Рыбья чешуя

– Рыбья чешуя это небольшая жесткая пластинка, вырастающая из кожи рыбы. Кожа большинства челюстных рыб покрыта этими защитными чешуйками , которые также могут обеспечить эффективную маскировку за счет отражения и окраски , а также возможных гидродинамических преимуществ. Термин «чешуя» происходит от старофранцузского слова «escale» , означающего стручок ракушки или шелуху. [1]
Чешуя сильно различается по размеру, форме, структуре и протяженности: от прочных и жестких панцирных пластин у таких рыб, как креветки и коробочки , до микроскопических или отсутствующих у таких рыб, как угри и удильщики . Морфологию . чешуи можно использовать для определения вида рыбы, от которой она произошла Чешуя возникла у бесчелюстных остракодерм , предков всех сегодняшних челюстных рыб.Большинство костистых рыб покрыто циклоидной чешуей лососей и карпов , ктеноидной чешуей окуня или ганоидной чешуей осетровых и щук . Хрящевые рыбы ( акулы и скаты ) покрыты плакоидной чешуей. Некоторые виды вместо этого покрыты щитками , а у других нет внешнего покрытия на части или всей коже.
рыб Рыбья чешуя является частью покровной системы и образуется из мезодермального слоя дермы , что отличает ее от чешуи рептилий . [2] [3] Те же гены, которые участвуют в развитии зубов и волос у млекопитающих, также участвуют в развитии чешуи. Плакоидные чешуи хрящевых рыб также называются дермальными зубцами и структурно гомологичны зубам позвоночных. Большинство рыб также покрыты слоем слизи или слизи, который может защитить от таких патогенов, как бактерии, грибки и вирусы, а также снизить поверхностное сопротивление во время плавания рыбы.
Телодонтовые чешуйки
[ редактировать ]
Костная чешуя телодонтов , самой распространенной формы ископаемых рыб , хорошо изучена. Чешуя формировалась и отпадала на протяжении всей жизни организмов и быстро отделялась после их смерти. [4]
Кость, ткань, устойчивая к механическим повреждениям и относительно склонная к окаменению, часто сохраняет внутренние детали, что позволяет гистологию детально изучать и рост чешуи. Чешуйки представляют собой нерастущую «коронку», состоящую из дентина , с иногда орнаментированной эмалоидной верхней поверхностью и аспидиновым основанием. [5] Его растущая основа состоит из бесклеточной кости, в которой иногда образуются структуры крепления, фиксирующие ее в боку рыбы. [6] Помимо этого, по-видимому, существует пять типов роста костей, которые могут представлять собой пять естественных групп внутри телодонтов — или спектр, варьирующийся между конечными членами мета- (или орто-) дентина и мезодентина. [7] Каждая из пяти чешуйчатых морф, по-видимому, напоминает чешую более продвинутых групп рыб, что позволяет предположить, что группы телодонтов могли быть стволовыми группами последующих клад рыб. [6]
Однако использование только морфологии чешуи для различения видов имеет некоторые подводные камни. В каждом организме форма чешуи сильно варьируется в зависимости от площади тела. [8] при этом промежуточные формы появляются между разными областями - и, что еще хуже, морфология чешуи может даже не быть постоянной в пределах одной области. Еще больше запутывает ситуацию то, что морфология чешуи не уникальна для таксонов и может быть неотличима на одной и той же территории у двух разных видов. [9]
Морфология и гистология телодонтов являются основным инструментом для количественной оценки их разнообразия и различения видов, хотя в конечном итоге использование таких конвергентных признаков подвержено ошибкам. Тем не менее, на основе морфологии и гистологии чешуи была предложена структура, состоящая из трех групп. [7] Сравнение с современными видами акул показало, что чешуя телодонтов функционально аналогична чешуе современных хрящевых рыб, а также позволило провести обширное сравнение между экологическими нишами. [10]
Космоидные весы
[ редактировать ]
Космоидные чешуи встречаются только у древних лопастных рыб , включая некоторых из самых ранних двоякодышащих рыб (подкласс Dipnoi ), и у Crossopterygii , включая живую латимерию в модифицированной форме (см. эласмоидные чешуи ниже). Вероятно, они произошли от слияния плакоидно-ганоидных чешуек. Внутренняя часть чешуи состоит из плотной пластинчатой кости, называемой изопедином. Поверх него лежит слой губчатой или сосудистой кости, снабженный кровеносными сосудами, за которым следует сложный дентиноподобный слой, называемый космином, с поверхностным внешним покрытием из витродентина . Верхняя поверхность кератиновая . Космоидные чешуи увеличиваются в размерах за счет роста пластинчатого костного слоя. [11]
Эласмоидные чешуйки
[ редактировать ]
Эласмоидные чешуи представляют собой тонкие, черепитчатые чешуйки, состоящие из слоя плотной пластинчатой коллагеновой кости, называемой изопедином, над которым находится слой бугорков, обычно состоящих из кости, как у Eusthenopteron . Слой дентина, имевшийся у первых лопастно-перых рыб, обычно редуцирован, как у современных целакантов , или вовсе отсутствовал, как у современных двоякодышащих рыб и у девонских Eusthenopteron . [12] Эласмоидная чешуя появлялась несколько раз в ходе эволюции рыб. Они присутствуют у некоторых лопастных рыб , таких как все современные и некоторые вымершие двоякодышащие рыбы , а также у латимерий, которые имеют модифицированную космоидную чешую, в которой отсутствует космин и которая тоньше настоящей космоидной чешуи. Они также присутствуют у некоторых тетраподоморфов, таких как Eusthenopteron , амииды и костистые кости, чьи циклоидные и ктеноидные чешуи представляют собой наименее минерализованные эласмоидные чешуи.
Эластоидная чешуя рыбок данио используется в лаборатории для изучения процесса минерализации костей и может культивироваться (храниться) вне организма. [13] [14]
Ганоидные чешуйки
[ редактировать ]

Ганоидные чешуи встречаются у осетровых , веслоносов , щук , боуфинов и бичиров . Они произошли от космоидных чешуек и часто имеют зазубренные края. Они покрыты слоем твердого эмалевоподобного дентина вместо космина и слоем неорганической костной соли, называемой ганоином, вместо витродентина .
Ганоин является характерным компонентом ганоидной чешуи. Это стекловидная, часто многослойная минерализованная ткань , покрывающая чешую, а также черепные кости и плавниковые лучи у некоторых некостистых лучепёрых рыб . [15] такие как гары , бичиры и целаканты . [16] [17] Он состоит из стержневидных кристаллитов апатита . [18] Ганойн — древняя особенность лучепёрых рыб, обнаруженная, например, на чешуе группы стеблей актиноптеригианской Cheirolepis . [17] Хотя ганоин или ганоиноподобные ткани часто считаются синапоморфным признаком лучепёрых рыб, они также встречаются на вымерших акантодиях . [17] Было высказано предположение, что ганоин зубной гомологичен эмали позвоночных. [15] или даже считается разновидностью эмали. [18]
 Амблиптерус полосатый | Ганоидные чешуи вымершей каменноугольной рыбы Amblypterus striatus . (а) показана внешняя поверхность четырех чешуек, а (б) показана внутренняя поверхность двух чешуек. Каждая из ганоидных чешуек Amblypterus ромбовидной формы имеет выступ на внутренней поверхности, который на одном конце превращается в выступающий штифт, который входит в выемку на следующей чешуе, подобно тому, как черепицы соединяются друг с другом на крыше. дома. |  |
Большинство ганоидных чешуек имеют ромбовидную (ромбовидную) форму и соединены шарнирными соединениями. Они обычно толстые и больше похожи на мозаику, чем накладываются друг на друга, как другие чешуйки. [19] Таким образом, чешуя ганоидов практически непроницаема и является отличной защитой от хищников.

