Захват конформации хромосомы

Данная научная статья нуждается в дополнительных ссылках на вторичные или третичные источники . ( Апрель 2016 г. ) |
Методы захвата конформации хромосом (часто сокращенно называемые технологиями 3C или методами на основе 3C). [1] ) — набор методов молекулярной биологии, используемых для анализа пространственной в клетке организации хроматина . Эти методы количественно определяют количество взаимодействий между геномными локусами , которые находятся рядом в трехмерном пространстве, но могут быть разделены многими нуклеотидами в линейном геноме. [2] Такие взаимодействия могут быть результатом биологических функций, таких как взаимодействия промотор - энхансер , или случайного образования петель полимера, когда ненаправленное физическое движение хроматина вызывает столкновение локусов. [3] Частоты взаимодействия можно анализировать напрямую, [4] или их можно преобразовать в расстояния и использовать для реконструкции трехмерных структур. [5]
Основное различие между методами на основе 3C заключается в их сфере применения. Например, при использовании ПЦР для обнаружения взаимодействия в эксперименте 3C количественно оцениваются взаимодействия между двумя конкретными фрагментами. Напротив, Hi-C количественно определяет взаимодействия между всеми возможными парами фрагментов одновременно. Глубокое секвенирование материала, произведенного 3C, также позволяет получить карты полногеномных взаимодействий.
История
[ редактировать ]Исторически микроскопия была основным методом исследования ядерной организации . [6] который можно датировать 1590 годом. [7]
- В 1879 году Вальтер Флемминг ввел термин хроматин. [ нужна ссылка ]
- В 1883 году Август Вейсман связал хроматин с наследственностью.
- В 1884 году Альбрехт Коссель открыл гистоны.
- В 1888 году Саттон и Бовери предложили теорию непрерывности хроматина в течение клеточного цикла. [8]
- В 1889 году Вильгельм фон Вальдемейер ввел термин « хромосома ». [9]
- В 1928 году Эмиль Хейтц ввел термины гетерохроматин и эухроматин . [10]
- В 1942 году Конрад Уоддингтон постулировал эпигенетические ландшафты . [11]
- В 1948 году Роллин Хочкисс обнаружил метилирование ДНК. [12]
- В 1953 году Уотсон и Крик сообщили о двойной спиральной структуре ДНК на основе Розалинды Франклин . рентгеновских дифракционных изображений [13] [14]
- В 1961 году Мэри Лайон постулировала принцип Х-инактивации .
- В 1973/1974 году были открыты хроматиновые волокна. [11]
- В 1975 году Пьер Шамбон ввёл термин нуклеосомы . [11]
- В 1982 году хромосомные территории . были открыты [15]
- В 1984 году Джон Т. Лис разработал метод иммунопреципитации хроматина .
- В 1993 году был опубликован метод ядерного лигирования — метод, позволяющий определять частоту циркуляризации ДНК в растворе. Этот анализ был использован, чтобы показать, что эстроген индуцирует взаимодействие между промотором гена пролактина и близлежащим энхансером . [16]
- В 2002 году Джоб Деккер выдвинул новую идею о том, что плотные матрицы частот взаимодействия между локусами можно использовать для вывода о пространственной организации геномов. Эта идея легла в основу его разработки анализа захвата конформации хромосом (3C), опубликованного в 2002 году Джобом Деккером и его коллегами в Клекнера лаборатории Гарвардского университета . [17] [18]
- В 2003 году проект «Геном человека» был завершен.
- В 2006 году Марике Симонис изобрела 4C. [19] Дости из лаборатории Деккера изобрел 5C. [20]
- В 2007 году Б. Франклин Пью разработал инновационную технику ChIP-seq . [21]
- В 2009 году Эрез Либерман Эйден и Джоб Деккер изобрели Hi-C. [22] Мелисса Дж. Фуллвуд и Иджун Руан изобрели ChIA-PET. [23]
- В 2012 году группа Рена и группы под руководством Эдит Херд и Джоба Деккера обнаружили топологически ассоциированные домены (TAD) у млекопитающих. [24] [25]
- В 2013 году Такаши Нагано и Питер Фрейзер представили внутриядерное лигирование для Hi-C и одноклеточного Hi-C. [26]
- В 2014 году Сухас Рао, Мириам Хантли и др. разработали in-situ Hi-C и использование ферментов рестрикции с 4 резцами, а также выпустили первые наборы данных с высоким разрешением вплоть до тысячных оснований для нескольких линий клеток человека. Они также идентифицировали первые явные доказательства образования петель CTCF-Cohesin в картах Hi-C и определили правило конвергентного мотива CTCF, лежащее в основе этих петель. [27]
- В 2018 году С. Шенфельдер и др. разработали комплексный метод Hi-C захвата промотора для создания атласа дальнодействующих взаимодействий промоторов в десятках типов клеток человека и мыши в непредвзятых взаимодействиях в десятках типов клеток человека и мыши в непредвзятых взаимодействиях. -предвзятый ва .
Экспериментальные методы
[ редактировать ]Все методы 3C начинаются с аналогичного набора шагов, выполняемых на образце клеток.
