Плавательный пузырь


S: передняя, S': задняя часть воздушного пузыря.
œ: пищевод; l: проходимость воздушного пузыря
, Плавательный пузырь газовый пузырь , рыбья пасть или воздушный пузырь — это внутренний газонаполненный орган , который способствует способности многих костных рыб (но не хрящевых рыб) [1] ), чтобы контролировать свою плавучесть и, таким образом, оставаться на текущей глубине воды, не тратя энергию на плавание. [2] Кроме того, дорсальное положение плавательного пузыря означает, что расширение пузыря смещает центр массы вниз, что позволяет ему действовать как стабилизирующий агент у некоторых видов. Кроме того, плавательный пузырь функционирует как резонирующая камера , производящая или воспринимающая звук.
Плавательный пузырь гомологичен легким рыб четвероногих . и двоякодышащих эволюционно Чарльз Дарвин заметил это в «Происхождении видов» . [3] Дарвин полагал, что легкие у позвоночных, дышащих воздухом, произошли от более примитивного плавательного пузыря и стали специализированной формой энтерального дыхания .
На эмбриональных стадиях некоторые виды, например красногубая собачка , [4] снова потеряли плавательный пузырь, в основном это донные обитатели, такие как рыба-погодка . Другие рыбы, такие как опа и помфрет , используют свои грудные плавники , чтобы плавать и балансировать вес головы, чтобы сохранять горизонтальное положение. обычно обитающая на дне, Морская малиновка, может использовать свои грудные плавники для подъема силы во время плавания.
Граница раздела газ/ткань в плавательном пузыре производит сильное отражение звука, которое используется в гидролокаторах для поиска рыбы.
Хрящевые рыбы , такие как акулы и скаты , не имеют плавательного пузыря. Некоторые из них могут контролировать свою глубину только плавая (с помощью динамической подъемной силы ); другие хранят жиры или масла с плотностью меньшей, чем плотность морской воды, для создания нейтральной или почти нейтральной плавучести, которая не меняется с глубиной.
Структура и функции
[ редактировать ]
Плавательный пузырь обычно состоит из двух газонаполненных мешков, расположенных в спинной части рыб, хотя у некоторых примитивных видов он имеет только один мешок. Он имеет гибкие стенки, которые сжимаются или расширяются в зависимости от давления окружающей среды . Стенки мочевого пузыря содержат очень мало кровеносных сосудов и выстланы кристаллами гуанина , что делает их непроницаемыми для газов. Регулируя орган давления газа с помощью газового сальника или овального окна, рыба может получить нейтральную плавучесть, подниматься и опускаться в широком диапазоне глубин. Благодаря спинному положению придает рыбе боковую устойчивость.
В физостомных плавательных пузырях сохраняется связь между плавательным пузырем и кишкой — воздуховодом, позволяющая рыбам наполнять плавательный пузырь, «заглатывая» воздух. Излишек газа можно удалить аналогичным образом.
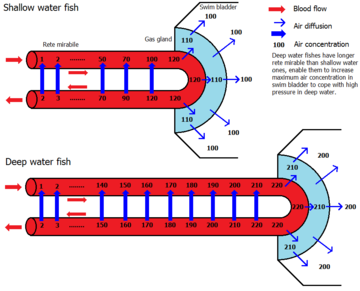
У более производных разновидностей рыб ( физоклистов ) связь с пищеварительным трактом утрачена. На ранних стадиях жизни этим рыбам приходится подниматься на поверхность, чтобы наполнить плавательный пузырь; на более поздних стадиях воздуховод исчезает, и газовой железе приходится подавать газ (обычно кислород ) в мочевой пузырь, чтобы увеличить его объем и тем самым повысить плавучесть . Этот процесс начинается с закисления крови в сетчатке, когда газовая железа выделяет молочную кислоту и выделяет углекислый газ , последний из которых подкисляет кровь через бикарбонатную буферную систему . Возникающая в результате кислотность приводит к тому, что гемоглобин крови теряет кислород ( эффект Рута ), который затем частично диффундирует в плавательный пузырь. Прежде чем вернуться в организм, кровь вновь поступает в сетчатую сеть мирабиле , и в результате практически весь избыток углекислого газа и кислорода, образующихся в газовой железе, диффундирует обратно в артерии, снабжающие газовую железу, через противоточную петлю умножения . Таким образом можно получить очень высокое давление газа кислорода, что может даже объяснить наличие газа в плавательных пузырях глубоководных рыб, таких как угорь , требующий давления в сотни бар . [5] В другом месте, в аналогичной структуре, известной как «овальное окно», мочевой пузырь контактирует с кровью, и кислород может снова диффундировать наружу. Вместе с кислородом высаливаются и другие газы. [ нужны разъяснения ] в плавательном пузыре, что также является причиной высокого давления других газов. [6]
Комбинация газов в мочевом пузыре варьируется. У мелководных рыб это соотношение близко к атмосферному , в то время как глубоководные рыбы, как правило, имеют более высокий процент кислорода. Например, у угря Synaphobranchus содержится 75,1% кислорода, 20,5% азота , 3,1% углекислого газа и 0,4% аргона в плавательном пузыре .
