Аллохронное видообразование
| Часть серии о |
| Эволюционная биология |
|---|
 |
Аллохронное видообразование (также известное как аллохронная изоляция или временная изоляция ) — это форма видообразования (в частности, экологическое видообразование ), возникающая в результате репродуктивной изоляции , которая возникает из-за изменения времени размножения, которое уменьшает или устраняет поток генов между двумя популяциями вида. Термин аллохрония используется для описания общего экологического явления, заключающегося в различиях в фенологии , возникающих между двумя или более видами: видообразование, вызванное аллохронией, по сути, является аллохронным видообразованием.
Изменения окружающей среды, действующие на популяцию или популяции видов, могут привести к изоляции. Важной формой изоляции является разделение популяций не географически, а во времени (по времени). Генетические изменения ( мутации ) с течением времени могут привести к различиям между двумя популяциями, особенно в фенологии (события в жизни вида, диктуемые временем, такие как сезоны размножения); демонстрирующие уникальные фенотипы (наблюдаемые характеристики или черты организма).
Ученые разработали модели, объясняющие, как происходит этот процесс и как он обнаруживается в природных популяциях. Существует множество исследований, посвященных видам в аллохронии, причем некоторые из них убедительно предполагают, что виды видоизменяются или уже видоизменились как прямое следствие этого способа изоляции.
Модель
[ редактировать ]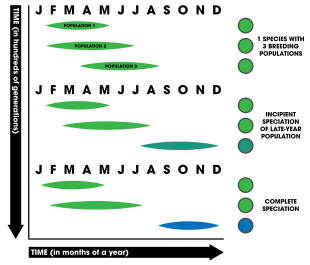
Видообразование в конечном итоге происходит из-за репродуктивной изоляции между двумя популяциями. Это может происходить множеством способов, общий способ которых известен как аллопатрическое видообразование . Географический режим, при котором два вида становятся физически изолированными и неспособными скрещиваться, позволяет отбору действовать на обе популяции независимо. Со временем это дает начало новому виду. [1] : 86 Аллохронное видообразование — это форма изоляции, которая может включать аллопатрию; однако это не требуется. [1] : 208
Аллохрония может включать ряд факторов, индуцирующих образование нового вида. Организмы развили различные репродуктивные стратегии (например, семелпарность и итеропарность , один или несколько репродуктивных циклов в течение жизни), которые могут приводить к различным результатам для аллохронии. Многие организмы также размножаются в разное время суток, в разные сезоны года и даже в течение нескольких лет или десятилетий. [2] Сезонное размножение у животных – обычное явление, как и время нереста (у водных животных). [1] : 202 У растений размножение во времени может включать в себя восприимчивость рыльца ( женской части цветка) к принятию спермы , периоды высвобождения пыльцы (например, у хвойных деревьев, где мужские шишки рассеивают пыльцу, полагаясь на ветер, чтобы направить пыльцу к женским особям). шишки) или общее время цветения (в зависимости от возможных факторов окружающей среды, таких как уровень влажности, тип или качество почвы, температура или фотопериод ). [1] : 202–206 Даже характер миграции может сыграть свою роль, поскольку виды могут оказаться изолированными из-за миграции в разное время и в разные места. [3] Считается, что изменение климата оказывает значительное влияние на аллохронию, в частности на сезонное размножение видов. [2] Моделирование изменений в моделях размножения видов, вызванных климатом, а также понимание генетических механизмов, которые его контролируют, оказались важными. [2]
Из-за этих многочисленных факторов незначительные или серьезные изменения в фенологии могут привести к расхождению между двумя популяциями. Например, вид, у которого в году несколько сезонов размножения, может смещать это время в зависимости от внешних условий, таких как температура или хищничество. Если бы популяции (аллопатрически или симпатрически распределенные ) начали размножаться в разное время, это помешало бы членам каждой популяции обмениваться генами друг с другом. Со временем, если гены не обмениваются, в каждой популяции возникают генетические различия. отбор сильно действует на две популяции, они могут оказаться репродуктивно изолированными, неспособными воспроизводить жизнеспособное, плодовитое потомство. [1] : 202–210
Чтобы считать, что аллохронное видообразование действительно произошло, модель предъявляет три основных требования: [2]
- Филогенетический анализ должен показать, что два рассматриваемых таксона являются зарождающимися видами или явно родственными таксонами.
- Время размножения должно быть генетически обусловленным ( наследственным ), а не изменяемым на протяжении всей жизни ( фенотипическая пластичность) .
- Источником дивергенции можно определить как явную аллохронию, а не результат подкрепления или других эволюционных механизмов.
Считается, что аллохрония развивается тем легче, чем выше наследственность сроков размножения, то есть чем больше связь между генами и временем размножения, тем более вероятно, что произойдет видообразование. [4] Аллохрония может быть негенетической; [1] : 203 однако для изоляции должны быть задействованы генетические факторы, чтобы привести к полной репродуктивной изоляции и последующему видообразованию. Временные рамки, связанные с аллохронией, обычно делятся на три категории (распространенность в природе, а также примеры приведены рядом с каждой категорией): [2]
- Ежедневно (считается обычным явлением) примеры включают каменные кораллы, такие как Acropora. [5] или Орбичелла . [6]
- Сезонное время размножения (считающееся наиболее распространенным) часто совпадает с зимой, весной, осенью или летом; примеры включают разведение лосося , например, нерки . [7]
- Ежегодно (считается более редким), примеры включают периодические цикады. [8] и бамбук , [9] оба из которых воспроизводятся в течение десятилетий.
Структура населения
[ редактировать ]
Ответ: В отсутствие аллохронии только географический выбор и выбор партнера вызывают изоляцию.
Б: Начинается с географического разделения, выбор партнера способствует изоляции и завершается аллохронией.
C: Начинается с дифференциации выбора партнера, за которой следует аллохрония.
D: Аллохронию сопровождают факторы спаривания и экологические факторы.
Часто обнаруживается, что другие фенотипические признаки сопутствуют срокам размножения, такие как количество цветения, размер кладки яиц, продолжительность репродуктивной жизни или размер тела - то, что можно определить как временные фенотипические клины . [4] Существуют два объяснения существования этих клин: фенотипическая пластичность или фенотипическая наследственность (или, возможно, комбинация того и другого). В случае пластичности клины возникают, когда определенные фенотипические признаки влияют на время размножения, например, при размножении в то время, когда их признаки лучше всего подходят, или если условия способствуют проявлению признаков. [4] Если они передаются по наследству, то могут быть выражены те же факторы, что и при пластическом объяснении; однако ограничения потока генов позволяют адаптироваться к конкретным условиям репродуктивного периода. Это означает, что «человек с наследственной тенденцией к раннему воспроизводству, который вместо этого воспроизводит позднее, может проявлять черты, типичные для ранних воспроизводителей». [4]
Изоляция по времени (IBT) частично аналогична концепции изоляции по расстоянию (IBD). [4] при этом генетические различия между популяциями увеличиваются с увеличением пространственного расстояния. [11] Когда IBT присутствует в популяции, изменения естественного отбора в течение сезона размножения вызывают адаптацию по времени (ABT), вызывая адаптивные временные вариации фенотипических признаков. Эти две концепции описаны в следующих разделах. Исследования лососевых рыб (с учетом продолжительности репродуктивной жизни, размера во взрослом состоянии, возраста, запаса энергии, массы яичников , размеров яиц, количества яиц в кладке, плодовитости и темпов развития) и цветковых растений (с учетом размера растений, продолжительности время цветения, количество цветков, количество плодов, время плодоношения и размер листьев) предоставили убедительные доказательства того, что ИБТ приводит к АБТ. [4] а также исследования годовой аллохронии. [2]
Изоляция по времени
