G-квадруплекс
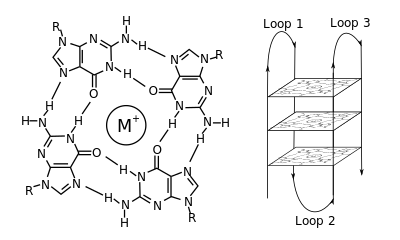
В молекулярной биологии G-квадруплекса вторичные структуры (G4) образуются в нуклеиновых кислотах с помощью последовательностей, богатых гуанином . [2] Они имеют спиральную форму и содержат гуаниновые тетрады, которые могут образовываться из одной, [3] два [4] или четыре нити. [5] Мономолекулярные формы часто встречаются в природе вблизи концов хромосом, более известных как теломерные области, а также в областях регуляции транскрипции множества генов, как у микробов, так и в регионах, регулирующих транскрипцию. [6] [7] и среди позвоночных [8] [7] включая онкогены у человека. [9] Четыре основания гуанина могут связываться посредством водородной связи Хугстина с образованием плоской квадратной структуры, называемой тетрадой гуанина (G-тетрада или G-квартет), а две или более тетрады гуанина (из G-трактов, непрерывных цепочек гуанина) могут складываться сверху. друг друга, образуя G-квадруплекс.
Расположение и соединение с образованием G-квадруплексов не случайно и служат весьма необычным функциональным целям. Структура квадруплекса дополнительно стабилизируется наличием катиона , особенно калия , который находится в центральном канале между каждой парой тетрад. [3] Они могут состоять из ДНК , РНК , LNA и PNA и могут быть внутримолекулярными , бимолекулярными или тетрамолекулярными. [10] В зависимости от направления нитей или частей нитей, образующих тетрады, структуры можно охарактеризовать как параллельные или антипараллельные . Структуры G-квадруплексов можно предсказать с помощью вычислений на основе мотивов последовательностей ДНК или РНК. [11] [12] но их фактическая структура может весьма варьироваться внутри мотивов и между ними, число которых может превышать 100 000 на геном. Их деятельность в основных генетических процессах является активной областью исследований в области теломер, регуляции генов и исследований функциональной геномики. [13] [14]
История
[ редактировать ]Идентификация структур с высокой ассоциацией гуанина стала очевидной в начале 1960-х годов благодаря идентификации гелеобразных веществ, связанных с гуанинами. [15] В частности, это исследование детализировало четырехцепочечные структуры ДНК с высокой ассоциацией гуанинов, которые позже были идентифицированы в теломерных областях ДНК эукариот в 1980-х годах. [16] Важность открытия структуры G-квадруплекса была описана в заявлении: «Если G-квадруплексы образуются так легко in vitro , природа нашла способ использовать их in vivo » - Аарон Клуг , лауреат Нобелевской премии по химии (1982). Интерес к функции G-квадруплексов in vivo резко возрос после того, как крупномасштабный полногеномный анализ показал преобладание потенциальных последовательностей, образующих G-квадруплекс (pG4), в промоторах генов человека, шимпанзе, мыши и крысы, представленных на Первом Международном G-квадруплексе (pG4). -квадруплекс Встреча состоялась в апреле 2007 года в Луисвилле, Кентукки. [7] В 2006 году сообщалось о преобладании G-квадруплексов в промоторах генов нескольких бактериальных геномов, что предсказывает регуляцию генов, опосредованную G-квадруплексами. [6] Учитывая обилие G-квадруплексов in vivo , эти структуры играют биологически значимую роль посредством взаимодействия с промоторными областями онкогенов и теломерными областями нитей ДНК. Текущие исследования заключаются в выявлении биологической функции этих структур G-квадруплекса для конкретных онкогенов и открытии эффективных терапевтических методов лечения рака, основанных на взаимодействии с G-квадруплексами. Ранние доказательства образования G-квадруплексов in vivo в клетках были установлены путем их выделения из клеток. [17] а позже благодаря наблюдению, что специфические ДНК-хеликазы могут быть идентифицированы там, где в клетках накапливаются небольшие молекулы, специфичные для этих структур ДНК. [18]
Топология
[ редактировать ]Длина последовательностей нуклеиновых кислот, участвующих в образовании тетрад, определяет, как сворачивается квадруплекс. Короткие последовательности, состоящие только из одной непрерывной цепочки трех или более гуаниновых оснований, требуют четырех отдельных цепей для образования квадруплекса. Такой квадруплекс описывается как тетрамолекулярный, что отражает необходимость наличия четырех отдельных цепей. Термин ДНК G4 первоначально был зарезервирован для этих тетрамолекулярных структур, которые могут играть роль в мейозе . [5] Однако в настоящее время термин G4, используемый в молекулярной биологии, может означать G-квадруплексы любой молекулярности. Более длинные последовательности, которые содержат две смежные серии из трех или более гуаниновых оснований, где гуаниновые области разделены одним или несколькими основаниями, требуют только двух таких последовательностей, чтобы обеспечить достаточное количество гуаниновых оснований для образования квадруплекса. Эти структуры, образованные из двух отдельных нитей, богатых G, называются бимолекулярными квадруплексами. Наконец, последовательности, которые содержат четыре различных ряда гуаниновых оснований, сами по себе могут образовывать стабильные квадруплексные структуры, а квадруплекс, образованный полностью из одной цепи, называется внутримолекулярным квадруплексом. [19]
В зависимости от того, как отдельные группы гуаниновых оснований расположены в бимолекулярном или внутримолекулярном квадруплексе, квадруплекс может принимать одну из множества топологий с различными конфигурациями петель. [20] Если все нити ДНК идут в одном направлении, квадруплекс называется параллельным. Для внутримолекулярных квадруплексов это означает, что любые присутствующие петлевые области должны быть пропеллерного типа, расположенными по бокам квадруплекса. Если одна или несколько цепочек гуаниновых оснований имеют направление 5'-3', противоположное другим цепочкам гуаниновых оснований, говорят, что квадруплекс принял антипараллельную топологию. Петли, соединяющие цепочки гуаниновых оснований во внутримолекулярных антипараллельных квадруплексах, бывают либо диагональными, соединяющими две диагонально противоположные цепочки гуаниновых оснований, либо петлями латерального (реберного) типа, соединяющими две соседние цепочки пар оснований гуанина.
В квадруплексах, образованных из двухцепочечной ДНК, также обсуждались возможные межцепочечные топологии. [21] . [22] Межнитевые квадруплексы содержат гуанины, происходящие из обеих цепей дцДНК.
Структура и функциональная роль в геноме
[ редактировать ]человека После секвенирования генома было обнаружено множество последовательностей, богатых гуанином, которые потенциально могли образовывать квадруплексы. [23] опосредующие факторы, такие как ДНК-связывающие белки на хроматине , состоящие из ДНК, плотно намотанной вокруг белков -гистонов В зависимости от типа клеток и клеточного цикла на динамическое образование квадруплексов влияют , а также другие условия окружающей среды и стрессы. Например, количественные оценки термодинамики молекулярного краудинга показывают , что антипараллельный g-квадруплекс стабилизируется молекулярным краудингом. [24] Этот эффект, по-видимому, опосредован изменением гидратации ДНК и ее влиянием на пар оснований Хугстина . связывание [25] Эти квадруплексы, казалось, легко возникали на концах хромосом . Кроме того, склонность к образованию g-квадруплексов во время транскрипции в последовательностях РНК с потенциалом образования взаимоисключающих структур шпилек или G-квадруплексов сильно зависит от положения последовательности, образующей шпильку. [26]
Поскольку ферменты репарации естественным образом распознают концы линейных хромосом как поврежденную ДНК и обрабатывают их как таковые, оказывая вредное воздействие на клетку, на концах линейных хромосом необходимы четкая передача сигналов и жесткая регуляция. Теломеры обеспечивают эту передачу сигналов. Теломеры, богатые гуанином и склонные к образованию g-квадруплексов, расположены на концевых концах хромосом и помогают поддерживать целостность генома, защищая эти уязвимые концевые концы от нестабильности.
Эти теломерные регионы характеризуются длинными участками двухцепочечных повторов CCCTAA:TTAGGG. Повторы заканчиваются 3'-выступом, содержащим от 10 до 50 одноцепочечных повторов TTAGGG. Гетеродимерный комплекс рибонуклеопротеинового фермента теломеразы добавляет повторы TTAGGG на 3'-конце нити ДНК. На этих 3'-концевых выступах G-богатый выступ может образовывать вторичные структуры, такие как G-квадруплексы, если выступ длиннее четырех повторов TTAGGG. Наличие этих структур предотвращает удлинение теломер теломеразным комплексом. [27]
Теломерные квадруплексы
[ редактировать ]Было показано, что теломерные повторы у различных организмов образуют эти квадруплексные структуры in vitro , а впоследствии также было показано, что они формируются in vivo . [28] [29] Теломерный повтор человека (одинаковый для всех позвоночных ) состоит из множества секвенированных повторов (TTAGGG), а квадруплексы, образованные этой структурой, могут иметь форму бусинок размером от 5 до 8 нм и хорошо изучены. изучен методами ЯМР , ПЭМ и рентгеновского определения кристаллической структуры . [30] Было показано, что образование этих квадруплексов в теломерах снижает активность фермента теломеразы , который отвечает за поддержание длины теломер и участвует примерно в 85% всех случаев рака . Это активная мишень для открытия новых лекарств, в том числе теломестатина .
Нетеломерные квадруплексы
[ редактировать ]Квадруплексы присутствуют не только в теломерах, но и в других местах . Анализ геномов человека, шимпанзе, мыши и крысы показал огромное количество потенциальных последовательностей, образующих G-квадруплекс (pG4), в нетеломерных областях. Большое количество нетеломерных G-квадруплексов было обнаружено в промоторах генов и консервативно у всех видов. [6] [7] Точно так же большое количество G-квадруплексов было обнаружено в геномах E. coli и сотнях других микробов. Здесь также, как и у позвоночных, G-квадруплексы были обогащены промоторами генов. [6] Кроме того, у растений и водорослей был обнаружен консервативный локус G-квадруплекса, сохраняющийся более одного миллиарда лет, в гене, кодирующем большую субъединицу РНК-полимеразы II. [31] Хотя эти исследования предсказали регуляцию генов, опосредованную G-квадруплексом, маловероятно, что все pG4 будут образовываться in vivo. Протоонкоген области , c-myc образует квадруплекс в гиперчувствительной к нуклеазе критической для активности гена. [32] [33] Другие гены, которые, как показано, образуют G-квадруплексы в своих промоторных областях, включают курицы β-глобина ген , человеческую убиквитин -лигазу RFP2 и протоонкогены c-kit , bcl-2 , VEGF , H-ras и N-ras . [34] [35] [36]
геноме Были проведены полногеномные исследования, основанные на правиле квадруплексной складки, которые выявили 376 000 предполагаемых квадруплексных последовательностей (PQS) в , человека хотя не все из них, вероятно, формируются in vivo . [37] Подобные исследования выявили предполагаемые G-квадруплексы у прокариот , а именно у бактерии E. coli . [38] Существует несколько возможных моделей того, как квадруплексы могут влиять на активность генов путем усиления или подавления . Одна модель показана ниже: образование G-квадруплекса внутри или рядом с промотором блокирует транскрипцию гена и, следовательно, деактивирует его. В другой модели квадруплекс, образующийся на некодирующей цепи ДНК, помогает поддерживать открытую конформацию кодирующей цепи ДНК и усиливать экспрессию соответствующего гена.
