Топоизомераза типа II
| ДНК-топоизомераза II (АТФ-гидролизующая) | |||
|---|---|---|---|
 Структура фрагмента N-концевой АТФазы массой 42 кДа и преобразовательных доменов ДНК-гиразы гомологична всем остальным топоизомеразам типа IIA. | |||
| Идентификаторы | |||
| Номер ЕС. | 5.6.2.2 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| |||
Топоизомеразы типа II — это топоизомеразы , которые одновременно разрезают обе цепи спирали ДНК, чтобы управлять ДНК клубками и суперспиралями . Они используют гидролиз АТФ , в отличие от топоизомеразы I типа . В этом процессе эти ферменты изменяют число связей кольцевой ДНК на ±2. Топоизомеразы — широко распространенные ферменты, обнаруженные во всех живых организмах. [ 1 ]
У животных мишенью химиотерапии является топоизомераза II. У прокариот гираза является антибактериальной мишенью. [ 2 ] Действительно, эти ферменты представляют интерес из-за широкого спектра эффектов.
Функция
[ редактировать ]Топоизомеразы типа II увеличивают или уменьшают число связей петли ДНК на 2 единицы, что способствует распутыванию хромосом. Например, ДНК-гираза , топоизомераза типа II, наблюдаемая у E. coli и большинства других прокариот , образует отрицательные суперспирали и уменьшает число связей на 2. Гираза также способна удалять узлы из бактериальной хромосомы . Наряду с гиразой большинство прокариот также содержат топоизомеразу второго типа IIA, называемую топоизомеразой IV. Гираза и топоизомераза IV различаются по С-концевым доменам, что, как полагают, определяет субстратную специфичность и функциональность этих двух ферментов. Футпринтинг показывает, что гираза, которая образует след из 140 пар оснований и оборачивает ДНК, образует отрицательные суперспирали , тогда как топоизомераза IV, которая образует след из 28 пар оснований, не оборачивает ДНК.
Эукариотическая топоизомераза типа II не может образовывать суперспирали; это может только расслабить их.
Роль топоизомераз типа IIB менее изучена. В отличие от топоизомераз типа IIA, топоизомеразы типа IIB не могут упростить топологию ДНК (см. Ниже), но они имеют некоторые общие структурные особенности с топоизомеразами типа IIA.
Упрощение топологии
[ редактировать ]Топоизомеразы типа IIA необходимы для разделения запутанных дочерних цепей во время репликации. Считается, что эту функцию выполняет топоизомераза II у эукариот и топоизомераза IV у прокариот. Неспособность разделить эти нити приводит к гибели клеток. Топоизомеразы типа IIA обладают особой способностью релаксировать ДНК до состояния ниже состояния термодинамического равновесия, в отличие от топоизомераз типов IA, IB и IIB. Эта способность, известная как упрощение топологии, была впервые обнаружена Рыбенковым и др. [ 3 ] Гидролиз АТФ приводит к такому упрощению, но четкий молекулярный механизм этого упрощения до сих пор отсутствует. Было предложено несколько моделей для объяснения этого явления, в том числе две модели, основанные на способности топоизомераз типа IIA распознавать изогнутые дуплексы ДНК. [ 4 ] Биохимия, электронная микроскопия и недавние структуры топоизомеразы II, связанной с ДНК, показывают, что топоизомеразы типа IIA связываются на вершинах ДНК, что подтверждает эту модель.
Классификация
[ редактировать ]В этом разделе отсутствует информация о новых членах МИБ [doi:10.1093/nargab/lqz021]. ( октябрь 2021 г. ) |
Существует два подкласса топоизомераз типа II: типа IIA и IIB.
- Топоизомеразы типа IIA включают ферменты ДНК-гиразу , эукариотическую топоизомеразу II (топо II) и бактериальную топоизомеразу IV (топо IV). Эти ферменты охватывают все сферы жизни и необходимы для функционирования. [ 5 ]
- Топоизомеразы типа IIB структурно и биохимически различны и включают одного члена семейства, топоизомеразу VI (топо VI). Топоизомеразы типа IIB обнаружены у архей и некоторых высших растений.