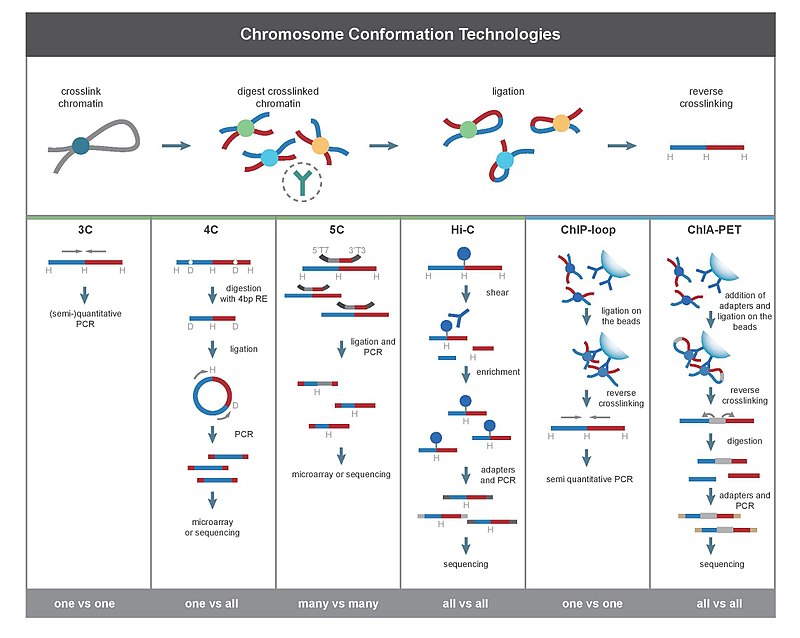
-первых, клеточные геномы сшиваются формальдегидом Во . [28] который вводит связи, которые «замораживают» взаимодействия между геномными локусами. Обработка клеток 1-3% формальдегидом в течение 10-30 минут при комнатной температуре является наиболее распространенной, однако необходима стандартизация для предотвращения высокого перекрестного сшивания белок-ДНК, поскольку это может отрицательно повлиять на эффективность рестрикционного расщепления на последующем этапе. [29] Затем геном разрезается на фрагменты с помощью эндонуклеазы рестрикции . Размер рестрикционных фрагментов определяет разрешение картирования взаимодействия. ферменты рестрикции (RE), которые разрезают последовательности распознавания длиной 6 пар оснований, такие как EcoR1 или HindIII , поскольку они разрезают геном один раз каждые 4000 пар оснований, давая ~ 1 миллион фрагментов в геноме человека. Для этой цели используются [29] [30] Для более точного отображения взаимодействия также можно использовать 4bp-распознающий RE. Следующим шагом является лигирование на основе близости . Это происходит при низких концентрациях ДНК или в интактных, проницаемых ядрах. [26] в присутствии ДНК-лигазы Т4 , [31] так что лигирование между сшитыми взаимодействующими фрагментами предпочтительнее лигирования между несшитыми фрагментами. Впоследствии взаимодействующие локусы количественно оцениваются путем амплификации лигированных соединений методами ПЦР. [29] [31]
Оригинальные методы
[ редактировать ]3C (один против одного)
[ редактировать ]Эксперимент по захвату конформации хромосомы (3C) позволяет количественно оценить взаимодействие между одной парой геномных локусов. Например, 3C можно использовать для проверки взаимодействия потенциального промотора и энхансера. Лигированные фрагменты выявляют с помощью ПЦР с известными праймерами . [2] [17] Вот почему этот метод требует предварительного знания взаимодействующих регионов.
4C (один против всех)
[ редактировать ]Захват конформации хромосомы на чипе (4C) (также известный как захват конформации кольцевой хромосомы) фиксирует взаимодействия между одним локусом и всеми другими локусами генома. Он включает второй этап лигирования для создания самоциркулярных фрагментов ДНК, которые используются для выполнения обратной ПЦР . Обратная ПЦР позволяет использовать известную последовательность для амплификации связанной с ней неизвестной последовательности. [32] [2] [19] В отличие от 3C и 5C, метод 4C не требует предварительного знания обеих взаимодействующих хромосомных областей. Результаты, полученные с помощью 4C, высоко воспроизводимы, причем большинство взаимодействий обнаруживаются между областями, проксимальными друг к другу. На одном микрочипе можно проанализировать около миллиона взаимодействий. [ нужна ссылка ]
5C (многие против многих)
[ редактировать ]Карбоновая копия захвата конформации хромосомы (5C) обнаруживает взаимодействия между всеми рестрикционными фрагментами в пределах данной области, при этом размер этой области обычно не превышает мегабазы. [2] [20] Это делается путем лигирования универсальных праймеров ко всем фрагментам. Однако 5C имеет относительно низкий охват. Техника 5C позволяет преодолеть проблемы соединения на этапе внутримолекулярного лигирования и полезна для построения сложных взаимодействий конкретных представляющих интерес локусов. Этот подход непригоден для проведения сложных полногеномных взаимодействий, поскольку для этого потребуется использование миллионов праймеров 5C. [ нужна ссылка ]
Привет-C (все против всех)
[ редактировать ]Hi-C использует высокопроизводительное секвенирование для поиска нуклеотидной последовательности фрагментов. [2] [22] и использует секвенирование парных концов , которое извлекает короткую последовательность с каждого конца каждого лигированного фрагмента. Таким образом, для данного лигированного фрагмента две полученные последовательности должны представлять собой два разных рестрикционных фрагмента, которые были лигированы вместе на этапе лигирования на основе близости. Пара последовательностей индивидуально выравнивается с геномом, тем самым определяя фрагменты, участвующие в этом событии лигирования. Таким образом, проверяются все возможные парные взаимодействия между фрагментами.