Функция
[ редактировать ]Было высказано предположение, что образование квадруплексов играет роль в переключении тяжелой цепи иммуноглобулина . [5] Поскольку клетки развили механизмы разрешения (т. е. раскручивания) образующихся квадруплексов. Образование квадруплексов может быть потенциально разрушительным для клетки; хеликазы WRN и белок синдрома Блума обладают высоким сродством к расщеплению G-квадруплексов ДНК. [39] Хеликаза DEAH/RHA, DHX36 , также была идентифицирована как ключевая резольваза G-квадруплекса. [40] [41] В 2009 году было обнаружено, что белок-супрессор метастазирования NM23H2 (также известный как NME2) напрямую взаимодействует с G-квадруплексом в промоторе гена c-myc и регулирует транскрипцию c-myc. [42] [43] Совсем недавно сообщалось, что NM23H2 взаимодействует с G-квадруплексом в промоторе гена теломеразы человека (hTERT) и регулирует экспрессию hTERT. [44] В 2019 году с помощью TRF2 ChIP-seq было показано, что теломер-связывающий фактор-2 (TRF2 или TERF2) связывается с тысячами нетеломерных G-квадруплексов в геноме человека. [45] Существует множество исследований, которые подтверждают участие квадруплексов как в положительной, так и в отрицательной регуляции транскрипции, включая эпигенетическую регуляцию таких генов, как hTERT. [44] Сообщалось также, что функция G-квадруплексов позволяет запрограммировать рекомбинацию тяжелых генов иммуноглобина и системы антигенных вариаций пилина патогенных Neisseria . [46] Роль квадруплексной структуры в контроле трансляции недостаточно изучена. Прямая визуализация структур G-квадруплексов в клетках человека [47] а также сокристаллическая структура РНК-хеликазы, связанной с G-квадруплексом. [48] предоставили важные подтверждения их значимости для клеточной биологии. Потенциальная положительная и отрицательная роль квадруплексов в репликации и функционировании теломер остается спорной. Т-петли и G-квадруплексы описываются как две третичные структуры ДНК, которые защищают концы теломер и регулируют длину теломер. [49]
Регуляция генома посредством формирования структур G-квадруплекса
[ редактировать ]Многие регуляторные процессы генома связаны с образованием структур G-квадруплекса, что объясняется огромной ролью, которую он играет в репарации ДНК апуриновых/апиримидиновых сайтов, также известных как AP-сайты. [50] Был разработан новый метод картирования AP-сайтов, известный как AP-seq, в котором используется меченный биотином альдегид-реактивный зонд (ARP) для маркировки определенных областей генома, где повреждение AP-сайтов было значительным. [51] Для картирования обоих был использован другой метод полногеномного секвенирования, известный как ChIP-секвенирование ; повреждение сайтов AP и фермент, ответственный за его восстановление, эндонуклеаза 1 AP (APE1). Оба эти метода полногеномного секвенирования, ChIP-секвенирование и ARP, показали, что возникновение повреждений AP-сайта неслучайно. Повреждение AP-сайта также было более распространено в определенных областях генома, которые содержат специфические активные маркеры промотора и энхансера, некоторые из которых были связаны с областями, ответственными за аденокарциному легких и рак толстой кишки. [52] Было обнаружено, что повреждение AP-сайта преобладает в PQS-районах генома, где образование G-квадруплексных структур регулируется и стимулируется процессом репарации ДНК – эксцизионной репарацией оснований (BER). [52] Доказано, что процессы эксцизионной репарации базы в клетках снижаются с возрастом, поскольку ее компоненты в митохондриях начинают уменьшаться, что может привести к формированию многих заболеваний, таких как болезнь Альцгеймера (БА). [53] Говорят, что эти G-квадруплексные структуры образуются в промоторных областях ДНК за счет сверхспиральности, которая способствует раскручиванию двойной спиральной структуры ДНК и, в свою очередь, закручивает цепи с образованием G-квадруплексных структур в богатых гуанином регионах. [54] Сигнал о пути BER подается, когда он указывает на окислительное повреждение основания ДНК, где такие структуры, как 8-оксогуанин-ДНК-гликозилаза 1 (OGG1), APE1 и G-квадруплекс, играют огромную роль в его восстановлении. Эти ферменты участвуют в BER для восстановления определенных повреждений ДНК, таких как 7,8-дигидро-8-оксогуанин (8-oxoG), который образуется при окислительном стрессе с образованием гуаниновых оснований. [55]
Роль эндогенного окисленного повреждения основания ДНК в формировании G4
[ редактировать ]Основания гуанина (G) в G-квадруплексе имеют самый низкий окислительно-восстановительный потенциал, что делает его более восприимчивым к образованию 8-оксогуанина (8-oxoG), эндогенного окисленного повреждения основания ДНК в геноме. Из-за того, что гуанин имеет более низкий потенциал восстановления электронов, чем другие нуклеотидные основания, [56] 8-оксо-2'-дезоксигуанозин (8-оксо-dG) является известным основным продуктом окисления ДНК. Его концентрация используется для измерения окислительного стресса внутри клетки. [57] Когда ДНК подвергается окислительному повреждению, возможное структурное изменение гуанина после ионизирующего излучения приводит к образованию енольной формы 8-OH-Gua. Этот окислительный продукт образуется в результате таутомерного сдвига исходного поврежденного гуанина, 8-оксо-Гуа, и представляет собой повреждение ДНК, вызывающее изменения в структуре. Эта форма позволяет базовой эксцизионной репарации (BER) ферменту OGG1 связывать и удалять окислительные повреждения с помощью APE1, что приводит к образованию AP-сайта. [55] [53] Более того, AP-сайт — это участок ДНК, который не имеет ни пуринового, ни пиримидинового основания из-за повреждения ДНК. Это наиболее распространенный тип эндогенного повреждения ДНК в клетках. AP-сайты могут образовываться спонтанно или после расщепления модифицированных оснований, таких как 8-OH-Gua. [51] Генерация AP-сайта позволяет плавить дуплексную ДНК, чтобы демаскировать PQS, принимая [53] G-квадруплексная складка. С использованием полногеномного анализа ChIP-секвенирования , клеточных анализов и биохимических анализов in vitro была установлена связь между AP-сайтами, полученными из окисленных оснований ДНК, и образованием G-квадруплекса. [52]
Вклад окисления ДНК в болезни
Кроме того, концентрация 8-оксо-dG является известным биомаркером окислительного стресса внутри клетки, а чрезмерное количество окислительного стресса связано с канцерогенезом и другими заболеваниями. [58] При производстве 8-оксо-dG обладает способностью инактивировать OGG1, предотвращая тем самым восстановление повреждений ДНК, вызванных окислением гуанина. [52] Возможная инактивация позволяет невосстановленным повреждениям ДНК собираться в нереплицирующихся клетках, таких как мышцы, а также может вызывать старение. [57] Более того, окислительное повреждение ДНК, такое как 8-оксо-dG, способствует канцерогенезу посредством модуляции экспрессии генов или индукции мутаций. [57] При условии, что 8-oxo-dG восстанавливается с помощью BER, части репарационного белка остаются, что может привести к эпигенетическим изменениям или модуляции экспрессии генов. [59] При вставке 8-oxo-dG в ген тимидинкиназы человека было установлено, что если 8-oxo-dG оставить неконтролируемым и не восстановить с помощью BER, это может привести к частым мутациям и, в конечном итоге, к канцерогенезу. [52] [53]
Роль APE1 в регуляции генов
[ редактировать ]AP-эндонуклеаза 1 (APE1) — фермент, ответственный за продвижение и формирование структур G-квадруплекса. APE1 в основном отвечает за устранение повреждений, нанесенных сайтам AP через путь BER. APE1 считается очень важным, поскольку известно, что повреждение AP-сайта является наиболее повторяющимся типом эндогенного повреждения ДНК. [59] Окисление некоторых пуриновых оснований, таких как гуанин, приводит к образованию окисленных нуклеотидов, которые нарушают функцию ДНК из-за несовпадения нуклеотидов в последовательностях. [57] Это чаще встречается в последовательностях PQS, которые образуют окисленные структуры, такие как 8-оксогуанин . Как только клетка узнает об окислительном стрессе и повреждении, она привлекает к этому сайту OGG1 , основная функция которого — инициировать путь BER. [52] OGG1 делает это, расщепляя окисленное основание и создавая таким образом AP-сайт, в первую очередь за счет процесса отрицательной сверхспиральности. [54] Этот сайт AP затем сигнализирует клеткам о необходимости задействования связывания APE1, который связывается с открытой дуплексной областью. [58] Связывание APE1 затем играет важную роль, стабилизируя образование структур G-квадруплекса в этой области. Это способствует образованию структур G-квадруплекса путем складывания подставки. [60] В результате этого процесса образования петель четыре основания оказываются в непосредственной близости, которые будут удерживаться вместе с помощью пары оснований Хугстина. После этой стадии APE1 ацетилируется несколькими остатками лизина на хроматине, образуя ацетилированный APE1 (AcAPE1). [60] AcAPE1 очень важен для пути BER, поскольку он действует как коактиватор транскрипции или корепрессор, загружая факторы транскрипции (TF) в место повреждения, позволяя ему регулировать экспрессию генов. [61] AcAPE1 также очень важен, поскольку он позволяет APE1 связываться в течение более длительных периодов времени за счет задержки его диссоциации от последовательности, что позволяет процессу репарации быть более эффективным. [62] Деацетилирование AcAPE1 является движущей силой загрузки этих ТФ, при которой APE1 диссоциирует от структур G-квадруплекса. [63] Когда исследование выявило снижение присутствия APE1 и AcAPE1 в клетке, образование G-квадруплексных структур ингибировалось, что доказывает важность APE1 для формирования этих структур. Однако не все структуры G-квадруплексов нуждаются в APE1 для формирования, фактически некоторые из них образовывали более крупные структуры G-квадруплексов в его отсутствие. [52] Таким образом, мы можем заключить, что APE1 выполняет две важные роли в регуляции генома: стабилизирует образование структур g-квадруплекса и загрузку транскрипционных факторов на сайт AP.
Рак
[ редактировать ]Теломеры
[ редактировать ]Последовательности, образующие G-квадруплексы, преобладают в эукариотических клетках, особенно в теломерах, 5'-нетранслируемых цепях и горячих точках транслокации. G-квадруплексы могут ингибировать нормальную функцию клеток, а в здоровых клетках легко и быстро раскручиваются хеликазой . Однако в раковых клетках с мутировавшей хеликазой эти комплексы не могут быть раскручены, что приводит к потенциальному повреждению клетки. Это вызывает репликацию поврежденных и раковых клеток. Что касается терапевтических достижений, стабилизация G-квадруплексов раковых клеток может ингибировать рост и репликацию клеток, что приводит к их гибели . [64]
Промоутерские регионы
[ редактировать ]Наряду с ассоциацией G-квадруплексов в теломерных областях ДНК, структуры G-квадруплексов были идентифицированы в различных областях промотора протоонкогена человека . Структуры, наиболее присутствующие в промоторных областях этих онкогенов, обычно представляют собой структуры G-квадруплексной ДНК с параллельными цепями. [65] Некоторые из этих онкогенов включают c-KIT, PDGF-A, c-Myc и VEGF, что показывает важность этой вторичной структуры в росте и развитии рака. Хотя формирование структуры G-квадруплекса в некоторой степени различается для разных промоторных областей онкогенов, при развитии рака была обнаружена последовательная стабилизация этих структур. [66] Текущие терапевтические исследования активно фокусируются на стабилизации структур G-квадруплексов для остановки нерегулируемого роста и деления клеток.