Некоторые организмы, включая человека, имеют две изоформы топоизомеразы II: альфа и бета . При раке топоизомераза IIα высоко экспрессируется в пролиферирующих клетках. При некоторых видах рака, таких как опухоли оболочек периферических нервов, высокая экспрессия кодируемого им белка также связана с плохой выживаемостью пациентов.
Два класса топоизомераз обладают сходным механизмом прохождения цепи и доменной структурой (см. ниже), однако они также имеют несколько важных различий. Топоизомеразы типа IIA образуют двухцепочечные разрывы с выступами из четырех пар оснований, тогда как топоизомеразы типа IIB образуют двухцепочечные разрывы с выступами из двух оснований. [ 6 ] Кроме того, топоизомеразы типа IIA способны упрощать топологию ДНК, [ 3 ] тогда как топоизомеразы типа IIB этого не делают. [ 7 ]
Структура
[ редактировать ]Тип IIA
[ редактировать ]
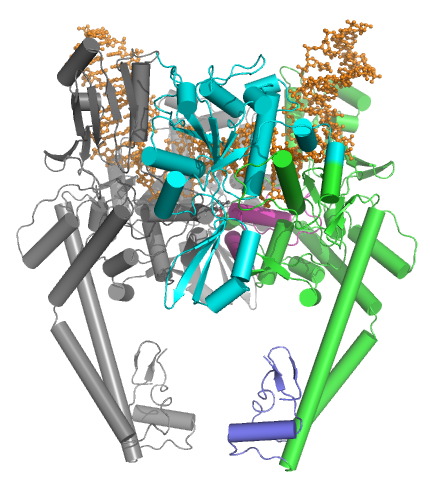
Топоизомеразы типа IIA состоят из нескольких ключевых мотивов:
- N-концевой домен АТФазы GHKL (для гиразы, Hsp, киназы и MutL),
- домен Toprim ( подкласс складки Россмана ), который существует как в топоизомеразах типа II, топоизомеразах типа IA, так и в бактериальных примазах ( DnaG ),
- центральное ДНК-связывающее ядро (которое структурно образует структуру в форме сердца) и
- вариабельный С-концевой домен.
Эукариотические топоизомеразы типа II являются гомодимерами (А 2 ), тогда как прокариотические топоизомеразы типа II являются гетеротетрамерами (А 2 В 2 ). Прокариоты имеют АТФазный домен и складку Топрима на одном полипептиде ( Pfam PF00204 ), тогда как ядро расщепления ДНК и CTD расположены на втором полипептиде ( Pfam PF00521 ). Для гиразы первый полипептид называется GyrB, а второй полипептид — GyrA. Для topo IV первый полипептид называется ParE, а второй полипептид — ParC. Обе сигнатуры Pfam обнаружены в одноцепочечной эукаиотической топоизомеразе.
Структуры N-концевого АТФазного домена гиразы [ 8 ] и дрожжевая топоизомераза II [ 9 ] были решены в комплексе с AMPPNP (аналогом АТФ), показав, что два домена АТФазы димеризуются с образованием закрытой конформации. Что касается гиразы, структура имеет значительное отверстие посередине, которое, как предполагается, предназначено для размещения Т-сегмента.
Связывание домена АТФазы со складкой Топрима представляет собой спиральный элемент, известный как домен-преобразователь. Считается, что этот домен передает нуклеотидное состояние домена АТФазы остальной части белка. Модификации этого домена влияют на активность топоизомеразы, а структурные исследования, проведенные группой Verdine, показывают, что состояние АТФ влияет на ориентацию домена-преобразователя. [ 10 ]
Центральное ядро белка содержит складку Toprim и ДНК-связывающее ядро, которое содержит домен крылатой спирали (WHD), часто называемый доменом CAP, поскольку впервые было идентифицировано, что он напоминает WHD белка-активатора катаболита. Каталитический тирозин находится на этом WHD. Складка Топрима представляет собой складку Россмана, которая содержит три инвариантных кислотных остатка, которые координируют ионы магния, участвующие в расщеплении ДНК и повторном лигировании ДНК. [ 11 ] Структура складки Топрима и ДНК-связывающего ядра дрожжевой топоизомеразы II была впервые раскрыта Бергером и Вангом. [ 12 ] а первое ядро, связывающее ДНК гиразы, было обнаружено Morais Cabral et al. [ 13 ] Структура, раскрытая Бергером, позволила получить важную информацию о функции фермента. ДНК-связывающее ядро состоит из WHD, который ведет к домену башни. Область спиральной спирали ведет к C-концевому домену, который образует основной интерфейс димера для этого кристаллического состояния (часто называемый C-затвором). В то время как исходная структура топоизомеразы II демонстрирует ситуацию, когда WHD разделены большим расстоянием, структура гиразы демонстрирует закрытую конформацию, в которой WHD закрываются.