 есть У осетра ряды ганоидных чешуек, увеличенных в щитковидные панцирные пластины.
есть У осетра ряды ганоидных чешуек, увеличенных в щитковидные панцирные пластины. Ганоидные чешуи на боуфине уменьшены в размерах и напоминают циклоидные чешуи .
Ганоидные чешуи на боуфине уменьшены в размерах и напоминают циклоидные чешуи .




У осетровых чешуя сильно увеличена в панцирные пластинки по бокам и спине, тогда как у боуфиновых чешуя сильно уменьшена в толщине и напоминает циклоидную чешую .
 Серьги из ганоидной чешуи аллигатора.
Серьги из ганоидной чешуи аллигатора. Ископаемое примитивного лучевого плавника с ганоидной чешуей
Ископаемое примитивного лучевого плавника с ганоидной чешуей Ганоидные чешуйки на окаменелом лепидоте , ок. 130 млн лет назад
Ганоидные чешуйки на окаменелом лепидоте , ок. 130 млн лет назад



Коренные американцы и жители Карибского бассейна использовали жесткую ганоидную чешую аллигатора для изготовления наконечников стрел, нагрудников и в качестве защиты для плугов. В настоящее время из этих чешуек изготавливают украшения. [20]
Лептоидные чешуйки
[ редактировать ]Лептоидные (костно-гребневые) чешуи встречаются у костистых рыб высшего порядка, костистых рыб (более развитая группа лучепёрых рыб). Внешняя часть этих чешуек разветвлена с костными гребнями, а внутренняя часть покрыта волокнистой соединительной тканью. Лептоидные чешуйки тоньше и прозрачнее, чем другие типы чешуек, и не имеют затвердевших слоев эмали или дентина. В отличие от чешуи ганоида, по мере роста рыбы дополнительные чешуи добавляются концентрическими слоями. [21]
Чешуйки лептоидов перекрываются в конфигурации «от головы к хвосту», как черепица на крыше, что делает их более гибкими, чем чешуи космоидов и ганоидов. Такое расположение обеспечивает более плавный поток воды по корпусу и снижает сопротивление . [22] На чешуе некоторых видов наблюдаются полосы неравномерного сезонного роста, называемые кольцами (единственное кольцо ). Эти ленты можно использовать для старения рыбы .
Лептоидные чешуи бывают двух форм: циклоидная (гладкая) и ктеноидная (гребневидная). [23]
Циклоидные весы
[ редактировать ]Циклоидные (круглые) чешуи имеют гладкую текстуру, однородные, с гладким наружным краем или краем. Чаще всего они встречаются у рыб с мягкими лучами в плавниках, таких как лосось и карп .
 |  |
У азиатской арованы большие циклоидные чешуи, расположенные на рыбе мозаикой из приподнятых ребер (слева). Сами чешуйки покрыты нежным сетчатым узором (справа). [24] [25] | |


Циклоидная (круглая) чешуя обычно встречается у карпообразных или лососевых рыб. |





Ктеноидные чешуи
[ редактировать ]Ктеноидные (зубчатые) чешуи похожи на циклоидные чешуи, за исключением того, что у них есть маленькие зубцы или шипики, называемые ктениями, вдоль их внешнего или заднего края. Из-за этих зубов чешуя имеет грубую текстуру. Обычно они встречаются у рыб с колючими плавниками, например у окуневых рыб. Эти чешуйки почти не содержат кости и состоят из поверхностного слоя, содержащего гидроксиапатит и карбонат кальция , и более глубокого слоя, состоящего в основном из коллагена . Эмаль чешуек других типов редуцирована до поверхностных гребней и ктений.


 |  |
Размер зубов на ктеноидных чешуях может варьироваться в зависимости от положения, как показывают чешуи гремучего хвоста Cetonurus crassiceps . | |
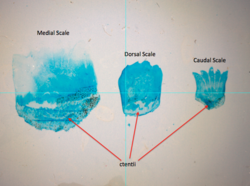

Ктеноидная (зубчатая) чешуя обычно встречается у окуневых рыб. |





Ктеноидные чешуйки, как и другие эпидермальные структуры, происходят из плакод , а отличительная клеточная дифференциация делает их исключительными из других структур, возникающих из покровов . [27] Развитие начинается возле хвостового плавника , по боковой линии рыбы. [28] Процесс развития начинается с скопления фибробластов между эпидермисом и дермой . [27] Коллагеновые фибриллы начинают самоорганизовываться в дермальном слое, что приводит к инициации минерализации . [27] Сначала растет окружность чешуек, а затем толщина, когда перекрывающиеся слои минерализуются вместе. [27]
Ктеноидные чешуи можно разделить на три типа:
- Городчатые чешуи, края которых имеют углубления и выступы.
- Спиноидные чешуи, где чешуя имеет шипы, продолжающие саму чешую.
- Настоящие ктеноидные чешуи, у которых шипы на чешуе представляют собой отдельные структуры.
Большинство лучепёрых рыб имеют ктеноидную чешую. Некоторые виды камбал имеют ктеноидную чешую на глазной стороне и циклоидную чешую на слепой стороне, тогда как у других видов ктеноидная чешуя у самцов и циклоидная чешуя у самок.
Отражение
[ редактировать ]