Одна конкретная область гена, путь c-myc, играет важную роль в регуляции белкового продукта c-Myc. С помощью этого продукта белок c-Myc участвует в процессах апоптоза и роста или развития клеток, а также осуществляет контроль транскрипции обратной транскриптазы теломеразы человека . [67] В 2009 году было показано, что взаимодействие G-квадруплекса промотора c-Myc с NM23H2 регулирует c-Myc в раковых клетках. [42]
Регуляция c-myc посредством обратной транскриптазы теломеразы человека (hTERT) также напрямую регулируется через промотор G-квадруплекса путем взаимодействия с фактором транскрипции NM23H2, где эпигенетические модификации зависели от ассоциации NM23H2-G-квадруплекса. [44] Недавно сообщалось, что эпигенетическая регуляция hTERT опосредована взаимодействием G-квадруплекса промотора hTERT с теломерным фактором TRF2. [68]
Другой генный путь связан с геном VEGF, фактором роста эндотелия сосудов, который продолжает участвовать в процессе ангиогенеза или образования новых кровеносных сосудов. Формирование внутримолекулярной структуры G-квадруплекса показано при исследовании полипуринового тракта промоторной области гена VEGF. Благодаря недавним исследованиям роли функции G-квадруплекса in vivo было показано, что стабилизация структур G-квадруплекса регулирует транскрипцию гена VEGF с ингибированием факторов транскрипции на этом пути. Внутримолекулярные структуры G-квадруплекса образуются в основном за счет обильной последовательности гуанина в промоторной области этого специфического пути. [69] Ген циклин-зависимого ингибитора киназы контрольной точки клеточного цикла-1 CDKN1A (также известного как p21) содержит промотор G-квадруплекса. Взаимодействие этого G-квадруплекса с TRF2 (также известным как TERF2) привело к эпигенетической регуляции p21, что было протестировано с использованием G-квадруплекс-связывающего лиганда 360A. [70]
Фактор 1ɑ, индуцируемый гипоксией, HIF-1ɑ, продолжает участвовать в передаче сигналов рака посредством его связывания с элементом ответа на гипоксию, HRE, в присутствии гипоксии, чтобы начать процесс ангиогенеза . Благодаря недавним исследованиям этого специфического генного пути полипуриновая и полипиримидиновая области позволяют осуществлять транскрипцию этого специфического гена и формирование внутримолекулярной структуры G-квадруплекса. Однако необходимы дополнительные исследования, чтобы определить, регулирует ли образование G-квадруплекса экспрессию этого гена положительным или отрицательным образом. [71]
Онкоген c-kit связан с путем, кодирующим RTK, уровень экспрессии которого, как было показано, повышен при некоторых типах рака. Богатая гуаниновая последовательность этой промоторной области продемонстрировала способность образовывать различные квадруплексы. Текущие исследования этого пути сосредоточены на обнаружении биологической функции этого специфического образования квадруплекса на пути c-kit, хотя эта квадруплексная последовательность наблюдалась у различных видов. [36]
Онкоген RET участвует в транскрипции киназы , которая широко распространена при некоторых типах рака. Богатая гуанином последовательность в промоторной области этого пути указывает на необходимость базовой транскрипции тирозинкиназы этого рецептора. При некоторых типах рака уровень экспрессии белка RET повышен. Исследование этого пути позволило предположить образование G-квадруплекса в области промотора и сделать его подходящей мишенью для терапевтического лечения. [72]
Другой путь онкогена, включающий PDGF-A, фактор роста тромбоцитов, включает процесс заживления ран и действует как митогенный фактор роста клеток. Высокие уровни экспрессии PDGF связаны с повышенным ростом клеток и раком. Присутствие богатой гуанином последовательности в промоторной области PDGF-A демонстрирует способность образовывать внутримолекулярные параллельные структуры G-квадруплекса и, по-прежнему, предположительно играет роль в регуляции транскрипции PDGF-A. Однако исследования также выявили наличие структур G-квадруплекса в этой области благодаря взаимодействию TMPyP4 с этой последовательностью промотора. [73]
Терапия
[ редактировать ]Теломеры обычно состоят из G-квадруплексов и остаются важным объектом терапевтических исследований и открытий. Эти комплексы обладают высоким сродством к порфириновым кольцам , что делает их эффективными противораковыми средствами. Однако использование TMPyP4 было ограничено из-за его неселективности по отношению к теломерам раковых клеток и нормальной двухцепочечной ДНК (дцДНК). Чтобы решить эту проблему, был синтезирован аналог TMPyP4, известный как 5Me, который нацелен только на G-квадруплексную ДНК, которая подавляет рост рака более эффективно, чем TMPyP4. [74]
Проектирование и разработка лигандов остается важной областью исследований терапевтических реагентов из-за обилия G-квадруплексов и их многочисленных конформационных различий. Один тип лиганда, включающий производное хиндолина, SYUIQ-05, использует стабилизацию G-квадруплексов в промоторных областях для ингибирования продукции как белкового продукта c-Myc, так и обратной транскриптазы теломеразы человека (hTERT). Этот основной путь воздействия на эту область приводит к отсутствию удлинения теломеразы, что приводит к остановке развития клеток. Дальнейшие исследования по-прежнему необходимы для открытия единственного гена-мишени, позволяющего минимизировать нежелательную реактивность и обеспечить более эффективную противоопухолевую активность. [67]
Лиганды, связывающие квадруплексы
[ редактировать ]Одним из способов индуцирования или стабилизации образования G-квадруплекса является введение молекулы, которая может связываться со структурой G-квадруплекса. Ряд лигандов , которые могут быть как небольшими молекулами, так и белками , могут связываться с G-квадруплексом. Эти лиганды могут быть природными или синтетическими. Это становится все более обширной областью исследований в области генетики, биохимии и фармакологии.
катионные порфирины Было показано, что интеркалативно связываются с G-квадруплексами, а также с молекулой теломестатина .
Связывание лигандов с G-квадруплексами жизненно важно для борьбы с раком, поскольку G-квадруплексы обычно обнаруживаются в горячих точках транслокации. MM41, лиганд, который избирательно связывается с квадруплексом на промоторе BCL-2 , имеет форму центрального ядра и 4 боковых цепей, стерически разветвляющихся наружу. Форма лиганда имеет жизненно важное значение, поскольку она точно соответствует квадруплексу, который состоит из сложенных друг на друга квартетов и петель нуклеиновых кислот, удерживающих его вместе. При связывании центральный хромофор MM41 располагается поверх 3'-концевого G-квартета, а боковые цепи лиганда присоединяются к петлям квадруплекса. Квартет и хромофор связаны π-π-связью , тогда как боковые цепи и петли не связаны, а находятся в непосредственной близости. Что делает это связывание прочным, так это подвижность положения петель, позволяющая лучше связываться с боковыми цепями лиганда. [75]
TMPyP4, катионный порфирин, является более известным лигандом, связывающим G4, который помогает подавлять c-Myc. Способ связывания TMPyP4 с G4 аналогичен способу связывания MM41: кольцо укладывается на внешний G-квартет, а боковые цепи связаны с петлями G4. [76]
При разработке лигандов для связывания с G-квадруплексами лиганды обладают более высоким сродством к параллельно свернутым G-квадруплексам. Было обнаружено, что лиганды с меньшими боковыми цепями лучше связываются с квадруплексом, поскольку лиганды меньшего размера имеют более концентрированную электронную плотность . Кроме того, водородные связи лигандов с меньшими боковыми цепями короче и, следовательно, прочнее. Лиганды с подвижными боковыми цепями, которые способны вращаться вокруг своего центрального хромофора, сильнее связываются с G-квадруплексами, поскольку конформация петель G4 и боковых цепей лиганда может выравниваться. [77]
Методы квадруплексного прогнозирования
[ редактировать ]Идентификация и прогнозирование последовательностей, способных образовывать квадруплексы, является важным инструментом для дальнейшего понимания их роли. Обычно для поиска возможных последовательностей, образующих внутрицепочечный квадруплекс, используется простое сопоставление с образцом: d(G 3+ N 1-7 G 3+ N 1-7 G 3+ N 1-7 G 3+ ), где N - любой нуклеотид. основание (включая гуанин ). [78] Это правило широко используется в онлайн- алгоритмах . Хотя это правило эффективно идентифицирует места образования G-квадруплексов, оно также определяет подмножество несовершенных гомопуриновых зеркальных повторов, способных к образованию триплекса. [79] и образование i-мотива C-цепи. [80] Более того, эти последовательности также обладают способностью образовывать структуры со скольжением и фолдбэком, которые являются неявными промежуточными звеньями в формировании как квадруплексов, так и фолдбеков. [4] и триплекс ДНК [81] структуры. В одном исследовании [82] было обнаружено, что наблюдаемое количество этих мотивов на пару оснований (т.е. частота) быстро увеличивается у эуметазоа , для которых доступны полные геномные последовательности. Это предполагает, что последовательности могут находиться под положительным отбором, обусловленным эволюцией систем, способных подавлять образование структур, отличных от B.
Совсем недавно были разработаны усовершенствованные веб-инструменты для идентификации последовательностей формирования G-квадруплексов, включая удобную для пользователя версию G4Hunter с открытым доступом, основанную на подходе скользящего окна. [83] или G4RNA Screener на основе алгоритма машинного обучения. [84]
Методы исследования G-квадруплексов
[ редактировать ]Для идентификации G-квадруплексов разработан ряд экспериментальных методов. Эти методы можно разделить на два класса: биофизические и биохимические методы. [85]
Биохимические методы
[ редактировать ]Биохимические методы были использованы для изучения образования G-квадруплексов в контексте более длинных последовательностей. В анализе остановки ДНК-полимеразы образование G-квадруплекса в матрице ДНК может действовать как препятствие и вызывать остановку полимеразы, что останавливает расширение праймера. [86] Диметилсульфат (ДМС) с последующим анализом расщепления пиперидина основан на том факте, что образование G-квадруплекса препятствует метилированию гуанина N7, вызванному ДМС, что приводит к защитному паттерну, наблюдаемому в области G-квадруплекса ДНК после пиперидина. расщепление. [87]
Биофизические методы
[ редактировать ]Топологию структуры G-квадруплекса можно определить путем мониторинга сигналов положительного или отрицательного кругового дихроизма (CD) на определенных длинах волн. [88] Параллельные G-квадруплексы имеют отрицательные и положительные сигналы CD при 240 и 262 нм соответственно, тогда как антипараллельные G-квадруплексы размещают эти сигналы при 262 и 295 нм соответственно. Чтобы проверить образование G-квадруплекса, следует также провести эксперименты по КД в условиях, стабилизирующих не-G-квадруплекс (Li+) и стабилизирующих G-квадруплекс (таких как K+ или с лигандами G-квадруплекса), и сканировать в сторону дальней УФ-области. (180–230 нм). Аналогично, термостабильность структуры G-квадруплекса можно определить, наблюдая УФ-сигнал при длине волны 295 нм. [89] При плавлении G-квадруплекса УФ-поглощение при 295 нм уменьшается, что приводит к гипохромному сдвигу, который является отличительной чертой структуры G-квадруплекса. Другой подход к обнаружению G-квадруплексов включает методы на основе нанопор . Во-первых, было показано, что биологические нанопоры могут обнаруживать G-квадруплексы на основе исключения размера и специфического взаимодействия G-квадруплекса и белковой нанополости. [90] Новый подход сочетает в себе твердотельные нанопоры и нанотехнологию ДНК для обнаружения G-квадруплексов без меток, их картирования на дцДНК и мониторинга образования G-квадруплексов. [91]
Роль в неврологических расстройствах
[ редактировать ]G-квадруплексы участвуют в неврологических расстройствах посредством двух основных механизмов. Первый заключается в расширении G-повторов внутри генов, что приводит к образованию структур G-квадруплексов, непосредственно вызывающих заболевание, как в случае с геном C9orf72 и боковым амиотрофическим склерозом (БАС) или лобно-височной деменцией (ЛВД). Второй механизм заключается в мутациях, которые влияют на экспрессию белков, связывающих G-квадруплекс, как видно из гена ломкой Х-гена умственной отсталости 1 (FMR1) и синдрома ломкой Х-хромосомы . [92]
