Тубулин
| Тубулин | |||
|---|---|---|---|
 Структура комплекса головка-микротрубочки kif1a в АТП-форме | |||
| Идентификаторы | |||
| Символ | Тубулин | ||
| Пфам | PF00091 | ||
| Пфам Клан | CL0442 | ||
| ИнтерПро | ИПР003008 | ||
| PROSITE | PDOC00201 | ||
| ОБЛАСТЬ ПРИМЕНЕНИЯ 2 | 1туба / ОБЛАСТЬ ПРИМЕНЕНИЯ / СУПФАМ | ||
| |||
Тубулин в молекулярной биологии может относиться либо к суперсемейству тубулиновых белков глобулярных белков , либо к одному из белков-членов этого суперсемейства. α- и β-тубулины полимеризуются в микротрубочки , основной компонент эукариот цитоскелета . [ 1 ] Он был открыт и назван Хидео Мори в 1968 году. [ 2 ] Микротрубочки участвуют во многих важных клеточных процессах, включая митоз . Тубулин-связывающие препараты убивают раковые клетки, ингибируя динамику микротрубочек, которая необходима для ДНК сегрегации и, следовательно, деления клеток .
У эукариот имеется шесть членов суперсемейства тубулинов, хотя не все они присутствуют у всех видов. [ 3 ] [ 4 ] И α, и β тубулины имеют массу около 50 кДа и, таким образом, находятся в том же диапазоне, что и актин (с массой ~ 42 кДа). Напротив, полимеры тубулина (микротрубочки), как правило, намного больше актиновых нитей из-за их цилиндрической природы.
Долгое время считалось, что тубулин свойственен только эукариотам. Однако совсем недавно было показано, что несколько прокариотических белков связаны с тубулином. [ 5 ] [ 6 ] [ 7 ] [ 8 ]
Характеристика
[ редактировать ]Тубулин характеризуется эволюционно консервативным семейством тубулинов/FtsZ, ГТФазы белковым доменом .
Этот белковый домен ГТФазы обнаружен во всех цепях тубулина эукариот. [ 9 ] а также бактериальный белок TubZ, [ 8 ] архейный белок CetZ, [ 10 ] и семейство белков FtsZ , широко распространенное у бактерий и архей . [ 5 ] [ 11 ]
Функция
[ редактировать ]Микротрубочки
[ редактировать ]
α- и β-тубулины полимеризуются в динамические микротрубочки. У эукариот микротрубочки являются одним из основных компонентов цитоскелета и участвуют во многих процессах, включая структурную поддержку, внутриклеточный транспорт и сегрегацию ДНК .
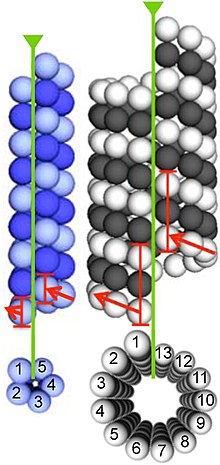
Микротрубочки собраны из димеров α- и β-тубулина. Эти субъединицы слегка кислые, с изоэлектрической точкой от 5,2 до 5,8. [ 14 ] Каждый из них имеет молекулярную массу примерно 50 кДа. [ 15 ]
Для формирования микротрубочек димеры α- и β-тубулина связываются с GTP и собираются на (+)-концах микротрубочек, находясь в GTP-связанном состоянии. [ 16 ] Субъединица β-тубулина экспонируется на плюс-конце микротрубочки, а субъединица α-тубулина — на минус-конце. После того, как димер включен в микротрубочку, молекула GTP, связанная с субъединицей β-тубулина, в конечном итоге гидролизуется до GDP через междимерные контакты вдоль протофиламента микротрубочки . [ 17 ] Молекула GTP, связанная с субъединицей α-тубулина, не гидролизуется в течение всего процесса. Связан ли β-тубулиновый член димера тубулина с GTP или GDP, влияет на стабильность димера в микротрубочках. Димеры, связанные с GTP, имеют тенденцию собираться в микротрубочки, тогда как димеры, связанные с GDP, имеют тенденцию распадаться; таким образом, этот цикл GTP важен для динамической нестабильности микротрубочек.