Ядро топоизомеразы II позже было решено в новых конформациях, в том числе Fass et al. [ 14 ] и один от Dong et al. [ 15 ] Структура Fass показывает, что домен Toprim является гибким и что эта гибкость может позволить домену Toprim координироваться с WHD с образованием компетентного комплекса расщепления. В конечном итоге это было подтверждено Dong et al. структура, которая была решена в присутствии ДНК. Эта последняя структура показала, что домен Toprim и WHD образуют комплекс расщепления, очень похожий на комплекс топоизомераз типа IA, и указала на то, как связывание ДНК и расщепление могут быть разделены, а структура показала, что ДНК изгибалась на ~ 150 градусов через инвариантный изолейцин (у топоизомеразы II — I833, у гиразы — I172). Этот механизм изгиба очень похож на механизм интеграции фактора хозяина (IHF) и HU, двух архитектурных белков бактерий. Кроме того, в то время как предыдущие структуры ДНК-связывающего ядра имели закрытые C-ворота, эта структура захватывала ворота открытыми, что является ключевым этапом в двухворотном механизме (см. ниже).
Совсем недавно было решено несколько структур ДНК-связанной структуры в попытке понять как химический механизм расщепления ДНК, так и структурную основу ингибирования топоизомеразы антибактериальными ядами. Первая полная архитектура ДНК-гиразы E. coli была раскрыта с помощью криоэлектронной микроскопии с разрешением, близким к атомному. [ 16 ] Нуклеопротеиновый комплекс был захвачен с помощью длинного дуплекса ДНК и гепотидацина , нового ингибитора бактериальной топоизомеразы.
С-концевая область прокариотических топоизомераз определена для нескольких видов. Первая структура С-концевого домена гиразы была раскрыта Corbett et al. [ 17 ] а С-концевой домен топоизомеразы IV был решен Corbett et al. [ 7 ] Структуры сформировали новый бета-цилиндр, который изгибает ДНК, обертывая нуклеиновую кислоту вокруг себя. Изгиб ДНК под действием гиразы был предложен как ключевой механизм способности гиразы создавать отрицательные суперспирали в ДНК. Это согласуется с данными о следе, которые показывают, что гираза имеет след из 140 пар оснований. CTD как гиразы, так и топоизомеразы IV изгибают ДНК, но только гираза образует отрицательные суперспирали.
В отличие от функции С-концевого домена прокариотических топоизомераз, функция С-концевого участка эукариотической топоизомеразы II до сих пор не ясна. Исследования показали, что эта область регулируется фосфорилированием, и это модулирует активность топоизомеразы, однако для изучения этого необходимо провести дополнительные исследования.
Тип IIВ
[ редактировать ]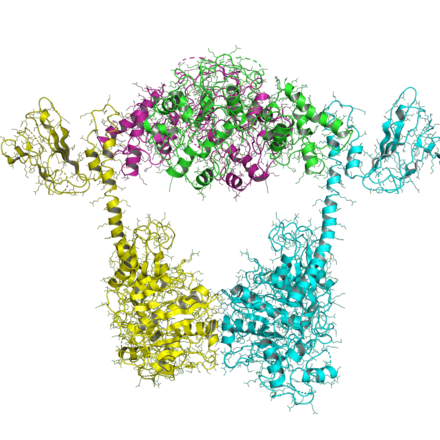
Организация топоизомераз типа IIB аналогична организации топоизомераз типа IIA, за исключением того, что все типы IIB имеют два гена и образуют гетеротетрамеры. Один ген, названный topo VI-B (поскольку он напоминает gyrB), содержит домен АТФазы, домен преобразователя ( Pfam PF09239 ) и C-концевой Ig-фолд-подобный домен H2TH ( Pfam PF18000 ). Второй ген, названный topo VI-A ( Pfam PF04406 ), содержит WHD и домен Toprim.