Ген C9orf72 кодирует белок C9orf72 , который обнаруживается по всему мозгу в нейронов цитоплазме и на пресинаптических окончаниях. [93] Мутации гена C9orf72 связаны с развитием ЛВД и БАС. [94] Эти два заболевания имеют причинную связь с повторами GGGGCC (G 4 C 2 ) в 1-м интроне гена C9orf72. Нормальные люди обычно имеют от 2 до 8 повторов G 4 C 2 , но люди с ЛВД или БАС имеют от 500 до нескольких тысяч повторов G 4 C 2 . [95] [96] Было показано, что транскрибируемая РНК этих повторов образует стабильные G-квадруплексы, причем данные показывают, что повторы G 4 C 2 в ДНК также обладают способностью образовывать смешанные параллельно-антипараллельные структуры G-квадруплексов. [97] [98] Было показано, что эти РНК-транскрипты, содержащие повторы G 4 C 2 , связывают и разделяют широкий спектр белков, включая нуклеолин . Нуклеолин участвует в синтезе и созревании рибосом внутри ядра, а разделение нуклеолина мутировавшими транскриптами РНК нарушает функцию ядрышка и синтез рибосомальной РНК. [99]
Белок умственной отсталости Fragile X (FMRP) представляет собой широко экспрессируемый белок, кодируемый геном FMR1, который связывается с вторичными структурами G-квадруплексов в нейронах и участвует в синаптической пластичности . [100] FMRP действует как негативный регулятор трансляции, и его связывание стабилизирует структуры G-квадруплексов в транскриптах мРНК, ингибируя рибосомальное удлинение нейрона мРНК в дендритах и контролируя время экспрессии транскрипта. [101] [102] Мутации этого гена могут вызвать развитие синдрома ломкой Х-хромосомы, аутизма и других неврологических расстройств. [103] В частности, синдром ломкой Х-хромосомы вызван увеличением количества повторов CGG с 50 до более 200 в экзоне 13 гена FMR1. Это расширение повторов способствует метилированию ДНК и другим эпигенетическим модификациям гетерохроматина FMR1, которые предотвращают транскрипцию гена, что приводит к патологически низким уровням FMRP. [104] [105]
Терапевтические подходы
[ редактировать ]Антисмысловые вмешательства и низкомолекулярные лиганды являются распространенными стратегиями, используемыми для борьбы с неврологическими заболеваниями, связанными с повторами расширения G-квадруплекса. Следовательно, эти методы особенно выгодны для борьбы с неврологическими заболеваниями, которые имеют механизм усиления функции, то есть когда измененный генный продукт имеет новую функцию или новую экспрессию гена; это было обнаружено в C9orf72 (открытая рамка считывания хромосомы 9 72). [106]
Антисмысловая терапия — это процесс, при котором синтезированные цепи нуклеиновых кислот используются для прямого и специфического связывания с мРНК, продуцируемой определенным геном, что инактивирует ее. Антисмысловые олигонуклеотиды (ASO) обычно используются для нацеливания на РНК C9orf72 области повторов расширения GGGGCC G-квадруплекса, что снижает токсичность в клеточных моделях C9orf72. [107] [108] [109] ASO ранее использовались для восстановления нормальных фенотипов при других неврологических заболеваниях, которые имеют механизмы усиления функции, с той лишь разницей, что они использовались при отсутствии повторяющихся областей расширения G-квадруплекса. [110] [111] [112] [113]
Стратегия-приманка G-квадруплекса — еще один многообещающий подход к нацеливанию на раковые клетки за счет использования уникальных структурных особенностей G-квадруплекса. Стратегия предполагает создание синтетических олигонуклеотидов , которые имитируют структуру G-квадруплексов и конкурируют с эндогенными G-квадруплексами за связывание с факторами транскрипции. Эти ловушки обычно состоят из G-богатой последовательности, которая может образовывать стабильную структуру G-квадруплекса, и короткой линкерной области, которую можно модифицировать для оптимизации их свойств. [114] При введении в раковые клетки приманка может перехватывать связанные факторы транскрипции и связывать их, что приводит к регуляции экспрессии генов. Было успешно продемонстрировано, что приманки ингибируют онкогенный KRAS у мышей SCID, что приводит к снижению роста опухоли и увеличению медианного времени выживаемости. [115]
Другим широко используемым методом является использование низкомолекулярных лигандов . Их можно использовать для воздействия на области G-квадруплексов, которые вызывают неврологические расстройства. Существует около 1000 различных G-квадруплексных лигандов, с которыми они способны взаимодействовать через свои ароматические кольца ; это позволяет низкомолекулярным лигандам штабелироваться на плоских концевых тетрадах внутри областей G-квадруплекса. Недостатком использования низкомолекулярных лигандов в качестве терапевтического метода является то, что специфичностью трудно управлять из-за вариабельности G-квадруплексов в их первичных последовательностях, ориентации, термодинамической стабильности и стехиометрии цепи нуклеиновой кислоты. На данный момент, [ когда? ] ни один низкомолекулярный лиганд не смог быть совершенно специфичным для одной последовательности G-квадруплекса. [116] [117] Однако катионный порфирин, известный как TMPyP4, способен связываться с областью повторов C9orf72 GGGGCC, что приводит к разворачиванию области повторов G-квадруплекса и потере взаимодействия с белками, в результате чего она теряет свою функциональность. [118] Низкомолекулярные лиганды, состоящие в основном из свинца, также могут нацеливаться на области повторов GGGGCC и в конечном итоге уменьшать как связанную с повторами не-ATG-трансляцию, так и фокусы РНК в нейронных клетках, полученных от пациентов с боковым амиотрофическим склерозом (БАС). Это доказывает, что низкомолекулярные лиганды являются эффективным и действенным процессом воздействия на регионы GGGGCC и что специфичность связывания низкомолекулярных лигандов является достижимой целью для научного сообщества.
Металлокомплексы обладают рядом особенностей, которые делают их особенно подходящими в качестве связующих ДНК G4 и, следовательно, в качестве потенциальных лекарств. Хотя металл играет в основном структурную роль в большинстве связующих G4, есть также примеры, когда он напрямую взаимодействует с G4 посредством электростатических взаимодействий или прямой координации с азотистыми основаниями. [119]
Ссылки
[ редактировать ]- ^ Капра Дж.А., Паешке К., Сингх М., Закян В.А. (июль 2010 г.). «Последовательности ДНК G-квадруплекса эволюционно консервативны и связаны с различными геномными особенностями Saccharomyces cerevisiae» . PLOS Вычислительная биология . 6 (7): e1000861. Бибкод : 2010PLSCB...6E0861C . дои : 10.1371/journal.pcbi.1000861 . ПМЦ 2908698 . ПМИД 20676380 .
- ^ Рут Э.Д., Криси С.Д., Бирбауэр П.Е., Акман С.А., Вон Дж.П., Смальдино П.Дж. (март 2017 г.). «Подход G-квадруплексного ДНК-сродства для очистки ферментативно активной резольвазы G4» . Журнал визуализированных экспериментов . 121 (121). дои : 10.3791/55496 . ПМК 5409278 . ПМИД 28362374 .
- ^ Jump up to: а б Ларджи Э., Мергни Дж., Габелика В. (2016). «Глава 7. Роль ионов щелочных металлов в структуре и стабильности G-квадруплексной нуклеиновой кислоты». В Астрид С., Хельмут С., Роланд КО (ред.). Ионы щелочных металлов: их роль в жизни (PDF) . Ионы металлов в науках о жизни. Том. 16. Спрингер. стр. 203–258. дои : 10.1007/978-3-319-21756-7_7 . ISBN 978-3-319-21755-0 . ПМИД 26860303 .
- ^ Jump up to: а б Сандквист, Висконсин, Клуг А. (декабрь 1989 г.). «Теломерная ДНК димеризуется за счет образования гуаниновых тетрад между шпильками». Природа . 342 (6251): 825–9. Бибкод : 1989Natur.342..825S . дои : 10.1038/342825a0 . ПМИД 2601741 . S2CID 4357161 .
- ^ Jump up to: а б с Сен Д., Гилберт В. (июль 1988 г.). «Образование параллельных четырехцепочечных комплексов с помощью богатых гуанином мотивов в ДНК и его значение для мейоза». Природа . 334 (6180): 364–6. Бибкод : 1988Natur.334..364S . дои : 10.1038/334364a0 . ПМИД 3393228 . S2CID 4351855 .
- ^ Jump up to: а б с д Равал П., Куммарасетти В.Б., Равиндран Р., Кумар Н., Хальдер К., Шарма Р., Мукерджи М., Дас С.К., Чоудхури С. (2006). «Пологеномное предсказание ДНК G4 как регуляторных мотивов: роль в глобальной регуляции Escherichia Coli» . Геномные исследования . 16 (5): 644–655. дои : 10.1101/гр.4508806 . ПМЦ 1457047 . ПМИД 16651665 .
- ^ Jump up to: а б с д Борман С. (28 мая 2007 г.). «Восхождение структур нуклеиновых кислот квадруплексов становится перспективными мишенями для лекарств». Новости химии и техники . 85 (22): 12–17. doi : 10.1021/cen-v085n009.p012a .
- ^ Верма А., Хальдер К., Хальдер Р., Ядав В.К., Равал П., Тхакур Р.К., Мохд Ф., Шарма А., Чоудхури С. (2008). «Полногеномный компьютерный анализ и анализ экспрессии выявили мотивы G-квадруплексной ДНК как консервативные цис-регуляторные элементы у человека и родственных ему видов». Журнал медицинской химии . 51 (18): 5641–5649. дои : 10.1021/jm800448a . ПМИД 18767830 .
- ^ Хан Х., Херли Л.Х. (апрель 2000 г.). «ДНК G-квадруплекса: потенциальная мишень для разработки противораковых лекарств». Тенденции в фармакологических науках . 21 (4): 136–42. дои : 10.1016/s0165-6147(00)01457-7 . ПМИД 10740289 .
- ^ Бохман М.Л., Паешке К., Закян В.А. (ноябрь 2012 г.). «Вторичные структуры ДНК: стабильность и функция G-квадруплексных структур» . Обзоры природы. Генетика . 13 (11): 770–80. дои : 10.1038/nrg3296 . ПМЦ 3725559 . ПМИД 23032257 .
- ^ Ядав В.К., Авраам Дж.К., Мани П., Кулшреста Р., Чоудхури С. (2008). «QuadBase: Полногеномная база данных ДНК G4 - возникновение и сохранение у промоутеров человека, шимпанзе, мышей и крыс и 146 микробов» . Исследования нуклеиновых кислот . 36 (База данных): D381–D385. дои : 10.1093/nar/gkm781 . ПМК 2238983 . ПМИД 17962308 .
- ^ Дапола П., Чоудхури С. (июль 2016 г.). «QuadBase2: веб-сервер для мультиплексного квадруплексного майнинга и визуализации гуанина» . Исследования нуклеиновых кислот . 44 (П1): W277–W283. дои : 10.1093/nar/gkw425 . ПМЦ 4987949 . ПМИД 27185890 .
- ^ Роудс Д., Липпс Х.Дж. (октябрь 2015 г.). «G-квадруплексы и их регуляторная роль в биологии» . Исследования нуклеиновых кислот . 43 (18): 8627–37. дои : 10.1093/nar/gkv862 . ПМК 4605312 . ПМИД 26350216 .
- ^ Борман С. (ноябрь 2009 г.). «Промоторные квадруплексы, сложенные структуры ДНК в сайтах активации генов, могут быть полезными мишенями для лечения рака». Новости химии и техники . 87 (44): 28–30. doi : 10.1021/cen-v087n044.p028 .
- ^ Геллерт М., Липсетт М.Н., Дэвис Д.Р. (декабрь 1962 г.). «Образование спирали гуаниловой кислотой» . Труды Национальной академии наук Соединенных Штатов Америки . 48 (12): 2013–8. Бибкод : 1962ПНАС...48.2013Г . дои : 10.1073/pnas.48.12.2013 . ПМК 221115 . ПМИД 13947099 .
- ^ Хендерсон Э., Хардин CC, Уок С.К., Тиноко I, Блэкберн Э.Х. (декабрь 1987 г.). «Теломерные ДНК-олигонуклеотиды образуют новые внутримолекулярные структуры, содержащие пары оснований гуанин-гуанин». Клетка . 51 (6): 899–908. дои : 10.1016/0092-8674(87)90577-0 . ПМИД 3690664 . S2CID 37343642 .
- ^ Мюллер, Себастьян; Кумари, Сунита; Родригес, Рафаэль; Баласубраманян, Шанкар (10 октября 2010 г.). «Низкомолекулярное выделение G-квадруплексов из клеток человека» . Природная химия . 2 (12): 1095–1098. Бибкод : 2010НатЧ...2.1095М . дои : 10.1038/nchem.842 . ПМК 3119466 . ПМИД 21107376 .
- ^ Родригес, Рафаэль; Миллер, Кайл М; Формент, Хосеп V; Брэдшоу, Чарльз Р.; Никан, Мехран; Бриттон, Себастьен; Эльшлегель, Тобиас; Кшемальче, Блерта; Баласубраманиан, Шанкар; Джексон, Стивен П. (5 февраля 2012 г.). «Повреждение ДНК, вызванное малыми молекулами, идентифицирует альтернативные структуры ДНК в генах человека» . Химическая биология природы . 8 (3): 301–310. дои : 10.1038/nchembio.780 . ПМЦ 3433707 . ПМИД 22306580 .
- ^ Симонссон Т. (апрель 2001 г.). «G-квадруплексные структуры ДНК — вариации на тему». Биологическая химия . 382 (4): 621–8. дои : 10.1515/BC.2001.073 . ПМИД 11405224 . S2CID 43536134 .
- ^ Бердж С., Паркинсон Г.Н., Хейзел П., Тодд А.К., Нидл С. (2006). «Квадруплексная ДНК: последовательность, топология и структура» . Исследования нуклеиновых кислот . 34 (19): 5402–15. дои : 10.1093/нар/gkl655 . ПМЦ 1636468 . ПМИД 17012276 .