Многие костистые рыбы покрыты чешуей с высокой отражающей способностью, которые действуют как маленькие зеркала и создают вид посеребренного стекла. Отражение через серебрение широко распространено или доминирует у рыб открытого моря, особенно у тех, которые обитают на глубине 100 метров. Эффекта прозрачности можно добиться за счет серебрения, придающего телу животного высокую отражающую способность. На средних глубинах в море свет падает сверху, поэтому зеркало, ориентированное вертикально, делает животных, например рыб, невидимыми сбоку. [29]
Морской топорик чрезвычайно сплющен с боков (из стороны в сторону), оставляя тело толщиной всего несколько миллиметров, а тело настолько серебристое, что напоминает алюминиевую фольгу . Зеркала состоят из микроскопических структур, подобных тем, которые используются для структурной окраски : стопки из 5-10 кристаллов гуанина, расположенных на расстоянии примерно ¼ длины волны друг от друга, что позволяет конструктивно интерферировать и достигать почти 100-процентного отражения. В глубоких водах, где обитает рыба-топорик, только синий свет с длиной волны 500 нанометров просачивается вниз и должен отражаться, поэтому зеркала, расположенные на расстоянии 125 нанометров друг от друга, обеспечивают хорошую маскировку. [29]
Большинство рыб в верхних слоях океана замаскированы серебрением. У рыб, таких как сельдь , которая обитает на мелководье, зеркала должны отражать смесь волн, и, соответственно, рыба имеет стопки кристаллов с разными расстояниями между ними. Еще одна сложность для рыб с закругленным в поперечном сечении телом заключается в том, что зеркала будут неэффективны, если их положить плоско на кожу, поскольку они не смогут отражать горизонтально. Общий зеркальный эффект достигается за счет множества небольших отражателей, ориентированных вертикально. [29]
Рыбья чешуя, обладающая этими свойствами, используется в некоторых косметических средствах, поскольку способна придавать мерцающий эффект макияжу и помаде. [30]
Плакоидные чешуи
[ редактировать ]
Плакоидные (заостренные, зубчатые) чешуи встречаются у хрящевых рыб : акул , скатов . Их еще называют дермальными зубчиками . Плакоидные чешуйки структурно гомологичны зубам позвоночных («дентикул» переводится как «маленький зуб» ) , имеют центральную полость пульпы, снабженную кровеносными сосудами , окруженную коническим слоем дентина , каждый из которых расположен поверх прямоугольной базальной пластинки, которая лежит на дерме . Внешний слой состоит из витродентина , в значительной степени неорганического вещества, похожего на эмаль . Плакоидная чешуя не может увеличиваться в размерах, а по мере увеличения размера рыбы увеличивается ее количество.
Подобные чешуйки можно найти и под головой зубчатой сельди . Степень покрытия чешуи у лучей гораздо меньше.
Предполагается, что ромбовидные чешуи со свойствами как плакоидных, так и ганоидных чешуек существуют у современных предков челюстных рыб: бесчелюстных остракодерм , а затем и челюстных плакодерм .
Кожа акулы
[ редактировать ]
Кожа акулы почти полностью покрыта мелкой плакоидной чешуей. Чешуя поддерживается шипами, которые кажутся грубыми при движении назад, но когда они сплющиваются под действием движения воды вперед, они создают крошечные вихри , которые уменьшают гидродинамическое сопротивление и уменьшают турбулентность , что делает плавание более эффективным и тихим по сравнению с плаванием костистых животных. Рыбы. [31] Он также защищает от обрастания, демонстрируя эффект лотоса . [32]
Все зубцы состоят из внутренней полости пульпы с нервным и артериальным кровоснабжением, расположенным в дерме и обеспечивающим снабжение зубца слизью. [33] Зубцы содержат ребрышки, выступающие из поверхности чешуи; под микроскопом этот ребрышек может выглядеть как крючок или выступы, выходящие из чешуи. Общая форма выступа зубца зависит от типа акулы и обычно может быть описана двумя способами. [34] Первый представляет собой шкалу, в которой гребни расположены сбоку от акулы и параллельно потоку воды. Вторая форма представляет собой гладкую чешуйку с чем-то похожим на крючковатый выступ, выходящий из поверхности и направленный к задней стороне акулы. [34] Обе формы ребрышек способствуют созданию турбулентного пограничного слоя, отталкивающего ламинарный поток дальше от кожи акулы. [35]
В отличие от костистых рыб, акулы имеют сложный кожный корсет, состоящий из гибких коллагеновых волокон , расположенных в виде спиральной сети, окружающей их тело. Корсет работает как внешний скелет, обеспечивая крепление плавательных мышц и тем самым экономя энергию. [36] В зависимости от положения этих плакоидных чешуек на теле они могут быть гибкими и пассивно подниматься, что позволяет им менять угол атаки. Эти чешуйки также имеют ребрышки, которые ориентированы в направлении потока. Эти ребрышки уменьшают силу сопротивления, действующую на кожу акулы, отталкивая вихрь дальше от поверхности кожи, препятствуя любому высокоскоростному поперечному потоку. [37]
Морфология чешуи
[ редактировать ]Общая анатомия чешуек различается, но всех их можно разделить на три части: темя, шейка и основание. Податливость шкалы связана с размером основания шкалы. Чешуйки с более высокой гибкостью имеют меньшее основание и, следовательно, менее прочно прикреплены к слабому слою. На темени быстроплавающих акул имеется ряд параллельных ребрышек или гребней, идущих спереди назад. [38]
Анализируя три компонента чешуи, можно сделать вывод, что основание зубца не контактирует ни с какой частью потока жидкости. [39] Однако коронка и шейка зубчиков играют ключевую роль и ответственны за создание турбулентных вихрей и водоворотов, возникающих у поверхности кожи. [39] Поскольку зубчики бывают самых разных форм и размеров, можно ожидать, что не все формы будут создавать одинаковый тип турбулентного потока . Во время недавнего исследовательского эксперимента биомиметические образцы зубцов акул с серповидной микроструктурой были протестированы в резервуаре с водой с использованием тягового стола в качестве предметного стекла. Эксперимент показал, что сопротивление поверхности с зубчиками в целом снизилось на 10% по сравнению с гладким образцом. Причиной такого уменьшения сопротивления было то, что турбулентные вихри захватывались между зубчиками, создавая «подушкообразный» барьер против ламинарного потока. [40] Эксперимент того же типа был проведен другой исследовательской группой, которая применила больше вариаций в своем биомиметическом образце. Вторая группа пришла к тому же выводу, что и первая. Однако, поскольку их эксперимент содержал больше вариаций внутри образцов, они смогли достичь высокой степени экспериментальной точности. В заключение они заявили, что более практичные формы более долговечны, чем формы со сложными ребристыми линиями. Практичные формы были низкопрофильными, имели трапециевидное или полукруглое поперечное сечение, напоминающее желоб, и были менее эффективными, но, тем не менее, снижали сопротивление на 6 или 7%. [41]
Уменьшение сопротивления
[ редактировать ]

Акулы уменьшают сопротивление и общую стоимость транспортировки (COT) по нескольким направлениям. Сопротивление давления создается за счет разницы давления между передней и задней сторонами акулы из-за количества объема, который проходит мимо акулы, чтобы двигаться вперед. [42] Этот тип сопротивления также прямо пропорционален ламинарному потоку . Когда ламинарный поток вокруг рыбы увеличивается, увеличивается и сопротивление давления. [43] Сопротивление трения является результатом взаимодействия жидкости с кожей акулы и может варьироваться в зависимости от того, как изменяется пограничный слой на поверхности рыбы. [42]
Рибреты препятствуют поперечному перемещению продольных вихрей в вязком подслое. Механизм сложен и еще до конца не изучен. По сути, ребрышки препятствуют образованию вихрей у поверхности, поскольку вихрь не может поместиться в впадинах, образованных ребрышками. Это толкает вихрь дальше от поверхности, взаимодействуя только с кончиками ребрышек, не вызывая высокоскоростного потока во впадинах. Поскольку этот высокоскоростной поток теперь взаимодействует только с кончиком ребрышки, который представляет собой очень небольшую площадь поверхности, передача импульса, вызывающая сопротивление, теперь намного ниже, чем раньше, тем самым эффективно уменьшая сопротивление. Кроме того, это уменьшает колебания скорости поперечного потока, что также способствует передаче импульса. [38]
Недавние исследования показали, что в пристеночном пограничном слое существует режим до и после пробоя, когда подслой утолщается с уменьшающейся скоростью, а затем резко разрушается на турбулентные вихри, прежде чем окончательно схлопнуться. Эта система полностью саморегулируется и обеспечивает цикл роста и упадка; вихри накапливаются в период роста и внезапно превращаются в массивы шпилек Струхаля, отрывающиеся от стенки. Подъемные вихри — это то, что выталкивает пограничный слой от поверхности акулы, что приводит к уменьшению общего сопротивления, испытываемого рыбой. [44]
Техническое применение
[ редактировать ]Грубая текстура кожи акулы и ската, напоминающая наждачную бумагу , в сочетании с ее прочностью привела к тому, что ее стали ценить как источник сыромятной кожи , называемой шагренью . Одним из многих исторических применений акульей шагрени было изготовление рукояток для мечей . Грубая текстура кожи также используется в японской кухне для изготовления терок, называемых орошики , путем прикрепления кусочков акульей кожи к деревянным доскам. Небольшой размер весов позволяет очень мелко натирать продукты.