Методы, основанные на захвате последовательности
[ редактировать ]В ряде методов используется захват олигонуклеотидов для обогащения библиотек 3C и Hi-C конкретными представляющими интерес локусами. [33] [34] Эти методы включают Capture-C, [35] ИЗ Capture-C, [36] Захват-3С, [35] ПриветКап, [33] [37] Захват Hi-C. [38] и Микро Каптюр-С . [39] Эти методы способны обеспечить более высокое разрешение и чувствительность, чем методы на основе 4C. [40] Micro Capture-C обеспечивает самое высокое разрешение среди доступных методов 3C и позволяет генерировать данные о разрешении пар оснований. [39]
Одноклеточные методы
[ редактировать ]Адаптации этих методов для отдельных клеток, такие как ChIP-seq и Hi-C, можно использовать для исследования взаимодействий, происходящих в отдельных клетках. [41] [42]
Методы мультивзаимодействия
[ редактировать ]Ряд методов секвенируют несколько лигирующих соединений одновременно для обнаружения структур более высокого порядка, в которых могут взаимодействовать несколько областей хроматина. Эти методы включают Tri-C, [43] 3-сторонние 4C/C-проходы, [44] и многоконтактный 4С (МС-4С). [45]
Методы на основе иммунопреципитации
[ редактировать ]ЧИП-петля
[ редактировать ]ChIP-петля сочетает в себе 3C с ChIP-seq для обнаружения взаимодействий между двумя представляющими интерес локусами, опосредованных интересующим белком. [2] [46] ChIP-петля может быть полезна для выявления дальних цис -взаимодействий и транс -взаимодействий, опосредованных белками, поскольку при этом не происходит частых столкновений ДНК. [ нужна ссылка ]
Полногеномные методы
[ редактировать ]ChIA-PET сочетает Hi-C с ChIP-seq для обнаружения всех взаимодействий, опосредованных интересующим белком. [2] [23] HiChIP был разработан для обеспечения возможности анализа, аналогичного ChIA-PET, с меньшим количеством входного материала. [47]
Биологическое воздействие
[ редактировать ]Методы 3C привели к ряду биологических открытий, включая открытие новых структурных особенностей хромосом, каталогизацию петель хроматина и лучшее понимание механизмов регуляции транскрипции (нарушение которых может привести к заболеванию). [6]
Методы 3C продемонстрировали важность пространственной близости регуляторных элементов к генам, которые они регулируют. Например, в тканях, экспрессирующих глобиновые гены, контрольная область β-глобинового локуса образует петлю с этими генами. Эта петля не обнаруживается в тканях, где ген не экспрессируется. [48] Эта технология еще больше помогла генетическому и эпигенетическому изучению хромосом как у модельных организмов, так и у людей. [ не проверено в теле ]
Эти методы выявили крупномасштабную организацию генома в топологически ассоциированные домены (TAD), которые коррелируют с эпигенетическими маркерами. Некоторые TAD транскрипционно активны, тогда как другие репрессированы. [49] Многие TAD были обнаружены у D. melanogaster, мыши и человека. [50] Более того, CTCF и cohesin играют важную роль в определении TAD и взаимодействиях энхансер-промотор. Результат показывает, что ориентация мотивов связывания CTCF в петле энхансер-промотор должна быть обращена друг к другу, чтобы энхансер мог найти свою правильную мишень. [51]
Болезнь человека
[ редактировать ]Существует несколько заболеваний, вызванных дефектами взаимодействий промотор-энхансер, которые рассматриваются в этой статье. [52]
Бета-талассемия — это определенный тип заболевания крови, вызванный делецией энхансерного элемента LCR. [53] [54]
Голопрозэнцефалия — это заболевание головного мозга, вызванное мутацией энхансерного элемента SBE2, что, в свою очередь, ослабляет выработку гена SHH. [55]
PPD2 (полидактилия трехфалангового большого пальца) вызвана мутацией энхансера ZRS, что, в свою очередь, усиливает выработку гена SHH. [56] [57]
Аденокарцинома легкого может быть вызвана дупликацией энхансерного элемента гена MYC. [58]
Т-клеточный острый лимфобластный лейкоз вызван введением нового энхансера. [59]
Анализ данных
[ редактировать ]
Различные эксперименты в стиле 3C дают данные с очень разными структурами и статистическими свойствами. Таким образом, для каждого типа эксперимента существуют специальные пакеты анализа. [34]
Данные Hi-C часто используются для анализа общегеномной организации хроматина, такой как топологически ассоциированные домены (TAD), линейно смежные области генома, которые связаны в трехмерном пространстве. [49] Было разработано несколько алгоритмов для идентификации TAD по данным Hi-C. [4] [64]
Hi-C и его последующий анализ развиваются. Fit-Hi-C [3] это метод, основанный на подходе дискретного группирования с модификациями добавления расстояния взаимодействия (начальная аппроксимация сплайном, также известная как сплайн-1) и уточнением нулевой модели (сплайн-2). Результатом Fit-Hi-C является список парных внутрихромосомных взаимодействий с их значениями p и q. [63]
Трехмерную организацию генома также можно анализировать посредством собственного разложения контактной матрицы. Каждый собственный вектор соответствует набору локусов, которые не обязательно линейно смежны и имеют общие структурные особенности. [65]
Существенным мешающим фактором в технологиях 3C являются частые неспецифические взаимодействия между геномными локусами, которые происходят из-за случайного поведения полимера . Взаимодействие между двумя локусами должно быть подтверждено как специфическое посредством тестирования статистической значимости. [3]
Нормализация карты контактов Hi-C
[ редактировать ]Существует два основных способа нормализации необработанных тепловых карт контактов Hi-C. Первый способ — предположить равную видимость, что означает, что каждая хромосомная позиция имеет равные шансы на взаимодействие. Следовательно, истинный сигнал карты контактов Hi-C должен быть сбалансированной матрицей (сбалансированная матрица имеет постоянные суммы строк и суммы столбцов). Примером алгоритмов, предполагающих равную видимость, является алгоритм Синкхорна-Кноппа , который масштабирует необработанную карту контактов Hi-C в сбалансированную матрицу.
Другой способ — предположить, что с каждой хромосомной позицией связана предвзятость. Значение карты контактов в каждой координате будет истинным сигналом в этой позиции, умноженным на смещение, связанное с двумя позициями контактов. Примером алгоритмов, направленных на решение этой модели смещения, является итеративная коррекция, которая итеративно устраняет смещение строк и столбцов из необработанной карты контактов Hi-C. Существует ряд программных инструментов для анализа данных Hi-C. [66]
Анализ мотивов ДНК
[ редактировать ]Мотивы ДНК представляют собой специфические короткие последовательности ДНК, часто длиной 8-20 нуклеотидов. [67] которые статистически перепредставлены в наборе последовательностей с общей биологической функцией. В настоящее время регуляторные мотивы дальнодействующих взаимодействий хроматина недостаточно изучены. Несколько исследований были сосредоточены на выяснении влияния мотивов ДНК на взаимодействия промотор-энхансер.
Бэйли и др. определили, что мотив ZNF143 в областях промотора обеспечивает специфичность последовательности для взаимодействий промотор-энхансер. [68] Мутация мотива ZNF143 снизила частоту взаимодействий промотор-энхансер, что позволяет предположить, что ZNF143 является новым фактором зацикливания хроматина.
Для анализа мотивов в масштабе генома в 2016 г. Wong et al. сообщили о списке из 19 491 пары мотивов ДНК для клеточной линии K562 о взаимодействиях промотор-энхансер. [69] В результате они предположили, что множественность спаривания мотивов (количество мотивов, спаренных с данным мотивом) связана с расстоянием взаимодействия и типом регуляторной области. В следующем году Вонг опубликовал еще одну статью, в которой сообщил о 18 879 парах мотивов в 6 клеточных линиях человека. [70] Новым вкладом этой работы является MotifHyades, инструмент обнаружения мотивов , который можно напрямую применять к парным последовательностям.