[ редактировать ]Концепция IBT гарантирует две вероятности: в случае, если среди популяций, размножающихся в разные сезоны, существует наследственность по срокам размножения, вероятность спаривания будет «обратно пропорциональна разнице в наследственном компоненте их репродуктивного времени». [4] [12] Вероятность спаривания также может быть пропорциональна племенной ценности (фенотипический признак, выраженный как признак потомства) в течение репродуктивного времени в случае, если наследственность является аддитивной (более одного гена контролирует фенотипический признак). [4] В популяции потомство унаследует признаки репродуктивного времени, что приводит к уменьшению потока генов, в то время как различия во времени репродуктивного возраста увеличиваются. [4]
Адаптация по времени
[ редактировать ]Адаптация со временем — это расширение дивергенции из-за ограниченного потока генов между популяциями, испытывающими различное давление отбора. [13] Обычно это ограничивается пространственными вариациями, например, экологическим видообразованием; однако при аллохронии отбор варьируется не только в пространстве, но и в репродуктивном времени, что приводит к появлению адаптивных временных клин в фенотипических признаках, которые передаются по наследству. Изоляция во времени эффективно позволяет эволюционировать адаптивным темпоральным клинам до тех пор, пока репродуктивный сезон имеет избирательные вариации. Доказательства адаптации во времени требуют четырех факторов: 1) время ограничивает поток генов, 2) репродуктивный сезон выражает вариации фенотипических признаков, 3) временные вариации контролируются генетически (они не пластичны) и 4) временные вариации являются адаптивными. [4] ABT увеличивается, «поскольку (i) усиливается отбор по признаку; (ii) уменьшается влияние окружающей среды на репродуктивное время; (iii) увеличивается наследуемость репродуктивного времени; и (iv) временное распределение репродуктивной активности становится все более однородным». [4]
Обнаружение аллохронии
[ редактировать ]

Поскольку аллохронное видообразование может происходить в сочетании с другими способами и формами видообразования, исследователи должны попытаться определить, были ли начальные стадии изоляции результатом аллохронии. Континуум видообразования аллопатрии, парапатрии и симпатрии был вовлечен в исследования временной изоляции. [1] : 206 Аллохрония также может способствовать подкреплению после вторичного контакта . [2] Считается, что частота аллохронного видообразования обычна, но недостаточно изучена, поскольку аллохрония широко распространена в природе. [14]
Проверить, предотвращает ли аллохрония поток генов, может быть сложно из-за множества неизвестных переменных в диких популяциях и невозможности воспроизвести и манипулировать ею в лабораторных условиях. [1] : 203 Производство жизнеспособного и плодовитого потомства (или его отсутствие) не всегда возможно; к счастью, тесты на озере партнеров не обязательно указывают на отсутствие временной изоляции. [1] : 203 Как указывалось ранее, одним из необходимых критериев является то, что рассматриваемый вид должен быть родственным таксоном (или явно зарождающимся). Это означает, что точная филогения жизненно важна для определения начальной стадии видообразования. [1] : 203
Несмотря на множество исследований, не всегда очевидно, является ли аллохрония единственной движущей силой видообразования или за это ответственны другие факторы, действующие одновременно. [2] Это может быть более сложной задачей, когда видообразование уже произошло (поскольку рассматриваемые таксоны репродуктивно изолированы и больше не находятся в зачаточном состоянии). [2] Определение того, насколько важна аллохрония как историческая причина видообразования, может быть проверено с помощью: 1) сравнительных исследований, которые показывают, что молодые пары сестринских таксонов изолированы во времени, и 2) тестирования случаев зарождающихся видов в симпатрии, где репродуктивная изоляция неполная без временной изоляции. [1] : 206
Чтобы определить, является ли аллохрония источником дивергенции, необходимо измерить ключевую закономерность: изоляция (и, следовательно, видообразование) должна коррелировать с уменьшением перекрывающихся периодов размножения. [2] Эта закономерность указывает на то, что ежедневная аллохрония более склонна к потоку генов (близость времени размножения может допускать случайное смешивание популяций), тогда как годовая аллохрония наименее склонна к потоку генов (случайное смешение редко, если между периодами спаривания существуют большие временные рамки). [2]
Примеры дивергенции, вызванной аллохронией
[ редактировать ]В следующей таблице описаны случаи аллохронного видообразования. Существуют различные степени уверенности, поскольку не все случаи полностью соответствуют трем основным критериям, необходимым для аллохронного видообразования. Виды, отмеченные звездочкой (*), указывают на более высокую достоверность по оценке Ребекки Тейлор и Вики Фризен (2017). [2]
| Разновидность | Описание |
|---|---|
| Acropora samoensis * и другие виды Acropora * | Японские кораллы оказались репродуктивно изолированными по времени их нереста. [5] Популяции симпатрических видов кораллов A. samoensis нерестятся отдельно осенью и весной, причем нерест является наследственным, вероятно, с участием гена PaxC. [15] |
| Montastraea annularis *, M. faveolata * и M. Franksi * | Три родственных вида кораллов , которые видоизменились в зависимости от времени нереста. [6] |
| Oncorhynchus nerka *, O. gorbuscha * и O. tshawytscha | Ежегодные ходы размножения нерки, происходящие в течение двух периодов года (позднего и раннего), привели к генетической изоляции зарождающихся популяций. Известно, что разведение лосося является генетическим, но специфические гены этого вида неизвестны. [7] [16] [17] Четные и нечетные двухлетние жизненные циклы в сочетании с сезонным размножением горбуши ( O. gorbuscha ) привели к генетической дифференциации между двумя популяциями. [18] [19] [20] [21] [22] Время размножения также варьируется в зависимости от ареала популяции чавычи ( O. tshawytscha ). [23] [24] |
| Тауметопея питиокампа * | Кодоминирование генов связано со временем вылета личиночных стадий этого вида бабочек. Зимние и летние популяции личинок находятся в процессе видообразования. [25] [26] [27] |
| Inurois punctigera * | Размножение предотвращается в районах, где температура середины зимы не подходит для этих видов бабочек. Это привело к появлению поздних и ранних популяций. [28] |
| Pemphigus populi-transversus * и P. obesinymphae * | Галлообразующие тли образуют галлы на разных листьях одного и того же вида деревьев-хозяев. P. populi-transversus образует галлы на листьях ранней весной, а P. obesinymphae образует их на листьях летом. Это привело к полной репродуктивной изоляции. [29] |
| виды Asphondylia * | Три вида мошек поражают стебли Larrea tridentata , A. auripila летом, A. reinosa зимой и A. foliosa весной. [30] |
| Виды Целланы * | Обитая на разной глубине в пределах нескольких сантиметров, блюда стали репродуктивно изолированными, вероятно, из-за сочетания парапатрического видообразования и сигналов нереста (например, нерест в зависимости от уровня воды. [31] |
| Виды гидробатов * | В группе буревестников есть репродуктивно изолированные (на Азорских островах ) и зарождающиеся виды (на других архипелагах), вызванные прохладным и теплым сезоном размножения. [32] [33] [34] |
| Howea belmoreana * и H. forsteriana * | Генетически контролируемое время цветения привело (в сочетании с различным уровнем pH почвы ) к репродуктивной изоляции двух видов пальм на острове Лорд-Хау . [35] |
| Эрисифа-убийца * | Обладает доказательствами изоляции из-за временных различий между видами-хозяевами Vitis vinifera . [36] |
| виды Magicicada * | Группы пар видов цикад с 13- и 17-летним жизненным циклом (всего семь видов) появляются для размножения, разделенные большими временными интервалами между сезонами размножения. [37] [38] [8] Только каждые 221 год 13-летний и 17-летний циклы совпадают, когда обе пары возникают одновременно. [2] |
| Маленький Антитрогус * | Две когорты жуков демонстрируют генетическую дифференциацию жизненных циклов, разделенных двухлетними интервалами. [39] |
| Оэней полудья * | Двухлетний жизненный цикл селекционных групп видов бабочек привел к генетической дифференциации. [40] |