- ^ Цао К., Рывкин П., Джонсон Ф.Б. (май 2012 г.). «Вычислительное обнаружение и анализ последовательностей с потенциалом образования межцепочечных G-квадруплексов дуплексного происхождения» . Методы . 57 (1): 3–10. дои : 10.1016/j.ymeth.2012.05.002 . ПМК 3701776 . ПМИД 22652626 .
- ^ Кудлицкий А.С. (2016). «G-квадруплексы, включающие обе цепи геномной ДНК, широко распространены и колокализуются с функциональными участками генома человека» . ПЛОС ОДИН . 11 (1): e0146174. Бибкод : 2016PLoSO..1146174K . дои : 10.1371/journal.pone.0146174 . ПМЦ 4699641 . ПМИД 26727593 .
- ^ Мурат П., Баласубраманян С. (апрель 2014 г.). «Существование и последствия G-квадруплексных структур в ДНК» . Текущее мнение в области генетики и развития . 25 (25): 22–9. дои : 10.1016/j.gde.2013.10.012 . ПМИД 24584093 .
- ^ Миёси Д., Каримата Х., Сугимото Н. (июнь 2006 г.). «Гидратация регулирует термодинамику образования G-квадруплекса в условиях молекулярной скученности». Журнал Американского химического общества . 128 (24): 7957–63. дои : 10.1021/ja061267m . ПМИД 16771510 .
- ^ Чжэн К.В., Чен З., Хао Ю.Х., Тан З. (январь 2010 г.). «Молекулярная скученность создает необходимую среду для образования стабильных G-квадруплексов в длинной двухцепочечной ДНК» . Исследования нуклеиновых кислот . 38 (1): 327–38. дои : 10.1093/nar/gkp898 . ПМК 2800236 . ПМИД 19858105 .
- ^ Эндо Т., Роде А.Б., Такахаши С., Катаока Ю., Кувахара М., Сугимото Н. (февраль 2016 г.). «Мониторинг образования G-квадруплексов в реальном времени во время транскрипции» . Аналитическая химия . 88 (4): 1984–9. дои : 10.1021/acs.analchem.5b04396 . ПМИД 26810457 .
- ^ Ван Ц, Лю JQ, Чэнь Z, Чжэн К.В., Чэнь С.И., Хао Ю.Х., Тан З. (август 2011 г.). «Образование G-квадруплекса на 3'-конце теломерной ДНК ингибирует ее удлинение теломеразой, полимеразой и раскручивание геликазой» . Исследования нуклеиновых кислот . 39 (14): 6229–37. дои : 10.1093/nar/gkr164 . ПМЦ 3152327 . ПМИД 21441540 .
- ^ Шаффицель С., Бергер И., Постберг Дж., Ханес Дж., Липпс Х.Дж., Плюктхун А. (июль 2001 г.). «Сгенерированные in vitro антитела, специфичные к теломерной гуанин-квадруплексной ДНК, реагируют с макронуклеусами Stylonychia lemnae» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (15): 8572–7. Бибкод : 2001PNAS...98.8572S . дои : 10.1073/pnas.141229498 . ПМЦ 37477 . ПМИД 11438689 .
- ^ Паешке К., Симонссон Т., Постберг Дж., Роудс Д., Липпс Х.Дж. (октябрь 2005 г.). «Белки, связывающие концы теломер, контролируют образование структур ДНК G-квадруплекса in vivo». Структурная и молекулярная биология природы . 12 (10): 847–54. дои : 10.1038/nsmb982 . ПМИД 16142245 . S2CID 6079323 .
- ^ Кар А., Джонс Н., Арат Н.О., Фишел Р., Гриффит Дж.Д. (июнь 2018 г.). «Длинно повторяющаяся (TTAGGG) n одноцепочечная ДНК самоконденсируется в компактные бисерные нити, стабилизированные образованием G-квадруплекса» . Журнал биологической химии . 293 (24): 9473–9485. дои : 10.1074/jbc.RA118.002158 . ПМК 6005428 . ПМИД 29674319 .
- ^ Волна А, Бартас М, Карлицкий В, Незваль Дж, Кундратова К, Печинка П и др. (июль 2021 г.). «G-квадруплекс в гене, кодирующем большую субъединицу растительной РНК-полимеразы II: история миллиардной давности» . Международный журнал молекулярных наук . 22 (14): 7381. doi : 10.3390/ijms22147381 . ПМЦ 8306923 . ПМИД 34299001 .
- ^ Симонссон Т., Печинка П., Кубиста М. (март 1998 г.). «Образование тетраплекса ДНК в контрольной области c-myc» . Исследования нуклеиновых кислот . 26 (5): 1167–72. дои : 10.1093/нар/26.5.1167 . ПМК 147388 . ПМИД 9469822 .
- ^ Сиддики-Джейн А., Grand CL, Bearss DJ, Херли Л.Х. (сентябрь 2002 г.). «Прямое доказательство наличия G-квадруплекса в промоторной области и его нацеливания с помощью небольшой молекулы для подавления транскрипции c-MYC» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (18): 11593–8. Бибкод : 2002PNAS...9911593S . дои : 10.1073/pnas.182256799 . ПМК 129314 . ПМИД 12195017 .
- ^ Юпперт Дж.Л., Баласубраманян С. (14 декабря 2006 г.). «G-квадруплексы в промоторах по всему геному человека» . Исследования нуклеиновых кислот . 35 (2): 406–13. дои : 10.1093/нар/gkl1057 . ПМК 1802602 . ПМИД 17169996 .
- ^ Дай Дж., Дексхаймер Т.С., Чен Д., Карвер М., Амбрус А., Джонс Р.А., Ян Д. (февраль 2006 г.). «Внутримолекулярная структура G-квадруплекса со смешанными параллельными/антипараллельными G-нитями, образующимися в области промотора BCL-2 человека в растворе» . Журнал Американского химического общества . 128 (4): 1096–8. дои : 10.1021/ja055636a . ПМК 2556172 . ПМИД 16433524 .
- ^ Jump up to: а б Фернандо Х., Решка А.П., Юпперт Дж., Ладам С., Ранкин С., Венкитараман А.Р., Нейдл С., Баласубраманиан С. (июнь 2006 г.). «Консервативный квадруплексный мотив, расположенный в сайте активации транскрипции онкогена c-kit человека» . Биохимия . 45 (25): 7854–60. дои : 10.1021/bi0601510 . ПМК 2195898 . ПМИД 16784237 .
- ^ Юпперт Дж.Л., Баласубраманян С. (2005). «Распространенность квадруплексов в геноме человека» . Исследования нуклеиновых кислот . 33 (9): 2908–16. дои : 10.1093/nar/gki609 . ПМК 1140081 . ПМИД 15914667 .
- ^ Равал П., Куммарасетти В.Б., Равиндран Дж., Кумар Н., Хальдер К., Шарма Р., Мукерджи М., Дас С.К., Чоудхури С. (май 2006 г.). «Пологеномное предсказание ДНК G4 как регуляторного мотива: роль в глобальной регуляции Escherichia coli» . Геномные исследования . 16 (5): 644–55. дои : 10.1101/гр.4508806 . ПМЦ 1457047 . ПМИД 16651665 .
- ^ Камат-Лёб А., Леб Л.А., Фрай М. (2012). Коттерилл С. (ред.). «Белок синдрома Вернера отличается от белка синдрома Блума своей способностью прочно связывать различные структуры ДНК» . ПЛОС ОДИН . 7 (1): e30189. Бибкод : 2012PLoSO...730189K . дои : 10.1371/journal.pone.0030189 . ПМК 3260238 . ПМИД 22272300 .
- ^ Вон Дж.П., Криси С.Д., Раут Э.Д., Джойнер-Батт С., Дженкинс Г.С., Паули С., Нагамин Ю., Акман С.А. (ноябрь 2005 г.). «Белковый продукт DEXH гена DHX36 является основным источником тетрамолекулярной квадруплексной активности разрешения G4-ДНК в лизатах клеток HeLa» . Журнал биологической химии . 280 (46): 38117–20. дои : 10.1074/jbc.C500348200 . ПМИД 16150737 .
- ^ Чен MC, Ферре-Д'Амаре, Арканзас (15 августа 2017 г.). «Структурная основа активности геликазы DEAH/RHA» . Кристаллы . 7 (8): 253. дои : 10.3390/cryst7080253 .
- ^ Jump up to: а б Тхакур Р.К., Кумар П., Хальдер К., Верма А., Кар А., Пэрент Дж.Л., Басундра Р., Кумар А., Чоудхури С. (январь 2009 г.). «Взаимодействие супрессора метастазов NM23-H2 с G-квадруплексной ДНК в гиперчувствительном элементе нуклеазы промотора c-MYC индуцирует экспрессию c-MYC» . Исследования нуклеиновых кислот . 37 (1): 172–183. дои : 10.1093/нар/gkn919 . ПМЦ 2615625 . ПМИД 19033359 .
- ^ Борман С. (ноябрь 2009 г.). «Промоторные квадруплексы. Свернутые структуры ДНК в сайтах активации генов могут быть полезными мишенями для лечения рака». Новости химии и техники . 87 (44): 28–30. doi : 10.1021/cen-v087n044.p028 .
- ^ Jump up to: а б с Саха Д., Сингх А., Хусейн Т., Шривастава В., Сенгупта С., Кар А., Дхапола П., Умманни Р., Чоудхури С. (июль 2017 г.). «Эпигенетическая супрессия теломеразы человека (hTERT) опосредована супрессором метастазов NME2 G-квадруплекс-зависимым способом» . Журнал биологической химии . 292 (37): 15205–15215. дои : 10.1074/jbc.M117.792077 . ПМК 5602382 . ПМИД 28717007 .
- ^ Мукерджи А.К., Шарма С., Багри С., Кутум Р., Кумар П., Хусейн А., Сингх П., Саха Д., Кар А., Даш Д., Чоудхури С. (ноябрь 2019 г.). «Фактор 2, связывающий теломерные повторы, активно связывается с внетеломерными G-квадруплексами и регулирует эпигенетический статус нескольких промоторов генов» . Журнал биологической химии . 294 (47): 17709–17722. дои : 10.1074/jbc.RA119.008687 . ПМЦ 6879327 . ПМИД 31575660 .
- ^ Майзелс Н., Грей LT (апрель 2013 г.). Розенберг С.М. (ред.). «Геном G4» . ПЛОС Генетика . 9 (4): e1003468. дои : 10.1371/journal.pgen.1003468 . ПМК 3630100 . ПМИД 23637633 .
- ^ Биффи Дж., Таннахилл Д., Маккафферти Дж., Баласубраманиан С. (март 2013 г.). «Количественная визуализация структур G-квадруплекса ДНК в клетках человека» . Природная химия . 5 (3): 182–6. Бибкод : 2013НатЧ...5..182Б . дои : 10.1038/nchem.1548 . ПМЦ 3622242 . ПМИД 23422559 .
- ^ Чен М.С., Типпана Р., Демешкина Н.А., Мурат П., Баласубраманян С., Мьонг С., Ферре-Д'Амаре А.Р. (июнь 2018 г.). «Структурная основа разворачивания G-квадруплекса хеликазой DEAH/RHA DHX36» . Природа . 558 (7710): 465–469. Бибкод : 2018Natur.558..465C . дои : 10.1038/s41586-018-0209-9 . ПМК 6261253 . ПМИД 29899445 .
- ^ Райс С., Скордалакес Э (2016). «Структура и функции теломерного комплекса CST» . Журнал вычислительной и структурной биотехнологии . 14 : 161–7. дои : 10.1016/j.csbj.2016.04.002 . ПМЦ 4872678 . ПМИД 27239262 .
- ^ Гензель-Херч Р., Беральди Д., Ленсинг С.В., Марсико Г., Зайнер К., Парри А. и др. (октябрь 2016 г.). «Структуры G-квадруплекса отмечают регуляторный хроматин человека» . Природная генетика . 48 (10): 1267–1272. дои : 10.1038/ng.3662 . ПМИД 27618450 . S2CID 20967177 .