Анализ генома рака
[ редактировать ]Методы на основе 3C могут дать представление о хромосомных перестройках в геномах рака. [71] Более того, они могут показывать изменения пространственной близости регуляторных элементов и их генов-мишеней, что способствует более глубокому пониманию структурной и функциональной основы генома. [72]
Ссылки
[ редактировать ]- ^ де Вит Э, де Лаат В (январь 2012 г.). «Десятилетие технологий 3C: взгляд на ядерную организацию» . Гены и развитие . 26 (1): 11–24. дои : 10.1101/gad.179804.111 . ПМЦ 3258961 . ПМИД 22215806 .
- ^ Jump up to: а б с д и ж г Хаким О, Мистели Т (март 2012 г.). «SnapShot: захват подтверждения хромосомы» . Клетка . 148 (5): 1068.e1–2. дои : 10.1016/j.cell.2012.02.019 . ПМК 6374129 . ПМИД 22385969 .
- ^ Jump up to: а б с Ай Ф, Бейли ТЛ, Нобл ВС (июнь 2014 г.). «Статистическая оценка достоверности данных Hi-C выявляет регуляторные контакты хроматина» . Геномные исследования . 24 (6): 999–1011. дои : 10.1101/гр.160374.113 . ПМК 4032863 . ПМИД 24501021 .
- ^ Jump up to: а б с Рао С.С., Хантли М.Х., Дюран Н.К., Стаменова Е.К., Бочков И.Д., Робинсон Дж.Т., Сэнборн А.Л., Махол И., Омер А.Д., Ландер Э.С., Эйден Э.Л. (декабрь 2014 г.). «Трехмерная карта человеческого генома с разрешением в тысячных базах раскрывает принципы образования петель хроматина» . Клетка . 159 (7): 1665–80. дои : 10.1016/j.cell.2014.11.021 . ПМК 5635824 . ПМИД 25497547 .
- ^ Варокво Н., Ай Ф., Нобл В.С., Верт Дж.П. (июнь 2014 г.). «Статистический подход к определению трехмерной структуры генома» . Биоинформатика . 30 (12): i26–33. doi : 10.1093/биоинформатика/btu268 . ПМК 4229903 . ПМИД 24931992 .
- ^ Jump up to: а б Денкер А., де Лаат В. (июнь 2016 г.). «Второе десятилетие технологий 3C: детальный взгляд на ядерную организацию» . Гены и развитие . 30 (12): 1357–82. дои : 10.1101/gad.281964.116 . ПМК 4926860 . ПМИД 27340173 .
- ^ «Кто изобрел микроскоп? Полная история микроскопа» . Vision Engineering Ltd. Архивировано из оригинала 22 апреля 2018 года.
- ^ Мартинс Л.А. (1999). «Предложили ли Саттон и Бовери так называемую хромосомную гипотезу Саттона-Бовери?» . Жене. Мол. Биол . 22 (2): 261–272. дои : 10.1590/S1415-47571999000200022 .
- ^ «Гены и генетика: язык научных открытий» . Оксфордский словарь английского языка . Издательство Оксфордского университета. 16 августа 2012 г. Архивировано из оригинала 29 января 2018 г. Проверено 7 декабря 2017 г.
- ^ Харрис М (05 февраля 2015 г.). «Основы гетерохроматина и эухроматина» .
- ^ Jump up to: а б с Дайхманн У (август 2016 г.). «Эпигенетика: Истоки и эволюция модной темы» . Биология развития . 416 (1): 249–254. дои : 10.1016/j.ydbio.2016.06.005 . ПМИД 27291929 .
- ^ Лу Х, Лю X, Дэн Ю, Цин Х (декабрь 2013 г.). «Метилирование ДНК, причина нейродегенеративных заболеваний» . Границы стареющей неврологии . 5 : 85. дои : 10.3389/fnagi.2013.00085 . ПМЦ 3851782 . ПМИД 24367332 .
- ^ «Документы Фрэнсиса Крика: открытие двойной спирали, 1951–1953» .
- ^ «Фрэнсис Крик, Розалинда Франклин, Джеймс Уотсон и Морис Уилкинс» . Институт истории науки . 01.06.2016 . Проверено 28 февраля 2023 г.
- ^ Кремер Т. , Кремер М. (март 2010 г.). «Хромосомные территории» . Перспективы Колд-Спринг-Харбор в биологии . 2 (3): а003889. doi : 10.1101/cshperspect.a003889 . ПМЦ 2829961 . ПМИД 20300217 .
- ^ Каллен К.Е., Кладде член парламента, Сейфред М.А. (июль 1993 г.). «Взаимодействие между областями регуляции транскрипции хроматина пролактина». Наука . 261 (5118): 203–6. Бибкод : 1993Sci...261..203C . дои : 10.1126/science.8327891 . ПМИД 8327891 .
- ^ Jump up to: а б Деккер Дж., Риппе К., Деккер М., Клекнер Н. (февраль 2002 г.). «Захват конформации хромосомы». Наука . 295 (5558): 1306–11. Бибкод : 2002Sci...295.1306D . дои : 10.1126/science.1067799 . ПМИД 11847345 . S2CID 3561891 .
- ^ Осборн К.С., Юэлс П.А., Янг А.Н. (январь 2011 г.). «Знакомьтесь с соседями: инструменты для анализа структуры и функций ядра» . Брифинги по функциональной геномике . 10 (1): 11–7. дои : 10.1093/bfgp/elq034 . ПМК 3080762 . ПМИД 21258046 .
- ^ Jump up to: а б Симонис М., Клоус П., Сплинтер Э., Мошкин Ю., Виллемсен Р., де Вит Э., ван Стинсел Б., де Лаат В. (ноябрь 2006 г.). «Ядерная организация активных и неактивных доменов хроматина, обнаруженная с помощью захвата конформации хромосомы на чипе (4C)». Природная генетика . 38 (11): 1348–54. дои : 10.1038/ng1896 . ПМИД 17033623 . S2CID 22787572 .
- ^ Jump up to: а б Дости Дж., Ричмонд Т.А., Арнаут Р.А., Зельцер Р.Р., Ли В.Л., Хонан Т.А. и др. (октябрь 2006 г.). «Углеродная копия захвата конформации хромосомы (5C): массово-параллельное решение для картирования взаимодействий между геномными элементами» . Геномные исследования . 16 (10): 1299–309. дои : 10.1101/гр.5571506 . ПМЦ 1581439 . ПМИД 16954542 .