| Бамбусовые * | Бамбук подвергается семелпарному размножению , при котором они живут годами, прежде чем начнут массовое цветение сразу же . Это может произойти в разные годы и в разных местах. Считается, что аллохронные пятна привели к диверсификации глобальных видов бамбука. [41] [9] [42] |
| Spodoptera frugiperda | Пример фитофага: два штамма личинок моли, размножающиеся на кукурузе или рисе в разное время ночи. На вид могут действовать и другие причины изоляции. Население Соединенных Штатов, по-видимому, видоизменяется посредством аллохронии; [43] [44] однако население Колумбии этого не делает. [45] |
| Anopheles gambiae и A. coluzzii | Контролируемые циркадными ритмами, которые стимулируют спаривание, комары роятся в сумерках в несколько разное время , демонстрируя некоторые признаки аллохронии, хотя возможно, что подкрепление или микроаллопатрическое видообразование . здесь играет роль [46] [47] |
| Bactrocera tryoni и B. neohumeralis | Между двумя плодовыми мухами Квинсленда наблюдалась только лабораторная гибридизация, последняя из которых спаривается только в дневное время, а первая - только ночью. [48] |
| Anastrepha bistrigata и A. striata | Плодовые мушки спариваются утром и днем соответственно. Поведение ухаживания также может привести к изоляции этих двух видов. [49] |
| Салмо Салар | Возраст полного созревания, а также генетическая дифференциация варьируются от однолетнего до трехлетнего атлантического лосося (это годы, в которые молодь уходит в океан и возвращается в места размножения). [50] |
| Ангилья Ангилья | Европейские угри имеют разную скорость взросления в зависимости от факторов окружающей среды. Это создает отдельные гнездящиеся популяции, которые демонстрируют некоторую генетическую дифференциацию, особенно между 2–3-летними интервалами размножения. [51] |
| Поющая кукушка | Аллохрония, вероятно, играет роль у птиц-кукушек, поскольку они зависят от вида-хозяина в воспитании птенцов. Хозяева откладывают яйца в разное время, и кукушки зависят от этих сроков, чтобы заменить яйца птицы-хозяина. [52] |
| Antechinus spp. | Сумчатые мыши сильно реагируют на фотопериодические сигналы , и в симпатрических популяциях репродуктивно изолированные виды размножаются в разное время. [53] |
| Coregonus clupeaformis | Озерный сиг имеет две известные формы: нормальную и карликовую. У них разное время нереста, но они могли различаться по аллопатрии. [54] [55] |
| Exapion ulicis и E. lemovicinum | E. lemovicinum инфицирует Ulex major и U. Gallii, растения а E. ulicis инфицирует U. europaeus . Время откладки яиц на растение приходится на осень и весну соответственно. [56] |
| Meconopsis Autumnalis и M. метельчатая | Гималайский мак — это полностью репродуктивно изолированный вид, который, как полагают, возник в результате аллохронии, поскольку они существуют в симпатии и цветут в разное время сезона. [57] |
| Кордия виды. | Некоторые виды этого рода демонстрируют значительные различия во времени цветения. [58] |
| Гесперииды | Считается, что временная изоляция ответственна за видообразование многих из 400 изученных видов бабочек-шкиперов. [59] |
| Bryopsidales spp. | Зеленые водоросли размножаются, выпуская гаметы в разное время. Считается, что это привело к репродуктивной изоляции, но неясно, контролируется ли она генетически или основано исключительно на факторах окружающей среды. [60] |
| Чило Супрессалис | Время спаривания происходит в разные промежутки времени в ночное время, а также зависит от разных растений-хозяев. [61] |
| Продоксус quinquepuntellus | Расы-хозяева бабочек населяют юкку Адамова иголка и нить, причем появление личинок происходит одновременно со временем цветения. Считается, что морфология и смена хозяина способствуют наряду с аллохронией. [62] |
| Gryllus pennsylvanicus и G. veletis | Весенние полевые сверчки были описаны как особи аллохронии из-за сроков их созревания. [63] Однако более поздние исследования показывают, что это не родственные виды. [64] |
| виды Haemaphysalis . и Dermacentor spp. | Три вида венгерских клещей рода Haemaphysalis проявляют симпатическую брачную активность в течение трех сезонных периодов: поздней осени, поздней весны и ранней весны. Два вида клещей рода Dermacentor демонстрируют пик активности осенью и весной. [65] |
| Страуция длинная пеннис | Генетическая изменчивость обнаружена у трех вариантов личинки подсолнечника, обитающих на одном и том же растении-хозяине. Вылет их личинок происходит в три разных периода лета, что делает их частично изолированными. Экспериментальные манипуляции предполагают, что аллохрония будет увеличиваться по мере того, как они продолжают расходиться. [66] [67] |
| Энченопа бинотата Комплекс | Фенология представителей видового комплекса цикадок коррелирует с их растениями-хозяевами: при смене вида-хозяина в эксперименте время вылупления яиц цикадки меняется, что способствует ассортативному спариванию. [68] |
| возвышенное ополчение | Поскольку время цветения африканского тика считается генетически контролируемым, популяция демонстрирует изоляцию. [69] |
| Астералобия сасакии | Две популяции галлиц Cecidomyiidae существенно различаются по времени вылета (без перекрытия) на двух разных Ilex . хозяевах [70] |
| Salix виды. | Виды канадской ивы изолированы по времени цветения; три из них цветут рано ( S. bebbiana , S. discolor , S. eriocephala и S. petiolaris ) и поздно ( S. amygdaloides , S. exigua и S. lucida ). Гибриды неизвестны за пределами лабораторных условий и имеют промежуточное время цветения. Все семь видов существуют в симпатрическом распространении. [71] |
| Юнкус сарай | Симпатрические популяции генетически дифференцированных растений цветут в разное время, что предотвращает гибридизацию. Неясно, происходит ли видообразование путем аллохронии, поскольку подкрепление может быть более сильным объяснением. [72] |
| Агростис тонкий | Вид травы A. tenuis растет на почве, загрязненной высоким содержанием меди, выщелоченной из неиспользуемой шахты. Рядом находится незагрязненная почва. Популяции развивают репродуктивную изоляцию из-за различий во времени цветения. [73] |
| Антоксантус душистый | Вид травы A. odoratum растет на почве, загрязненной высоким содержанием свинца и цинка, выщелоченных из неиспользуемой шахты. Рядом находится незагрязненная почва. Популяции развивают репродуктивную изоляцию из-за различий во времени цветения. [73] |
| Хирономус нудитарсис | Некусающая мошка (род Chironomus ) демонстрирует различия в жизненном цикле в зависимости от высоты. [74] |
| Терелия фусикорнис | Различия в ухаживающем поведении, а также в морфологии обнаружены в популяциях, заражающих разных хозяев ( Silybum marianum и Cynara ), цветущих в разное время. [75] |
| виды Ampelomyces . | Генетически разные штаммы микопаразитного гриба, поражающего мучнистую росу яблони, завершают свой жизненный цикл раньше других штаммов, поражающих других хозяев мучнистой росы. [76] |
| Глицин макс и Г. соя | Гибридизацию дикой сои ( G. soja ) и культивируемой сои ( G. max ) можно предотвратить, вызывая асинхронность во времени цветения. [77] Это исследование уникально тем, что оно не является примером аллохронного видообразования, а является экспериментом, демонстрирующим, что аллохрония может быть экспериментально применена для индукции изоляции. |
| Паразитоидные осы: Rhagoletis pomonella , Rhagoletis mendax , Diachasma alloeum , Diachasmimorpha mellea и Utetes canaliculatus. | У R. pomonella (одного из наиболее изученных модельных организмов) генетические данные указывают на наследственность вылета и связанное с ним время полета. [78] На коммерческих полях черники по сравнению с дикими популяции R. mendax различаются по периодам полета, что приводит к снижению потока генов. [79] Другие виды Rhagoletis , обитающие на Crataegus, демонстрируют аналогичные закономерности. [80] Совместное видообразование паразитоидных ос ( D. alloeum , D. mellea и U. canaliculatus ) и яблочной личинки их растения-хозяина было вызвано сменой хозяев, вызванной различными факторами, такими как время вылупления яиц, предпочтение запаха фруктов, филопатрия , и избегание . Фактор времени вылупления яиц предполагает аллохронию. [81] |
| Из евро солидагиниса | Две популяции золотарника-галловой мухи различаются по срокам вылета на растениях-хозяевах Solidago altissima и S. gigantea на 10–14 дней, что препятствует гибридизации и вызывает изоляцию. [82] |
| Фалько-перепелятник | Пустельги Айдахо имеют как мигрирующих, так и круглогодичных жителей, причем круглогодичная популяция, гнездящаяся раньше, приводит к ассортативному спариванию. [83] |