АТФазный домен топо VI B был решен в нескольких нуклеотидных состояниях. [ 18 ] Он очень похож на домен GHKL topo II и MutL и показывает, что состояние нуклеотидов (ADP или ATP) влияет на ориентацию домена преобразователя (и 1MX0).
Структура топо VI-A была раскрыта Bergerat et al. [ 19 ] показывая, что складка HTH и Toprim имела новую конформацию по сравнению с topo IIA.
Была расшифрована недавняя структура комплекса topo VI A/B, показывающая открытую и закрытую конформацию, два состояния, которые предсказаны в двухворотном механизме (см. ниже). Эти структуры, одна из которых представляет собой рентгеновскую кристаллическую структуру, а другая представляет собой реконструкцию малоуглового рентгеновского рассеяния (SAXS), показывают, что домен АТФазы может быть либо открытым, либо закрытым. [ 20 ]
Механизм действия
[ редактировать ]Береговой проход
[ редактировать ]Топоизомераза типа IIA действует посредством «двухворотного» механизма (хотя это историческое обозначение), механизма, поддерживаемого биохимией. [ 21 ] а также структурной работой. [ 22 ]
Нить ДНК, называемая воротами или G-сегментом, связана центральными воротами связывания ДНК (ДНК-воротами). Вторая цепь ДНК, называемая транспортной, или Т-сегментом, захватывается в результате димеризации N-концевого домена АТФазы (ворота АТФазы), когда связываются две молекулы АТФ. Гидролиз АТФ и высвобождение неорганического фосфата приводит к расщеплению G-сегмента, поскольку каталитические тирозины образуют ковалентную фосфотирозиновую связь с 5'-концом ДНК. Это создает выступ из четырех оснований и двухцепочечный разрыв G-сегмента. Когда ворота связывания ДНК разделяются, Т-сегмент передается через G-сегмент. G-сегмент запечатан, что приводит к открытию C-концевых ворот (или C-ворот), позволяя освободить Т-сегмент. Выпуск продукта ADP приводит к перезагрузке системы и позволяет захватить второй Т-сегмент.
Топоизомеразы типа IIB действуют аналогичным образом, за исключением того, что белок образует выступ из двух оснований в G-сегменте, а С-концевые ворота полностью отсутствуют.
расщепление ДНК
[ редактировать ]В механизме прохождения цепи расщепление ДНК является ключевым моментом, позволяющим Т-сегменту пройти через G-сегмент. Механизм расщепления ДНК топоизомеразами типа IIA в последнее время находится в центре внимания многих биохимических и структурно-биологических исследований.
катенация
[ редактировать ]Катенация — это процесс, при котором две кольцевые цепи ДНК соединяются друг с другом, как звенья цепи. Это происходит после репликации ДНК, когда две одиночные цепи соединяются и все еще могут реплицироваться, но не могут разделиться на две дочерние клетки. Поскольку топоизомеразы II типа разрывают двойную цепь, они могут зафиксировать это состояние (топоизомеразы I типа могли бы сделать это только в том случае, если уже был одноцепочечный разрыв), и правильное число хромосом может остаться в дочерних клетках. Линейная ДНК у эукариот настолько длинная, что ее можно считать не имеющей концов; Топоизомеразы типа II необходимы по той же причине.
Торможение
[ редактировать ]Малые молекулы, нацеленные на топоизомеразу типа II, делятся на два класса: ингибиторы и яды. Из-за их частого присутствия в пролиферирующих эукариотических клетках ингибиторы топоизомераз типа II широко изучались и использовались в качестве противораковых препаратов. [ 23 ]
- Ингибиторы топоизомеразы типа II включают HU-331 , ICRF-187 , ICRF-193 и митиндомид . Эти молекулы действуют путем ингибирования активности АТФазы, действуя как неконкурентные ингибиторы АТФ. Это было показано структурными исследованиями. [ 9 ] и биохимические исследования, проведенные группой Линдсли.