В морской отрасли существует чрезвычайно большой рынок и потребность в необрастающих поверхностях . С точки зрения непрофессионала, загрязнение — это процесс, при котором что-то покрывается коркой материалов из окружающей среды, таких как ракушки , водоросли и зеленый ил . Кожные зубцы являются чрезвычайно многообещающей областью исследований для такого типа применения, поскольку акулы являются одной из немногих рыб, у которых нет наростов или роста чешуи. Исследования ВМС США показали, что если можно будет разработать биомиметический материал, это потенциально может привести к экономии затрат на топливо для военных кораблей до 45%. [45]
Существует множество примеров биомиметических материалов и поверхностей, основанных на строении водных организмов, в том числе акул. Такие приложения призваны обеспечить более эффективное движение через жидкие среды, такие как воздух, вода и масло.
Поверхности, имитирующие кожу акул, также использовались для предотвращения попадания микроорганизмов и водорослей на корпуса подводных лодок и кораблей. Один сорт продается как « шарклет ». [46] [47]
Многие новые методы репликации кожи акулы предполагают использование полидиметилсилоксана (ПДМС) для создания формы. Обычно процесс включает в себя взятие плоского куска акульей кожи, покрытие его ПДМС для формирования формы и повторное заливание ПДМС в эту форму, чтобы получить копию кожи акулы. Этот метод был использован для создания биомиметической поверхности, обладающей супергидрофобными свойствами и проявляющей эффект лотоса . [46] Одно исследование показало, что эти биомиметические поверхности снижают сопротивление до 9%. [37] при маховом движении снижение сопротивления достигало 12,3%. [48]
Зубцы также обеспечивают снижение сопротивления объектов, основная форма сопротивления которых вызвана турбулентным потоком на поверхности. Большая часть общего сопротивления длинных объектов с относительно плоскими сторонами обычно возникает из-за турбулентности у стенки, поэтому ребрышки будут иметь заметный эффект. Наряду с морскими применениями, аэрокосмическая промышленность может получить большую выгоду от этих биомиметических разработок. Другие области применения включают трубы, где они придают внутренней поверхности шероховатость, напоминающую шероховатость, и обнаруживают снижение сопротивления на 5%, а снижение на несколько процентов заявлено для конкурентоспособных купальных костюмов. [49]
Параметрическое моделирование было выполнено на зубах акулы с широким спектром конструктивных вариантов, таких как генераторы вихрей низкого и высокого профиля. [50] С помощью этого метода была выполнена наиболее тщательная характеристика симметричных двумерных ребрышек с пилообразным, фестончатым и лопастным поперечным сечением. [49] Эти биомиметические модели были разработаны и проанализированы, чтобы увидеть последствия применения зубцеподобных структур на крыльях различных самолетов. В ходе моделирования было отмечено, что образец изменил реакцию на низкие и высокие углы атаки . Как геометрия зубцов, так и их расположение оказывают глубокое влияние на аэродинамические характеристики аэродинамических профилей. Из протестированных как низкопрофильных, так и высокопрофильных образцов низкопрофильные вихревые генераторы превзошли нынешние конструкции с гладким крылом на 323%. Такое увеличение производительности происходит за счет разделительного пузыря в следе за зубчиком и продольных вихрей, которые восполняют импульс, потерянный в пограничном слое из-за поверхностного трения. [50]
Щиты
[ редактировать ]
Щитки похожи на чешуи и выполняют ту же функцию. В отличие от чешуи рыб, которая формируется из эпидермиса, щитки образуются в нижнем сосудистом слое кожи, а эпидермальным элементом является только верхняя поверхность. Формируясь в живой дерме, щитки образуют роговой внешний слой, внешне похожий на чешуйчатый.
Scute происходит от латинского слова « щит» и может принимать форму:
- внешняя щитовидная костная пластина или
- видоизмененная, утолщенная чешуя, часто килеватая или колючая, или
- выступающая видоизмененная (грубая и сильно ребристая) чешуя, обычно приуроченная к боковой линии, или на хвостовом стебле, образующая хвостовые кили, или вдоль вентрального профиля.
Некоторые рыбы, например , сосновая шишка , полностью или частично покрыты щитками. Речные сельди и плавники имеют брюшной ряд щитков, представляющих собой чешуйки с приподнятыми острыми концами, служащие для защиты. У некоторых гнездов есть ряд жучков, идущих по боковой линии с обеих сторон.
Масштабное развитие
[ редактировать ]Чешуя обычно появляется на поздних стадиях развития рыб. В случае с рыбками данио проходит 30 дней после оплодотворения, прежде чем различные слои, необходимые для формирования чешуи, дифференцируются и организуются. Для этого необходимо, чтобы произошла консолидация мезенхимы , затем индуцировался морфогенез и, наконец, произошел процесс дифференцировки или позднего метаморфоза . [51] [52]
- Консолидация мезенхимы. Консолидация или структурирование мезенхимы возникает во время развития дермы . Этот процесс зависит от того, хрящевая рыба или костистая. У хрящевых рыб структурирование происходит за счет образования двух слоев. Первый поверхностный и широкий, второй тонкий и компактный. Эти два слоя разделены мезенхимальными клетками . Костные рыбы образуют бесклеточный субстрат, организованный перпендикулярно коллагеновыми волокнами . В дальнейшем у обеих рыб фибробласты удлиняются. Они проникают в компактный слой мезенхимы, который консолидируется до образования чешуек, чтобы инициировать дермальную пластинку. [51] [52] [53]
- Индукция морфогенеза: Морфогенез обусловлен образованием эпидермального сосочка , который образуется путем соединения эпидермиса и дермы в процессе инвагинации . Морфогенез начинается с момента перемещения фибробластов в верхнюю часть компактной мезенхимы. В ходе этого процесса базальные клетки эпителия образуют отграничительный слой, который располагается в верхней части мезенхимы. В дальнейшем эти клетки будут дифференцироваться в том месте, где зачаток чешуи . возникнет [51] [52] [53]
- Дифференциация или поздний метаморфоз: эта дифференциация возникает в двух разных формах в зависимости от типа формируемой чешуи. Образование эласмоидных чешуек (циклоидов и ктеноидов) происходит за счет образования пространства между матриксом эпидермального сосочка. Это пространство содержит коллагеновые волокна. Вокруг этого пространства дифференцируются эластобласты, отвечающие за выработку необходимого материала для образования чешуек. Впоследствии происходит матричная минерализация , позволяющая окалине приобрести жесткую характеристику, которая ее идентифицирует. [51] [52] [53]
В отличие от эласмоидных чешуек, ганоидные чешуйки состоят из минерализованного и неминерализованного коллагена в разных регионах. Образование их происходит за счет проникновения поверхностных клеток мезенхимы в матрикс, последний состоит из коллагеновых волокон и располагается вокруг сосудистых капилляров, давая начало сосудистым полостям. На этом этапе эластобласты заменяются остеобластами , образуя кость. участки матрикса чешуи Неокостеневшие состоят из уплотненного коллагена, что позволяет ему сохранять соединение с мезенхимой. Они известны как волокна Шарпи . [51] [52] [53]
Одним из генов, регулирующих развитие чешуйообразования у рыб, является ген звукового ежа (shh), который посредством белка (shh), участвующего в органогенезе и в процессе клеточной коммуникации , обеспечивает образование чешуи. [54] [55] Аполипопротеин E (ApoE), который обеспечивает транспортировку и метаболизм триглицеридов и холестерина , взаимодействует с shh, поскольку ApoE обеспечивает холестерин для сигнального пути shh . Показано, что в процессе клеток дифференцировки и взаимодействия уровень транскрипции АроЕ высок, что позволило сделать вывод о важности этого белка для позднего развития чешуи. [54] [55]
Модифицированные весы
[ редактировать ]



Различные группы рыб развили ряд модифицированных чешуек, выполняющих различные функции.
- Почти у всех рыб есть боковая линия — система механорецепторов , улавливающих движение воды. У костистых рыб чешуя вдоль боковой линии имеет центральные поры, которые позволяют воде контактировать с чувствительными клетками.
- Шипы спинных плавников акул-собачек и химер, жалящие шипы хвостов скатов и «пилильные» зубы рыб-пил и акул-пил представляют собой сросшиеся и видоизмененные плакоидные чешуи.
- У рыб-хирургов имеется лезвие, напоминающее скальпель, представляющее собой видоизмененную чешуйку по обе стороны от хвостового стебля . [56]
- У некоторых сельдей , анчоусов и полуклювов есть лиственная чешуя , которая легко сбрасывается и помогает спастись от хищников.
- Самцы дротиков Percina имеют ряд увеличенных кадудообразных чешуек между брюшными плавниками и анальным отверстием .
- У рыб-дикобразов чешуя видоизменилась в большие внешние шипы .
- Напротив, у рыбы-фугу более тонкие и более скрытые шипы, чем у рыбы-дикобраза, которые становятся видимыми только тогда, когда рыба надувается. В отличие от рыбы-дикобраза, эти шипы не представляют собой модифицированную чешую, а развиваются под контролем той же сети генов, которая производит перья и волосы у других позвоночных. [57] [58]
 У рыбы-дикобраза чешуя превратилась в шипы .
У рыбы-дикобраза чешуя превратилась в шипы .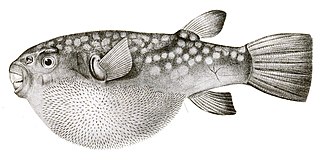 Шипы рыбы фугу не представляют собой модифицированную чешую, а развиваются независимой генной сетью.
Шипы рыбы фугу не представляют собой модифицированную чешую, а развиваются независимой генной сетью.