| Сильвия атрикап | Существует некоторая генетическая дифференциация между популяциями черной шапки, мигрирующими в Соединенное Королевство и Ирландию, и популяциями, мигрирующими в Иберию. Птицы дружно размножаются в Германии; однако популяции Великобритании и Ирландии мигрируют обратно раньше, вызывая ассортативное спаривание. Гибриды имеют промежуточные пути миграции. [84] |
| Junco hyemalis hyemalis и J. h. Каролина | Фотопериодические сигналы способствуют более раннему развитию гонад у J. h. carolinensis , из которых мигрируют в другой регион для размножения, проживая только симпатически с J. h. hyemalis птиц в течение полугода. [85] |
| Daphnia pulex и D. pulicaria | Между двумя видами водяных блох обнаружена очень ограниченная изоляция ( D. pulicaria входит в комплекс D. pulex ), возможно, в результате определения времени размножения, основанного на фотопериодических сигналах. [86] |
| Трубка | Изоляция местообитаний, аллохрония и аллопатрия выявлены у различных пар семи видов и подвидов рода ( T. Corsica Corsica , T. Corsica fairmairei , T. garricola , Thaematodes , T. nigronervosa , T. Quadrisignata и T. tomentosa). ). [87] |
| Кальтенбачиелла японская | Размножение тли зависит от появления почек у вяза японского . Зарождающиеся популяции возникли вследствие изменения сроков бутонизации растения-хозяина. [88] |
| Scolioneura betuleti и сосед S. | Листогрызущие пилильщики, несмотря на большое сходство, демонстрируют некоторые признаки расхождения из-за сезонного лета (соответственно осенью и весной). [89] [90] |
| Papilio canadensis и P. glaucus | Два вида бабочек гибридизировались, создав гибридную популяцию, которая размножается в другое время, чем родительские популяции. Генетические данные показывают, что гены контролируют время размножения. [91] [92] |
| Саванна тиран | Популяции вилкохвостых мухоловок расходятся из-за изоляции из-за изменения времени размножения и мест размножения в результате утраты миграционного поведения. [93] |
| Остриния nubilalis | Штаммы кукурузной мотыльки, размножающиеся в разное время ночи, считаются зарождающимися, однако неясно, является ли изоляция исключительной причиной аллохронии. [94] Сезонное размножение может привести к изоляции североамериканских популяций из-за количества ежегодных выводков ( вольтинизм ) между двумя линиями. Симпатричные друг другу штамм Z является моновольтинным (имеет один выводок в год), а штамм E - дивольтинным (имеет два выводка в год). [14] В Европе существует корреляция между временем появления личинок и растением-хозяином. [95] |
| Виды птиц Нового Света | В эксперименте по проверке гипотезы асинхронности сезонов (см. раздел ниже) было обнаружено, что 57 различных видов птиц, обитающих в Новом Свете (Северная и Южная Америка), демонстрируют повышенную генетическую дифференциацию в корреляции с проживанием в районах с асинхронными осадками. [96] |
Гипотеза асинхронности сезонов
[ редактировать ]Примечательной и важной закономерностью в природе являются широтные градиенты видового разнообразия . [97] [98] где видовое богатство ( биоразнообразие Земли ) увеличивается ближе к экватору . Считается, что одним из факторов, способствующих этому, является то, что в этих регионах по всей планете темпы видообразования выше. [99] Предполагается, что гипотеза асинхронности сезонов является фактором, способствующим более высокой скорости видообразования, поскольку она напрямую связана с аллохронным видообразованием. [100] Гипотеза предполагает, что такая закономерность является результатом отсутствия синхронности с сезонными колебаниями в тропических регионах. [100]
В высоких широт регионах различные таксоны испытывают схожие температуры и солнечную радиацию в циклическом порядке из-за наклона оси Земли , создавая сезоны, которых нет на экваторе. Из-за этого фенология популяций вида обычно синхронна на всем ареале, что обеспечивает беспрепятственное распространение и последующий поток генов. Это приводит к уменьшению различий между популяциями; в конечном итоге снижая скорость видообразования (и общее биоразнообразие). [100]
В отличие от этого, в широтах вблизи экватора или в тропиках наблюдается асинхронность сезонных изменений, поскольку регионы получают одинаковое количество солнечной радиации и поддерживают постоянную температуру. Что меняется, так это характер осадков , поскольку они могут быть спорадическими, рассеянными и варьироваться на очень коротких расстояниях. Эта закономерность вызывает асинхронность фенологии популяций видов, что в конечном итоге может выступать в качестве временного изолирующего барьера для воспроизводства. Это предотвращает поток генов между популяциями и может способствовать дивергенции, видообразованию и увеличению биоразнообразия. [100]
Масштабная проверка гипотезы была проведена на пятидесяти семи видах птиц Нового Света в Южной, Центральной и Северной Америке. [2] Результаты, полученные с использованием ДНК, географических и экологических расстояний, а также климатических данных, показали, что генетическая дифференциация усилилась в популяциях видов, где присутствовала асинхронность осадков. [96] Похожая картина была обнаружена в другом исследовании сорока восьми неотропических лягушек. [101]
Ссылки
[ редактировать ]- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к Джерри А. Койн ; Х. Аллен Орр (2004), Видообразование , Sinauer Associates, стр. 1–545, ISBN 0-87893-091-4
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л м н тот п Ребекка С. Тейлор и Вики Л. Фризен (2017), «Роль аллохронии в видообразовании», Molecular Ecology , 26 (13): 3330–3342, Бибкод : 2017MolEc..26.3330T , doi : 10.1111/mec.14126 , ПМИД 28370658 , С2КИД 46852358
- ^ Турбек, Шила П.; Скордато, Элизабет СК; Сафран, Ребекка Дж. (2018), «Роль сезонной миграции в демографической дивергенции и репродуктивной изоляции», Trends in Ecology & Evolution , 33 (3): 164–175, doi : 10.1016/j.tree.2017.11.008 , ПМИД 29289354
- ↑ Перейти обратно: Перейти обратно: а б с д и ж г час я дж к л Хендри, Эндрю П.; Дэй, Трой (2005), «Структура населения, обусловленная репродуктивным временем: изоляция по времени и адаптация по времени», Molecular Ecology , 14 (4): 901–916, Бибкод : 2005MolEc..14..901H , doi : 10.1111/ j.1365-294X.2005.02480.x , PMID 15773924 , S2CID 8226535
- ↑ Перейти обратно: Перейти обратно: а б Фуками, Х.; Омори, М.; Симойке, К.; Хаясибара, Т.; Хатта, М. (2003), «Экологические и генетические аспекты репродуктивной изоляции кораллов Acropora в разное время нереста», Marine Biology , 142 (4): 679–684, Bibcode : 2003MarBi.142..679F , doi : 10.1007/ s00227-002-1001-8 , S2CID 81981786
- ↑ Перейти обратно: Перейти обратно: а б Ноултон, Н.; Мате, Дж.Л.; Гусман, Ее Величество; Роуэн, Р.; Хара, Дж. (1997), «Прямые доказательства репродуктивной изоляции трех видов комплекса Montastraea annularis в Центральной Америке (Панама и Гондурас)», Marine Biology , 127 (4): 705–711, Bibcode : 1997MarBi.127 ..705K , doi : 10.1007/s002270050061 , S2CID 37997956
- ↑ Перейти обратно: Перейти обратно: а б Хендри, Эндрю П.; Берг, Оле К.; Куинн, Томас П. (1999), «Зависимость от условий и адаптация ко времени: дата размножения, история жизни и распределение энергии в популяции лосося», Oikos , 85 (3): 499–514, Bibcode : 1999Oikos. .85..499H , doi : 10.2307/3546699 , JSTOR 3546699
- ↑ Перейти обратно: Перейти обратно: а б Сота, Тейджи; Ямамото, Сатоши; Кули, Джон Р.; Хилл, Кэти БР; Саймон, Крис; Ёсимура, Джин (2013), «Независимое расхождение 13- и 17-летних жизненных циклов среди трех периодических линий цикад», PNAS , 110 (17): 6919–6924, Bibcode : 2013PNAS..110.6919S , doi : 10.1073/pnas .1220060110 , PMC 3637745 , PMID 23509294
- ↑ Перейти обратно: Перейти обратно: а б Дональд К. Франклин (2004), «Синхронность и асинхронность: наблюдения и гипотезы волны цветения у долгоживущего семелеродящего бамбука», Journal of Biogeography , 31 (5): 773–786, Bibcode : 2004JBiog..31.. 773F , doi : 10.1111/j.1365-2699.2003.01057.x , S2CID 55279438
- ^ Дикманн, У.; Добели, М.; Мец, JAJ; Таутц, Д. (2004), Адаптивное видообразование (PDF) , Кембридж, Великобритания: Издательство Кембриджского университета, ISBN 9781107404182
- ^ Монтгомери Слаткин (1993). «Изоляция расстоянием в равновесных и неравновесных популяциях». Эволюция . 47 (1): 264–279. дои : 10.2307/2410134 . JSTOR 2410134 . ПМИД 28568097 .