Бактериальные микротрубочки
[ редактировать ]Гомологи α- и β-тубулина идентифицированы у Prosthecobacter бактерий рода . [ 6 ] Они обозначены BtubA и BtubB, чтобы идентифицировать их как бактериальные тубулины. Оба обладают гомологией как с α-, так и с β-тубулином. [ 18 ] Хотя структурно они очень похожи на эукариотические тубулины, они обладают несколькими уникальными особенностями, включая сворачивание без шаперонов и слабую димеризацию. [ 19 ] Криогенная электронная микроскопия показала, что BtubA/B образует микротрубочки in vivo , и предположила, что эти микротрубочки содержат только пять протофиламентов, в отличие от эукариотических микротрубочек, которые обычно содержат 13. [ 13 ] Последующие исследования in vitro показали, что BtubA/B образует четырехцепочечные «мини-микротрубочки». [ 20 ]
сегрегация ДНК
[ редактировать ]Деление клеток
[ редактировать ]Прокариотический отдел
[ редактировать ]FtsZ обнаружен почти у всех бактерий и архей , где он участвует в делении клеток , локализуясь в кольце в середине делящейся клетки и вовлекая в работу другие компоненты дивисомы, группы белков, которые вместе сужают клеточную оболочку, отщипывая ее. ячейку, давая две дочерние клетки. FtsZ может полимеризоваться в трубки, листы и кольца in vitro и образовывать динамические нити in vivo .
с низким числом копий Функция TubZ заключается в разделении плазмид во время деления бактериальных клеток. Белок образует структуру, необычную для гомолога тубулина; две спиральные нити обвивают друг друга. [ 21 ] Это может отражать оптимальную структуру для этой роли, поскольку неродственный белок ParM , разделяющий плазмиды, демонстрирует аналогичную структуру. [ 22 ]
Форма ячейки
[ редактировать ]CetZ участвует в изменении формы клеток у плеоморфных Haloarchaea . У Haloferax volcanii CetZ образует динамические цитоскелетные структуры, необходимые для дифференцировки из пластинчатой формы клеток в палочковидную форму, проявляющую плавательную подвижность. [ 10 ]
Типы
[ редактировать ]Эукариотический
[ редактировать ]Суперсемейство тубулинов включает шесть семейств (альфа-(α), бета-(β), гамма-(γ), дельта-(δ), эпсилон-(ε) и зета-(ζ) тубулины). [ 23 ]
α-Тубулин
[ редактировать ]Подтипы альфа-тубулина человека включают: [ нужна ссылка ]
β-тубулин
[ редактировать ]
Все лекарства, которые, как известно, связываются с тубулином человека, связываются с β-тубулином. [ 24 ] К ним относятся паклитаксел , колхицин и барвинка алкалоиды , каждый из которых имеет отдельный участок связывания на β-тубулине. [ 24 ]
Кроме того, некоторые противогельминтные препараты преимущественно нацелены на колхициновый участок β-тубулина у червей, а не у высших эукариот. Хотя мебендазол все еще сохраняет некоторое сродство связывания с человека и дрозофилы , β-тубулином [ 25 ] альбендазол почти исключительно связывается с β-тубулином червей и других низших эукариот. [ 26 ] [ 27 ]
β-тубулин класса III представляет собой элемент микротрубочек, экспрессирующийся исключительно в нейронах . [ 28 ] и является популярным идентификатором, специфичным для нейронов нервной ткани. Он связывает колхицин гораздо медленнее, чем другие изотипы β-тубулина. [ 29 ]