- Яды топоизомераз II типа включают доксорубицин , этопозид , новобиоцин , хинолоны (в том числе ципрофлоксацин ) и тенипозид . Эти небольшие молекулы нацелены на комплекс ДНК-белок. Некоторые из этих молекул приводят к усилению расщепления, тогда как другие, такие как этопозид, ингибируют повторное лигирование.
Экспериментальный противоопухолевый препарат м-АМСА (4'-(9'-акридиниламино)метансульфон-м-анизидид) также ингибирует топоизомеразу 2 типа. [ 24 ]
Яды топоизомеразы широко используются как в противораковых, так и в антибактериальных целях. Хотя антибактериальные соединения, такие как ципрофлоксацин, нацелены на бактериальную гиразу, они не способны ингибировать эукариотические топоизомеразы типа IIA. Кроме того, устойчивые к лекарствам бактерии часто имеют точечную мутацию гиразы (серин79аланин в E. coli ), которая делает хинолоны неэффективными. [ нужна ссылка ] Недавние структурные исследования привели к открытию соединения, которое больше не зависит от этого остатка и, следовательно, обладает эффективностью против устойчивых к лекарствам бактерий. [ нужна ссылка ]
Гираза бактериофага Т4
[ редактировать ]Гираза бактериофага (фага) Т4 (топоисмераза II типа) представляет собой многосубъединичный белок, состоящий из продуктов генов 39, 52 и, вероятно, 60. [ 25 ] [ 26 ] Он катализирует релаксацию отрицательно или положительно суперспиральной ДНК и используется в репликации ДНК фага во время заражения бактериального хозяина E. coli . [ 27 ] Белок фагового гена 52 имеет гомологию с E. coli. субъединицей gyrA гиразы [ 28 ] а белок фагового гена 39 имеет гомологию с субъединицей gyr B. [ 29 ] хозяина Поскольку ДНК-гираза E. coli может частично компенсировать потерю продуктов гена фага Т4, мутанты с дефектами в генах 39, 52 или 60 не полностью отменяют репликацию ДНК фага, а скорее задерживают ее инициацию. [ 27 ] Скорость элонгации ДНК при таких мутантных инфекциях не ниже, чем у дикого типа. [ 30 ] Мутанты с дефектами в генах 39, 52 или 60 демонстрируют повышенную генетическую рекомбинацию , а также повышенную мутацию замены оснований и делеции, что позволяет предположить, что компенсируемый хозяином синтез ДНК менее точен, чем синтез, направляемый фагом дикого типа. [ 31 ] Мутант с дефектом гена 39 демонстрирует повышенную чувствительность к инактивации ультрафиолетовым множественные копии фаговой хромосомы . облучением на стадии фаговой инфекции после инициации репликации ДНК, когда присутствуют [ 32 ] Мутанты с дефектами генов 39, 52 и 60 имеют пониженную способность осуществлять реактивацию множественности — форму рекомбинационной репарации, которая может справляться с различными типами повреждений ДНК. [ 33 ] Гираза, специфичная в геноме неинфицированной E. coli , также, по-видимому, участвует в рекомбинационной репарации, обеспечивая точку инициации взаимного обмена цепей, управляемого белком RecA. [ 34 ]
Ссылки
[ редактировать ]- ^ Дьюиз Дж. Э., Ошерофф Н. (февраль 2009 г.). «Реакция расщепления ДНК топоизомеразой II: волк в овечьей шкуре» . Исследования нуклеиновых кислот . 37 (3): 738–748. дои : 10.1093/нар/gkn937 . ПМК 2647315 . ПМИД 19042970 .
- ^ Рис Р.Дж., Максвелл А. (1991). «ДНК-гираза: строение и функции». Критические обзоры по биохимии и молекулярной биологии . 26 (3–4): 335–375. дои : 10.3109/10409239109114072 . ПМИД 1657531 .