- ^ Гордон А. Фокс (2003), «Ассортативное спаривание и фенология растений: эволюционные и практические последствия» (PDF) , Evolutionary Ecology Research , 5 : 1–18
- ^ Дольф Шлютер (2000), Экология адаптивной радиации , Oxford University Press, ISBN 0198505221
- ↑ Перейти обратно: Перейти обратно: а б Допман, Э.Б.; Роббинс, PS; Симэн, А. (2010), «Компоненты репродуктивной изоляции между североамериканскими штаммами феромонов европейского кукурузного мотылька», Evolution , 64 (4): 881–902, doi : 10.1111/j.1558-5646.2009.00883.x , PMC 2857697 , ПМИД 19895559 , С2КИД 9909878
- ^ Натали Л. Россер (2015), «Асинхронный нерест в симпатрических популяциях твердого коралла выявляет загадочные виды и древние генетические линии», Molecular Ecology , 24 (19): 5006–5019, Bibcode : 2015MolEc..24.5006R , doi : 10.1111/ mec.13372 , PMID 26339867 , S2CID 13151100
- ^ Эндрю П. Хендри, Иоланда Э. Морби, Оле К. Берг и Джон К. Венбург (2004), «Адаптивная изменчивость старения: продолжительность репродуктивной жизни в популяции дикого лосося», Proceedings of the Royal Society B: Biological Sciences , 271 (1536): 259–266, doi : 10.1098/rspb.2003.2600 , PMC 1691593 , PMID 15058436
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Филлатр, ЕК; Этертон, П.; Хит, Д.Д. (2003), «Бимодальное распределение пробега в северной популяции нерки (Oncorhynchus nerka): история жизни и генетический анализ во временном масштабе», Molecular Ecology , 12 (7): 1793–1805, Bibcode : 2003MolEc. .12.1793F , doi : 10.1046/j.1365-294x.2003.01869.x , PMID 12803632 , S2CID 25772120
- ^ Животовский Лев А.; Гарретт, Эй Джей; МакГрегор, AJ; Глубоковский, М.К.; Фельдман, Маркус В. (1994), «Дифференциация генов у тихоокеанского лосося (Oncorhynchus Sp.): факты и модели применительно к горбуше (O. Gorbuscha)», Canadian Journal of Fisheries and Aquatic Sciences , 51 : 223–232, дои : 10.1139/f94-308
- ^ Д. Чуриков и А.Дж. Гарретт (2002), «Сравнительная филогеография двух выводков горбуши: анализ, основанный на генеалогии митохондриальной ДНК», Molecular Ecology , 11 (6): 1077–1101, Bibcode : 2002MolEc..11.1077C , doi : 10.1046/j.1365-294x.2002.01506.x , PMID 12030984 , S2CID 24965183
- ^ Лимборг, Мортен Т.; Уэплс, Райан К.; Сиб, Джеймс Э.; Сиб, Лиза В. (2014), «Временно изолированные линии горбуши обнаруживают уникальные признаки отбора в различных пулах постоянных генетических вариаций», Journal of Heredity , 105 (6): 835–845, doi : 10.1093/jhered/esu063 , PMID 25292170
- ^ Курильщик, WW; Гарретт, Эй Джей; Стеколл, М.С. (1998), «Генетическая изменчивость даты возвращения в популяции горбуши: следствие меняющейся окружающей среды и дисперсионного отбора?», Бюллетень исследований рыбного хозяйства Аляски , 5 : 46–54.
- ^ Гарретт, Эй Джей; Лейн, С; МакГрегор, AJ; Тейлор, С.Г. (2001), «Использование генетического маркера для изучения генетического взаимодействия между субпопуляциями горбуши ( Oncorhynchus gorbuscha )», Genetica , 111 (1–3): 259–267, doi : 10.1023/A:1013791314900 , PMID 11841171 , S2CID 19278664
- ^ Куинн, ТП; Анвин, MJ; Киннисон, М.Т. (2000), «Эволюция временной изоляции в дикой природе: генетическое расхождение во времени миграции и размножения интродуцированных популяций чавычи», Evolution , 54 (4): 1372–1385, doi : 10.1111/j.0014- 3820.2000.tb00569.x , PMID 11005303 , S2CID 23316021
- ^ Бриё, MSO; Оно, К; Дринан, ДП; Нэйш, К.А. (2015), «Интеграция случайного леса с популяционным анализом выбросов дает представление о геномной основе и эволюции времени пробега чавычи ( Oncorhynchus tshawytscha )», Molecular Ecology , 24 (11): 2729–2746, Bibcode : 2015MolEc..24.2729B , doi : 10.1111/mec.13211 , PMID 25913096 , S2CID 206182207
- ^ Пиментель, К.; Кальвао, Т.; Сантос, М.; Феррейра, К.; Невес, М.; Нильссон, Дж.О. (2006), «Создание и расширение популяции Thaumetopoeapityocampa (Den. & Schiff.) (Lep. Notodontidae) со смещенным жизненным циклом в продуктивном сосновом лесу, Центрально-Прибрежная Португалия», Лесная экология и управление , 233 (1 ): 108–115, doi : 10.1016/j.foreco.2006.06.005
- ^ Сантос, Хелена М; Пайва, Мария-Роза; Роча, Сусана; Кердельуэ, Кэрол; Бранко, Мануэла (2013), «Фенотипическое расхождение репродуктивных признаков популяции бабочек, переживающих фенологический сдвиг», Ecology and Evolution , 3 (15): 5098–5108, Bibcode : 2013EcoEv...3.5098S , doi : 10.1002/ece3 .865 , PMC 3892371 , PMID 24455139
- ^ Бранко, Мануэла; Пайва, Мария-Роза; Сантос, Хелена Мария; Бурбан, Кристиан; Кердельюэ, Кэрол (2017), «Экспериментальные доказательства наследственного репродуктивного времени в двух аллохронных популяциях сосновой процессионной бабочки», Insect Science , 24 (2): 325–335, Бибкод : 2017InsSc..24..325B , doi : 10.1111/ 1744-7917.12287 , ПМИД 26530538 , С2КИД 9091980
- ^ Ямамото, Сатоши; Сота, Тейджи (2012), «Параллельная аллохронная дивергенция у зимней бабочки из-за нарушения репродуктивного периода из-за суровых зим», Molecular Ecology , 21 (1): 174–183, Bibcode : 2012MolEc..21..174Y , doi : 10.1111/j.1365-294X.2011.05371.x , PMID 22098106 , S2CID 23572464
- ^ Патрик Эббот и Джеймс Х. Уитготт (2004), «Филогенетические и молекулярные доказательства аллохронного видообразования у тлей, образующих галлы (пузырчатка)», Evolution , 58 (3): 539–553, doi : 10.1111/j.0014-3820.2004.tb01677 .x , PMID 15119438 , S2CID 25277034
- ^ Джеффри Б. Джой и Бернард Дж. Креспи (2007), «Адаптивная радиация насекомых, вызывающих галл, в пределах одного вида растений-хозяев», Evolution , 61 (4): 784–795, doi : 10.1111/j.1558-5646.2007.00069 .x , PMID 17439611 , S2CID 16864372
- ^ Берд, Кристофер Э.; Холланд, Бренден С.; Боуэн, Брайан В.; Тунен, Роберт Дж. (2011), «Диверсификация симпатрических блюдечек, нерестящихся широковещательно (Cellana spp.) на Гавайском архипелаге», Molecular Ecology , 20 (10): 2128–2141, Бибкод : 2011MolEc..20.2128B , doi : 10.1111 /j.1365-294X.2011.05081.x , PMID 21481050 , S2CID 23432529
- ^ Л. Р. Монтейро (1998), «Видообразование посредством временной сегрегации популяций мадейрских кастро (Oceanodroma castro) на Азорских островах?», « Философские труды Лондонского королевского общества». Серия B, Биологические науки , 353 (1371): 945–953, doi : 10.1098/rstb.1998.0259 , PMC 1692297.