- ^ Альберт I, Маврич Т.Н., Томшо Л.П., Ци Дж, Зантон С.Дж., Шустер С.К., Пью Б.Ф. (март 2007 г.). «Настройки трансляции и вращения нуклеосом H2A.Z в геноме Saccharomyces cerevisiae». Природа . 446 (7135): 572–6. Бибкод : 2007Natur.446..572A . дои : 10.1038/nature05632 . ПМИД 17392789 . S2CID 4416890 .
- ^ Jump up to: а б Либерман-Эйден Э., ван Беркум Н.Л., Уильямс Л., Имакаев М., Рагоци Т., Теллинг А. и др. (октябрь 2009 г.). «Комплексное картирование долгосрочных взаимодействий раскрывает принципы свертывания человеческого генома» . Наука . 326 (5950): 289–93. Бибкод : 2009Sci...326..289L . дои : 10.1126/science.1181369 . ПМЦ 2858594 . ПМИД 19815776 .
- ^ Jump up to: а б Фуллвуд М.Дж., Лю М.Х., Пан Ю.Ф., Лю Дж., Сюй Х., Мохамед Ю.Б. и др. (ноябрь 2009 г.). «Интерактивный хроматин человека, связанный с эстрогеновым рецептором-альфа» . Природа . 462 (7269): 58–64. Бибкод : 2009Natur.462...58F . дои : 10.1038/nature08497 . ПМЦ 2774924 . ПМИД 19890323 .
- ^ Диксон-младший, Сельварадж С., Юэ Ф, Ким А, Ли Ю, Шен Ю, Ху М, Лю Дж. С., Рен Б (апрель 2012 г.). «Топологические домены в геномах млекопитающих, выявленные путем анализа взаимодействий хроматина» . Природа . 485 (7398): 376–80. Бибкод : 2012Natur.485..376D . дои : 10.1038/nature11082 . ПМЦ 3356448 . ПМИД 22495300 .
- ^ Нора Э.П., Ладжуа Б.Р., Шульц Э.Г., Джорджетти Л., Окамото I, Слуга Н., Пиолот Т., ван Беркум Н.Л., Мейзиг Дж., Седат Дж., Грибнау Дж., Барилло Э., Блютген Н., Деккер Дж., Херд Э. (апрель 2012 г.). «Пространственное разделение регуляторного ландшафта центра Х-инактивации» . Природа . 485 (7398): 381–5. Бибкод : 2012Natur.485..381N . дои : 10.1038/nature11049 . ПМЦ 3555144 . ПМИД 22495304 .
- ^ Jump up to: а б Нагано, Такаши; Люблинг, Янив; Стивенс, Тим Дж.; Шенфельдер, Стефан; Яффе, Эйтан; Дин, Венди; Лауэ, Эрнест Д.; Танай, Амос; Фрейзер, Питер (октябрь 2013 г.). «Одноклеточный Hi-C обнаруживает межклеточную изменчивость структуры хромосом» . Природа . 502 (7469): 59–64. Бибкод : 2013Natur.502...59N . дои : 10.1038/nature12593 . ПМЦ 3869051 . ПМИД 24067610 .
- ^ Рао, Сухас; Хантли, Мириам (декабрь 2014 г.). «Трехмерная карта человеческого генома с разрешением в тысячных базах раскрывает принципы образования петель хроматина» . Клетка . 159 (7): 1665–1680. дои : 10.1016/j.cell.2014.11.021 . ПМК 5635824 . ПМИД 25497547 .
- ^ Гаврилов А, Эйвазова Е, Приожкова И, Липинский М, Разин С, Васецкий Ю (2009). «Захват конформации хромосомы (от 3C до 5C) и его модификация на основе чипа». Анализы иммунопреципитации хроматина . обзор. Методы молекулярной биологии. Том. 567. стр. 171–88. дои : 10.1007/978-1-60327-414-2_12 . ISBN 978-1-60327-413-5 . ПМИД 19588093 .
- ^ Jump up to: а б с Наумова Н., Смит Э.М., Жан Ю., Деккер Дж. (ноябрь 2012 г.). «Анализ дальнодействующих взаимодействий хроматина с использованием захвата конформации хромосом» . Методы . 58 (3): 192–203. дои : 10.1016/j.ymeth.2012.07.022 . ПМЦ 3874837 . ПМИД 22903059 .
- ^ Белтон Дж. М., Деккер Дж. (июнь 2015 г.). «Захват конформации хромосом (3C) у почкующихся дрожжей» . Протоколы Колд-Спринг-Харбора . 2015 (6): 580–6. дои : 10.1101/pdb.prot085175 . ПМИД 26034304 .
- ^ Jump up to: а б Гаврилов А.А., Голов А.К., Разин С.В. (26 марта 2013 г.). «Фактические частоты лигирования в процедуре захвата конформации хромосомы» . ПЛОС ОДИН . 8 (3): e60403. Бибкод : 2013PLoSO...860403G . дои : 10.1371/journal.pone.0060403 . ПМЦ 3608588 . ПМИД 23555968 .
- ^ Чжао, Чжиху; Тавусидана, Голамреза; Шолиндер, Микаэль; Гондор, Анита; Мариано, Пьеро; Ван, Ша; Кандури, Чандрасекхар; Лескано, Магда; Сандху, Кулджит Сингх; Сингх, Умашанкар; Пант, Винод; Тивари, Виджай; Курукути, Шринивасулу; Олссон, Рольф (2006). «Захват конформации круговой хромосомы (4C) раскрывает обширную сеть эпигенетически регулируемых внутри- и межхромосомных взаимодействий». Природная генетика . 38 (11): 1341–7. дои : 10.1038/ng1891 . ПМИД 17033624 . S2CID 2660843 .
- ^ Jump up to: а б Патент США 10287621.
- ^ Jump up to: а б Шмитт А.Д., Ху М., Рен Б. (декабрь 2016 г.). «Полногеномное картирование и анализ архитектуры хромосом» . Nature Reviews Молекулярно-клеточная биология . 17 (12): 743–755. дои : 10.1038/номер.2016.104 . ПМК 5763923 . ПМИД 27580841 .