β1-тубулин , иногда называемый β-тубулином класса VI, [ 30 ] является наиболее дивергентным на уровне аминокислотной последовательности. [ 31 ] Он экспрессируется исключительно в мегакариоцитах и тромбоцитах у человека и, по-видимому, играет важную роль в образовании тромбоцитов. [ 31 ] Когда β-тубулин класса VI экспрессируется в клетках млекопитающих, он вызывает разрушение сети микротрубочек, образование фрагментов микротрубочек и в конечном итоге может вызывать структуры, подобные маргинальным полосам, присутствующие в мегакариоцитах и тромбоцитах. [ 32 ]
Катанин представляет собой белковый комплекс, который разрывает микротрубочки по субъединицам β-тубулина и необходим для быстрого транспорта микротрубочек в нейронах и у высших растений. [ 33 ]
Подтипы β-тубулинов человека включают: [ нужна ссылка ]
γ-тубулин
[ редактировать ]
γ-Тубулин, еще один член семейства тубулинов, играет важную роль в зарождении и полярной ориентации микротрубочек. Он обнаруживается преимущественно в центросомах и телах полюсов веретена , поскольку это области наиболее обильного зарождения микротрубочек. В этих органеллах несколько молекул γ-тубулина и других белков находятся в комплексах, известных как кольцевые комплексы γ-тубулина (γ-TuRC), которые химически имитируют (+) конец микротрубочки и, таким образом, позволяют микротрубочкам связываться. γ-тубулин также был выделен в виде димера и как часть небольшого комплекса γ-тубулина (γTuSC), промежуточного по размеру между димером и γTuRC. γ-тубулин является наиболее изученным механизмом зарождения микротрубочек, но некоторые исследования показали, что определенные клетки могут быть способны адаптироваться к его отсутствию, о чем свидетельствуют исследования мутаций и РНКи, которые ингибируют его правильную экспрессию. Помимо образования γ-TuRC для зарождения и организации микротрубочек, γ-тубулин может полимеризоваться в нити, которые собираются в пучки и сети. [ 34 ]
Подтипы человеческого γ-тубулина включают:
Члены кольцевого комплекса γ-тубулина:
d и e-тубулин
[ редактировать ]Было обнаружено, что дельта (δ) и эпсилон (ε) тубулин локализуются в центриолях и могут играть роль в структуре и функции центриолей, хотя ни один из них не изучен так хорошо, как α- и β-формы.
Гены δ- и ε-тубулина человека включают: [ нужна ссылка ]
з-тубулин
[ редактировать ]Дзета-тубулин ( IPR004058 ) присутствует у многих эукариот, но отсутствует у других, включая плацентарных млекопитающих. Было показано, что это связано со структурой центриолей базальной ножки в многореснитчатых эпителиальных клетках. [ 4 ]
Прокариотический
[ редактировать ]БтубА/Б
[ редактировать ]BtubA ( Q8GCC5 ) и BtubB ( Q8GCC1 ) обнаружены у некоторых видов бактерий Verrucomicrobiota рода Prosthecobacter . [ 6 ] Их эволюционное родство с эукариотическими тубулинами неясно, хотя они, возможно, произошли от эукариотической линии путем латерального переноса генов . [ 19 ] [ 18 ] По сравнению с другими бактериальными гомологами они гораздо больше похожи на эукариотические тубулины. В собранной структуре BtubB действует как α-тубулин, а BtubA действует как β-тубулин. [ 35 ]
ФтсЗ
[ редактировать ]Многие бактериальные и эвриархеотные клетки используют FtsZ для деления посредством бинарного деления . Все хлоропласты и некоторые митохондрии , обе органеллы, полученные в результате эндосимбиоза бактерий, также используют FtsZ. [ 36 ] Это был первый идентифицированный прокариотический цитоскелетный белок.