- ^ Фризен, В.Л.; Смит, Алабама; Гомес-Диас, Э.; Болтон, М.; Фернесс, РВ; Гонсалес-Солис, Дж.; Монтейро, ЛР (2007), «Симпатрическое видообразование путем аллохронии у морских птиц» (PDF) , PNAS , 104 (47): 18589–18594, Бибкод : 2007PNAS..10418589F , doi : 10.1073/pnas.0700446104 , PMC 214 1821 г. , ПМИД 18006662
- ^ Болтон, Марк; Смит, Андреа Л.; Гомес-Диас, Елена; Фризен, Вики Л.; Медейрос, Рената; Брид, Жоэль; Роскалес, Хосе Л.; Фернесс, Роберт В. (2008), «Буревестник Монтейро Oceanodroma monteiroi: новый вид с Азорских островов», Ibis , 150 (4): 717–727, doi : 10.1111/j.1474-919X.2008.00854.x
- ^ Саволайнен, Винсент; Анстетт, Мари-Шарлотта; Лексер, Кристиан; Хаттон, Ян; Кларксон, Джеймс Дж; Норуп, Мария V; Пауэлл, Мартин П.; Спрингейт, Дэвид; Саламин, Николас; Бейкер, Уильям Дж. (2006), «Симпатрическое видообразование на пальмах на океаническом острове», Nature , 441 (7090): 210–213, Бибкод : 2006Natur.441..210S , doi : 10.1038/nature04566 , PMID 16467788 , S2CID 867216
- ^ Монтарри, Жослен; Картоларо, Филипп; Ричард-Сервера, Сильви; Дельмотт, Франсуа (2009), «Пространственно-временное распределение генетических групп Erysiphe necator и их связь с уровнем заболеваемости виноградников», Европейский журнал патологии растений , 123 (1): 61–70, Bibcode : 2009EJPP..123.. .61M , CiteSeerX 10.1.1.511.8215 , doi : 10.1007/s10658-008-9343-9 , S2CID 13114251
- ^ Д. К. Маршалл и Дж. Р. Кули (2000), «Смещение репродуктивных признаков и видообразование у периодических цикад с описанием новых видов, 13-летняя Magicicada neotredecem», Evolution , 54 (4): 1313–1325, doi : 10.1111/j.0014 -3820.2000.tb00564.x , hdl : 2027.42/73691 , PMID 11005298 , S2CID 28276015
- ^ Саймон, К.; Тан, Дж.; Далвади, С.; Стейли, Г.; Дениега, Дж.; Уннаш, Т.Р. (2000), «Генетические доказательства ассортативного спаривания между 13-летними цикадами и симпатрическими «17-летними цикадами с 13-летним жизненным циклом» подтверждают аллохронное видообразование», Evolution , 54 (4): 1326–1336, doi : 10.1111/j.0014-3820.2000.tb00565.x , PMID 11005299 , S2CID 19105047
- ^ Д. П. Логан, П. Г. Олсопп и М. П. Залуки (2003), «Зимовка, распределение почвы и фенология Canegrub Чайлдерса, Antitrogus parvulus (Coleoptera: Scarabaeidae) в сахарном тростнике Квинсленда», Бюллетень энтомологических исследований , 93 (4): 307–314, doi : 10.1079/ber2003245 , PMID 12908916
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ А.Э. Градиш, Н. Кейгобади и Г.В. Отис (2015), «Генетическая структура популяции и генетическое разнообразие находящейся под угрозой исчезновения арктической бабочки Белой горы (Oeneis melissa semidea)», Conservation Genetics , 16 (5): 1253–1264, Bibcode : 2015ConG ...16.1253G , doi : 10.1007/s10592-015-0736-y , S2CID 13307002
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мадхав Гаджил и С. Нарендра Прасад (1984), «Экологические детерминанты эволюции жизненного цикла двух индийских видов бамбука», Biotropica , 16 (3): 161–172, Бибкод : 1984Biotr..16..161G , doi : 10.2307/ 2388050 , JSTOR 2388050
- ^ Анелена Л. де Карвалью, Брюс В. Нельсон, Милтон К. Бьянкини, Даниэла Планьоль, Татьяна М. Куплич и Дуглас К. Дейли (2013), «Леса юго-западной Амазонки с преобладанием бамбука: обнаружение, пространственный масштаб, жизненный цикл Длина и волны цветения", PLOS ONE , 8 (1): e54852, Bibcode : 2013PLoSO...854852C , doi : 10.1371/journal.pone.0054852 , PMC 3554598 , PMID 23359438
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Шёфл Г., Дилл А., Хекель Д.Г. и Грут А.Т. (2011), «Аллохронное разделение против выбора партнера: неслучайные закономерности спаривания между штаммами-хозяевами кукурузной совки», American Naturalist , 177 (4): 470–485, doi : 10.1086/ 658904 , ПМИД 21460569 , С2КИД 12528391
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Шёфл Г., Хекель Д.Г. и Грут А.Т. (2009), «Смещенное во времени репродуктивное поведение среди штаммов-хозяев осенней совки (Noctuidae: Spodoptera frugiperda ): доказательства различных способов наследования», Журнал эволюционной биологии , 22 (7): 1447 –1459, doi : 10.1111/j.1420-9101.2009.01759.x , PMID 19467132 , S2CID 22004781
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Салдамандо-Бенхумеа С.И., Эстрада-Пиедрахита К., Веласкес-Велес М.И. и Бейли Р.И. (2014), «Ассортативное спаривание и отсутствие временного соответствия между штаммами кукурузы и риса Spodoptera frugiperda (Lepidoptera, Noctuidae) из центральной Колумбии», Журнал насекомых Поведение , 27 (5): 555–566, Бибкод : 2014JIBeh..27..555S , doi : 10.1007/s10905-014-9451-7 , S2CID 16310075
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сэмюэл С.С. Рунд, Сэмюэл Дж. Ли, Брайан Р. Буш и Джайлс Э. Даффилд (2012), «Деформационные и половые различия в ежедневной полетной активности и циркадных часах комаров Anopheles gambiae », Журнал физиологии насекомых , 58 (12 ): 1609–1619, doi : 10.1016/j.jinsphys.2012.09.016 , PMID 23068991
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Савадого, Саймон П; Константини, Карло; Пеннетье, Седрик; Диабате, Абдулай; Гибсон, Габриэлла ; Дабире, Рох К. (2013), «Различия во времени спаривания роев в симпатрических популяциях Anopheles coluzzii и Anopheles gambiae ss (ранее молекулярные формы Anopheles gambiae M и S) в Буркина-Фасо, Западная Африка», Parasites & Vectors , 6 ( 275): 275–288, doi : 10.1186/1756-3305-6-275 , PMC 3851435 , PMID 24330578
- ^ Н. Пайк, ВИС Ван и А. Митс (2003), «Вероятная судьба гибридов Bactrocera tryoni и Bactrocera neohumeralis », Heredity , 90 (5): 365–370, doi : 10.1038/sj.hdy.6800253 , PMID 12714981 , S2CID 34631721
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Дениз Селивон и Жоао С. Морганте (1997), «Репродуктивная изоляция между Anastrepha bistrigata и A. striata (Diptera, Tephritidae)», Бразильский журнал генетики , 20 (4): 583–585, doi : 10.1590/S0100-84551997000400005
- ^ Сьюзан Э. Джонстон, Пану Орелл, Виктория Л. Притчард, Мэтью П. Кент, Сигбьёрн Лиен, Ээро Ниемеля, Яакко Эркинаро и Крейг Р. Прайммер (2014), «Полногеномный анализ SNP выявляет генетическую основу изменений морского возраста в дикой природе». популяция атлантического лосося ( Salmo salar )», Molecular Ecology , 23 (14): 3452–3468, doi : 10.1111/mec.12832 , PMID 24931807 , S2CID 11737380
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Маес Г.Э., Пуйолар Дж.М., Хеллеманс Б. и Волкарт ФАМ (2006), «Доказательства изоляции по времени у европейского угря ( Anguilla anguilla L.)», Molecular Ecology , 15 (8): 2095–2107, Bibcode : 2006MolEc. .15.2095M , doi : 10.1111/j.1365-294X.2006.02925.x , PMID 16780427 , S2CID 10586483
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ AP Møller, A Antonov, BG Stokke, F Fossøy, A Moksnes, E Røskaft и F Takasu (2011), «Изоляция по времени и среде обитания и сосуществование различных рас-хозяев обыкновенной кукушки», Journal of Evolutionary Biology , 24 ( 3): 676–684, doi : 10.1111/j.1420-9101.2010.02202.x , PMID 21214656 , S2CID 24056512
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Макаллан Б.М., Дикман Ч.Р. и Кроутер М.С. (2006), «Фотопериод как репродуктивный сигнал у сумчатых рода Antechinus: экологические и эволюционные последствия», Биологический журнал Линнеевского общества , 87 (3): 365–379, doi : 10.1111 /j.1095-8312.2006.00571.x
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Киркпатрик М. и Селандер Р.К. (1979), «Генетика видообразования озерных сигов в бассейне Аллегаш», Evolution , 33 (1Part2): 478–485, doi : 10.1111/j.1558-5646.1979.tb04700.x , PMID 28568177 , S2CID 32131280