Плавательные пузыри Физоклиста имеют один важный недостаток: они не позволяют быстро подниматься, так как пузырь лопнет. Физостомы могут «вырыгивать» газ, но это усложняет процесс повторного погружения.
Плавательный пузырь у некоторых видов, преимущественно пресноводных рыб ( карп , сом , боуфин ), соединен между собой с внутренним ухом рыбы. Они соединены четырьмя костями, называемыми веберовскими косточками веберовского аппарата . Эти кости могут передавать вибрации мешочку и лагене . Они подходят для обнаружения звука и вибраций благодаря своей низкой плотности по сравнению с плотностью тканей тела рыбы. Это увеличивает способность обнаружения звука. [7] Плавательный пузырь может излучать звуковое давление, что помогает повысить его чувствительность и расширить слух. У некоторых глубоководных рыб, таких как Антимора , плавательный пузырь может быть также соединен с макулой мешочка , чтобы внутреннее ухо могло воспринимать звуковое давление. [8] У краснобрюхих пираний плавательный пузырь может играть важную роль в производстве звука в качестве резонатора. Звуки, издаваемые пираньями, генерируются за счет быстрых сокращений звуковых мышц и связаны с плавательным пузырем. [9]
Считается, что костистые кости лишены чувства абсолютного гидростатического давления , которое можно было бы использовать для определения абсолютной глубины. [10] Однако было высказано предположение, что костистые особи могут определять свою глубину, ощущая скорость изменения объема плавательного пузыря. [11]
Эволюция
[ редактировать ]
Пример плавательного пузыря у рыб... ясно показывает нам тот чрезвычайно важный факт, что орган, первоначально созданный для одной цели, а именно для плавучести, может быть преобразован в орган, предназначенный для совершенно другой цели, а именно для дыхания. Плавательный пузырь также использовался как дополнение к слуховым органам некоторых рыб. Все физиологи признают, что плавательный пузырь гомологичен, или «идеально сходен» по положению и строению с легкими высших позвоночных животных: следовательно, нет никаких оснований сомневаться в том, что плавательный пузырь действительно превращен в легкие или является органом, используемым исключительно для дыхания. Согласно этой точке зрения, можно заключить, что все позвоночные животные с настоящими легкими произошли обычным поколением от древнего и неизвестного прототипа, который был снабжен плавучим аппаратом или плавательным пузырем.
Чарльз Дарвин , 1859 г. [3]
Плавательные пузыри эволюционно тесно связаны (т. е. гомологичны ) легким . Первые легкие возникли у последнего общего предка Actinopterygii (рыбы с лучевыми плавниками) и Sarcopterygii (рыбы с лопастными плавниками и четвероногих ) как расширение верхнего пищеварительного тракта, которое позволяло им глотать воздух в условиях недостатка кислорода. [12] У Actinopteri (рыбы с лучеплавными плавниками, за исключением bichirs ) легкие развились в плавательный пузырь (вторично отсутствующий в некоторых линиях), который, в отличие от легких, отпочковающихся вентрально, отпочковывается дорсально из передней передней кишки. [13] [14] У целакантов есть «жировой орган», который иногда называют плавательным пузырем, но он структурно отличается и имеет отдельную эволюционную историю. [15]
В 1997 году Фармер предположил, что легкие эволюционировали, чтобы снабжать сердце кислородом. У рыб кровь циркулирует от жабр к скелетным мышцам и только потом к сердцу. Во время интенсивных физических упражнений кислород из крови используется скелетными мышцами до того, как кровь достигнет сердца. Примитивные легкие давали преимущество, снабжая сердце насыщенной кислородом кровью через сердечный шунт. Эта теория убедительно подтверждается летописью окаменелостей, экологией современных дышащих воздухом рыб и физиологией современных рыб. [16] В эмбриональном развитии и легкие, и плавательный пузырь возникают как выходы из кишечника; в случае плавательных пузырей эта связь с кишечником продолжает существовать в виде воздуховода у более «примитивных» лучепёрых рыб и теряется у некоторых более производных отрядов костистых рыб. Не существует животных, у которых были бы одновременно легкие и плавательный пузырь.