Рыба без чешуи
[ редактировать ] У мандаринок нет чешуи, и они защищаются слоем вонючей и горькой слизи.
У мандаринок нет чешуи, и они защищаются слоем вонючей и горькой слизи.

У рыб без чешуи обычно появляются альтернативы защитной чешуе, такие как жесткая кожистая кожа или костные пластины.
- У бесчелюстных рыб ( миноги и миксины ) кожа гладкая, без чешуи и кожных костей. [59] Минога получает некоторую защиту от жесткой кожистой кожи. Миксы выделяют обильное количество слизи или слизи , если им угрожает опасность. [60] Они могут завязать себя в узел , соскребая на ходу слизь и освобождаясь от хищника. [61]
- Большинство угрей не имеют чешуи, хотя некоторые виды покрыты крошечной гладкой циклоидной чешуей.
- У большинства сомов нет чешуи, хотя у некоторых семейств имеется бронежилет в виде кожных пластин или своего рода щитка. [62]
- У мандаринок нет чешуи, и у них есть слой вонючей и горькой слизи, который блокирует болезни и, вероятно, отпугивает хищников, подразумевая, что их яркая окраска апосематическая . [63]
- У удильщиков рыхлая, тонкая кожа, часто покрытая мелкими раздвоенными кожными колючками или бугорками , но у них нет правильной чешуи. Они полагаются на камуфляж, чтобы избежать внимания хищников, а их дряблая кожа мешает хищникам их схватить.
Многие группы костных рыб, включая игл , морских коньков , самцов , браконьеров и несколько семейств колюшек , развили внешние костные пластинки, структурно напоминающие плакоидные чешуйки, в качестве защитной брони от хищников.
- У морских коньков нет чешуи, но у них тонкая кожа, натянутая на костяную пластинчатую броню, расположенную кольцами по всей длине тела.
- У самцов пластины сливаются вместе, образуя жесткий панцирь или экзоскелет, охватывающий все тело. Эти костные пластины представляют собой не видоизмененные чешуйки, а окостеневшую кожу . Из-за этой тяжелой брони кузовок ограничен в медленных движениях, но немногие другие рыбы способны съесть взрослых особей.
 | |
Угри кажутся бесчешуйными, но некоторые виды покрыты крошечной гладкой циклоидной чешуей. | |
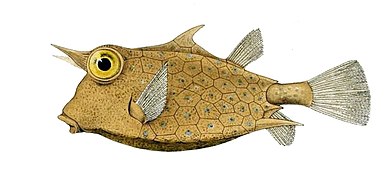 Кузовок имеет пластинки окостеневшей кожи, сросшиеся вместе и образующие твердый панцирь.
Кузовок имеет пластинки окостеневшей кожи, сросшиеся вместе и образующие твердый панцирь. У морских коньков тонкая кожа, натянутая на костные пластины, расположенные кольцами.
У морских коньков тонкая кожа, натянутая на костные пластины, расположенные кольцами.


Некоторые рыбы, такие как хоки и рыба-меч, рождаются с чешуей, но сбрасывают ее по мере роста.
Файлфиш имеет грубую, неперекрывающуюся чешую с небольшими шипами, поэтому его называют филефишом. Некоторые рыбы-филиалы кажутся бесчешуйными, потому что их чешуя очень мала.
Заметная чешуйка появляется у тунца только вдоль боковой линии и на панцире — защитной полосе из утолщенных и увеличенных чешуек в области плеч. Большая часть тела тунца покрыта чешуей настолько мелкой, что при случайном осмотре она кажется бесчешуйной. [64]
 Некоторые рыбки кажутся бесчешуйными, потому что их чешуя очень мала.
Некоторые рыбки кажутся бесчешуйными, потому что их чешуя очень мала. При беглом осмотре кажется, что тунец практически лишен чешуи, но это не так.
При беглом осмотре кажется, что тунец практически лишен чешуи, но это не так.