- ^ Jump up to: а б Рыбенков В.В., Улльспергер С., Вологодский А.В., Коцарелли Н.Р. (август 1997 г.). «Упрощение топологии ДНК ниже равновесных значений топоизомеразами типа II». Наука . 277 (5326). Нью-Йорк, штат Нью-Йорк: 690–3. дои : 10.1126/science.277.5326.690 . ПМИД 9235892 .
- ^ Вологодский А.В., Чжан В., Рыбенков В.В., Подтележников А.А., Субраманиан Д., Гриффит Дж.Д., Коццарелли Н.Р. (март 2001 г.). «Механизм упрощения топологии ДНК-топоизомеразами типа II» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (6): 3045–9. Бибкод : 2001PNAS...98.3045V . дои : 10.1073/pnas.061029098 . ПМК 30604 . ПМИД 11248029 .
- ^ Рис Р.Дж., Максвелл А. (январь 1991 г.). «ДНК-гираза: строение и функции». Критические обзоры по биохимии и молекулярной биологии . 26 (3–4): 335–75. дои : 10.3109/10409239109114072 . ПМИД 1657531 .
- ^ Бюлер С., Леббинк Дж. Х., Бокс С., Ладенштайн Р., Фортер П. (октябрь 2001 г.). «ДНК-топоизомераза VI генерирует АТФ-зависимые двухцепочечные разрывы с двумя нуклеотидными выступами» . Журнал биологической химии . 276 (40): 37215–22. дои : 10.1074/jbc.M101823200 . ПМИД 11485995 . S2CID 24354635 .
- ^ Jump up to: а б ПДБ : 1звт ; Корбетт К.Д., Шеффлер А.Дж., Томсен Н.Д., Бергер Дж.М. (август 2005 г.). «Структурные основы субстратной специфичности ДНК-топоизомеразы IV». Журнал молекулярной биологии . 351 (3): 545–61. дои : 10.1016/j.jmb.2005.06.029 . ПМИД 16023670 .
- ^ Вигли Д.Б., Дэвис Г.Дж., Додсон Э.Дж., Максвелл А., Додсон Дж. (июнь 1991 г.). «Кристаллическая структура N-концевого фрагмента белка ДНК-гиразы B». Природа . 351 (6328): 624–9. Бибкод : 1991Natur.351..624W . дои : 10.1038/351624a0 . ПМИД 1646964 . S2CID 4373125 .
- ^ Jump up to: а б ПДБ : 1ПВГ ; Классен С., Олланд С., Бергер Дж. М. (сентябрь 2003 г.). «Структура АТФазной области топоизомеразы II и механизм ее ингибирования химиотерапевтическим средством ICRF-187» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (19): 10629–34. Бибкод : 2003PNAS..10010629C . дои : 10.1073/pnas.1832879100 . ЧВК 196855 . ПМИД 12963818 .
- ^ Вэй Х., Рутенбург А.Дж., Бечис С.К., Вердин Г.Л. (ноябрь 2005 г.). «Нуклеотид-зависимое движение домена в АТФазном домене ДНК-топоизомеразы типа IIA человека» (PDF) . Журнал биологической химии . 280 (44): 37041–7. дои : 10.1074/jbc.M506520200 . ПМИД 16100112 . S2CID 35186716 .
- ^ Аравинд Л., Лейпе Д.Д., Кунин Е.В. (сентябрь 1998 г.). «Топрим - консервативный каталитический домен в топоизомеразах типа IA и II, примазах типа DnaG, нуклеазах семейства OLD и белках RecR» . Исследования нуклеиновых кислот . 26 (18): 4205–13. дои : 10.1093/нар/26.18.4205 . ПМЦ 147817 . ПМИД 9722641 .
- ^ ВВП : 1BGW ; Бергер Дж. М., Гамблин С. Дж., Харрисон С. К., Ван Дж. К. (январь 1996 г.). «Структура и механизм ДНК-топоизомеразы II». Природа . 379 (6562): 225–32. Бибкод : 1996Natur.379..225B . дои : 10.1038/379225a0 . ПМИД 8538787 . S2CID 4360011 .