ТубЗ
[ редактировать ]TubZ ( Q8KNP3 ; pBt156) был идентифицирован у Bacillus thuringiensis как необходимый для поддержания плазмиды . [ 8 ] Он связывается с ДНК-связывающим белком под названием TubR ( Q8KNP2 ; pBt157), перетягивая плазмиду. [ 37 ]
ЦетЗ
[ редактировать ]CetZ ( D4GVD7 ) обнаружен в эуриархейных кладах Methanomicrobia и Halobacteria , где он участвует в дифференциации формы клеток. [ 10 ]
Фаговые тубулины
[ редактировать ]Фаги рода Phikzlikevirus , а также фаг Serratia PCH45 используют белок оболочки ( Q8SDA8 ) для построения ядроподобной структуры, называемой фаговым ядром. Эта структура включает в себя ДНК, а также механизмы репликации и транскрипции. Он защищает ДНК фага от защитных сил хозяина, таких как ферменты рестрикции и CRISPR системы -Cas типа I. Тубулин , образующий веретено , называемый по-разному PhuZ ( B3FK34 ) и gp187 , центрирует ядро в клетке. [ 38 ] [ 39 ]
Тубулин одинархеоты
[ редактировать ]Тубулин архей Асгарда из гидротермальных обитателей Odinarchaeota (OdinTubulin) был идентифицирован как настоящий тубулин. OdinTubulin образует протомеры и протофиламенты, наиболее похожие на эукариотические микротрубочки, но при этом собирается в кольцевые системы, более похожие на FtsZ , что указывает на то, что OdinTubulin может представлять собой промежуточное звено в эволюции между FtsZ и тубулинами, образующими микротрубочки. [ 40 ]
Фармакология
[ редактировать ]Тубулины — мишень для противораковых препаратов [ 41 ] [ 42 ] [ 43 ] такие как винбластин и винкристин , [ 44 ] [ 45 ] и паклитаксел . [ 46 ] Противогельминтные препараты мебендазол и альбендазол, а также против подагры средство колхицин связываются с тубулином и ингибируют образование микротрубочек. В то время как первое в конечном итоге приводит к гибели клеток червей, второе останавливает подвижность нейтрофилов и уменьшает воспаление у людей. Противогрибковый препарат гризеофульвин воздействует на образование микротрубочек и применяется при лечении рака.
Посттрансляционные модификации
[ редактировать ]При включении в микротрубочки тубулин накапливает ряд посттрансляционных модификаций , многие из которых уникальны для этих белков. Эти модификации включают детирозинирование , ацетилирование , полиглутамилирование , полиглицилирование , фосфорилирование , убиквитинирование , сумойлирование и пальмитоилирование . Тубулин также склонен к окислительной модификации и агрегации, например, во время острого клеточного повреждения. [ 47 ]
В настоящее время проводится множество научных исследований ацетилирования, осуществляемого в некоторых микротрубочках, особенно ацетилирования с помощью α-тубулина N-ацетилтрансферазы (АТАТ1), которая, как было показано, играет важную роль во многих биологических и молекулярных функциях и, следовательно, также связана с ацетилированием. при многих заболеваниях человека, особенно неврологических заболеваниях .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Ганнинг П.В., Гошдастидер Ю., Уитакер С., Попп Д., Робинсон Р.К. (июнь 2015 г.). «Эволюция композиционно и функционально различных актиновых нитей» . Журнал клеточной науки . 128 (11): 2009–19. дои : 10.1242/jcs.165563 . ПМИД 25788699 .
- ^ Мори, Х. (16 марта 1968 г.). «Аминокислотный состав «Тубулина», входящего в состав микротрубочек жгутиков спермиев» . Природа . 217 (5133): 1053–1054. дои : 10.1038/2171053a0 . ISSN 0028-0836 . ПМИД 4296139 .
- ^ Финдейзен П., Мюльхаузен С., Демпевольф С., Герцог Дж., Зитлоу А., Карломаньо Т., Коллмар М. «Шесть подгрупп и обширные недавние дупликации характеризуют эволюцию семейства эукариотических белков тубулина» Genome Biol Evol (2014) 6 : 2274-2288.