- ^ Jump up to: а б Хьюз Дж.Р., Робертс Н., Макгоуэн С., Хэй Д., Джаннулату Э., Линч М. и др. (февраль 2014 г.). «Анализ сотен цис-регуляторных ландшафтов с высоким разрешением в одном высокопроизводительном эксперименте» . Природная генетика . 46 (2): 205–12. дои : 10.1038/ng.2871 . hdl : 2318/144575 . ПМИД 24413732 . S2CID 205348099 .
- ^ Дэвис Дж.О., Телениус Дж.М., Макгоуэн С.Дж., Робертс Н.А., Тейлор С., Хиггс Д.Р., Хьюз Дж.Р. (январь 2016 г.). «Мультиплексный анализ конформации хромосом со значительно повышенной чувствительностью» . Природные методы . 13 (1): 74–80. дои : 10.1038/nmeth.3664 . ПМЦ 4724891 . ПМИД 26595209 .
- ^ Сален, Пелин; Абдуллаев, Ильгар; Рамшельд, Дэниел; Мацкова, Людмила; Рилакович, Неманья; Лётстедт, Бритта; Альберт, Томас Дж.; Лундеберг, Йоаким; Сэндберг, Рикард (3 августа 2015 г.). «Полногеномное картирование взаимодействий, закрепленных на промоторе, с разрешением, близким к одному энхансеру» . Геномная биология . 16 (1): 156. дои : 10.1186/s13059-015-0727-9 . ISSN 1474-760X . ПМЦ 4557751 . ПМИД 26313521 .
- ^ Джагер Р., Мильорини Дж., Генрион М., Кандасвами Р., Спиди Х.Э., Хейндл А., Уиффин Н., Карнисер М.Дж., Брум Л., Драйден Н., Нагано Т., Шенфельдер С., Энге М., Юань Й., Тайпале Дж., Фрейзер П., Флетчер О. , Хоулстон RS (февраль 2015 г.). «Capture Hi-C идентифицирует хроматиновый интерактом локусов риска колоректального рака» . Природные коммуникации . 6 : 6178. Бибкод : 2015NatCo...6.6178J . дои : 10.1038/ncomms7178 . ПМЦ 4346635 . ПМИД 25695508 .
- ^ Jump up to: а б Хуа П., Бадат М., Ханссен Л., Хентгес Л., Крамп Н., Даунс Д., Езиорска Д.М., Уделаар А.М., Швессингер Р., Тейлор С., Милн Т.А., Хьюз Дж.Р., Хиггс Д.Р., Дэвис, Дж.О. (июнь 2021 г.). «Определение архитектуры генома с разрешением пары оснований» . Природа . 595 (7865): 125–129. Бибкод : 2021Natur.595..125H . дои : 10.1038/s41586-021-03639-4 . ПМИД 34108683 . S2CID 235394147 .
- ^ Дэвис Дж. О., Уделаар А. М., Хиггс Д. Р., Хьюз Дж. Р. (январь 2017 г.). «Как лучше всего выявить хромосомные взаимодействия: сравнение подходов» . Природные методы . 14 (2): 125–134. дои : 10.1038/nmeth.4146 . ПМИД 28139673 . S2CID 4136037 .
- ^ Нагано Т., Лублинг Ю., Стивенс Т.Дж., Шенфельдер С., Яффе Э., Дин В. и др. (октябрь 2013 г.). «Одноклеточный Hi-C обнаруживает межклеточную изменчивость структуры хромосом» . Природа . 502 (7469): 59–64. Бибкод : 2013Natur.502...59N . дои : 10.1038/nature12593 . ПМЦ 3869051 . ПМИД 24067610 .
- ^ Шварцман О., Танай А. (декабрь 2015 г.). «Одноклеточная эпигеномика: методы и новые приложения». Обзоры природы Генетика . 16 (12): 716–26. дои : 10.1038/nrg3980 . ПМИД 26460349 . S2CID 10326803 .
- ^ Уделаар, А. Марике; Дэвис, Джеймс О.Дж.; Ханссен, Ларс Л.П.; Телениус, Елена М.; Швессингер, Рон; Лю, Ю; Браун, Джилл М.; Даунс, Дэмиен Дж.; Кьяриелло, Андреа М.; Бьянко, Симона; Никодеми, Марио (2018). «Одноаллельные взаимодействия хроматина идентифицируют регуляторные центры в динамически разделенных доменах» . Природная генетика . 50 (12): 1744–1751. дои : 10.1038/s41588-018-0253-2 . ISSN 1546-1718 . ПМК 6265079 . ПМИД 30374068 .
- ^ Оливарес-Шове, Педро; Мукамель, Зоар; Лифшиц, Авиезер; Шварцман, Омер; Элькаям, Ноа Одед; Люблинг, Янив; Дейкус, Гинтарас; Себра, Роберт П.; Танай, Амос (2016). «Захват парных и многоходовых хромосомных конформаций с помощью хромосомных блужданий» . Природа . 540 (7632): 296–300. Бибкод : 2016Natur.540..296O . дои : 10.1038/nature20158 . ISSN 1476-4687 . ПМИД 27919068 . S2CID 786054 .
- ^ Аллахьяр, Амин; Вермюлен, Карло; Бауман, Бритта AM; Воин, Питер Х.Л.; Верстеген, Марджон ДЖЕМ; Дарение, Герт; ван Краненбург, Мелисса; Питерс, Марк; Стрейвер, Рой; Хаархейс, Джудит HI; Джалинк, Кес (2018). «Концентраторы энхансеров и коллизии петель, выявленные на основе одноаллельных топологий» . Природная генетика . 50 (8): 1151–1160. дои : 10.1038/s41588-018-0161-5 . ISSN 1546-1718 . ПМИД 29988121 . S2CID 49667747 .
- ^ Хорике С., Цай С., Мияно М., Ченг Дж.Ф., Кохви-Сигемацу Т. (январь 2005 г.). «Потеря петли молчащего хроматина и нарушение импринтинга DLX5 при синдроме Ретта». Природная генетика . 37 (1): 31–40. дои : 10.1038/ng1491 . ПМИД 15608638 . S2CID 2884412 .
- ^ Мумбах М.Р., Рубин А.Дж., Флинн Р.А., Дай С., Хавари П.А., Гринлиф У.Дж., Чанг Х.И. (ноябрь 2016 г.). «HiChIP: эффективный и чувствительный анализ белково-ориентированной архитектуры генома» . Природные методы . 13 (11): 919–922. дои : 10.1038/nmeth.3999 . ПМК 5501173 . ПМИД 27643841 .