- ^ Jump up to: а б Поетч А.Р. (2020). «AP-Seq: метод измерения апуриновых сайтов и аддуктов малых оснований по всему геному» . Ядро . Методы молекулярной биологии. Том. 2175. Клифтон, Нью-Джерси, стр. 95–108. дои : 10.1007/978-1-0716-0763-3_8 . ISBN 978-1-0716-0762-6 . ISSN 1940-6029 . ПМИД 32681486 . S2CID 220631202 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Jump up to: а б с д и ж г Ройчудри С., Праманик С., Харрис Х.Л., Тарпли М., Саркар А., Спаньол Г. и др. (май 2020 г.). «Эндогенные окисленные основания ДНК и APE1 регулируют образование G-квадруплексных структур в геноме» . Труды Национальной академии наук Соединенных Штатов Америки . 117 (21): 11409–11420. Бибкод : 2020PNAS..11711409R . дои : 10.1073/pnas.1912355117 . ПМК 7260947 . ПМИД 32404420 .
- ^ Jump up to: а б с д Канугови С., Шаманна Р.А., Крото Д.Л., Бор В.А. (июнь 2014 г.). «Базовые уровни репарации эксцизионной ДНК в митохондриальных лизатах болезни Альцгеймера» . Нейробиология старения . 35 (6): 1293–1300. doi : 10.1016/j.neurobiolaging.2014.01.004 . ПМК 5576885 . ПМИД 24485507 .
- ^ Jump up to: а б Сан Д., Херли Л.Х. (май 2009 г.). «Важность отрицательной сверхспиральности в индукции образования структур G-квадруплекса и i-мотива в промоторе c-Myc: значение для нацеливания лекарств и контроля экспрессии генов» . Журнал медицинской химии . 52 (9): 2863–2874. дои : 10.1021/jm900055s . ПМК 2757002 . ПМИД 19385599 .
- ^ Jump up to: а б Хилл Дж.В., Хазра Т.К., Изуми Т., Митра С. (январь 2001 г.). «Стимуляция 8-оксогуанин-ДНК-гликозилазы человека AP-эндонуклеазой: потенциальная координация начальных этапов эксцизионного восстановления оснований» . Исследования нуклеиновых кислот . 29 (2): 430–438. дои : 10.1093/нар/29.2.430 . ПМК 29662 . ПМИД 11139613 .
- ^ Берроуз С.Дж., Мюллер Дж.Г. (май 1998 г.). «Окислительные модификации нуклеиновых оснований, ведущие к разрыву цепи». Химические обзоры . 98 (3): 1109–1152. дои : 10.1021/cr960421s . ПМИД 11848927 .
- ^ Jump up to: а б с д Поетч АР (07.01.2020). «Геномика окислительного повреждения ДНК, репарации и последующего мутагенеза» . Журнал вычислительной и структурной биотехнологии . 18 : 207–219. дои : 10.1016/j.csbj.2019.12.013 . ПМК 6974700 . ПМИД 31993111 .
- ^ Jump up to: а б Флеминг AM, Берроуз CJ (октябрь 2017 г.). «Тандемные поражения 8-оксо-7,8-дигидро-2'-дезоксигуанозина и абазового сайта склонны к окислению с образованием гидантоиновых продуктов, которые сильно дестабилизируют дуплексную ДНК» . Органическая и биомолекулярная химия . 15 (39): 8341–8353. дои : 10.1039/C7OB02096A . ПМЦ 5636683 . ПМИД 28936535 .
- ^ Jump up to: а б Китсера Н., Родригес-Альварес М., Эммерт С., Карелл Т., Хобта А. (сентябрь 2019 г.). «Нуклеотидная эксцизионная репарация абазических повреждений ДНК» . Исследования нуклеиновых кислот . 47 (16): 8537–8547. дои : 10.1093/nar/gkz558 . ПМК 6895268 . ПМИД 31226203 .
- ^ Jump up to: а б Ройчудри С., Нат С., Сонг Х., Хегде М.Л., Беллот Л.Дж., Манта А.К. и др. (март 2017 г.). «Человеческая апуриновая/апиримидиновая эндонуклеаза (APE1) ацетилируется в местах повреждения ДНК в хроматине, и ацетилирование модулирует ее активность по восстановлению ДНК» . Молекулярная и клеточная биология . 37 (6). дои : 10.1128/mcb.00401-16 . ПМЦ 5335514 . ПМИД 27994014 .
- ^ Чаттопадьяй Р., Дас С., Маити А.К., Болдог И., Се Дж., Хазра Т.К. и др. (декабрь 2008 г.). «Регуляторная роль AP-эндонуклеазы человека (APE1/Ref-1) в YB-1-опосредованной активации гена множественной лекарственной устойчивости MDR1» . Молекулярная и клеточная биология . 28 (23): 7066–7080. дои : 10.1128/mcb.00244-08 . ПМК 2593380 . ПМИД 18809583 .
- ^ Бхакат К.К., Изуми Т., Ян Ш., Хазра Т.К., Митра С. (декабрь 2003 г.). «Роль ацетилированной AP-эндонуклеазы человека (APE1/Ref-1) в регуляции гена паратиреоидного гормона» . Журнал ЭМБО . 22 (23): 6299–6309. дои : 10.1093/emboj/cdg595 . ПМК 291836 . ПМИД 14633989 .
- ^ Ямамори Т., ДеРикко Дж., Накви А., Хоффман Т.А., Маттаджасингх И., Касуно К. и др. (январь 2010 г.). «SIRT1 деацетилирует APE1 и регулирует иссечение клеточного основания» . Исследования нуклеиновых кислот . 38 (3): 832–845. дои : 10.1093/нар/gkp1039 . ПМЦ 2817463 . ПМИД 19934257 .
- ^ Нидл С. (июль 2016 г.). «Квадруплексные нуклеиновые кислоты как новые терапевтические мишени» (PDF) . Журнал медицинской химии . 59 (13): 5987–6011. doi : 10.1021/acs.jmedchem.5b01835 . ПМИД 26840940 .
- ^ Чен Ю, Ян Д (сентябрь 2012 г.). Последовательность, стабильность и структура G-квадруплексов и их взаимодействие с лекарственными средствами . Том. Глава 17. С. 17.5.1–17.5.17. дои : 10.1002/0471142700.nc1705s50 . ISBN 978-0471142706 . ПМЦ 3463244 . ПМИД 22956454 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Брукс Т.А., Кендрик С., Херли Л. (сентябрь 2010 г.). «Понимание функций G-квадруплекса и i-мотива в промоторах онкогенов» . Журнал ФЭБС . 277 (17): 3459–69. дои : 10.1111/j.1742-4658.2010.07759.x . ПМК 2971675 . ПМИД 20670278 .
- ^ Jump up to: а б Оу ТМ, Линь Дж, Лу YJ, Хоу JQ, Тан Дж. Х., Чен Ш, Ли З, Ли ЮП, Ли Д, Гу LQ, Хуан З.С. (август 2011 г.). «Ингибирование пролиферации клеток производным хиндолина (SYUIQ-05) посредством его преимущественного взаимодействия с G-квадруплексом промотора c-myc». Журнал медицинской химии . 54 (16): 5671–9. дои : 10.1021/jm200062u . ПМИД 21774525 .
- ^ Шарма С., Мукерджи А.К., Рой С.С., Багри С., Лиер С., Верма М., Сенгупта А., Кумар М., Нессе Дж., Панди Д.П., Чоудхури С. (январь 2020 г.). «Экспрессия теломеразы человека находится под прямым транскрипционным контролем фактора связывания теломер TRF2» (PDF) . биоRxiv . дои : 10.1101/2020.01.15.907626 . S2CID 214472968 .
- ^ Сунь Д., Го К., Руше Дж.Дж., Херли Л.Х. (12 октября 2005 г.). «Облегчение структурного перехода в полипурин/полипиримидиновом тракте в проксимальной промоторной области человеческого гена VEGF за счет присутствия взаимодействующих с калием и G-квадруплексом агентов» . Исследования нуклеиновых кислот . 33 (18): 6070–80. дои : 10.1093/nar/gki917 . ПМК 1266068 . ПМИД 16239639 .
- ^ Хуссейн Т., Саха Д., Пурохит Г., Мукерджи А.К., Шарма С., Сенгупта С., Дхапола П., Маджи Б., Ведагопурам С., Хорикоши Н.Т., Хорикоши Н., Пандита Р.К., Бхаттачарья С., Баджадж А., Риу Дж.Ф., Пандита Т.К., Чоудхури С. (сентябрь 2017 г.). «Регуляция транскрипции CDKN1A (p21/CIP1/WAF1) с помощью TRF2 эпигенетически контролируется через репрессорный комплекс REST» . Научные отчеты . 7 (1): 11541. Бибкод : 2017NatSR...711541H . дои : 10.1038/s41598-017-11177-1 . ПМЦ 5599563 . ПМИД 28912501 .
- ^ Де Армонд Р., Вуд С., Сан Д., Херли Л.Х., Эббингауз С.В. (декабрь 2005 г.). «Доказательства присутствия области, образующей гуаниновый квадруплекс, в полипуриновом тракте промотора фактора 1альфа, индуцируемого гипоксией». Биохимия . 44 (49): 16341–50. дои : 10.1021/bi051618u . ПМИД 16331995 .
- ^ Го К., Пурпак А., Битц-Роджерс К., Гохале В., Сан Д., Херли Л.Х. (август 2007 г.). «Формирование псевдосимметричных структур G-квадруплекса и i-мотива в проксимальной промоторной области онкогена RET» . Журнал Американского химического общества . 129 (33): 10220–8. дои : 10.1021/ja072185g . ПМК 2566970 . ПМИД 17672459 .
- ^ Цинь Ю, Резлер Э.М., Гохале В., Сан Д., Херли Л.Х. (26 ноября 2007 г.). «Характеристика G-квадруплексов в гиперчувствительном к дуплексной нуклеазе элементе промотора PDGF-A и модуляция активности промотора PDGF-A с помощью TMPyP4» . Исследования нуклеиновых кислот . 35 (22): 7698–713. дои : 10.1093/нар/gkm538 . ПМК 2190695 . ПМИД 17984069 .
- ^ Чилакамарти У., Котешвар Д., Джинка С., Вамси Кришна Н., Шридхаран К., Нагеш Н., Гирибабу Л. (ноябрь 2018 г.). «Новое амфифильное синтетическое производное TMPyP4, связывающееся с G-квадруплексом, и его влияние на пролиферацию раковых клеток и индукцию апоптоза». Биохимия . 57 (46): 6514–6527. doi : 10.1021/acs.biochem.8b00843 . ПМИД 30369235 . S2CID 53093959 .
- ^ Онмахт С.А., Маркетти С., Гунаратнам М., Бессер Р.Дж., Хайдер С.М., Ди Вита Г., Лоу Х.Л., Меллинас-Гомез М., Диоку С., Робсон М., Шпонер Дж., Ислам Б., Педли Р.Б., Хартли Дж.А., Нидл С. (июнь 2015 г.) ). «Соединение, связывающее G-квадруплекс, проявляющее противоопухолевую активность на модели рака поджелудочной железы in vivo» . Научные отчеты . 5 : 11385. Бибкод : 2015NatSR...511385O . дои : 10.1038/srep11385 . ПМЦ 4468576 . ПМИД 26077929 .
- ^ Сиддики-Джейн А., Grand CL, Bearss DJ, Херли Л.Х. (сентябрь 2002 г.). «Прямое доказательство наличия G-квадруплекса в промоторной области и его нацеливания с помощью небольшой молекулы для подавления транскрипции c-MYC» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (18): 11593–8. Бибкод : 2002PNAS...9911593S . дои : 10.1073/pnas.182256799 . ПМК 129314 . ПМИД 12195017 .
- ^ Колли Г.В., Промонторио Р., Хампель С.М., Микко М., Нейдл С., Паркинсон Г.Н. (февраль 2012 г.). «Структурная основа нацеливания теломерного G-квадруплекса нафталиндиимидными лигандами» . Журнал Американского химического общества . 134 (5): 2723–31. дои : 10.1021/ja2102423 . ПМИД 22280460 .
- ^ Тодд А.К., Джонстон М., Нидл С. (2005). «Высоко распространенные предполагаемые мотивы квадруплексной последовательности в ДНК человека» . Исследования нуклеиновых кислот . 33 (9): 2901–7. дои : 10.1093/nar/gki553 . ПМК 1140077 . ПМИД 15914666 .