- ^ Роджерс С.М., Изабель Н. и Бернатчес Л. (2007), «Карты связей комплекса видов карликового и обычного озерного сига ( Coregonus clupeaformis ) и их гибридов раскрывают генетическую архитектуру популяционной дивергенции», Genetics , 175 (1): 375– 398, doi : 10.1534/genetics.106.061457 , PMC 1774998 , PMID 17110497
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Барат М., Тарайр М. и Атлан А. (2007), «Фенология растений и хищничество семян: взаимодействие между дроками и долгоносиками в Бретани (Франция)», Entomologia Experimentalis et Applicata , 124 (2): 167–176, Bibcode : 2007EEApp. 124..167B , doi : 10.1111/j.1570-7458.2007.00565.x , S2CID 85880513
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Иган П.А. (2011), « Meconopsis Autumnalis и M. manasluensis (Papaveraceae), два новых вида гималайского мака, эндемичных для центрального Непала с симпатрическими родственниками», Phytotaxa , 20 : 47–56, doi : 10.11646/phytotaxa.20.1.4 , HDL : 2262/56528
- ^ Оплер П.А., Бейкер Х.Г. и Фрэнки Г.В. (1975), «Репродуктивная биология некоторых видов Коста-Рики Cordia (Boraginaceae)», Biotropica , 7 (4): 234–247, Bibcode : 1975Biotr...7..234O , doi : 10.2307/2989736 , JSTOR 2989736
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Деврис П.Дж., Остин Г.Т. и Мартин Н.Х. (2008), «Дильская активность и репродуктивная изоляция в разнообразном сообществе неотропических шкиперов (Lepidoptera: Hesperiidae)», Биологический журнал Линнеевского общества , 94 (4): 723–736, doi : 10.1111/j.1095-8312.2008.01037.x
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Клифтон К. и Клифтон Л. (1999), «Фенология полового размножения зеленых водорослей (Bryopsidales) на коралловых рифах Карибского моря», Journal of Phycology , 35 (1): 24–34, Бибкод : 1999JPcgy..35...24C , doi : 10.1046/j.1529-8817.1999.3510024.x , S2CID 83704320
- ^ Уэно Х., Фурукава С. и Цучида К. (2006), «Разница во времени спаривания между популяциями рисового стеблевого мотыля, связанными с хозяином, Chilo compressalis (Walker)», Entomological Science , 9 (3): 255–259. , doi : 10.1111/j.1479-8298.2006.00171.x , S2CID 86106927
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Громан Дж. Д. и Пеллмир О. (2000), «Быстрая эволюция и специализация после колонизации хозяина бабочкой юкки», Журнал эволюционной биологии , 13 (2): 223–236, doi : 10.1046/j.1420-9101.2000.00159.x , S2CID 84556390
- ^ Александр Р.Д. и Бигелоу Р.С. (1960), «Аллохронное видообразование полевых сверчков и новый вид Acheta veletis », Evolution , 14 (3): 334–346, doi : 10.1111/j.1558-5646.1960.tb03095.x , hdl : 2027.42/137466 , S2CID 87867996
- ^ Харрисон Р.Г., Богданович С.М. и Холл С. (1995), «Филогения митохондриальной ДНК североамериканских полевых сверчков: взгляды на эволюцию жизненных циклов, песен и ассоциаций среды обитания», Журнал эволюционной биологии , 8 (2): 209– 232, номер документа : 10.1046/j.1420-9101.1995.8020209.x , S2CID 85777010
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хорнок, С. (2009), «Аллохронный сезонный пик активности видов Dermacentor и Haemaphysalis в условиях континентального климата в Венгрии», Veterinary Parasitology , 163 (4): 366–369, doi : 10.1016/j.vetpar.2009.03.048 , ПМИД 19410373
- ^ Хиппи AC, Элнес М.Э., Армента Дж.С., Кондон М.А. и Форбс А.А. (2016), «Дивергенция перед сменой хозяина? Презиготическая репродуктивная изоляция среди трех разновидностей специализированной мухи на одном растении-хозяине», Экологическая энтомология , 41 (4) : 389–399, Бибкод : 2016EcoEn..41..389H , doi : 10.1111/een.12309 , S2CID 87320676
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Форбс А.А., Келли П.Х., Миддлтон К.А. и Кондон М.А. (2013), «Генетически дифференцированные расы и видообразование с потоком генов у личинки подсолнечника Strauzia longipennis », Evolutionary Ecology , 27 (5): 1017–1032, Bibcode. : 2013EvEco..27.1017F , doi : 10.1007/s10682-012-9622-y , S2CID 18472327
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Вуд Т.К., Олмстед К.Л. и Гуттман С.И. (1990), «Фенология насекомых, опосредованная водными отношениями растения-хозяина», Evolution , 44 (3): 629–636, doi : 10.1111/j.1558-5646.1990.tb05943.x , ПМИД 28567982 , С2КИД 43689173
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кассо Дайну, Эрик Лоренти, Грегори Маи, Оливье Дж. Харди, Ив Бросто, Никки Тагг и Жан-Луи Дусе (2012), «Фенологические закономерности в естественной популяции тропических древесных пород Milicia excelsa (Moraceae): свидетельства изоляция во времени и ее взаимодействие со стратегиями питания расселителей» , American Journal of Botany , 99 (9): 1453–1463, doi : 10.3732/ajb.1200147 , PMID 22912370
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Табучи К., Амано Х. (2003), «Связанные с хозяином различия в характере появления, репродуктивном поведении и истории жизни Asteralobia sasakii (Monzen) (Diptera: Cecidomyiidae) между популяциями Ilex crenata и I. integra (Aquifoliaceae)», Прикладная энтомология и зоологии , 38 (4): 501–508, Bibcode : 2003AppEZ..38..501T , doi : 10.1303/aez.2003.501
- ^ А. Мосселер и К.С. Пападопол (1989), «Сезонная изоляция как репродуктивный барьер среди симпатрических видов Salix », Canadian Journal of Botany , 67 (9): 2563–2570, doi : 10.1139/b89-331.
- ^ Стефан Г. Михальски и Вальтер Дурка (2015), «Разделение во время цветения способствует поддержанию симпатрических загадочных линий растений», Ecology and Evolution , 5 (11): 2172–2184, Bibcode : 2015EcoEv...5.2172M , doi : 10.1002/ece3.1481 , PMC 4461419 , PMID 26078854
- ↑ Перейти обратно: Перейти обратно: а б Томас Макнейли и Янис Антоновичс (1968), «Эволюция в тесно соседних популяциях растений. IV. Барьеры на пути потока генов», Heredity , 23 (2): 205–218, doi : 10.1038/hdy.1968.29
- ^ Полуконова Н.В. и Кармоков М.К. (2013), «Микроэволюционные изменения в популяциях Chironomus nuditarsis Str. (Keyl, 1962) (Chironomidae, Diptera) Центрального Кавказа», Российский генетический журнал , 49 (2): 175–181, doi : 10.1134/S1022795413020099 , S2CID 12200060
- ^ Саяр Н.П., Смит К.А., Уайт И.М., Книо К.М. (2009), « Terellia fuscicornis (Diptera: Tephritidae): биологическая и морфологическая адаптация к артишоку и расторопше», Journal of Natural History , 43 (19–20): 1159–1181. , Bibcode : 2009JNatH..43.1159S , doi : 10.1080/00222930902807742 , S2CID 56004378
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Левенте Кисс, Александра Пинтье, Габор М. Ковач, Тюнде Янкович, Майкл С. Фонтейн, Ник Харви, Сянмин Сюй, Филипп С. Нико, Марк Барден, Жаки А. Шикофф и Татьяна Жиро (2011): «Временная изоляция объясняет генетическую дифференциацию, связанную с хозяином. в группе широко распространенных микопаразитических грибов» (PDF) , Molecular Ecology , 20 (7): 1492–1507, Bibcode : 2011MolEc..20.1492K , doi : 10.1111/j.1365-294X.2011.05007.x , PMID 21261766 S , 2CID 34557058
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Охигаши К., Мизугути А., Ёсимура Ю., Мацуо К. и Мива Т. (2014), «Новый метод оценки синхронности цветения для поддержки временной изоляции генетически модифицированных культур от их диких родственников», Journal of Plant Research , 127 (1 ): 109–117, Bibcode : 2014JPlR..127..109O , doi : 10.1007/s10265-013-0592-0 , PMID 24122370 , S2CID 13606001
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Скотт П. Иган, Грегори Дж. Рэгланд, Лорен Ассур, Томас Х. К. Пауэлл, Глен Р. Худ, Скотт Эмрих, Патрик Носил и Джеффри Л. Федер (2015), «Экспериментальные доказательства полногеномного воздействия экологического отбора на ранних стадиях видообразования». с потоком генов», Ecology Letters , 18 (8): 817–825, doi : 10.1111/ele.12460 , PMC 4744793 , PMID 26077935