- ^ ПДБ : 1AB4 ; Мораиш Кабрал Дж. Х., Джексон А. П., Смит К. В., Шикотра Н., Максвелл А., Лиддингтон Р. К. (август 1997 г.). «Кристаллическая структура домена разрыва-воссоединения ДНК-гиразы» . Природа . 388 (6645): 903–6. Бибкод : 1997Natur.388..903M . дои : 10.1038/42294 . ПМИД 9278055 . S2CID 4320715 .
- ^ PDB : 1БДжТ ; Фасс Д., Богден С.Э., Бергер Дж.М. (апрель 1999 г.). «Четвертичные изменения в топоизомеразе II могут направлять ортогональное движение двух цепей ДНК». Структурная биология природы . 6 (4): 322–6. дои : 10.1038/7556 . ПМИД 10201398 . S2CID 947461 .
- ^ PDB : 2RGR ; Донг К.К., Бергер Дж.М. (декабрь 2007 г.). «Структурные основы распознавания и изгибания воротной ДНК топоизомеразами типа IIA». Природа . 450 (7173): 1201–5. Бибкод : 2007Natur.450.1201D . дои : 10.1038/nature06396 . ПМИД 18097402 . S2CID 1756317 .
- ^ Ванден Брук А., Лотц С., Ортис Дж., Ламур В. (октябрь 2019 г.). «Крио-ЭМ структура полного нуклеопротеинового комплекса ДНК-гиразы E. coli» . Природные коммуникации . 10 (1): 4935. Бибкод : 2019NatCo..10.4935V . дои : 10.1038/s41467-019-12914-y . ПМК 6821735 . ПМИД 31666516 .
- ^ PDB : 1SU ; Корбетт К.Д., Шульцабергер Р.К., Бергер Дж.М. (май 2004 г.). «С-концевой домен ДНК-гиразы А имеет складку бета-вертушки, изгибающую ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (19): 7293–8. Бибкод : 2004PNAS..101.7293C . дои : 10.1073/pnas.0401595101 . ПМК 409912 . ПМИД 15123801 .
- ^ ПДБ : 1MU5 ; Корбетт К.Д., Бергер Дж.М. (январь 2003 г.). «Структура субъединицы топоизомеразы VI-B: значение для механизма и эволюции топоизомеразы II типа» . Журнал ЭМБО . 22 (1): 151–63. дои : 10.1093/emboj/cdg008 . ПМК 140052 . ПМИД 12505993 .
- ^ Бергера А, де Масси Б, Гадель Д, Варутас ПК, Николя А, Фортер П (март 1997 г.). «Атипичная топоизомераза II из архей, имеющая значение для мейотической рекомбинации». Природа . 386 (6623): 414–7. Бибкод : 1997Natur.386..414B . дои : 10.1038/386414a0 . ПМИД 9121560 . S2CID 4327493 .
- ^ PDB : 2Q2E ; Корбетт К.Д., Бенедетти П., Бергер Дж.М. (июль 2007 г.). «Сборка голофермента и АТФ-опосредованная конформационная динамика топоизомеразы VI». Структурная и молекулярная биология природы . 14 (7): 611–9. дои : 10.1038/nsmb1264 . ПМИД 17603498 . S2CID 2159631 .
- ^ Рока Дж., Ван Дж.К. (май 1994 г.). «Транспорт ДНК с помощью ДНК-топоизомеразы типа II: свидетельства в пользу двухворотного механизма». Клетка . 77 (4): 609–16. дои : 10.1016/0092-8674(94)90222-4 . ПМИД 8187179 . S2CID 19776252 .
- ^ Бергер Дж. М., Ван Дж. К. (февраль 1996 г.). «Последние разработки в области структуры и механизма ДНК-топоизомеразы II». Современное мнение в области структурной биологии . 6 (1): 84–90. дои : 10.1016/s0959-440x(96)80099-6 . ПМИД 8696977 .
- ^ Альбертс Б (18 ноября 2014 г.). Молекулярная биология клетки (Шестое изд.). Нью-Йорк, штат Нью-Йорк. ISBN 978-0-8153-4432-2 . OCLC 887605755 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Уиллмор Э., де Ко С., Сантер, Нью-Джерси, Тилби М.Дж., Джексон Г.Х., Остин Калифорния, Дуркач Б.В. (июнь 2004 г.). «Новый ДНК-зависимый ингибитор протеинкиназы NU7026 усиливает цитотоксичность ядов топоизомеразы II, используемых при лечении лейкемии» . Кровь . 103 (12): 4659–65. дои : 10.1182/кровь-2003-07-2527 . ПМИД 15010369 .