В качестве адаптации к миграции между поверхностными и более глубокими водами у некоторых рыб развился плавательный пузырь, в котором газ заменяется восковыми эфирами низкой плотности , чтобы справиться с законом Бойля . [17]
( Хрящевые рыбы например, акулы и скаты) отделились от других рыб около 420 миллионов лет назад, и у них отсутствуют легкие и плавательные пузыри, что позволяет предположить, что эти структуры развились после этого разделения. [16] Соответственно, у этих рыб также есть как гетероцеркальные , так и жесткие крыловидные грудные плавники , которые обеспечивают необходимую подъемную силу, необходимую из-за отсутствия плавательных пузырей. Костистые рыбы с плавательным пузырем обладают нейтральной плавучестью и не нуждаются в подъемной силе. [18]
Отражательная способность сонара
[ редактировать ]Плавательный пузырь рыбы может сильно отражать звук соответствующей частоты. Сильное отражение происходит, если частота настроена на объемный резонанс плавательного пузыря. Это можно рассчитать, зная ряд свойств рыбы, в частности объем плавательного пузыря, хотя общепринятый метод для этого [19] требуются поправочные коэффициенты для газосодержащего зоопланктона, у которого радиус плавательного пузыря составляет менее 5 см. [20] Это важно, поскольку рассеяние гидролокатора используется для оценки биомассы коммерчески и экологически важных видов рыб.
Глубокий рассеивающий слой
[ редактировать ]
Операторы гидролокаторов, использовавшие недавно разработанную гидролокационную технологию во время Второй мировой войны, были озадачены тем, что выглядело ложным морским дном глубиной 300–500 метров днем и меньшей глубиной ночью. Оказалось, что это произошло благодаря миллионам морских организмов, особенно мелких мезопелагических рыб, плавательные пузыри которых отражали сигналы сонара. Эти организмы мигрируют на мелководье в сумерках, чтобы питаться планктоном. Слой глубже, когда луна светит, и может стать меньше, когда облака закрывают луну. [21]
Большинство мезопелагических рыб совершают ежедневные вертикальные миграции , перемещаясь ночью в эпипелагическую зону, часто следуя за подобными миграциями зоопланктона, и возвращаясь на глубину в целях безопасности в течение дня. [22] [23] Эти вертикальные миграции часто происходят на большие вертикальные расстояния и осуществляются с помощью плавательного пузыря. Плавательный пузырь надувается, когда рыба хочет двигаться вверх, а, учитывая высокое давление в мезоплегической зоне, это требует значительных затрат энергии. По мере подъема рыбы давление в плавательном пузыре должно регулироваться, чтобы предотвратить его разрыв. Когда рыба хочет вернуться на глубину, плавательный пузырь сдувается. [24] Некоторые мезопелагические рыбы совершают ежедневные миграции через термоклин , где температура колеблется от 10 до 20 °C, демонстрируя тем самым значительную толерантность к изменению температуры.
Отбор проб с помощью глубоководного траления показывает, что на долю фонарей приходится до 65% всей биомассы глубоководных рыб . [25] Действительно, рыбы-фонарики являются одними из наиболее широко распространенных, густонаселенных и разнообразных среди всех позвоночных животных , играя важную экологическую роль в качестве добычи для более крупных организмов. По оценкам, глобальная биомасса фонарей составляет 550–660 миллионов тонн , что в несколько раз превышает годовой мировой вылов рыбы. Рыбы-фонарики также составляют большую часть биомассы, ответственной за глубокий рассеивающий слой мирового океана. Сонар отражается от миллионов плавательных пузырей рыб-фонарей, создавая впечатление ложного дна. [26]
Человеческое использование
[ редактировать ]В кулинарии Восточной Азии плавательные пузыри некоторых крупных рыб считаются пищевым деликатесом. В китайской кухне они известны как рыбья пасть , 花膠/鱼鳔. [27] и подаются в супах или рагу.