Левит
[ редактировать ]В книге Левит говорится, что «из всего, что находится в водах… в морях и реках», те, у которых нет ни плавников, ни чешуи, «будут мерзостью для вас» и их нельзя есть. [65] Это устраняет всех водных беспозвоночных как мерзостей и нечистых , а также любую рыбу, у которой нет чешуи (похоже, не существует рыб, у которых нет плавников).
Согласно чоку или божественным указам Торы и Талмуда , чтобы рыба была объявлена кошерной , она должна иметь чешую и плавники. [66] Определение «чешуя» отличается от определений, представленных в биологии, тем, что чешуя кошерной рыбы должна быть видна глазу и ее можно легко снять с кожи вручную или ножом для очистки чешуи. [66] По данным агентства по сертификации кошерности Православного союза , рыба является кошерной, если чешую можно снять, не разрывая кожицы. [67] Так, карп и лосось кошерны, тогда как акула, чешуя которой микроскопична, и осетр, чьи щитки невозможно легко удалить, не вырезав их из тела, — все они некошерны. К другим некошерным рыбам относятся сом, угри, пресноводная треска , змеиная скумбрия и рыба фугу. [66]
Лепидофагия
[ редактировать ]
Лепидофагия (по-древнегречески «поедание чешуи» ) — это особое пищевое поведение рыб, которое включает в себя поедание чешуи других рыб. [69] Лепидофагия независимо развилась как минимум в пяти пресноводных семействах и семи морских семействах. [70]
Рыбья чешуя может быть питательной: помимо слоев кератина и эмали она содержит кожную часть и слой богатой белком слизи . Они являются богатым источником фосфата кальция . [70] Однако соотношение энергии, затрачиваемой на удар, по сравнению с количеством чешуи, потребляемой за удар, накладывает ограничение на размер рыб-лепидофагов, и они обычно намного меньше своей добычи . [70] Масштабное пищевое поведение обычно развивается из-за нехватки еды и экстремальных условий окружающей среды. Поедание чешуи и кожи вокруг чешуи обеспечивает богатые белком питательные вещества, которые могут быть недоступны в других местах этой ниши. [71]
Рыбьи челюсти обычно демонстрируют двустороннюю симметрию . Исключением является чешуйчатая цихлида Perissodus microlepis . Челюсти этой рыбы встречаются в двух различных морфологических формах. У одного морфа челюсть повернута влево, что позволяет ему с большей готовностью поедать чешую на правом боку жертвы. У другого морфа челюсть повернута вправо, что облегчает поедание чешуи на левом боку жертвы. Относительная численность двух морфов в популяциях регулируется частотно-зависимым отбором . [68] [72] [73]
См. также
[ редактировать ]- Определение возраста у рыб
- Окраска животных
- Отражатели для животных
- Фотонные кристаллы
- Чешуя рептилии
- Масштаб (зоология)
- Чешуйчатая броня
- Змеиная чешуя
- Урокотори – японская рыбочистка.
Ссылки
[ редактировать ]- ^ Масштаб Этимонлин . Проверено 28 апреля 2019 г.
- ^ Монгера, А.; Нюсляйн-Фольхард, К. (2013). «Чешуя рыб возникает из мезодермы» . Современная биология . 23 (9): Р338–Р339. дои : 10.1016/j.cub.2013.02.056 . ПМИД 23660349 .
- ^ Шарп, ПТ (2001). «Развитие рыбьей чешуи: волосы сегодня, зубы и чешуя вчера?» . Современная биология . 11 (18): Р751–Р752. дои : 10.1016/S0960-9822(01)00438-9 . ПМИД 11566120 . S2CID 18868124 .
- ^ Тернер, С.; Тарлинг, Д.Х. (1982). «Распространение телодонта и других бесчелюстных как тест нижнепалеозойских континентальных реконструкций». Палеогеография, Палеоклиматология, Палеоэкология . 39 (3–4): 295–311. Бибкод : 1982PPP....39..295T . дои : 10.1016/0031-0182(82)90027-X .
- ^ Мерсс, Т. (2006). «Экзоскелетная ультраскульптура ранних позвоночных». Журнал палеонтологии позвоночных . 26 (2): 235–252. doi : 10.1671/0272-4634(2006)26[235:EUOEV]2.0.CO;2 . S2CID 85993241 .
- ^ Перейти обратно: а б Жанвье, Филипп (1998). «Ранние позвоночные и их современные родственники». Ранние позвоночные . Издательство Оксфордского университета . стр. 123–127. ISBN 978-0-19-854047-2 .
- ^ Перейти обратно: а б Тернер, С. (1991). «Монофилия и взаимоотношения Thelodonti». В ММ Чанг; Ю. Х. Лю; Г. Р. Чжан (ред.). Ранние позвоночные и связанные с ними проблемы эволюционной биологии . Наука Пресс, Пекин. стр. 87–119.
- ^ Мерсс, Т. (1986). «Чешуйка бесчелюстного телодонта Флеболеписа ». Журнал палеонтологии позвоночных . 6 (1): 1–11. дои : 10.1080/02724634.1986.10011593 .
- ^ Ботелла, Х.; JI Валенсуэла-Риос; П. Карлс (2006). «Новый телодонт раннего девона из Кельтиберии (Испания) с ревизией испанских телодонтов» . Палеонтология . 49 (1): 141–154. дои : 10.1111/j.1475-4983.2005.00534.x . S2CID 128939911 .
- ^ Феррон, Умберто Г.; Ботелла, Гектор (2017). «Сквамация и экология телодонтов» . ПЛОС ОДИН . 12 (2): e0172781. Бибкод : 2017PLoSO..1272781F . дои : 10.1371/journal.pone.0172781 . ПМК 5328365 . ПМИД 28241029 .
- ^ МАЙКЛ АЛЛАБИ «космоидный масштаб». Зоологический словарь. . Энциклопедия.com. 29 октября 2019 г. < https://www.encyclepedia.com >
- ^ Зильберберг, Л., Менье, Ф.Дж., Лорен, М. (2010). Микроанатомическое и гистологическое исследование посткраниального дермального скелета девонского саркоптерига Eusthenopteron foordi , Acta Palaeontologica Polonica 55: 459–470.
- ^ Берген, Дилан Дж. М.; Каге, Эрика; Хаммонд, Крисси Л. (2019). «Рыба данио как новая модель остеопороза: основная платформа тестирования для скрининга новых остеоактивных соединений» . Границы эндокринологии . 10 :6. дои : 10.3389/fendo.2019.00006 . ISSN 1664-2392 . ПМК 6361756 . ПМИД 30761080 .
- ^ де Вриз, Э.; ван Кессель, MAHJ; Питерс, HM; Пролеты, FAT; Флик, Г.; Мец, младший (1 февраля 2014 г.). «Преднизолон индуцирует остеопороз-подобный фенотип в регенерирующей чешуе рыбки данио». Международный остеопороз . 25 (2): 567–578. дои : 10.1007/s00198-013-2441-3 . ISSN 1433-2965 . ПМИД 23903952 . S2CID 21829206 .
- ^ Перейти обратно: а б Зильберберг, Л.; Сир, Ж.-Ю.; Нанси, А. (1997). «Иммунодетектирование амелогенинподобных белков в ганоине экспериментально регенерирующей чешуи Calamoichthys Calabaricus, примитивной актиноптеригической рыбы» . Анатомическая запись . 249 (1): 86–95. doi : 10.1002/(SICI)1097-0185(199709)249:1<86::AID-AR11>3.0.CO;2-X . ПМИД 9294653 .
- ^ Сир Жан-Ив; Донохью, Филип CJ; Викариус, Мэтьюз К. (2009). «Происхождение и эволюция покровного скелета у позвоночных, не являющихся четвероногими» . Журнал анатомии . 214 (4): 409–440. дои : 10.1111/j.1469-7580.2009.01046.x . ISSN 0021-8782 . ПМЦ 2736117 . ПМИД 19422423 .
- ^ Перейти обратно: а б с Рихтер, М. (1995). «Микроструктурное исследование ганоиновой ткани избранных низших позвоночных». Зоологический журнал Линнеевского общества . 114 (2): 173–212. дои : 10.1006/zjls.1995.0023 .