- ^ Франк-Каменецкий М.Д., Миркин С.М. (1995). «Триплексные структуры ДНК». Ежегодный обзор биохимии . 64 (9): 65–95. дои : 10.1146/annurev.bi.64.070195.000433 . ПМИД 7574496 .
- ^ Го К., Гохале В., Херли Л.Х., Сунь Д. (август 2008 г.). «Внутримолекулярно свернутые структуры G-квадруплекса и i-мотива в проксимальном промоторе гена фактора роста эндотелия сосудов» . Исследования нуклеиновых кислот . 36 (14): 4598–608. дои : 10.1093/нар/gkn380 . ПМК 2504309 . ПМИД 18614607 .
- ^ Миркин С.М., Лямичев В.И., Друшляк К.Н., Добрынин В.Н., Филиппов С.А., Франк-Каменецкий М.Д. (1987). «Н-форма ДНК требует зеркального повтора гомопурин-гомопиримидина». Природа . 330 (6147): 495–7. Бибкод : 1987Natur.330..495M . дои : 10.1038/330495a0 . ПМИД 2825028 . S2CID 4360764 .
- ^ Смит СС (2010). «Эволюционное расширение структурно сложных последовательностей ДНК». Геномика и протеомика рака . 7 (4): 207–15. ПМИД 20656986 .
- ^ Бразда, Вацлав; Коломазник, Ян; Лысек, Иржи; Бартас, Мартин; Фойта, Мирослав; Штястный, Иржи; Мерни, Жан-Луи (15 сентября 2019 г.). Хэнкок, Джон (ред.). «Веб-приложение G4Hunter: веб-сервер для прогнозирования G-квадруплексов» . Биоинформатика . 35 (18): 3493–3495. doi : 10.1093/биоинформатика/btz087 . ISSN 1367-4803 . ПМЦ 6748775 . ПМИД 30721922 .
- ^ Гарант, Жан-Мишель; Перро, Жан-Пьер; Скотт, Мишель С. (2018). «Веб-сервер скрининга G4RNA: ориентированный на пользователя интерфейс для прогнозирования G-квадруплекса РНК» . Биохимия . 151 : 115–118. дои : 10.1016/j.biochi.2018.06.002 . ПМИД 29885355 . S2CID 47005625 .
- ^ Квок С.К., Меррик С.Дж. (октябрь 2017 г.). «G-квадруплексы: прогнозирование, характеристика и биологическое применение» (PDF) . Тенденции в биотехнологии . 35 (10): 997–1013. дои : 10.1016/j.tibtech.2017.06.012 . ПМИД 28755976 .
- ^ Хан Х., Херли Л.Х., Салазар М. (январь 1999 г.). «Анализ остановки ДНК-полимеразы для соединений, взаимодействующих с G-квадруплексом» . Исследования нуклеиновых кислот . 27 (2): 537–542. дои : 10.1093/нар/27.2.537 . ПМК 148212 . ПМИД 9862977 .
- ^ Сан Д., Херли Л.Х. (23 октября 2009 г.). «Биохимические методы характеристики структур G-квадруплекса: EMSA, DMS Footprinting и стоп-анализ ДНК-полимеразы». G-квадруплексная ДНК . Методы молекулярной биологии. Том. 608. Хумана Пресс. стр. 65–79. дои : 10.1007/978-1-59745-363-9_5 . ISBN 9781588299505 . ПМЦ 2797547 . ПМИД 20012416 .
- ^ Парамасиван С., Ружан И., Болтон П.Х. (декабрь 2007 г.). «Круговой дихроизм квадруплексных ДНК: применение к структуре, катионным эффектам и связыванию лигандов». Методы . 43 (4): 324–331. дои : 10.1016/j.ymeth.2007.02.009 . ПМИД 17967702 .
- ^ Мергни Дж.Л., Фан А.Т., Лакруа Л. (сентябрь 1998 г.). «После образования G-квартета с помощью УФ-спектроскопии» . Письма ФЭБС . 435 (1): 74–78. дои : 10.1016/s0014-5793(98)01043-6 . ПМИД 9755862 . S2CID 1306129 .
- ^ Ан Н., Флеминг А.М., Миддлтон Э.Г., Берроуз С.Дж. (октябрь 2014 г.). «Одномолекулярное исследование G-квадруплексных складок теломерной последовательности человека в белковой нанополости» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (40): 14325–14331. Бибкод : 2014PNAS..11114325A . дои : 10.1073/pnas.1415944111 . ПМК 4209999 . ПМИД 25225404 .
- ^ Бошкович Ф., Чжу Дж., Чен К., Кейсер У.Ф. (ноябрь 2019 г.). «Мониторинг образования G-квадруплекса с помощью носителей ДНК и твердотельных нанопор». Нано-буквы . 19 (11): 7996–8001. Бибкод : 2019NanoL..19.7996B . дои : 10.1021/acs.nanolett.9b03184 . ПМИД 31577148 . S2CID 203638480 .
- ^ Симона Р., Фратта П., Нидл С., Паркинсон Г.Н., Исаакс А.М. (июнь 2015 г.). «G-квадруплексы: новая роль в нейродегенеративных заболеваниях и некодирующем транскриптоме» . Письма ФЭБС . 589 (14): 1653–68. doi : 10.1016/j.febslet.2015.05.003 . ПМИД 25979174 .
- ^ «C9orf72 хромосома 9 открытая рамка считывания 72 [Homo sapiens] - Джин» . Национальный центр биотехнологической информации . Национальная медицинская библиотека США.
- ^ Ратнавалли Э., Брейн С., Доусон К., Ходжес-младший (июнь 2002 г.). «Распространенность лобно-височной деменции». Неврология . 58 (11): 1615–21. дои : 10.1212/WNL.58.11.1615 . ПМИД 12058088 . S2CID 45904851 .
- ^ Резерфорд Н.Дж., Хекман М.Г., Дехесус-Эрнандес М., Бейкер М.К., Сото-Ортолаза А.И., Раяпролу С., Стюарт Х., Фингер Е., Волкенинг К., Сили У.В., Хатанпаа К.Дж., Ломен-Хёрт К., Кертес А., Бигио Э.Х., Липпа К. , Кнопман Д.С., Кречмар Х.А., Нойманн М., Казелли Р.Дж., Уайт КЛ, Маккензи И.Р., Петерсен Р.С., Стронг М.Дж., Миллер Б.Л., Боев Б.Ф., Уитти Р.Дж., Бойлан К.Б., Вшолек З.К., Графф-Рэдфорд Н.Р., Диксон Д.В., Росс О.А. , Радемакерс Р. (декабрь 2012 г.). «Длина нормальных аллелей повтора C9ORF72 GGGGCC не влияет на фенотип заболевания» . Нейробиология старения . 33 (12): 2950.e5–7. doi : 10.1016/j.neurobiolaging.2012.07.005 . ПМЦ 3617405 . ПМИД 22840558 .
- ^ Бек Дж., Поултер М., Хенсман Д., Рорер Дж.Д., Махони СиДжей, Адамсон Дж., Кэмпбелл Т., Апхилл Дж., Борг А., Фратта П., Оррелл Р.В., Маласпина А., Роу Дж., Браун Дж., Ходжес Дж., Сидл К., Полке Дж.М. , Хоулден Х., Шотт Дж.М., Фокс Н.К., Россор М.Н. , Тебризи С.Дж., Исаакс А.М., Харди Дж., Уоррен Дж.Д., Коллиндж Дж., Мид С. (март 2013 г.). «Большие экспансии гексануклеотидных повторов C9orf72 наблюдаются при множественных нейродегенеративных синдромах и встречаются чаще, чем ожидалось, в популяции Великобритании» . Американский журнал генетики человека . 92 (3): 345–53. дои : 10.1016/j.ajhg.2013.01.011 . ПМЦ 3591848 . ПМИД 23434116 .
- ^ Фратта П., Мизиелинска С., Николл А.Дж., Зло М., Фишер Э.М., Паркинсон Г., Исаакс А.М. (декабрь 2012 г.). «Гексануклеотидный повтор C9orf72, связанный с боковым амиотрофическим склерозом и лобно-височной деменцией, образует G-квадруплексы РНК» . Научные отчеты . 2 : 1016. Бибкод : 2012NatSR...2E1016F . дои : 10.1038/srep01016 . ПМЦ 3527825 . ПМИД 23264878 .
- ^ Редди К., Замири Б., Стэнли С.Ю., Макгрегор Р.Б., Пирсон CE (апрель 2013 г.). «Связанный с заболеванием повтор r(GGGGCC)n из гена C9orf72 образует зависимые от длины тракта одно- и мультимолекулярные G-квадруплексные структуры РНК» . Журнал биологической химии . 288 (14): 9860–6. дои : 10.1074/jbc.C113.452532 . ПМЦ 3617286 . ПМИД 23423380 .
- ^ Хеуслер А.Р., Доннелли С.Дж., Периз Г., Симко Е.А., Шоу П.Г., Ким М.С., Марагакис Н.Дж., Тронкосо Дж.К., Панди А., Саттлер Р., Ротштейн Дж.Д., Ван Дж. (март 2014 г.). «Структуры нуклеотидных повторов C9orf72 инициируют молекулярные каскады заболеваний» . Природа . 507 (7491): 195–200. Бибкод : 2014Natur.507..195H . дои : 10.1038/nature13124 . ПМК 4046618 . ПМИД 24598541 .
- ^ Дарнелл Дж.К., Дженсен К.Б., Джин П., Браун В., Уоррен С.Т., Дарнелл Р.Б. (ноябрь 2001 г.). «Хрупкий белок умственной отсталости X нацелен на мРНК G-квартета, важные для функции нейронов» . Клетка . 107 (4): 489–499. дои : 10.1016/S0092-8674(01)00566-9 . ПМИД 11719189 . S2CID 8203054 .
- ^ Семан С., О'Доннелл В.Т., Рид М., Паттон С., Пол Дж., Уоррен С.Т. (декабрь 2003 г.). «Фосфорилирование влияет на состояние трансляции FMRP-ассоциированных полирибосом» . Молекулярная генетика человека . 12 (24): 3295–3305. дои : 10.1093/hmg/ddg350 . ПМИД 14570712 .
- ^ Фэлинг М., Мровка Р., Стидж А., Киршнер К.М., Бенко Е., Фёрстера Б. и др. (февраль 2009 г.). «Трансляционная регуляция человеческого гомолога ахета-щитка-1 с помощью хрупкого X-белка умственной отсталости» . Журнал биологической химии . 284 (7): 4255–4266. дои : 10.1074/jbc.M807354200 . ПМИД 19097999 .
- ^ «Хрупкая умственная отсталость» . Генные карты .
- ^ Пьеретти М., Чжан Ф.П., Фу Ю.Х., Уоррен С.Т., Остра Б.А., Каски Коннектикут, Нельсон Д.Л. (август 1991 г.). «Отсутствие экспрессии гена FMR-1 при синдроме хрупкой Х». Клетка . 66 (4): 817–822. дои : 10.1016/0092-8674(91)90125-I . ПМИД 1878973 . S2CID 31455523 .
- ^ Сатклифф Дж.С., Нельсон Д.Л., Чжан Ф., Пьеретти М., Кэски КТ, Сакс Д., Уоррен С.Т. (сентябрь 1992 г.). «Метилирование ДНК подавляет транскрипцию FMR-1 при синдроме ломкой Х-хромосомы». Молекулярная генетика человека . 1 (6): 397–400. дои : 10.1093/hmg/1.6.397 . ПМИД 1301913 .
- ^ Мизелинска С., Исаакс А.М. (октябрь 2014 г.). «Боковой амиотрофический склероз C9orf72 и лобно-височная деменция: усиление или потеря функции?» . Современное мнение в неврологии . 27 (5): 515–23. doi : 10.1097/WCO.0000000000000130 . ПМК 4165481 . ПМИД 25188012 .
- ^ Доннелли С.Дж., Чжан П.В., Фам Дж.Т., Хеуслер А.Р., Хойслер А.Р., Мистри Н.А., Виденски С., Дэйли Э.Л., Пот Э.М., Гувер Б., Файнс Д.М., Марагакис Н., Тиенари П.Дж., Петручелли Л., Трейнор Б.Дж., Ван Дж., Риго Ф. , Беннетт К.Ф., Блэкшоу С., Сэттлер Р., Ротштейн Дж.Д. (октябрь 2013 г.). «Токсичность РНК, вызванная экспансией ALS/FTD C9ORF72, смягчается антисмысловым вмешательством» . Нейрон . 80 (2): 415–28. дои : 10.1016/j.neuron.2013.10.015 . ПМЦ 4098943 . ПМИД 24139042 .
- ^ Лажье-Туренн К, Бон М, Риго Ф, Сан С, Лю П, Ли ХР, Цзян Дж, Ватт А.Т., Чун С, Кац М, Цю Дж, Сунь Ю, Лин СК, Чжу Ц, Полимениду М, Дреннер К, Артатес Дж.В., МакАлонис-Даунс М., Маркмиллер С., Хатт К.Р., Пиццо Д.П., Кэди Дж., Хармс М.Б., Балох Р.Х., Ванденберг С.Р., Йео Г.В., Фу XD, Беннетт К.Ф., Кливленд Д.В., Равитс Дж. (ноябрь 2013 г.). «Направленная деградация очагов смысловой и антисмысловой РНК C9orf72 как терапия БАС и лобно-височной дегенерации» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (47): E4530–9. Бибкод : 2013PNAS..110E4530L . дои : 10.1073/pnas.1318835110 . ПМЦ 3839752 . ПМИД 24170860 .
- ^ Сарин Д., О'Рурк Дж.Г., Мира П., Мухаммад А.К., Грант С., Симпкинсон М., Белл С., Кармона С., Орнелас Л., Сахабиан А., Гендрон Т., Петручелли Л., Баун М., Равитс Дж., Хармс М.Б., Риго Ф., Беннетт К.Ф., Отис Т.С., Свендсен К.Н., Балох Р.Х. (октябрь 2013 г.). «Нацеливание на фокусы РНК в мотонейронах, полученных из ИПСК, у пациентов с БАС с расширением повторов C9ORF72» . Наука трансляционной медицины . 5 (208): 208ра149. doi : 10.1126/scitranslmed.3007529 . ПМК 4090945 . ПМИД 24154603 .
- ^ Уилер Т.М., Легер А.Дж., Панди С.К., Маклеод А.Р., Накамори М., Ченг Ш., Вентворт Б.М., Беннетт К.Ф., Торнтон, Калифорния (август 2012 г.). «Нацеливание ядерной РНК на коррекцию миотонической дистрофии in vivo» . Природа . 488 (7409): 111–5. Бибкод : 2012Natur.488..111W . дои : 10.1038/nature11362 . ПМЦ 4221572 . ПМИД 22859208 .
- ^ Ли Дж.Э., Беннетт К.Ф., Купер Т.А. (март 2012 г.). «Опосредованная РНКазой H деградация токсичной РНК при миотонической дистрофии 1 типа» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (11): 4221–6. Бибкод : 2012PNAS..109.4221L . дои : 10.1073/pnas.1117019109 . ПМК 3306674 . ПМИД 22371589 .
- ^ Кэрролл Дж.Б., Уорби СК, Саутвелл А.Л., Доти К.Н., Гринли С., Скотте Н., Хунг Дж., Беннетт К.Ф., Фрейер С.М., Хайден М.Р. (декабрь 2011 г.). «Мощные и селективные антисмысловые олигонуклеотиды, нацеленные на однонуклеотидные полиморфизмы в гене болезни Хантингтона / аллель-специфическое подавление мутантного хантингтина» . Молекулярная терапия . 19 (12): 2178–85. дои : 10.1038/mt.2011.201 . ПМЦ 3242664 . ПМИД 21971427 .
- ^ Ганьон К.Т., Пендерграф Х.М., Деливи Г.Ф., Суэйзи Э.Э., Потье П., Рэндольф Дж., Роеш Э.Б., Чаттопадхьяя Дж., Дамха М.Дж., Беннетт К.Ф., Монтайе С., Леметр М., Кори Д.Р. (ноябрь 2010 г.). «Аллель-селективное ингибирование экспрессии мутантного хантингтина с помощью антисмысловых олигонуклеотидов, нацеленных на расширенный повтор CAG» . Биохимия . 49 (47): 10166–78. дои : 10.1021/bi101208k . ПМК 2991413 . ПМИД 21028906 .
- ^ Когой С., Парамасивам М., Филичев В., Геси И., Педерсен Э.Б., Ходо Л.Е. (январь 2009 г.). «Идентификация нового мотива G-квадруплекса в промоторе KRAS и создание пирен-модифицированных G4-приманок с антипролиферативной активностью в клетках рака поджелудочной железы». Журнал медицинской химии . 52 (2): 564–568. дои : 10.1021/jm800874t . ПМИД 19099510 .
- ^ Когои С., Зорзе С., Рапоцци В., Геси И., Педерсен Э.Б., Ходо Л.Е. (апрель 2013 г.). «MAZ-связывающая G4-приманка с заблокированной нуклеиновой кислотой и скрученными интеркалирующими модификациями нуклеиновой кислоты подавляет KRAS в клетках рака поджелудочной железы и задерживает рост опухоли у мышей» . Исследования нуклеиновых кислот . 41 (7): 4049–4064. дои : 10.1093/нар/gkt127 . ПМЦ 3627599 . ПМИД 23471001 .
- ^ Кэмпбелл Н.Х., Патель М., Тофа А.Б., Гош Р., Паркинсон Г.Н., Нидл С. (март 2009 г.). «Селективность в распознавании лигандов петель G-квадруплекса». Биохимия . 48 (8): 1675–80. дои : 10.1021/bi802233v . ПМИД 19173611 .
- ^ Онмахт С.А., Нейдл С. (июнь 2014 г.). «Открытие низкомолекулярных лекарств, нацеленных на квадруплекс». Письма по биоорганической и медицинской химии . 24 (12): 2602–12. дои : 10.1016/j.bmcl.2014.04.029 . ПМИД 24814531 .
- ^ Замири Б., Редди К., Макгрегор Р.Б., Пирсон CE (февраль 2014 г.). «Порфирин TMPyP4 искажает G-квадруплексные структуры РНК ассоциированного с заболеванием повтора r(GGGGCC)n гена C9orf72 и блокирует взаимодействие РНК-связывающих белков» . Журнал биологической химии . 289 (8): 4653–9. дои : 10.1074/jbc.C113.502336 . ПМК 3931028 . ПМИД 24371143 .
- ^ Вилар Р. (2018). «Глава 12. Квадруплексы нуклеиновых кислот и металлопрепараты». В Sigel A, Sigel H, Freisinger E, Sigel RK (ред.). Металло-лекарства: разработка и действие противораковых средств . Том. 18. стр. 325–349. дои : 10.1515/9783110470734-018 . ISBN 9783110470734 . ПМИД 29394031 . S2CID 44981950 .
{{cite book}}:|journal=игнорируется ( помогите )
Дальнейшее чтение
[ редактировать ]- Рен Дж, Ван Дж, Хан Л, Ван Э, Ван Дж (октябрь 2011 г.). «Кинетическая прививка G-квадруплексов на наноструктуры ДНК для кодирования структуры и функций с помощью ДНК-машины». Химические коммуникации . 47 (38): 10563–5. дои : 10.1039/c1cc13973h . ПМИД 21858307 .
- Джонсон Дж. Э., Смит Дж. С., Козак М. Л., Джонсон Ф. Б. (август 2008 г.). «In vivo veritas: использование дрожжей для исследования биологических функций G-квадруплексов» . Биохимия . 90 (8): 1250–63. дои : 10.1016/j.biochi.2008.02.013 . ПМК 2585026 . ПМИД 18331848 .
- Юпперт Дж.Л., Баласубраманян С. (2005). «Распространенность квадруплексов в геноме человека» . Исследования нуклеиновых кислот . 33 (9): 2908–16. дои : 10.1093/nar/gki609 . ПМК 1140081 . ПМИД 15914667 .
- Тодд А.К., Джонстон М., Нидл С. (2005). «Высоко распространенные предполагаемые мотивы квадруплексной последовательности в ДНК человека» . Исследования нуклеиновых кислот . 33 (9): 2901–7. дои : 10.1093/nar/gki553 . ПМК 1140077 . ПМИД 15914666 .
- Бердж С., Паркинсон Г.Н., Хейзел П., Тодд А.К., Нидл С. (2006). «Квадруплексная ДНК: последовательность, топология и структура» . Исследования нуклеиновых кислот . 34 (19): 5402–15. дои : 10.1093/нар/gkl655 . ПМЦ 1636468 . ПМИД 17012276 .
- Сиддики-Джейн А., Grand CL, Bearss DJ, Херли Л.Х. (сентябрь 2002 г.). «Прямое доказательство наличия G-квадруплекса в промоторной области и его нацеливания с помощью небольшой молекулы для подавления транскрипции c-MYC» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (18): 11593–8. Бибкод : 2002PNAS...9911593S . дои : 10.1073/pnas.182256799 . ПМК 129314 . ПМИД 12195017 .
- Равал П., Куммарасетти В.Б., Равиндран Дж., Кумар Н., Хальдер К., Шарма Р., Мукерджи М., Дас С.К., Чоудхури С. (май 2006 г.). «Пологеномное предсказание ДНК G4 как регуляторного мотива: роль в глобальной регуляции Escherichia coli» . Геномные исследования . 16 (5): 644–55. дои : 10.1101/гр.4508806 . ПМЦ 1457047 . ПМИД 16651665 .
- Хоу X, Го W, Ся F, Не FQ, Донг Х, Тянь Ю, Вэнь Л, Ван Л, Цао Л, Ян Ю, Сюэ Дж, Сун Ю, Ван Ю, Лю Д, Цзян Л (июнь 2009 г.). «Биомиметический калий-чувствительный наноканал: конформационное переключение G-квадруплексной ДНК в синтетической нанопоре». Журнал Американского химического общества . 131 (22): 7800–5. дои : 10.1021/ja901574c . ПМИД 19435350 .
- Нидл и Баласубраманиан, изд. (2006). Квадруплексные нуклеиновые кислоты . Королевское химическое общество. ISBN 978-0-85404-374-3 . Архивировано из оригинала 30 сентября 2007 г.
- Роуленд ГБ, Барнетт К., Дюпон Дж.И., Акурати Дж., Ле В.Х., Льюис Э.А. (декабрь 2013 г.). «Влияние пиридильных заместителей на термодинамику связывания порфирина с ДНК G-квадруплекса». Биоорганическая и медицинская химия . 21 (23): 7515–22. дои : 10.1016/j.bmc.2013.09.036 . ПМИД 24148836 .
Внешние ссылки
[ редактировать ]Квадруплексные сайты
[ редактировать ]- Группа биосенсоров нанопор и аптамеров {NAB group}
- G-Quadruplex World – сайт для обсуждения публикаций и другой информации, интересующей тех, кто работает в области G-квадруплексов.
- Greglist - база данных, в которой перечислены потенциальные гены, регулируемые G-квадруплексом.
- База данных информации о квадруплексах: QuadBase от IGIB.
- GRSDB — база данных G-квадруплексов вблизи сайтов процессинга РНК.
- GRS_UTRdb. Архивировано 20 июля 2011 г. в Wayback Machine — базе данных G-квадруплексов в UTR.
- Ресурсный сайт G-квадруплекса
- Инструмент поиска мотивов, отличных от B, в базе данных не-B - веб-сервер для прогнозирования мотивов, образующих G-квадруплекс, и других мотивов, образующих ДНК, не относящихся к B, на основе последовательностей ДНК пользователей.
Инструменты для прогнозирования мотивов G-квадруплекса
[ редактировать ]- QGRS Mapper: веб-приложение для прогнозирования G-квадруплексов в нуклеотидных последовательностях и генах NCBI от группы Багги.
- Quadfinder: инструмент для прогнозирования и анализа мотивов G-квадруплекса в последовательностях ДНК/РНК от группы Маити, IGIB, Дели, Индия [ постоянная мертвая ссылка ]
- [1] G4Hunter из группы Мергни, но пользователю необходимо запустить код в R.
- [2] pqsfinder: исчерпывающий и устойчивый к несовершенствам инструмент поиска потенциальных последовательностей, образующих квадруплекс, в R.
- [3] pqsfinder: инструмент онлайн-поиска с использованием последней версии пакета R/Bioconductor.