- ^ Jump up to: а б Терк Э., Уиллс А.А., Квон Т., Седзински Дж., Уоллингфорд Дж.Б., Стернс Т. «Зета-тубулин является членом консервативного модуля тубулина и компонентом центриолярной базальной ножки в многоресничных клетках» Current Biology (2015) 25 :2177 -2183.
- ^ Jump up to: а б Ногалес Э., Даунинг К.Х., Амос Л.А., Лёве Дж. (июнь 1998 г.). «Тубулин и FtsZ образуют отдельное семейство GTPases». Структурная биология природы . 5 (6): 451–8. дои : 10.1038/nsb0698-451 . ПМИД 9628483 . S2CID 5945125 .
- ^ Jump up to: а б с Дженкинс С., Самудрала Р., Андерсон И., Хедлунд Б.П., Петрони Г., Михайлова Н. и др. (декабрь 2002 г.). «Гены цитоскелетного белка тубулина у бактерий рода Prosthecobacter» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (26): 17049–54. Бибкод : 2002PNAS...9917049J . дои : 10.1073/pnas.012516899 . ПМК 139267 . ПМИД 12486237 .
- ^ Ютин Н., Кунин Е.В. (март 2012 г.). «Археальное происхождение тубулина» . Биология Директ . 7:10 . дои : 10.1186/1745-6150-7-10 . ПМЦ 3349469 . ПМИД 22458654 .
- ^ Jump up to: а б с Ларсен Р.А., Кусумано С., Фудзиока А., Лим-Фонг Дж., Паттерсон П., Польяно Дж. (июнь 2007 г.). «Беговая дорожка прокариотического тубулиноподобного белка TubZ, необходимого для стабильности плазмиды в Bacillus thuringiensis» . Гены и развитие . 21 (11): 1340–52. дои : 10.1101/gad.1546107 . ПМЦ 1877747 . ПМИД 17510284 .
- ^ Ногалес Э., Вольф С.Г., Даунинг К.Х. (январь 1998 г.). «Структура димера альфа-бета-тубулина методом электронной кристаллографии». Природа . 391 (6663): 199–203. Бибкод : 1998Natur.391..199N . дои : 10.1038/34465 . ПМИД 9428769 . S2CID 4412367 .
- ^ Jump up to: а б с Даггин И.Г., Эйлетт Ч.С., Уолш Дж.К., Мичи К.А., Ван К., Тернбулл Л. и др. (март 2015 г.). «Тубулиноподобные белки CetZ контролируют форму клеток архей» . Природа . 519 (7543): 362–5. Бибкод : 2015Natur.519..362D . дои : 10.1038/nature13983 . ПМК 4369195 . ПМИД 25533961 .
- ^ Лёве Дж., Амос Л.А. (январь 1998 г.). «Кристаллическая структура белка деления бактериальных клеток FtsZ». Природа . 391 (6663): 203–6. Бибкод : 1998Natur.391..203L . дои : 10.1038/34472 . ПМИД 9428770 . S2CID 4330857 .
- ^ «Цифровые загрузки» . PurSolutions . Проверено 19 февраля 2020 г.
- ^ Jump up to: а б Пилхофер М., Ладинский М.С., Макдауэлл А.В., Петрони Дж., Дженсен Г.Дж. (декабрь 2011 г.). «Микротрубочки у бактерий: древние тубулины образуют пятипротофиламентный гомолог эукариотического цитоскелета» . ПЛОС Биология . 9 (12): e1001213. дои : 10.1371/journal.pbio.1001213 . ПМЦ 3232192 . ПМИД 22162949 .
- ^ Уильямс Р.К., Шах С., Сакетт Д. (ноябрь 1999 г.). «Разделение изоформ тубулина путем изоэлектрического фокусирования в иммобилизованных гелях с градиентом pH». Аналитическая биохимия . 275 (2): 265–7. дои : 10.1006/abio.1999.4326 . ПМИД 10552916 .
- ^ «Тубулин в белковых последовательностях» . ЭМБЛ-ЭБИ .