- ^ Толуис Б., Палстра Р.Дж., Сплинтер Э., Гросвельд Ф., де Лаат В. (декабрь 2002 г.). «Петля и взаимодействие между сверхчувствительными сайтами в активном локусе бета-глобина» . Молекулярная клетка . 10 (6): 1453–65. дои : 10.1016/S1097-2765(02)00781-5 . ПМИД 12504019 .
- ^ Jump up to: а б Кавалли Дж., Мистели Т. (март 2013 г.). «Функциональные последствия топологии генома» . Структурная и молекулярная биология природы . 20 (3): 290–9. дои : 10.1038/nsmb.2474 . ПМК 6320674 . ПМИД 23463314 .
- ^ Деккер Дж., Марти-Реном М.А., Мирный Л.А. (июнь 2013 г.). «Изучение трехмерной организации геномов: интерпретация данных о взаимодействии хроматина» . Обзоры природы Генетика . 14 (6): 390–403. дои : 10.1038/nrg3454 . ПМЦ 3874835 . ПМИД 23657480 .
- ^ Го Ю, Сюй Q, Канцио Д, Шоу Дж, Ли Дж, Горкин Д.У. и др. (август 2015 г.). «Инверсия CRISPR сайтов CTCF изменяет топологию генома и функцию энхансера/промотора» . Клетка . 162 (4): 900–10. дои : 10.1016/j.cell.2015.07.038 . ПМЦ 4642453 . ПМИД 26276636 .
- ^ Крийгер П.Х., де Лаат В. (декабрь 2016 г.). «Регуляция экспрессии генов, связанных с заболеваниями, в 3D-геноме». Nature Reviews Молекулярно-клеточная биология . 17 (12): 771–782. дои : 10.1038/номер.2016.138 . ПМИД 27826147 . S2CID 11484886 .
- ^ Фрич Э.Ф., Лоун Р.М., Маниатис Т. (июнь 1979 г.). «Характеристика делеций, влияющих на экспрессию генов фетального глобина у человека». Природа . 279 (5714): 598–603. Бибкод : 1979Natur.279..598F . дои : 10.1038/279598a0 . ПМИД 450109 . S2CID 4243029 .
- ^ Ван дер Плог Л.Х., Конингс А., Оорт М., Роос Д., Бернини Л., Флавелл Р.А. (февраль 1980 г.). «Исследования гамма-бета-талассемии, показывающие, что удаление гамма- и дельта-генов влияет на экспрессию гена бета-глобина у человека». Природа . 283 (5748): 637–42. Бибкод : 1980Natur.283..637V . дои : 10.1038/283637a0 . ПМИД 6153459 . S2CID 4371542 .
- ^ Чон Й., Эль-Джейк К., Росслер Э., Муенке М., Эпштейн DJ (февраль 2006 г.). «Функциональный скрининг регуляторных элементов звукового ежа в интервале 1 МБ идентифицирует дальнодействующие вентральные энхансеры переднего мозга» . Разработка . 133 (4): 761–72. дои : 10.1242/dev.02239 . ПМИД 16407397 .
- ^ Леттис Л.А., Хини С.Дж., Перди Л.А., Ли Л., де Бир П., Остра Б.А. и др. (июль 2003 г.). «Дальний энхансер Shh регулирует экспрессию в развивающихся конечностях и плавниках и связан с преаксиальной полидактилией» . Молекулярная генетика человека . 12 (14): 1725–35. дои : 10.1093/hmg/ddg180 . ПМИД 12837695 .
- ^ Вечорек Д., Павлик Б., Ли Ю., Акарсу Н.А., Калиебе А., Мэй К.Дж. и др. (январь 2010 г.). «Специфическая мутация в цис-регуляторе отдаленного звукового ежа (SHH) (ZRS) вызывает мезомелический синдром Вернера (WMS), в то время как полные дупликации ZRS лежат в основе полисиндактилии типа Хааса и преаксиальной полидактилии (PPD) с трехфаланговым большим пальцем или без него». Человеческая мутация . 31 (1): 81–9. дои : 10.1002/humu.21142 . ПМИД 19847792 . S2CID 1715146 .
- ^ Чжан X, Чой П.С., Фрэнсис Дж.М., Имелински М., Ватанабэ Х., Черняк А.Д., Мейерсон М. (февраль 2016 г.). «Идентификация фокально амплифицированных суперэнхансеров, специфичных для линии, при эпителиальном раке человека» . Природная генетика . 48 (2): 176–82. дои : 10.1038/ng.3470 . ПМЦ 4857881 . ПМИД 26656844 .
- ^ Мансур М.Р., Абрахам Б.Дж., Андерс Л., Березовская А., Гутьеррес А., Дурбин А.Д. и др. (декабрь 2014 г.). «Регуляция онкогена. Онкогенный суперэнхансер, образующийся в результате соматической мутации некодирующего межгенного элемента» . Наука . 346 (6215): 1373–7. дои : 10.1126/science.1259037 . ПМЦ 4720521 . ПМИД 25394790 .
- ^ Ладжуа Б.Р., ван Беркум Н.Л., Саньял А., Деккер Дж. (октябрь 2009 г.). «My5C: веб-инструменты для исследования конформации хромосом» . Природные методы . 6 (10): 690–1. дои : 10.1038/nmeth1009-690 . ПМК 2859197 . ПМИД 19789528 .
- ^ Дэн X, Ма В, Рамани В, Хилл А, Ян Ф, Ай Ф и др. (август 2015 г.). «Двудольная структура неактивной Х-хромосомы мыши» . Геномная биология . 16 (1): 152. дои : 10.1186/s13059-015-0728-8 . ПМЦ 4539712 . ПМИД 26248554 .
- ^ Чжоу X, Лоудон Р.Ф., Ли Д., Лоусон Х.А., Мэдден П.А., Костелло Дж.Ф., Ван Т (май 2013 г.). «Изучение долгосрочных взаимодействий генома с помощью браузера эпигенома WashU» . Природные методы . 10 (5): 375–6. дои : 10.1038/nmeth.2440 . ПМК 3820286 . ПМИД 23629413 .