- ^ Лю Л.Ф. , Лю CC, Альбертс Б.М. (октябрь 1979 г.). «ДНК-топоизомераза Т4: новый АТФ-зависимый фермент, необходимый для инициации репликации ДНК бактериофага Т4». Природа . 281 (5731): 456–61. Бибкод : 1979Natur.281..456L . дои : 10.1038/281456a0 . ПМИД 226889 . S2CID 4343962 .
- ^ Стетлер Г.Л., Кинг Г.Дж., Хуан В.М. (август 1979 г.). «Белки задержки ДНК Т4, необходимые для специфической репликации ДНК, образуют комплекс, обладающий АТФ-зависимой ДНК-топоизомеразной активностью» . Труды Национальной академии наук Соединенных Штатов Америки . 76 (8): 3737–41. Бибкод : 1979PNAS...76.3737S . дои : 10.1073/pnas.76.8.3737 . ПМЦ 383908 . ПМИД 226976 .
- ^ Jump up to: а б Маккарти Д. (январь 1979 г.). «Гиразозависимая инициация репликации ДНК бактериофага Т4: взаимодействие гиразы Escherichia coli с новобиоцином, кумермицином и продуктами генов задержки ДНК фага». Журнал молекулярной биологии . 127 (3): 265–83. дои : 10.1016/0022-2836(79)90329-2 . ПМИД 372540 .
- ^ Хуан В.М. (сентябрь 1986 г.). «52-белковая субъединица ДНК-топоизомеразы Т4 гомологична белку gyrA гиразы» . Исследования нуклеиновых кислот . 14 (18): 7379–90. ПМК 311757 . ПМИД 3020513 .
- ^ Хуан В.М. (октябрь 1986 г.). «Нуклеотидная последовательность гена ДНК-топоизомеразы типа II. Ген 39 бактериофага Т4» . Исследования нуклеиновых кислот . 14 (19): 7751–65. дои : 10.1093/нар/14.19.7751 . ПМК 311794 . ПМИД 3022233 .
- ^ Маккарти Д., Миннер С., Бернштейн Х., Бернштейн С. (октябрь 1976 г.). «Скорость элонгации ДНК и распределение точек роста фага Т4 дикого типа и янтарного мутанта с задержкой ДНК». Журнал молекулярной биологии . 106 (4): 963–81. дои : 10.1016/0022-2836(76)90346-6 . ПМИД 789903 .
- ^ Муфтий С., Бернштейн Х. (октябрь 1974 г.). «Мутанты с задержкой ДНК бактериофага Т4» . Журнал вирусологии . 14 (4): 860–71. doi : 10.1128/JVI.14.4.860-871.1974 . ПМЦ 355592 . ПМИД 4609406 .
- ^ Хайман П. (август 1993 г.). «Генетика эффекта Лурии-Латарже в бактериофаге Т4: доказательства участия множественных путей репарации ДНК» . Генетические исследования . 62 (1): 1–9. дои : 10.1017/s0016672300031499 . ПМИД 8405988 .
- ^ Мискиминс Р., Шнайдер С., Джонс В., Бернштейн Х. (июнь 1982 г.). «Участие топоизомеразы в множественной реактивации фага Т4» . Генетика . 101 (2): 157–77. doi : 10.1093/genetics/101.2.157 (неактивен 31 января 2024 г.). ПМК 1201854 . ПМИД 6293912 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Кассуто Э. (сентябрь 1984 г.). «Образование ковалентно замкнутой гетеродуплексной ДНК совместным действием гиразы и белка RecA» . Журнал ЭМБО . 3 (9): 2159–64. дои : 10.1002/j.1460-2075.1984.tb02106.x . ПМК 557658 . ПМИД 6092061 .
Дальнейшее чтение
[ редактировать ]Внешние ссылки
[ редактировать ]- ДНК + топоизомеразы + Тип + II в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)