Тщеславная цена исчезающего вида пасти стала причиной неминуемого исчезновения вакита , самого маленького вида морских свиней в мире. в Мексике Встречающиеся только в Калифорнийском заливе , некогда многочисленные виды вакита сейчас находятся под угрозой исчезновения. [28] Вакита погибает в жаберных сетях [29] набор для ловли тотоабы (самой большой в мире рыбы-барабана ). На Тотоабу ведется охота из-за его пасти, которую можно продать по цене до 10 000 долларов за килограмм.
Плавательные пузыри также используются в пищевой промышленности в качестве источника коллагена . Из них можно сделать прочный водостойкий клей или использовать для изготовления изингласа для осветления пива . [30] Раньше из них делали презервативы . [31]
Заболевание плавательного пузыря
[ редактировать ]Заболевание плавательного пузыря – распространенное заболевание аквариумных рыбок . Рыба с заболеванием плавательного пузыря может плавать носом вниз и вверх хвостом, всплывать наверх или опускаться на дно аквариума. [32]
Риск травмы
[ редактировать ]Многие антропогенные виды деятельности, такие как забивка свай или даже сейсмические волны , могут создавать звуковые волны высокой интенсивности, которые вызывают внутренние повреждения рыб, обладающих газовым пузырем. Physoclisti не могут достаточно быстро удалить воздух из газового пузыря, органа, наиболее восприимчивого к звуковому повреждению, что затрудняет им предотвращение серьезных травм. Физостомы, с другой стороны, могут достаточно быстро выпускать воздух из своего газового пузыря, чтобы защитить его; тем не менее, они не могут снизить давление в других жизненно важных органах и поэтому также уязвимы для травм. [33] Некоторые из часто встречающихся травм включают разрыв газового пузыря и почечное кровотечение . В основном это влияет на общее состояние здоровья рыб, но не на уровень их смертности. [33] Исследователи использовали заполненную жидкостью высокоинтенсивную импедансную трубку (HICI-FT), волновую трубку из нержавеющей стали с электромагнитным шейкером. Он имитирует звуковые волны высокой энергии в водной дальней зоне, в акустических условиях плоских волн. [34] [35]
Подобные структуры у других организмов
[ редактировать ]У сифонофоров есть специальный плавательный пузырь, который позволяет колониям, похожим на медуз, плавать по поверхности воды, в то время как их щупальца тянутся ниже. Этот орган не имеет отношения к рыбьему. [36]
Галерея
[ редактировать ] Выставка плавательного пузыря в Малакки торговом центре
Выставка плавательного пузыря в Малакки торговом центре Суп из рыбной пасти
Суп из рыбной пасти Заболевание плавательного пузыря привело к тому, что самка золотой рыбки рюкин плавает вверх ногами
Заболевание плавательного пузыря привело к тому, что самка золотой рыбки рюкин плавает вверх ногами



Ссылки
[ редактировать ]- ^ «Еще о морфологии» . www.ucmp.berkeley.edu .
- ^ "Рыба". Энциклопедия Microsoft Encarta Deluxe, 1999 г. Майкрософт. 1999.
- ^ Перейти обратно: а б Дарвин, Чарльз (1859 г.) «Происхождение видов», стр. 190, перепечатано в 1872 г. Д. Эпплтоном.
- ^ Нурсолл, младший (1989). «Плавучесть обеспечивают липиды личинок красногубой собачки Ophioblennius atlanticus». Копейя . 1989 (3): 614–621. дои : 10.2307/1445488 . JSTOR 1445488 .
- ^ Пелстер Б. (декабрь 2001 г.). «Поколение гипербарического напряжения кислорода у рыб». Новости Физиол. Наука . 16 (6): 287–91. doi : 10.1152/физиологияонлайн.2001.16.6.287 . ПМИД 11719607 . S2CID 11198182 .
- ^ «Секреция азота в плавательный пузырь рыб. II. Молекулярный механизм. Секреция благородных газов» . Биолбулл.орг. 01.12.1981 . Проверено 24 июня 2013 г.
- ^ Кардонг, Кеннет (16 февраля 2011 г.). Позвоночные животные: сравнительная анатомия, функции, эволюция . Нью-Йорк: Образование Макгроу-Хилл. п. 701. ИСБН 9780073524238 .
- ^ Дэн, Сяохун; Вагнер, Ханс-Иоахим; Поппер, Артур Н. (1 января 2011 г.). «Внутреннее ухо и его соединение с плавательным пузырем у глубоководной рыбы Antimora rostrata (Teleostei: Moridae)» . Глубоководные исследования. Часть I: Статьи океанографических исследований . 58 (1): 27–37. Бибкод : 2011DSRI...58...27D . дои : 10.1016/j.dsr.2010.11.001 . ПМК 3082141 . ПМИД 21532967 .
- ^ Онуки, А; Омори Ю.; Сомия Х. (январь 2006 г.). «Иннервация спинномозгового нерва звуковой мышцы и звукового моторного ядра у красной пираньи, Pygocentrus nattereri (Characiformes, Ostariophys)». Мозг, поведение и эволюция . 67 (2): 11–122. дои : 10.1159/000089185 . ПМИД 16254416 . S2CID 7395840 .
- ^ Боун, К.; Мур, Ричард Х. (2008). Биология рыб (3-е, тщательно обновленное и переработанное изд.). Тейлор и Фрэнсис. ISBN 9780415375627 .
- ^ Тейлор, Грэм К.; Холбрук, Роберт Иэн; де Перера, Тереза Берт (6 сентября 2010 г.). «Дробная скорость изменения объема плавательного пузыря достоверно связана с абсолютной глубиной при вертикальных перемещениях у костистых рыб» . Журнал интерфейса Королевского общества . 7 (50): 1379–1382. дои : 10.1098/rsif.2009.0522 . ПМЦ 2894882 . ПМИД 20190038 .
- ^ Райс, Стэнли А. (2009). Энциклопедия эволюции . Информационная база. ISBN 978-1-4381-1005-9 .
- ^ Фанк, Эмили С.; Бироль, Эда Б.; МакКьюн, Эми Р. (2021). «Является ли газовый пузырь боуфина промежуточной стадией эволюционного перехода от легких к газовому пузырю?» . Журнал морфологии . 282 (4): 600–611. дои : 10.1002/jmor.21330 . ПМИД 33538055 .
- ^ Би, Сюпэн, Кун; Пань, Хайлинь, Хайфэн; Фан, Мяоцюань; Чжу, Цай, Иран; Хунхуэй, Даци; Чжу, Юань (4 марта 2021 г.). генетических следов приземления позвоночных у лучеперых рыб» . Cell 184 . (5): « 1377–1391.e14 Отслеживание j.cell.2021.01.046 . PMID 33545088 .
- ^ Купелло, Камила; Брито, Пауло М.; Хербин, Марк; Менье, Франсуа Ж.; Жанвье, Филипп; Дютель, Хьюго; Клеман, Гаэль (2015). «Аллометрический рост существующих легких целаканта во время онтогенетического развития» . Природные коммуникации . 6 : 8222. Бибкод : 2015NatCo...6.8222C . дои : 10.1038/ncomms9222 . ПМЦ 4647851 . ПМИД 26372119 .
- ^ Перейти обратно: а б Фермер, Коллин (1997). «Развились ли легкие и внутрисердечный шунт для насыщения сердца кислородом у позвоночных» (PDF) . Палеобиология . 23 (3): 358–372. Бибкод : 1997Pbio...23..358F . дои : 10.1017/S0094837300019734 . S2CID 87285937 .
- ^ Боун, Квентин; Мур, Ричард (19 марта 2008 г.). Биология рыб . Тейлор и Фрэнсис. ISBN 978-1-134-18631-0 .
- ^ Кардонг, К.В. (1998) Позвоночные животные: сравнительная анатомия, функции, эволюция, 2-е издание, иллюстрированное, исправленное. Опубликовано WCB/McGraw-Hill, с. 12 ISBN 0-697-28654-1
- ^ Любовь Р.Х. (1978). «Резонансное акустическое рассеяние рыб с плавательным пузырем». Дж. Акуст. Соц. Являюсь . 64 (2): 571–580. Бибкод : 1978ASAJ...64..571L . дои : 10.1121/1.382009 .
- ^ Байк К. (2013). «Комментарий к «Резонансному акустическому рассеянию рыбами с плавательным пузырем» [J. Acoust. Soc. Am. 64, 571–580 (1978)] (L)». Дж. Акуст. Соц. Являюсь . 133 (1): 5–8. Бибкод : 2013ASAJ..133....5B . дои : 10.1121/1.4770261 . ПМИД 23297876 .
- ^ Райан П. «Глубоководные существа: Мезопелагическая зона» Те Ара — Энциклопедия Новой Зеландии . Обновлено 21 сентября 2007 г.
- ^ Мойл, Питер Б.; Чех, Джозеф Дж. (2004). Рыбы: введение в ихтиологию (5-е изд.). Река Аппер-Сэддл, Нью-Джерси: Пирсон/Прентис-Холл. п. 585. ИСБН 9780131008472 .