- ^ Хилд Р., Ногалес Э. (январь 2002 г.). «Динамика микротрубочек» . Журнал клеточной науки . 115 (Часть 1): 3–4. дои : 10.1242/jcs.115.1.3 . ПМИД 11801717 .
- ^ Ховард Дж., Хайман А.А. (апрель 2003 г.). «Динамика и механика плюс-конца микротрубочки». Природа . 422 (6933): 753–8. Бибкод : 2003Natur.422..753H . дои : 10.1038/nature01600 . ПМИД 12700769 . S2CID 4427406 .
- ^ Jump up to: а б Мартин-Галиано А.Дж., Олива М.А., Санс Л., Бхаттачарья А., Серна М., Йебенес Х. и др. (июнь 2011 г.). «Отличные последовательности петель бактериального тубулина и примитивные свойства сборки подтверждают его происхождение от эукариотического предка тубулина» . Журнал биологической химии . 286 (22): 19789–803. дои : 10.1074/jbc.M111.230094 . ПМК 3103357 . ПМИД 21467045 .
- ^ Jump up to: а б Шлипер Д., Олива М.А., Андреу Дж.М., Лёве Дж. (июнь 2005 г.). «Структура бактериального тубулина BtubA/B: доказательства горизонтального переноса генов» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (26): 9170–5. Бибкод : 2005PNAS..102.9170S . дои : 10.1073/pnas.0502859102 . ПМЦ 1166614 . ПМИД 15967998 .
- ^ Дэн Х, Финк Дж., Бхарат Т.А., Хе С., Курейсайте-Чизиене Д., Лёве Дж. (июль 2017 г.). «Простекобактерии BtubAB демонстрируют динамическую нестабильность» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (29): E5950–E5958. дои : 10.1073/pnas.1705062114 . ПМК 5530688 . ПМИД 28673988 .
- ^ Эйлетт Ч., Ван К., Мичи К.А., Амос Л.А., Лёве Дж. (ноябрь 2010 г.). «Филаментная структура гомолога бактериального тубулина TubZ» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (46): 19766–71. Бибкод : 2010PNAS..10719766A . дои : 10.1073/pnas.1010176107 . ПМЦ 2993389 . ПМИД 20974911 .
- ^ Бхарат Т.А., Муршудов Г.Н., Саксе С., Лёве Дж. (июль 2015 г.). «Структуры актиноподобных филаментов ParM демонстрируют архитектуру веретена разделения плазмид» . Природа . 523 (7558): 106–10. Бибкод : 2015Natur.523..106B . дои : 10.1038/nature14356 . ПМЦ 4493928 . ПМИД 25915019 .
- ^ NCBI CCD cd2186
- ^ Jump up to: а б Чжоу Дж., Яннакакоу П. (январь 2005 г.). «Нацеливание на микротрубочки для химиотерапии рака». Современная медицинская химия. Противораковые агенты . 5 (1): 65–71. дои : 10.2174/1568011053352569 . ПМИД 15720262 .
- ^ «Мебендазол» . Наркотики.com . Американское общество фармацевтов системы здравоохранения. Архивировано из оригинала 11 декабря 2019 года . Проверено 18 августа 2015 г.
- ^ «Альбендазол» . Наркотики.com . Американское общество фармацевтов системы здравоохранения. Архивировано из оригинала 23 сентября 2015 года . Проверено 18 августа 2015 г.
- ^ Сербус Л.Р., Ландманн Ф., Брей В.М., Уайт П.М., Руйбал Дж., Локи Р.С. и др. (сентябрь 2012 г.). «Клеточный скрининг показывает, что метаболит альбендазола, альбендазолсульфон, нацелен на Wolbachia» . ПЛОС Патогены . 8 (9): e1002922. дои : 10.1371/journal.ppat.1002922 . ПМЦ 3447747 . ПМИД 23028321 .
- ^ Карки Р., Мариани М., Андреоли М., Хе С., Скамбия Г., Шахаби С., Ферлини С. (апрель 2013 г.). «βIII-тубулин: биомаркер устойчивости к таксанам или мишень для лекарств?». Мнение экспертов о терапевтических целях . 17 (4): 461–72. дои : 10.1517/14728222.2013.766170 . ПМИД 23379899 . S2CID 26229777 .
- ^ Лудуэнья РФ (май 1993 г.). «Являются ли изотипы тубулина функционально значимыми» . Молекулярная биология клетки . 4 (5): 445–57. дои : 10.1091/mbc.4.5.445 . ПМК 300949 . ПМИД 8334301 .
- ^ «Тубулин TUBB1, бета 1 класс VI [Homo sapiens (человек)]» . Джин — NCBI .
- ^ Jump up to: а б Лецин П. и др. (август 2000 г.). «Гематопоэтически-специфичный бета-1-тубулин участвует в пути биогенеза тромбоцитов, зависящем от транскрипционного фактора NF-E2» . Кровь . 96 (4): 1366–73. дои : 10.1182/blood.V96.4.1366 . ПМИД 10942379 .
- ^ Ян Х., Гангули А., Инь С., Кабрал Ф. (март 2011 г.). «β-тубулин класса VI, специфичный для линии мегакариоцитов, подавляет динамику микротрубочек, фрагментирует микротрубочки и блокирует деление клеток» . Цитоскелет . 68 (3): 175–87. дои : 10.1002/см.20503 . ПМК 3082363 . ПМИД 21309084 .
- ^ МакНелли Ф.Дж., Вейл Р.Д. (ноябрь 1993 г.). «Идентификация катанин, АТФазы, которая разрезает и разбирает стабильные микротрубочки». Клетка . 75 (3): 419–29. дои : 10.1016/0092-8674(93)90377-3 . ПМИД 8221885 . S2CID 10264319 .
- ^ Чумова Ю, Трегелова Л, Коурова Х, Волц Дж, Сулименко В, Халада П, Кучера О, Бенада О, Кучаржова А, Клебанович А, Драбер П, Даниэль Г, Бинарова П (2018). «γ-Тубулин обладает консервативным внутренним свойством самополимеризации с образованием двухцепочечных нитей и фибриллярных сетей» . Biochimica et Biophysical Acta (BBA) - Исследования молекулярных клеток . 1865 (5): 734–748. дои : 10.1016/j.bbamcr.2018.02.009 . ПМИД 29499229 . S2CID 4053150 .
- ^ Зонтаг К.А., Сейдж Х., Эриксон Х.П. (сентябрь 2009 г.). «Гетеродимер BtubA-BtubB является важным промежуточным продуктом в сборке протофиламентов» . ПЛОС ОДИН . 4 (9): е7253. Бибкод : 2009PLoSO...4.7253S . дои : 10.1371/journal.pone.0007253 . ПМК 2746283 . ПМИД 19787042 .
- ^ Марголин В. (ноябрь 2005 г.). «ФцЗ и деление прокариотических клеток и органелл» . Обзоры природы. Молекулярно-клеточная биология . 6 (11): 862–71. дои : 10.1038/nrm1745 . ПМЦ 4757588 . ПМИД 16227976 .
- ^ Ни Л, Сюй В, Кумарасвами М, Шумахер М.А. (июнь 2010 г.). «Плазмидный белок TubR использует особый способ связывания HTH с ДНК и привлекает прокариотический гомолог тубулина TubZ для осуществления разделения ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (26): 11763–8. дои : 10.1073/pnas.1003817107 . ПМЦ 2900659 . ПМИД 20534443 .
- ^ Чайкеэратисак, В; Нгуен, К; Иган, Мэн; Эрб, МЛ; Вавилина А; Польяно, Дж. (15 августа 2017 г.). «Фаговое ядро и тубулиновое веретено консервативны среди крупных фагов Pseudomonas» . Отчеты по ячейкам . 20 (7): 1563–1571. дои : 10.1016/j.celrep.2017.07.064 . ПМК 6028189 . ПМИД 28813669 .
- ^ Мэлоун, Люсия М.; Уорринг, Сюзанна Л.; Джексон, Саймон А.; Варнеке, Кэролин; Гарднер, Пол П.; Гуми, Лаура Ф.; Финеран, Питер К. (9 декабря 2019 г.). «Большой фаг, образующий ядроподобную структуру, уклоняется от воздействия CRISPR-Cas на ДНК, но уязвим для иммунитета, основанного на РНК III типа». Природная микробиология . 5 (1): 48–55. bioRxiv 10.1101/782524 . дои : 10.1038/s41564-019-0612-5 . ПМИД 31819217 . S2CID 209164667 .
- ^ Акыл, Джанер; Али, Самсон; Тран, Линь Т.; Гайяр, Джереми; Ли, Вэньфэй; Хаясида, Кеничи; Хиросе, Мика; Като, Такаюки; Осима, Ацунори; Фудзисима, Косукэ; Бланшуан, Лоран; Нарита, Акихиро; Робинсон, Роберт С. (2021). «Структура и динамика тубулина Odinarchaeota и значение для эволюции эукариотических микротрубочек». bioRxiv 10.1101/2021.10.22.465531 . дои : 10.1101/2021.10.22.465531 . S2CID 239831170 .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ ван дер Хейден Р., Джейкобс Д.И., Снойер В., Халлард Д., Верпорте Р. (март 2004 г.). «Алкалоиды катаранта: фармакогнозия и биотехнология». Современная медицинская химия . 11 (5): 607–28. дои : 10.2174/0929867043455846 . ПМИД 15032608 .
- ^ Равинья, Энрике (2011). «Алкалоиды барвинка» . Эволюция открытия лекарств: от традиционных лекарств к современным лекарствам . Джон Уайли и сыновья . стр. 157–159. ISBN 9783527326693 .
- ^ Купер, Раймонд; Дикин, Джеффри Джон (2016). «Подарок Африки миру» . Ботанические чудеса: химия растений, изменивших мир . ЦРК Пресс . стр. 46–51. ISBN 9781498704304 .
- ^ Кеглевич П., Хазай Л., Калаус Г., Сантай С. (май 2012 г.). «Модификации основных скелетов винбластина и винкристина» . Молекулы . 17 (5): 5893–914. дои : 10.3390/molecules17055893 . ПМК 6268133 . ПМИД 22609781 .
- ^ Нго QA, Русси Ф, Кормье А, Торет С, Кноссов М, Генар Д, Герит Ф (январь 2009 г.). «Синтез и биологическая оценка алкалоидов барвинка и гибридов фомопсина». Журнал медицинской химии . 52 (1): 134–42. дои : 10.1021/jm801064y . ПМИД 19072542 .
- ^ Альтманн, Карл-Хайнц (2009). «Доклиническая фармакология и исследование структуры-активности эпотилонов» . В Мульцере, Иоганн Х. (ред.). Эпотилоны: выдающееся семейство противоопухолевых средств: от почвы к клинике . Springer Science & Business Media . стр. 157–220. ISBN 9783211782071 .
- ^ Самсон А.Л., Кнаупп А.С., Сашиндранат М., Борг Р.Дж., Ау А.Е., Копс Э.Дж. и др. (октябрь 2012 г.). «Нуклеоцитоплазматическая коагуляция: событие агрегации, вызванное повреждением, которое дисульфид сшивает белки и облегчает их удаление плазмином» . Отчеты по ячейкам . 2 (4): 889–901. дои : 10.1016/j.celrep.2012.08.026 . ПМИД 23041318 .
Внешние ссылки
[ редактировать ]- Тубулин в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- ЭК 3.6.5.6
- Протоколы экспериментов с тубулином
- Инфографика тубулина высокого разрешения
| Базы данных органов управления : Национальные |
|---|