- ^ Jump up to: а б Ярдымджи Г.Г., Нобл В.С. (февраль 2017 г.). «Программные средства визуализации данных Hi-C» . Геномная биология . 18 (1): 26. дои : 10.1186/s13059-017-1161-y . ПМК 5290626 . ПМИД 28159004 .
- ^ Диксон Дж.Р., Сельварадж С., Юэ Ф., Ким А., Ли Ю., Шен Ю. и др. (апрель 2012 г.). «Топологические домены в геномах млекопитающих, выявленные путем анализа взаимодействий хроматина» . Природа . 485 (7398): 376–80. Бибкод : 2012Natur.485..376D . дои : 10.1038/nature11082 . ПМЦ 3356448 . ПМИД 22495300 .
- ^ Имакаев М., Фуденберг Г., МакКорд Р.П., Наумова Н., Голобородько А., Ладжуа Б.Р. и др. (октябрь 2012 г.). «Итеративная коррекция данных Hi-C выявляет признаки организации хромосом» . Природные методы . 9 (10): 999–1003. дои : 10.1038/nmeth.2148 . ПМЦ 3816492 . ПМИД 22941365 .
- ^ Имакаев М., Фуденберг Г., МакКорд Р.П., Наумова Н., Голобородько А., Лажуа Б.Р., Деккер Дж., Мирный Л.А. (октябрь 2012 г.). «Итеративная коррекция данных Hi-C выявляет признаки организации хромосом» . Природные методы . 9 (10): 999–1003. дои : 10.1038/nmeth.2148 . ПМЦ 3816492 . ПМИД 22941365 .
- ^ Замбелли Ф., Песоле Дж., Павези Дж. (март 2013 г.). «Обнаружение мотивов и сайты связывания факторов транскрипции до и после эры секвенирования следующего поколения» . Брифинги по биоинформатике . 14 (2): 225–37. дои : 10.1093/нагрудник/bbs016 . ПМК 3603212 . ПМИД 22517426 .
- ^ Бэйли, С.Д., Чжан, X., Десаи, К., Эйд, М., Коррадин, О., Каупер-Саллари, Р.,… Люпиен, М. (2015). ZNF143 обеспечивает специфичность последовательности для обеспечения взаимодействия хроматина на промоторах генов. Nature Communications, 2, 6186. Получено с https://doi.org/10.1038/ncomms7186.
- ^ К. Вонг, Ю. Ли и К. Пэн, «Идентификация пар связывающих мотивов ДНК при взаимодействиях хроматина на большие расстояния у человека», том. 32, нет. Сентябрь 2015 г., стр. 321–324, 2016 г.
- ^ Ка-Чун Вонг; MotifHyades: максимизация ожиданий для обнаружения пар мотивов ДНК de novo в парных последовательностях, Биоинформатика, том 33, выпуск 19, 1 октября 2017 г., страницы 3028–3035, https://doi.org/10.1093/bioinformatics/btx381
- ^ Хэрвуд Л., Кишор К., Элдридж МД, Вингетт С., Пирсон Д., Шенфельдер С., Коллинз В.П., Фрейзер П. (июнь 2017 г.). «Hi-C как инструмент для точного обнаружения и характеристики хромосомных перестроек и изменения числа копий в опухолях человека» . Геномная биология . 18 (1): 125. дои : 10.1186/s13059-017-1253-8 . ПМЦ 5488307 . ПМИД 28655341 .
- ^ Taberlay PC, Achinger-Kawecka J, Lun AT, Buske FA, Sabir K, Gould CM и др. (июнь 2016 г.). «Трёхмерная дезорганизация ракового генома происходит одновременно с долгосрочными генетическими и эпигенетическими изменениями» . Геномные исследования . 26 (6): 719–31. дои : 10.1101/гр.201517.115 . ПМЦ 4889976 . ПМИД 27053337 .
Дальнейшее чтение
[ редактировать ]- Баруцу А.Р., Фриц А.Дж., Заиди СК, ван Вейнен А.Дж., Лиан Дж.Б., Стейн Дж.Л., Никерсон Дж.А., Имбальцано А.Н., Штейн Г.С. (январь 2016 г.). «Создание генома: сборник методов захвата конформации хромосом для изучения организации хроматина высшего порядка» . Журнал клеточной физиологии . 231 (1): 31–5. дои : 10.1002/jcp.25062 . ПМК 4586368 . ПМИД 26059817 .
- Марбути М., Кошул Р. (декабрь 2015 г.). «Метагеномный анализ с использованием данных высокопроизводительного захвата конформации хромосом (3C)» . обзор. Тенденции в генетике . 31 (12): 673–682. дои : 10.1016/j.tig.2015.10.003 . ПМК 6831814 . ПМИД 26608779 .
- Деккер Дж. (25 ноября 2014 г.). «Два способа сворачивания генома во время клеточного цикла: данные, полученные с помощью захвата конформации хромосом» . Эпигенетика и хроматин . 7 (1): 25. дои : 10.1186/1756-8935-7-25 . ПМК 4247682 . ПМИД 25435919 .
- О'Салливан Дж. М., Хенди М. Д., Пичугина Т., Уэйк Г. К., Ланговски Дж. (сентябрь – октябрь 2013 г.). «Статистическая механика захвата конформации хромосом» . Ядро . 4 (5): 390–8. дои : 10.4161/nucl.26513 . ПМЦ 3899129 . ПМИД 24051548 .
- Умбаргер М.А. (ноябрь 2012 г.). «Анализ захвата конформации хромосом у бактерий». обзор. Методы . 58 (3): 212–20. дои : 10.1016/j.ymeth.2012.06.017 . ПМИД 22776362 . S2CID 24234275 .
- Парельо В., Меркеншлагер М. (сентябрь 2005 г.). «Экспрессия генов: совместное взросление может помочь генам разойтись» . новости и комментарии. Европейский журнал генетики человека . 13 (9): 993–4. дои : 10.1038/sj.ejhg.5201464 . ПМИД 15999115 . S2CID 29714576 .
- Марвин М., Тан-Вонг С.М. (23 апреля 2016 г.). «Захват конформации хромосомы» (коммерческий метод) . ООО «Абкам» . Проверено 23 апреля 2016 г.
См. также
[ редактировать ]- Генетическое тестирование – Медицинский тест