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тейшейра Л.А.Ф. и Полаварапу С. (2003), «Эволюция фенологически различных популяций Rhagoletis mendax (Diptera: Tephritidae) на полях высокорослой черники», Анналы Энтомологического общества Америки , 96 (6): 818–827, doi : 10.1603/ 0013-8746(2003)096[0818:EOPDPO]2.0.CO;2 , S2CID 85974264
- ^ Томас Х.К. Пауэлл, Эндрю А. Форбс, Глен Р. Худ и Джеффри Л. Федер (2014), «Экологическая адаптация и репродуктивная изоляция в симпатии: генетические и фенотипические данные для местных рас-хозяев Rhagoletis pomonella », Molecular Ecology , 23 (3): 688–704, Bibcode : 2014MolEc..23..688P , doi : 10.1111/mec.12635 , PMID 24351094 , S2CID 2745741
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Глен Р. Худ, Эндрю А. Форбс, Томас Х. К. Пауэлл, Скотт П. Иган, Габриэла Хамерлинк, Джеймс Дж. Смит и Джеффри Л. Федер (2015), «Последовательное расхождение и мультипликативное происхождение разнообразия сообщества», Труды Национальной академии наук , 112 (44): E5980–E5989, Bibcode : 2015PNAS..112E5980H , doi : 10.1073/pnas.1424717112 , PMC 4640724 , PMID 26499247
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Крейг Т.П., Хорнер Дж.Д. и Итами Дж.К. (2001), «Генетика, опыт и предпочтение растения-хозяина у Eurosta Solidaginis : последствия для смены хозяев и видообразования», Evolution , 55 (4): 773–782, doi : 10.1554/ 0014-3820(2001)055[0773:GEAHPP]2.0.CO;2 , PMID 11392395 , S2CID 198157171
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Андерсон А.М., Новак С.Дж., Смит Дж.Ф., Стинхоф К., Хит Дж.А. (2016), «Фенология гнездования, выбор партнера и генетическая дивергенция в частично мигрирующей популяции американских пустельг», The Auk , 133 : 99–109, doi : 10.1642 /AUK-15-129.1 , S2CID 85683278
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Клаудия Гермес, Раэнн Меттлер, Диего Сантьяго-Аларкон, Гернот Сегельбахер и Х. Мартин Шефер (2015), «Пространственная изоляция и временные изменения в физической форме и состоянии способствуют расхождению в миграционном разделении», PLOS ONE , 10 (12): e0144264 , Bibcode : 2015PLoSO..1044264H , doi : 10.1371/journal.pone.0144264 , PMC 4681481 , PMID 26656955
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Фудикар А.М., Грейвс Т.Дж., Этвелл Дж.В., Стрикер К.А. и Кеттерсон Э.Д. (2016), «Репродуктивная аллохрония в сезонно симпатрических популяциях, поддерживаемая дифференциальной реакцией на фотопериод: последствия для дивергенции популяций и реакции на изменение климата» (PDF) , американский натуралист , 187 (4): 436–446, doi : 10.1086/685296 , PMID 27028072 , S2CID 16006406 , заархивировано из оригинала (PDF) 10 февраля 2020 г.
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ ХВ. Дэн (1997), «Фотопериодическая реакция полового размножения в группе Daphnia pulex обратная в двух различных средах обитания», Limnology and Oceanography , 42 (3): 609–611, Бибкод : 1997LimOc..42..609W , doi : 10.4319 /lo.1997.42.3.0609
- ^ Жером Сюёр и Стефан Пюиссан (2002), «Пространственная и экологическая изоляция цикад: первые данные по Tibicina (Hemiptera: Cicadoidea) во Франции», Европейский журнал энтомологии , 99 (4): 477–484, doi : 10.14411/EJE. 2002.063 , S2CID 56042738
- ^ Комацу Т. и Акимото С. (1995), «Генетическая дифференциация в результате адаптации к фенологии отдельных деревьев-хозяев у тли-раздражителя Kaltenbachiella japonica », Экологическая энтомология , 20 (1): 33–42, Bibcode : 1995EcoEn..20 ...33K , doi : 10.1111/j.1365-2311.1995.tb00426.x , S2CID 83596331
- ^ Леппянен С.А., Альтенхофер Э., Листон А.Д. и Найман Т. (2012), «Филогенетика и эволюция использования растений-хозяев у листовых пилильщиков (Hymenoptera: Tenthredinidae: Heterarthrinae)», Molecular Phylogenetics and Evolution , 64 (2): 331 –341, номер doi : 10.1016/j.ympev.2012.04.005 , PMID 22531610
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ МакКуорри С.Дж.К., Лангор Д.В. и Сперлинг ФАХ (2007), «Вариации митохондриальной ДНК у двух инвазивных березовых пилильщиков в Северной Америке», Канадский энтомолог , 139 (4): 545–553, doi : 10.4039/n06-084 , S2CID 85614594
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Scriber JM (2011), «Влияние потепления климата на перемещение гибридных зон: географически разбросанные и биологически пористые «видовые границы» », Insect Science , 18 (2): 121–159, doi : 10.1111/j.1744-7917.2010.01367 .x , S2CID 86586378
- ^ Ординг Г.Дж., Меркадер Р.Дж., Аардема М.Л. и Скрибер Дж.М. (2010), «Аллохронная изоляция и зарождающееся гибридное видообразование у тигровых бабочек-парусников», Oecologia , 162 (2): 523–531, Bibcode : 2010Oecol.162..523O , doi : 10.1007/s00442-009-1493-8 , PMID 19937057 , S2CID 22623508
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Валентина Гомес-Бахамон, Роберто Маркес, Алекс Э.Ян, Кристина Юми Мияки, Диего Т. Туэро, Оскар Лаверде-Р, Сильвия Рестрепо и Карлос Даниэль Кадена (2020), «Видообразование, связанное со сдвигами в миграционном поведении птиц в условиях радиации» ", Current Biology , 30 (7): 1312–1321, doi : 10.1016/j.cub.2020.01.064 , hdl : 11336/177388 , PMID 32197080 , S2CID 214585322
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Либхерр Дж. и Рулофс В. (1975), «Лабораторная гибридизация и исследования периода спаривания с использованием двух феромонных штаммов Ostrinia nubilalis », Анналы Энтомологического общества Америки , 68 (2): 305–309, doi : 10.1093/aesa/ 68.2.305
- ^ Томас Ю., Бетенод М.Т., Пелосуэло Л., Фреро Б. и Бурге Д. (2003), «Генетическая изоляция между двумя симпатическими расами растений-хозяев европейского кукурузного мотылька Ostrina nubilalis Hübner. I. Половой феромон, время появления моли и паразитизм Evolution (PDF) , 221734366 , 57 (2): 261–273, doi : 10.1111/ , PMID 12683523 , S2CID j.0014-3820.2003.tb00261.x
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ↑ Перейти обратно: Перейти обратно: а б Игнасио Кинтеро, Себастьян Гонсалес-Каро, Пол-Камило Саламеа и Карлос Даниэль Кадена (2014), «Асинхронность времен года: генетическая дифференциация, связанная с географическими изменениями климатической сезонности и репродуктивной фенологии», American Naturalist , 184 (3): 352– 363, номер doi : 10.1086/677261 , PMID 25141144 , S2CID 39670263
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ М. Р. Уиллиг, Д. М. Кауфман и Р. Д. Стивенс (2003), «Широтовые градиенты биоразнообразия: структура, процесс, масштаб и синтез», Ежегодный обзор экологии, эволюции и систематики , 34 : 273–309, doi : 10.1146/annurev. .ecolsys.34.012103.144032
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хиллебранд, Х. (февраль 2004 г.). «Об общности широтного градиента разнообразия» (PDF) . Американский натуралист . 163 (2): 192–211. дои : 10.1086/381004 . ПМИД 14970922 . S2CID 9886026 .
- ^ Шемске, Дуг (2009). «Биотические взаимодействия и видообразование в тропиках». В Батлине, Роджер К.; Уздечка, Джон и Шлютер, Дольф (ред.). Видообразование и закономерности разнообразия . Издательство Оксфордского университета. стр. 219–239. ISBN 9780511815683 .
- ↑ Перейти обратно: Перейти обратно: а б с д Пол Р. Мартин, Фрэнсис Бонье, Игнасио Т. Мур и Джошуа Дж. Тьюксбери (2009), «Широтовые изменения асинхронности сезонов: последствия для более высоких темпов дифференциации популяций и видообразования в тропиках», « Идеи в экологии и эволюции» , 2 : 9–17, doi : 10.4033/iee.2009.2.3.n
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Карлос Э. Гуарнизо, Паола Монтойя, Игнасио Кинтеро и Карлос Даниэль Кадена (2020), Аллохронная дивергенция, вызванная пространственной асинхронностью осадков у неотропических лягушек? , doi : 10.1101/2020.05.05.079210 , S2CID 218582518
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка )