДНК является сложной и включает в себя наносекундные и несколько десятков пикосекундных масштабов времени, тогда как динамика жидкого льда находится в пикосекундном масштабе времени, а динамика протонного обмена во льду - в миллисекундном масштабе времени. Скорость обмена протонов в ДНК и прикрепленных белках может варьироваться от пикосекунд до наносекунд, минут или лет, в зависимости от точного местоположения обмениваемых протонов в крупных биополимерах.
«Вибрация» простого гармонического осциллятора — это лишь упрощенное динамическое представление продольных колебаний переплетенных спиралей ДНК, которые оказались ангармоничными, а не гармоническими, как это часто предполагается в квантово- динамическом моделировании ДНК.
Структура ДНК
[ редактировать ]Структура ДНК имеет множество форм, как двухцепочечных, так и одноцепочечных. Механические свойства ДНК, которые напрямую связаны с ее структурой, представляют собой существенную проблему для клеток . Каждый процесс, который связывает или считывает ДНК, может использовать или модифицировать механические свойства ДНК в целях распознавания, упаковки и модификации. Чрезвычайная длина ( хромосома может содержать цепь ДНК длиной 10 см), относительная жесткость и спиральная структура ДНК привели к эволюции гистонов таких и ферментов, как топоизомеразы и геликазы, для управления ДНК клетки. Свойства ДНК тесно связаны с ее молекулярной структурой и последовательностью, особенно со слабостью водородных связей и электронных взаимодействий, которые удерживают цепи ДНК вместе, по сравнению с прочностью связей внутри каждой цепи.
Экспериментальные методы, позволяющие напрямую измерять механические свойства ДНК, относительно новы, и визуализация с высоким разрешением в растворе часто затруднена. Тем не менее, ученые обнаружили большое количество данных о механических свойствах этого полимера, а влияние механических свойств ДНК на клеточные процессы является темой активных текущих исследований.
ДНК, обнаруженная во многих клетках, может иметь макроскопическую длину: несколько сантиметров на каждую хромосому человека. Следовательно, клетки должны уплотнять или упаковывать ДНК, чтобы нести ее внутри себя. У эукариот это осуществляется катушечными белками, называемыми гистонами , вокруг которых наматывается ДНК. Дальнейшее уплотнение этого ДНК-белкового комплекса приводит к образованию хорошо известных митотических эукариотических хромосом .
В конце 1970-х годов альтернативные неспиральные модели структуры ДНК кратко рассматривались как потенциальное решение проблем репликации ДНК в плазмидах и хроматине . Однако модели были отброшены в пользу модели двойной спирали из-за последующих экспериментальных достижений, таких как рентгеновская кристаллография дуплексов ДНК, а затем и ядерных частиц нуклеосомы , а также открытие топоизомераз . Такие модели без двойной спирали в настоящее время не принимаются основным научным сообществом. [14] [15]
Определение структуры ДНК с использованием молекулярного моделирования и рентгенограмм ДНК
[ редактировать ]

После того, как ДНК была выделена и очищена стандартными биохимическими методами, у вас есть образец в банке, очень похожий на рисунок в верхней части этой статьи. Ниже приведены основные этапы получения структурной информации на основе рентгеновских дифракционных исследований ориентированных волокон ДНК, полученных из гидратированного образца ДНК, с помощью молекулярных моделей ДНК в сочетании с кристаллографическим и математическим анализом рентгеновских картин. .
Модели паракристаллической решетки структур B-ДНК
[ редактировать ]
Паракристаллическая решетка, или паракристалл, представляет собой молекулярную или атомную решетку со значительными уровнями (например, более нескольких процентов) частичного разупорядочения молекулярных структур. Предельными случаями паракристаллической модели являются наноструктуры , такие как стекла , жидкости и т. д., которые могут обладать только локальным упорядочением и не иметь глобального порядка. Простой пример паракристаллической решетки для кварцевого стекла показан на следующем рисунке:
Жидкие кристаллы также имеют паракристаллическую, а не кристаллическую структуру.
Высокогидратированная B-ДНК естественным образом встречается в живых клетках в таком паракристаллическом состоянии, которое является динамическим, несмотря на относительно жесткую двойную спираль ДНК, стабилизированную параллельными водородными связями между парами нуклеотидных оснований в двух комплементарных спиральных цепях ДНК (см. рисунки). ). Для простоты в большинстве молекулярных моделей ДНК не учитываются вода и ионы, динамически связанные с B-ДНК, и, таким образом, они менее полезны для понимания динамического поведения B-ДНК in vivo . Физико-математический анализ рентгеновских лучей [16] [17] и спектроскопические данные для паракристаллической B-ДНК, таким образом, гораздо более сложны, чем данные рентгеновской дифракции кристаллической A-ДНК. Паракристаллическая модель также важна для технологических приложений ДНК, таких как нанотехнология ДНК . В настоящее время также разрабатываются новые методы, сочетающие дифракцию рентгеновских лучей ДНК с рентгеновской микроскопией в гидратированных живых клетках. [18]
Геномные и биотехнологические применения молекулярного моделирования ДНК
[ редактировать ]
Существуют различные применения молекулярного моделирования ДНК в исследовательских приложениях в области геномики и биотехнологии: от восстановления ДНК до ПЦР и наноструктур ДНК . Двумерные массивы соединений ДНК были визуализированы с помощью атомно-силовой микроскопии . [19]
Молекулярное моделирование ДНК находит разнообразное применение в геномике и биотехнологии , а его исследовательские применения варьируются от восстановления ДНК до ПЦР и наноструктур ДНК. К ним относятся компьютерные молекулярные модели таких разнообразных молекул, как РНК-полимераза, E. coli, бактериальная ДНК-примаза, предполагающая очень сложную динамику на границах между ферментами и ДНК-матрицей, а также молекулярные модели мутагенного химического взаимодействия мощных канцерогенных молекул. с ДНК. Все они представлены в галерее ниже.
Технологические приложения включают биочипы ДНК и наноструктуры ДНК, предназначенные для вычислений ДНК и других динамических применений нанотехнологии ДНК . [20] [21] [22] [23] [24] [25] Изображение справа представляет собой самоорганизующиеся наноструктуры ДНК. «Плиточная» структура ДНК на этом изображении состоит из четырех разветвленных соединений, ориентированных под углами 90°. Каждая плитка состоит из девяти олигонуклеотидов ДНК, как показано на рисунке; такие плитки служат основным «строительным блоком» для сборки наносеток ДНК, показанных на микрофотографии АСМ.
Квадруплексная ДНК может быть вовлечена в развитие некоторых видов рака. [26] [27] Изображения квадруплексной ДНК находятся в галерее ниже.
Галерея моделей ДНК
[ редактировать ] Общая модель вращающейся ДНК.
Общая модель вращающейся ДНК. Упрощенный рисунок структуры двойной спирали А-ДНК.
Упрощенный рисунок структуры двойной спирали А-ДНК. Модель репликации ДНК, основанная на концепции двойной спирали.
Модель репликации ДНК, основанная на концепции двойной спирали. Анимированная заполняющая пространство молекулярная модель двойной спирали B-ДНК.
Анимированная заполняющая пространство молекулярная модель двойной спирали B-ДНК. Крупномасштабная модель ДНК Крика-Уотсона, представленная в Музее принца Фелипе.
Крупномасштабная модель ДНК Крика-Уотсона, представленная в Музее принца Фелипе. Вид сбоку на молекулярные модели A-, B-, Z-ДНК.
Вид сбоку на молекулярные модели A-, B-, Z-ДНК. Упрощенная модель двойной спирали А-ДНК.
Упрощенная модель двойной спирали А-ДНК. Молекулярное моделирование РНК-полимеразы.
Молекулярное моделирование РНК-полимеразы. Молекулярное моделирование матрицы бактериальной ДНК-примазы.
Молекулярное моделирование матрицы бактериальной ДНК-примазы. Молекулярное моделирование взаимодействия ДНК с молекулой канцерогена МГМТ.
Молекулярное моделирование взаимодействия ДНК с молекулой канцерогена МГМТ. 3D-молекулярная модель ДНК, поврежденной канцерогенным 2-аминофлуореном (АФ).
3D-молекулярная модель ДНК, поврежденной канцерогенным 2-аминофлуореном (АФ). Рис.6. Молекулярное моделирование репарации ДНК
Рис.6. Молекулярное моделирование репарации ДНК Анимированная скелетная модель А-ДНК.
Анимированная скелетная модель А-ДНК. Упрощенные модели хроматина.
Упрощенные модели хроматина. Упрощенная модель строения хромосом.
Упрощенная модель строения хромосом. Гипотетический квадруплекс структур ДНК, богатых гуанином, которые могут быть вовлечены в развитие рака.
Гипотетический квадруплекс структур ДНК, богатых гуанином, которые могут быть вовлечены в развитие рака. Трехмерная молекулярная структура внутримолекулярного теломерного G-квадруплекса человека в растворе калия.
Трехмерная молекулярная структура внутримолекулярного теломерного G-квадруплекса человека в растворе калия. Молекулярная модель заполнения пространства ДНК
Молекулярная модель заполнения пространства ДНК Модель спроектированного тетраэдра ДНК.
Модель спроектированного тетраэдра ДНК. Модель ДНК длиной 15 м, Центр биоразнообразия Naturalis
Модель ДНК длиной 15 м, Центр биоразнообразия Naturalis


















См. также
[ редактировать ]- G-квадруплекс
- Кристаллография
- Кристаллические решетки
- База данных свойств динуклеотидов ( DiProDB ), предназначенная для сбора и анализа термодинамических, структурных и других характеристик динуклеотидов.
- Рентгеновская микроскопия
- Рассеяние рентгеновских лучей
- Рассеяние нейтронов
- Последовательность нуклеиновой кислоты
- Колебательный круговой дихроизм (ВКД)
- Рамановская спектроскопия - микроскопия и когерентная антистоксова рамановская спектроскопия (CARS)
- Сэр Лоуренс Брэгг , ФРС
- Сравнение программного обеспечения для моделирования нуклеиновых кислот
- ЯНТАРЬ
- ШАРММ
- Морское ушко (молекулярная механика)
- Программа визуализации Сириус
- QMC@Домой
- ЯМР-спектроскопия (Фурье-ЯМР)
- ЯМР-визуальная микроскопия
- Микроволновая спектроскопия
- FT- инфракрасный (ИК)
- FT- спектроскопия ближнего инфракрасного диапазона (NIR)
- Спектральная визуализация , гиперспектральная визуализация , химическая визуализация
- Флуоресцентная корреляционная спектроскопия
- Флуоресцентная кросс-корреляционная спектроскопия и резонансный перенос энергии Фёрстера (FRET)
- Конфокальная микроскопия
- Устаревшие модели структуры ДНК
Ссылки
[ редактировать ]- ^ Франклин, Р.Э., Гослинг, Р.Г. (6 марта 1953 г.). «Структура волокон тимонуклеата натрия I. Влияние содержания воды» . Акта Кристаллогр . 6 (8): 673. дои : 10.1107/S0365110X53001939 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )
Франклин, Р.Э., Гослинг, Р.Г. (6 марта 1953 г.). «Структура волокон тимонуклеата натрия II. Цилиндрически симметричная функция Паттерсона» . Акта Кристаллогр . 6 (8): 678. дои : 10.1107/S0365110X53001940 .{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хаммер, ДИН Х.; Мэддокс, Бренда (2002). «Извилистая дорога к двойной спирали». Научный американец . 287 (6): 127–128. Бибкод : 2002SciAm.287f.127H . doi : 10.1038/scientificamerican1202-127 . JSTOR 26060090 .
- ^ Кохран, В., Крик, ФХК, Ванд В. (1952). «Строение синтетических полипептидов. 1. Преобразование атомов на спирали» . Акта Кристаллогр . 5 (5): 581–6. дои : 10.1107/S0365110X52001635 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Крик, FHC (1953a). «Преобразование Фурье спиральной катушки» . Акта Кристаллогр . 6 (8–9): 685–9. дои : 10.1107/S0365110X53001952 .
- ^ Уотсон, Джеймс Д., Крик, Фрэнсис Х.К. (25 апреля 1953 г.). «Структура нуклеиновой кислоты дезоксирибозы» (PDF) . Природа . 171 (4356): 737–8. Бибкод : 1953Natur.171..737W . дои : 10.1038/171737a0 . ПМИД 13054692 . S2CID 4253007 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) , . - ^ Уотсон, доктор медицинских наук; Крик FHC (1953b). «Структура ДНК». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 18 : 123–31. дои : 10.1101/SQB.1953.018.01.020 . ПМИД 13168976 .
- ^ Уилкинс МХФ, А.Р. Стоукс А.Р. и Уилсон, Х.Р. (1953). «Молекулярная структура дезоксипентозных нуклеиновых кислот» (PDF) . Природа . 171 (4356): 738–40. Бибкод : 1953Natur.171..738W . дои : 10.1038/171738a0 . ПМИД 13054693 . S2CID 4280080 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Элсон Д., Чаргафф Э. (1952). «О содержании дезоксирибонуклеиновой кислоты в гаметах морского ежа». Эксперименты . 8 (4): 143–5. дои : 10.1007/BF02170221 . ПМИД 14945441 . S2CID 36803326 .
- ^ Чаргафф Э., Липшиц Р., Грин С. (1952). «Состав дезоксипентозных нуклеиновых кислот четырех родов морских ежей» . J Биол Хим . 195 (1): 155–160. дои : 10.1016/S0021-9258(19)50884-5 . ПМИД 14938364 .
- ^ Чаргафф Э., Липшиц Р., Грин С., Ходс М.Э. (1951). «Состав дезоксирибонуклеиновой кислоты спермы лосося» . J Биол Хим . 192 (1): 223–230. дои : 10.1016/S0021-9258(18)55924-X . ПМИД 14917668 .
- ^ Чаргафф Э (1951). «Некоторые недавние исследования состава и структуры нуклеиновых кислот». J Cell Physiol Suppl . 38 (Приложение).
- ^ Магасаник Б., Вишер Э., Донигер Р., Элсон Д., Чаргафф Э. (1950). «Разделение и оценка рибонуклеотидов в ничтожных количествах» . J Биол Хим . 186 (1): 37–50. дои : 10.1016/S0021-9258(18)56284-0 . ПМИД 14778802 .
- ^ Чаргафф Э (1950). «Химическая специфичность нуклеиновых кислот и механизм их ферментативной деградации». Эксперименты . 6 (6): 201–9. дои : 10.1007/BF02173653 . ПМИД 15421335 . S2CID 2522535 .
- ^ Стоукс, Т.Д. (1982). «Двойная спираль и искривленная молния — образцовая история». Социальные исследования науки . 12 (2): 207–240. дои : 10.1177/030631282012002002 . ПМИД 11620855 . S2CID 29369576 .
- ^ Гаутам, Н. (25 мая 2004 г.). «Ответ на «Разнообразие вторичной структуры ДНК» » (PDF) . Современная наука . 86 (10): 1352–1353 . Проверено 25 мая 2012 г.
Однако открытие топоизомераз сняло остроту топологического возражения против плектонемической двойной спирали. Более позднее решение монокристаллической рентгеновской структуры коровой частицы нуклеосомы показало почти 150 пар оснований ДНК (т.е. около 15 полных оборотов) со структурой, которая во всех существенных отношениях совпадает со структурой модели Уотсона-Крика. . Это нанесло смертельный удар идее о том, что другие формы ДНК, особенно двуспиральная ДНК, существуют как нечто иное, чем локальные или временные структуры.
[ мертвая ссылка ] - ^ Хоземанн Р., Багчи Р.Н., Прямой анализ дифракции на веществе , издательства Северной Голландии, Амстердам – Нью-Йорк, 1962.
- ^ Баяну, IC (1978). «Рассеяние рентгеновских лучей частично неупорядоченными мембранными системами» . Акта Кристаллогр. А. 34 (5): 751–3. Бибкод : 1978AcCrA..34..751B . дои : 10.1107/S0567739478001540 .
- ^ Ямамото Ю., Синохара К. (октябрь 2002 г.). «Применение рентгеновской микроскопии в анализе живых гидратированных клеток». Анат. Рек . 269 (5): 217–23. дои : 10.1002/ar.10166 . ПМИД 12379938 . S2CID 43009840 .
- ^ Мао, Чэндэ; Сунь, Вэйцюн; Симан, Надриан К. (16 июня 1999 г.). «Спроектированные двумерные массивы соединений Холлидея ДНК, визуализированные с помощью атомно-силовой микроскопии». Журнал Американского химического общества . 121 (23): 5437–43. дои : 10.1021/ja9900398 .
- ^ Робинсон, Брюш Х.; Симан, Надриан К. (август 1987 г.). «Дизайн биочипа: самособирающееся устройство памяти молекулярного масштаба». Белковая инженерия . 1 (4): 295–300. дои : 10.1093/белок/1.4.295 . ISSN 0269-2139 . ПМИД 3508280 . Связь
- ^ Ротемунд, Пол В.К .; Экани-Нкодо, Аксель; Пападакис, Ник; Кумар, Ашиш; Файгенсон, Дебора Кучнир; Уинфри, Эрик (22 декабря 2004 г.). «Дизайн и характеристика программируемых ДНК-нанотрубок». Журнал Американского химического общества . 126 (50): 16344–52. дои : 10.1021/ja044319l . ПМИД 15600335 .
- ^ Керен, К.; Ротем С. Берман; Евгений Бухштаб; Ури Сиван ; Эрез Браун (ноябрь 2003 г.). «Полевой транзистор из углеродных нанотрубок на основе ДНК». Наука . 302 (6549): 1380–2. Бибкод : 2003Sci...302.1380K . CiteSeerX 10.1.1.454.2291 . дои : 10.1126/science.1091022 . ПМИД 14631035 . S2CID 29794525 .
- ^ Чжэн, Цзивэнь; Константину, Памела Э.; Майкл, Кристина; Аливисатос, А. Пол; Киль, Ричард А.; Симан Надриан К. (2006). «2D-массивы наночастиц демонстрируют организационную силу надежных мотивов ДНК» . Нано-буквы . 6 (7): 1502–4. Бибкод : 2006NanoL...6.1502Z . дои : 10.1021/nl060994c . ПМЦ 3465979 . ПМИД 16834438 .
- ^ Коэн, Джастин Д.; Садовски, Джон П.; Дерван, Питер Б. (2007). «Обращение к одиночным молекулам в наноструктурах ДНК» . Angewandte Chemie, международное издание . 46 (42): 7956–9. дои : 10.1002/anie.200702767 . ПМИД 17763481 .
- ^ Константину, Памела Э.; Ван, Тонг; Копач, Йенс; Израиль, Лиза Б.; Чжан, Сяопин; Дин, Баоцюань; Шерман, Уильям Б.; Ван, Син; Чжэн, Цзяньпин; Ша, Руоцзе; Симан, Надриан К. (2006). «Двойная когезия в структурных нанотехнологиях ДНК» . Органическая и биомолекулярная химия . 4 (18): 3414–9. дои : 10.1039/b605212f . ПМК 3491902 . ПМИД 17036134 .
- ^ «Отдел физики Кавендишской лаборатории – молекулярная биофизика» . Архивировано из оригинала 23 мая 2009 года . Проверено 17 мая 2009 г.
- ^ «Криптовалюты и физика – Планефизика» . Архивировано из оригинала 31 марта 2009 года . Проверено 17 мая 2009 г.
Дальнейшее чтение
[ редактировать ]- Применение новых технологий в производстве продуктов здорового питания, медицинской и сельскохозяйственной биотехнологии . (июнь 2004 г.) И. К. Байану, П. Р. Лозано, В. И. Присекару и Х. К. Лин., q-bio/0406047.
- Бессель Ф., Исследование части планетарных возмущений , Берлинские трактаты (1824), статья 14.
- Сэр Лоуренс Брэгг, ФРС. Кристаллическое состояние, общий обзор. Лондон: G. Bells and Sons, Ltd., тт. 1 и 2., 1966., 2024 стр.
- Кантор, Ч.Р. и Шиммель, П.Р. Биофизическая химия, части I и II ., Сан-Франциско: WH Freeman and Co., 1980. 1800 страниц.
- Воэт, Д. и Дж. Г. Воэт. Биохимия , 2-е изд., Нью-Йорк, Торонто, Сингапур: John Wiley & Sons, Inc., 1995, ISBN 0-471-58651-X ., 1361 стр.
- Уотсон, Дж. Н. Трактат по теории функций Бесселя ., (1995) Издательство Кембриджского университета. ISBN 0-521-48391-3 .
- Уотсон, Джеймс Д. Молекулярная биология гена. Нью-Йорк и Амстердам: WA Benjamin, Inc., 1965, 494 страницы.
- Вентворт, МЫ Физическая химия. Короткий курс. , Молден (Массачусетс): Blackwell Science, Inc., 2000.
- Герберт Р. Уилсон, ФРС. Дифракция рентгеновских лучей на белках, нуклеиновых кислотах и вирусах ., Лондон: Edward Arnold (Publishers) Ltd., 1966.
- Курт Вутрих. ЯМР белков и нуклеиновых кислот. , Нью-Йорк, Брисбен, Чичестер, Торонто, Сингапур: J. Wiley & Sons. 1986., 292 страницы.
- Халлин П.Ф., Дэвид Ассери Д. (2004). «База данных Атласа генома CBS: динамическое хранилище биоинформатических результатов и данных о последовательностях ДНК» . Биоинформатика . 20 (18): 3682–6. doi : 10.1093/биоинформатика/bth423 . ПМИД 15256401 .
- Чжан CT, Чжан Р, Оу HY (2003). «База данных Z-кривых: графическое представление последовательностей генома» . Биоинформатика . 19 (5): 593–599. doi : 10.1093/биоинформатика/btg041 . ПМИД 12651717 .
Внешние ссылки
[ редактировать ]- ДНК: игра «Двойная спираль» С официального сайта Нобелевской премии.
- MDDNA: Структурная биоинформатика ДНК
- Двойная спираль 1953–2003 Национальный центр биотехнологического образования
- DNAlive: веб-интерфейс для расчета физических свойств ДНК . Также позволяет сопоставлять результаты с браузером UCSC Genome и динамикой ДНК.
- Дальнейшие подробности математического и молекулярного анализа структуры ДНК на основе рентгеновских данных.
- Функции Бесселя, соответствующие преобразованиям Фурье атомных или молекулярных спиралей. [ мертвая ссылка ]
- обзор принципов STM/AFM/SNOM с обучающими видеороликами
Базы данных молекулярных моделей и последовательностей ДНК
[ редактировать ]- Рентгеновская дифракция
- Идентификатор NDB: UD0017 База данных
- Рентгеновский Атлас - база данных
- PDB-файлы координат структур нуклеиновых кислот по данным дифракции рентгеновских лучей на кристаллах NA (включая ДНК).
- Файлы структурных факторов, загружаемые в формате CIF.
- Рассеяние нейтронов
- Источник нейтронов ISIS : Импульсный источник нейтронов ISIS: Всемирный научный центр с нейтронами и мюонами в Харвелле, недалеко от Оксфорда, Великобритания.
- Рентгеновская микроскопия
- Электронная микроскопия
- Базы данных ЯМР
- Атлас ЯМР - база данных
- mmcif загружаемые файлы координат нуклеиновых кислот в растворе по данным 2D-FT ЯМР
- Файлы ограничений ЯМР для NA в формате PDB
- Геномные и структурные базы данных
- База данных атласа генома CBS - содержит примеры искажений базисов.
- База данных геномов Z-кривых — инструмент трехмерной визуализации и анализа геномов .
- Молекулярные модели ДНК и других нуклеиновых кислот: файлы координат моделей молекулярной структуры нуклеиновых кислот в форматах PDB и CIF.
- Атомно-силовая микроскопия
Виды нуклеиновых кислот |
|---|