- ^ Боун, Квентин; Мур, Ричард Х. (2008). «Глава 2.3. Морские среды обитания. Мезопелагические рыбы». Биология рыб (3-е изд.). Нью-Йорк: Тейлор и Фрэнсис. п. 38. ISBN 9780203885222 .
- ^ Дуглас, Эл.; Фридл, Вашингтон; Пиквелл, Г.В. (1976). «Рыбы в зонах кислородного минимума: особенности оксигенации крови» . Наука . 191 (4230): 957–959. Бибкод : 1976Sci...191..957D . дои : 10.1126/science.1251208 . ПМИД 1251208 .
- ^ Халли, П. Александр (1998). Пакстон, младший; Эшмейер, WN (ред.). Энциклопедия рыб . Сан-Диего: Академическая пресса. стр. 127–128. ISBN 978-0-12-547665-2 .
- ^ Р. Корнехо; Р. Коппельманн и Т. Саттон. «Разнообразие и экология глубоководных рыб в придонном пограничном слое» . Архивировано из оригинала 1 июня 2013 г. Проверено 26 марта 2015 г.
- ^ Тереза М. (2009) Традиция супа: ароматы дельты Жемчужной реки Китая, стр. 70, North Atlantic Books. ISBN 9781556437656 .
- ^ Рохас-Брачо, Л. и Тейлор, Б.Л. (2017). «Вакита ( Phocoena sinus )» . Красный список исчезающих видов МСОП . 2017 . doi : 10.2305/IUCN.UK.2022-1.RLTS.T17028A214541137.en . Проверено 14 октября 2022 г.
- ^ « Вымирание неизбежно»: опубликован новый отчет группы восстановления Vaquita (CIRVA)» . SSC МСОП — Группа специалистов по китообразным . 06.06.2016. Архивировано из оригинала 3 января 2019 г. Проверено 25 января 2017 г.
- ^ Bridge, TW (1905) [1] «Естественная история Изингласса»
- ^ Хаксли, Джулиан (1957). «Материал ранних противозачаточных чехлов» . Британский медицинский журнал . 1 (5018): 581–582. дои : 10.1136/bmj.1.5018.581-b . ЧВК 1974678 .
- ^ Джонсон, Эрик Л. и Ричард Э. Хесс (2006) Необычная золотая рыбка: Полное руководство по уходу и коллекционированию , Weatherhill, Shambhala Publications, Inc. ISBN 0-8348-0448-4
- ^ Перейти обратно: а б Халворсен, Мишель Б.; Каспер, Брэндон М.; Мэтьюз, Фрейзер; Карлсон, Томас Дж.; Поппер, Артур Н. (7 декабря 2012 г.). «Влияние звуков забивания свай на озерного осетра, нильскую тилапию и хогчокера» . Труды Королевского общества B: Биологические науки . 279 (1748): 4705–4714. дои : 10.1098/rspb.2012.1544 . ISSN 0962-8452 . ПМК 3497083 . ПМИД 23055066 .
- ^ Халворсен, Мишель Б.; Каспер, Брэндон М.; Вудли, Криста М.; Карлсон, Томас Дж.; Поппер, Артур Н. (20 июня 2012 г.). «Порог возникновения травмы чавычи из-за воздействия импульсивных звуков забивки свай» . ПЛОС ОДИН . 7 (6): e38968. Бибкод : 2012PLoSO...738968H . дои : 10.1371/journal.pone.0038968 . ISSN 1932-6203 . ПМК 3380060 . ПМИД 22745695 .
- ^ Поппер, Артур Н.; Хокинс, Энтони (26 января 2012 г.). Влияние шума на водную жизнь . Springer Science & Business Media. ISBN 9781441973115 .
- ^ Кларк, FE; CE Лейн (1961). «Состав плавающих газов Physalia physalis». Труды Общества экспериментальной биологии и медицины . 107 (3): 673–674. дои : 10.3181/00379727-107-26724 . ПМИД 13693830 . S2CID 2687386 .
т
Дальнейшие ссылки
[ редактировать ]- Бонд, Карл Э. (1996) Биология рыб , 2-е изд., Сондерс, стр. 283–290.
- Пелстер, Бернд (1997) «Плавучесть на глубине» В: WS Hoar, DJ Randall и AP Farrell (Eds) Deep-Sea Fishes , страницы 195–237, Academic Press. ISBN 9780080585406 .

| Базы данных органов управления : Национальные |
|---|