- ^ Перейти обратно: а б Брюэ, BJF; Сонг, Дж.; Бойс, MC; Ортис, К. (2008). «Принципы проектирования материалов древних рыбьих доспехов». Природные материалы . 7 (9): 748–756. Бибкод : 2008NatMa...7..748B . дои : 10.1038/nmat2231 . ПМИД 18660814 .
- ^ Перейти обратно: а б Шерман, Винсент Р.; Яраги, Николас А.; Кисаилус, Давид; Мейерс, Марк А. (1 декабря 2016 г.). «Микроструктурные и геометрические влияния в защитных чешуях лопаточки Atractosteus» . Журнал интерфейса Королевского общества . 13 (125): 20160595. doi : 10.1098/rsif.2016.0595 . ISSN 1742-5689 . ПМК 5221522 . ПМИД 27974575 .
- ^ «План управления и восстановления аллигатора штата Миссури» (PDF) . Отдел природоохранного рыболовства Департамента штата Миссури. 22 января 2013 г. Архивировано из оригинала (PDF) 6 мая 2016 г. . Проверено 12 апреля 2019 г.
- ^ Лаглер, К.Ф., Дж. Э. Бардах и Р. Р. Миллер (1962) Ихтиология . Нью-Йорк: Джон Уайли и сыновья.
- ^ Баллард, Бонни; Чик, Райан (2 июля 2016 г.). Медицина экзотических животных для ветеринарного техника . Джон Уайли и сыновья. ISBN 978-1-118-92421-1 .
- ^ МакГрутер, Марк (2 декабря 2019 г.). «Циклоидная и ктеноидная чешуя» . Австралийский музей . Проверено 29 декабря 2021 г.
- ^ Пуйо, Л.; Сударто, Гай Г. Тьюгельс (2003). «Разноцветные разновидности азиатской арованы Scleropages formosus (Osteoglossidae) представляют собой отдельные виды: морфологические и генетические доказательства». Цибиум . 27 (4): 287–305.
- ^ Исмаил, М. (1989). Систематика, зоогеография и охрана пресноводных рыб полуострова Малайзия (под ред. Докторской диссертации). Государственный университет Колорадо.
- ^ Э. Дж. Брилл (1953). Рыбы Индо-Австралийского архипелага . Э. Дж. Брилл. стр. 306–307.
- ^ Перейти обратно: а б с д Кавасаки, Кента К., «Генетический анализ морфологии чешуи цихлид» (2016). Магистерские диссертации май 2014 г. – по настоящее время. 425. http://scholarworks.umass.edu/masters_theses_2/425 .
- ^ Хельфман, Джин (2009). Разнообразие биологии, эволюции и экологии рыб . Уайли-Блэквелл.
- ^ Перейти обратно: а б с Сельдь, Питер (2002). Биология глубокого океана . Оксфорд: Издательство Оксфордского университета. стр. 193–195. ISBN 9780198549567 .
- ^ «В твоей помаде, наверное, рыбья чешуя» . HuffPost Индия . 23 апреля 2015 года . Проверено 6 мая 2019 г.
- ^ Мартин, Р. Эйдан. «Кожа зубов» . Проверено 28 августа 2007 г.
- ^ Фюрстнер, Райнер; Бартлотт, Вильгельм; Найнхейс, Кристоф; Уолцель, Питер (1 февраля 2005 г.). «Смачивающие и самоочищающиеся свойства искусственных супергидрофобных поверхностей». Ленгмюр . 21 (3): 956–961. дои : 10.1021/la0401011 . ISSN 0743-7463 . ПМИД 15667174 .
- ^ Лаудер, Джордж В.; Уэйнрайт, Дилан К.; Домель, Август Г.; Уивер, Джеймс К.; Вэнь, Ли; Бертольди, Катя (2016). «Структура, биомиметика и гидродинамика поверхности кожи рыб» . Физический обзор жидкостей . 1 (6): 060502. Бибкод : 2016PhRvF...1f0502L . doi : 10.1103/PhysRevFluids.1.060502 . S2CID 18118663 .
- ^ Перейти обратно: а б Фельд, Катрин; Колборг, Анна Ноер; Нюборг, Камилла Мари; Салевский, Мирко; Стеффенсен, Джон Фленг; Берг-Сёренсен, Кирстин (24 мая 2019 г.). «Кожные зубчики трех медленно плавающих видов акул: микроскопия и визуализация потока» . Биомиметика . 4 (2): 38. doi : 10.3390/biomimetics4020038 . ISSN 2313-7673 . ПМК 6631580 . ПМИД 31137624 .
- ^ Флетчер, Томас; Альтрингем, Джон; Пикалл, Джеффри; Виналл, Пол; Доррелл, Роберт (7 августа 2014 г.). «Гидродинамика ископаемых рыб» . Труды Королевского общества B: Биологические науки . 281 (1788): 20140703. doi : 10.1098/rspb.2014.0703 . ISSN 0962-8452 . ПМК 4083790 . ПМИД 24943377 .
- ^ Мартин, Р. Эйдан. «Как важно быть хрящевым» . Центр исследований акул ReefQuest . Проверено 29 августа 2009 г.
- ^ Перейти обратно: а б Хаге, В.; Брюс, М.; Бехерт, Д.В. (1 мая 2000 г.). «Эксперименты с трехмерными ребрышками как идеализированной моделью акульей кожи». Эксперименты с жидкостями . 28 (5): 403–412. Бибкод : 2000ExFl...28..403B . дои : 10.1007/s003480050400 . ISSN 1432-1114 . S2CID 122574419 .
- ^ Перейти обратно: а б Мотта, Филип; Хабеггер, Мария Лаура; Лэнг, Эми; Хютер, Роберт; Дэвис, Джессика (1 октября 2012 г.). «Морфология и гибкость чешуи короткоплавникового мако Isurus oxyrinchus и черноперой акулы Carcharhinuslimbatus». Журнал морфологии . 273 (10): 1096–1110. дои : 10.1002/Jmor.20047 . ISSN 1097-4687 . ПМИД 22730019 . S2CID 23881820 .
- ^ Перейти обратно: а б Доу, Чжаолян; Ван, Цзядао; Чен, Даронг (1 декабря 2012 г.). «Бионическое исследование рыбьей чешуи для уменьшения сопротивления» . Журнал бионической инженерии . 9 (4): 457–464. дои : 10.1016/S1672-6529(11)60140-6 . ISSN 1672-6529 . S2CID 137143652 .
- ^ «Экспериментальные исследования характеристик снижения сопротивления бионической поверхности с водоудерживающими микроструктурами рыбьей чешуи» (PDF) .
- ^ Палмер, Колин; Янг, Марк Т. (14 января 2015 г.). «Уменьшение поверхностного сопротивления и контроль разделения потоков у пелагических позвоночных, что имеет значение для интерпретации морфологии чешуи ископаемых таксонов» . Королевское общество открытой науки . 2 (1): 140163. Бибкод : 2015RSOS....240163P . дои : 10.1098/rsos.140163 . ISSN 2054-5703 . ПМЦ 4448786 . ПМИД 26064576 .
- ^ Перейти обратно: а б Лаудер, Джордж В.; Уэйнрайт, Дилан К.; Домель, Август Г.; Уивер, Джеймс К.; Вэнь, Ли; Бертольди, Катя (18 октября 2016 г.). «Структура, биомиметика и гидродинамика поверхности кожи рыб» . Физический обзор жидкостей . 1 (6): 060502. Бибкод : 2016PhRvF...1f0502L . doi : 10.1103/PhysRevFluids.1.060502 .
- ^ Мутурамалингам, Мутхукумар; Виллемин, Лео С.; Брюкер, Кристоф (29 апреля 2019 г.). «Формирование полос при обтекании биомиметических массивов рыбьей чешуи». Журнал экспериментальной биологии . 222 (Пт 16): джеб2 arXiv : 1904.12752 . Бибкод : 2019arXiv190412752M дои : 10.1242/jeb.205963 . ПМИД 31375542 . S2CID 139103148 .
- ^ Бандиопадьяй, Промод Р.; Хеллум, Арен М. (23 октября 2014 г.). «Моделирование того, как узоры кожи акулы и дельфина контролируют переходные модели завихренности стенок и турбулентности с использованием механизмов пространственно-временного сброса фазы» . Научные отчеты . 4 : 6650. Бибкод : 2014NatSR...4E6650B . дои : 10.1038/srep06650 . ISSN 2045-2322 . ПМК 4206846 . ПМИД 25338940 .
- ^ Магин, Челси М.; Купер, Скотт П.; Бреннан, Энтони Б. (1 апреля 2010 г.). «Нетоксичные стратегии борьбы с обрастанием» . Материалы сегодня . 13 (4): 36–44. дои : 10.1016/S1369-7021(10)70058-4 . ISSN 1369-7021 .
- ^ Перейти обратно: а б Лю, Юнхун; Ли, Гуанцзи (15 декабря 2012 г.). «Новый метод получения «эффекта лотоса» на биомиметической коже акулы». Журнал коллоидной и интерфейсной науки . 388 (1): 235–242. Бибкод : 2012JCIS..388..235L . дои : 10.1016/j.jcis.2012.08.033 . ISSN 0021-9797 . ПМИД 22995249 .
- ^ «Sharklet Discovery | Sharklet Technologies, Inc» . www.sharklet.com . Проверено 26 сентября 2018 г.
- ^ Лаудер, Джордж В.; Оффнер, Йоханнес (1 марта 2012 г.). «Гидродинамическая функция акульей кожи и два биомиметических применения» . Журнал экспериментальной биологии . 215 (5): 785–795. дои : 10.1242/jeb.063040 . ISSN 1477-9145 . ПМИД 22323201 .
- ^ Перейти обратно: а б Дин, Брайан и Бхушан, Бхарат. (2010). Поверхности из акульей кожи для снижения сопротивления жидкости в турбулентном потоке: обзор. Философские труды. Серия А. Математические, физические и технические науки. 368. 4775-806. 10.1098/рста.2010.0201.
- ^ Перейти обратно: а б Домель, Август Г.; Саадат, Мехди; Уивер, Джеймс К.; Хадж-Харири, Хосейн; Бертольди, Катя; Лаудер, Джордж В. (28 февраля 2018 г.). «Дизайн, вдохновленный кожей акулы, улучшает аэродинамические характеристики» . Журнал интерфейса Королевского общества . 15 (139): 20170828. doi : 10.1098/rsif.2017.0828 . ПМЦ 5832729 . ПМИД 29436512 .
- ^ Перейти обратно: а б с д и Сир, JY; Юисён, ANN (2003). «Формирование кожного скелета и зубных тканей у рыб: сравнительный и эволюционный подход». Биологические обзоры . 78 (2): 219–249. дои : 10.1017/S1464793102006073 . ПМИД 12803422 . S2CID 19556201 .
- ^ Перейти обратно: а б с д и Ле Гельек, Д.; Морван-Дюбуа, Ж.; Сир, JY (2004). «Развитие кожи костистых рыб с особым акцентом на отложение коллагена в дерме рыбок данио ( Danio rerio )» . Международный журнал биологии развития . 48 (2–3): 217–231. дои : 10.1387/ijdb.15272388 . ПМИД 15272388 .
- ^ Перейти обратно: а б с д Сир, JY (2001). «Зубы вне рта у костистых рыб: как извлечь выгоду из ошибки развития». Эволюция и развитие . 3 (2): 104–108. дои : 10.1046/j.1525-142x.2001.003002104.x . ПМИД 11341672 . S2CID 13353402 .
- ^ Перейти обратно: а б Сир, JY; Акименко, М.А. (2003). «Развитие чешуи у рыб: обзор с описанием экспрессии звукового ежа (шш) у рыбок данио ( Danio rerio )» . Международный журнал биологии развития . 48 (2–3): 233–247. дои : 10.1387/ijdb.15272389 . ПМИД 15272389 .
- ^ Перейти обратно: а б Монно, MJ; Бабин, П.Дж.; Полео, Г.; Андре, М.; Лафорест, Л.; Балланьи, К.; Акименко, М.А. (1999). «Эпидермальная экспрессия гена аполипопротеина Е во время развития плавников и чешуи, а также регенерации плавников у рыбок данио» . Динамика развития . 214 (3): 207–215. doi : 10.1002/(SICI)1097-0177(199903)214:3<207::AID-AJA4>3.0.CO;2-5 . ПМИД 10090147 .
- ^ Соренсон, Л.; Сантини, Ф.; Карневейл, Г.; Альфаро, Мэн (2013). «Мультилокусное древо рыб-хирургов (Acanthuridae, Percomorpha) с пересмотренной семейной таксономией». Молекулярная филогенетика и эволюция . 68 (1): 150–160. дои : 10.1016/j.ympev.2013.03.014 . ПМИД 23542000 .
- ↑ Как у рыбы-фугу появились дурацкие шипы Phys.org , 25 июля 2019 г.
- ^ Шоно, Т.; Тьери, AP; Купер, РЛ; Курокава, Д.; Бритц, Р.; Окабе, М.; Фрейзер, Дж.Дж. (2019). «Эволюция и разнообразие развития кожных шипов у рыб-фугу» . iScience . 19 : 1248–1259. Бибкод : 2019iSci...19.1248S . дои : 10.1016/j.isci.2019.06.003 . ПМЦ 6831732 . ПМИД 31353167 .
- ^ Кулидж Э., Хедрик М.С. и Милсом В.К. (2011) «Вентиляционные системы» . В: McKenzie DJ, Farrell AP и Brauner CJ (редакторы) Физиология рыб: примитивные рыбы , Elsevier, стр. 182–213. ISBN 9780080549521
- ^ Ротшильд, Анна (1 апреля 2013 г.). «Слизь миксины: одежда будущего?» . Новости Би-би-си . Проверено 2 апреля 2013 г.
- ^ Йонг, Эд (23 января 2019 г.). «Никто не готов к слизи миксины» . Атлантика . Проверено 26 января 2019 г.
- ^ Фрил, JP; Лундберг, Дж. Г. (1996). « Micromyzon akamai , gen. et sp. nov., маленький безглазый сом-банджо (Siluriformes: Aspredinidae) из речных русел нижнего бассейна Амазонки». Копейя . 1996 (3): 641–648. дои : 10.2307/1447528 . JSTOR 1447528 .
- ^ Садовый Ю.; Рэндалл, Дж. Э.; Расотто, Мария Б. (май 2005 г.). «Структура кожи шести видов дракончиков (Gobiesociformes; Callionymidae): межвидовые различия в типах железистых клеток и секреции слизи». Журнал биологии рыб . 66 (5): 1411–1418. дои : 10.1111/j.0022-1112.2005.00692.x .
- ^ Есть ли у тунца чешуя? Северо-восточный научный центр рыболовства, Рыболовное управление NOAA. По состоянию на 4 августа 2019 г.
- ^ Левит 11: 9–10.
- ^ Перейти обратно: а б с Арье Цитрон, «Все о кошерной рыбе»
- ^ Проверка кошерной рыбы или сертификации кошерности OU . Проверено 9 августа 2019 г.
- ^ Перейти обратно: а б Ли, HJ; Куше, Х.; Мейер, А. (2012). «Ручное добывание пищи у рыб-цихлид, питающихся чешуей: его потенциальная роль в формировании морфологической асимметрии» . ПЛОС ОДИН . 7 (9): e44670. Бибкод : 2012PLoSO...744670L . дои : 10.1371/journal.pone.0044670 . ПМЦ 3435272 . ПМИД 22970282 .
- ^ Фрёзе Р. и Д. Поли. Редакторы. «Глоссарий: Лепидофагия» . ФишБаза . Проверено 12 апреля 2007 г.
{{cite web}}:|author=имеет общее имя ( справка ) - ^ Перейти обратно: а б с Яновец, Джефф (2005). «Функциональная морфология питания у специалиста по поеданию чешуек Catoprion mento » (PDF) . Журнал экспериментальной биологии . 208 (Часть 24): 4757–4768. дои : 10.1242/jeb.01938 . ПМИД 16326957 . S2CID 15566769 .
- ^ Мартин, К.; ПК Уэйнрайт (2011). «Трофическая новизна связана с исключительными темпами морфологического разнообразия двух адаптивных разновидностей куколок карповых карповых» . Эволюция . 65 (8): 2197–2212. дои : 10.1111/j.1558-5646.2011.01294.x . ПМИД 21790569 . S2CID 23695342 .
- ^ Хори, М. (1993). «Частотозависимый естественный отбор в отношении рук цихлид, питающихся чешуей». Наука . 260 (5105): 216–219. Бибкод : 1993Sci...260..216H . дои : 10.1126/science.260.5105.216 . ПМИД 17807183 . S2CID 33113282 .
- ^ Стюарт, штат Калифорния; Альбертсон, RC (2010). «Эволюция уникального хищного пищевого аппарата: функциональная анатомия, развитие и генетический локус латерализации челюстей у цихлид, питающихся чешуей озера Танганьика» . БМК Биология . 8 (1): 8. дои : 10.1186/1741-7007-8-8 . ПМЦ 2828976 . ПМИД 20102595 .
Дальнейшее чтение
[ редактировать ]- Хелфман, Г.С., Б.Б. Коллетт и Д.Е. Фейси (1997). Разнообразие рыб . Блэквелл Наука. стр. 33–36. ISBN 978-0-86542-256-8 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - Шульце, HP (2016). «Чешуя, эмаль, космин, ганоин и ранние остихтианы» . Comptes Рендус Палевол . 15 (1–2): 83–102. дои : 10.1016/j.crpv.2015.04.001 .

Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|