Неурула

Нейрула происходит — позвоночных эмбрион на ранней стадии развития, в котором нейруляция . Стадии нейрулы предшествует стадия гаструлы ; следовательно, нейруляции предшествует гаструляция . [1] Нейруляция знаменует начало процесса органогенеза . [2]
Мыши, цыплята и лягушки являются распространенными экспериментальными моделями для изучения нейрулы. В зависимости от вида эмбрионы достигают стадии нейрулы в разные моменты времени и проводят на этой стадии разное количество времени. [3] [4] Для яйцекладущих организмов температура инкубации также влияет на продолжительность нейруляции. [2] Помимо развития нервной трубки , в эмбрионе на стадии нейрулы в зависимости от вида происходят и другие процессы. Например, у рептилий внеэмбриональные мембранные ткани становятся отличными от эмбриона. [2]
Зародыш нейрулы имеет пять участков мезодермы , окружающих нервную трубку. [5] Передняя мезодерма развивается в область головы, а задняя мезодерма развивается в туловище. [1] Различные молекулы, включая протеогликаны во внеклеточном матриксе и гены, включая факторы транскрипции Pax , необходимы для развития и закрытия нервной трубки у эмбриона на стадии нейрулы. [6] [7]
нейруляция
[ редактировать ]Нейруляция — это процесс у эмбрионов позвоночных на стадии нейрулы, на которой нервная трубка . формируется [6] [8] Различают два типа нейруляции: первичную и вторичную нейруляцию . Первичная нейруляция относится к образованию и складыванию нервной пластинки внутрь с образованием нервной трубки. [6] [8] При вторичной нейруляции нервная трубка образуется в результате слияния полостей мозгового канатика. [6] [8] [9] У амфибий и рептилий первичная нейруляция образует всю нервную трубку, причем нервная трубка закрывается одновременно по всей своей длине. [8] Напротив, у рыб вторичная нейруляция образует нервную трубку. [10] У птиц и млекопитающих встречается как первичная, так и вторичная нейруляция, хотя и с небольшими различиями. Первичная нейруляция происходит в краниальном и верхнем отделах спинного мозга, что дает начало головному и верхним отделам спинного мозга. Вторичная нейруляция происходит в нижних крестцовых и каудальных отделах, в результате чего формируются нижние отделы спинного мозга. [6] [10] У птиц нервная трубка закрывается спереди назад, а у млекопитающих сначала закрывается середина, а затем закрываются оба конца. [8]
Сроки развития
[ редактировать ]Точка, в которой эмбрион достигает стадии нейрулы, различается у разных видов, тогда как для яйцекладущих организмов на продолжительность нейруляции дополнительно влияет температура инкубации. В целом, чем ниже температура, тем больше продолжительность нейруляции. Куриные эмбрионы достигают стадии нейрулы на 2-й день после оплодотворения и подвергаются нейруляции до 5-го дня. Рептилии, включая крокодилов, ящериц и черепах, как правило, проводят более длительное время на стадии нейрулы. [2] Типичный эмбрион лягушки, инкубированный при 18 ° C, имеет нейрулу ранней стадии через 50 часов после оплодотворения и нейрулу поздней стадии через 67 часов. [3] У эмбриона мыши нейруляция начинается на 7,5 день беременности и остается на стадии нейрулы до 9 дня. [4]
Морфология
[ редактировать ]Мезодерму . эмбриона позвоночного на стадии нейрулы можно разделить на пять областей Вентральнее нервной трубки находится хордамезодерма . Латеральнее каждой стороны нервной трубки находится параксиальная мезодерма , а промежуточно-латеральная область нервной трубки — промежуточная мезодерма . Четвертая область — мезодерма латеральной пластинки , а последняя — головная мезенхима. [5] Передние части мезодермы развиваются в ростральные области организма, такие как голова, а задняя мезодерма развивается в каудальные области, такие как туловище или хвост. [1] Параксиальная мезодерма, также называемая сомитной мезодермой, развивается в сомиты , блоки ткани, которые имеют сегментарную структуру. Сомиты, в свою очередь, дают начало позвонкам, ребрам, скелетным мышцам, хрящам, сухожилиям и коже. [8] [11]
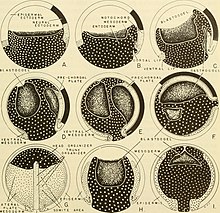
У Xenopus laevis переход от гаструлы к нейруле включает морфологические изменения в двух областях, окружающих бластопор: дорсальной инволюционной маргинальной зоне (IMZ) и вышележащей неинволюционирующей маргинальной зоне (NIMZ) гаструлы. После инволюции на стадии средней гаструлы IMZ подвергается конвергентному расширению , при котором латеральные области сужаются и перемещаются к средней линии, а передний конец удлиняется. Это приводит к сужению бластопора . НИМЗ, который не является эвольвентным, одновременно распространяется в противоположном направлении и с большей скоростью, чтобы охватить области, больше не занимаемые ИМЗ. Конвергентное расширение ИМЗ и НИМЗ начинается во второй половине гаструляции и продолжается до поздней стадии нейрулы. В конце концов, глубокая ткань ИМЗ образует центральную хорду и окружающую параксиальную мезодерму . На ранней стадии нейрулы хорда четко выражена. Нотохордальные клетки образуют образование, представляющее собой стопку монет, в процессе, называемом периферической интеркаляцией. Поверхностный слой ИМЗ перерастает в кровлю. архентерон , или примитивная кишка, в то время как подлежащая энтодерма образует дно архентерона. НИМЗ развивается в структуру, напоминающую раннюю нервную трубку . Внешний эктодермальный слой нейрулы формируется путем равномерного расширения клеток анимального полюса , известного как анимальная шапочка. Затем эктодерма ткань дифференцируется на нервную и эпидермальную . [12]
У эмбрионов рептилий, начиная с поздней стадии нейрулы и переходя на ранние стадии органогенеза, внеэмбриональные мембранные ткани, включающие желточный мешок , хорион и амнион, становятся отличными от тканей эмбриона. Мезодерма разделяется , образуя внеэмбриональный целом, состоящий из двух слоев. Васкуляризированный внутренний слой мезодермы-энтодермы, называемый спланхноплеврой, развивается в желточный мешок, а неваскуляризированный наружный слой эктодермы-мезодермы, называемый соматоплеврой, становится амнионом и хорионом. Во время органогенеза эти три внеэмбриональные ткани полностью развиваются. Кроме того, внутри рептильной нейрулы начинают дифференцироваться ткани головного мозга и начинают формироваться сердце и кровеносные сосуды. [2]
Химический состав
[ редактировать ]Ткани нейрулы мыши делятся быстро, средний клеточный цикл длится 8–10 часов. Протеогликаны во внеклеточном матриксе (ECM) клеток на стадии нейрулы играют важную роль в обеспечении функциональной краниальной нейруляции и поднятии нервных складок; гиалуроновая кислота (ГК) синтезируется и накапливается, при этом в клетке сохраняется низкий уровень сульфатированных гликозаминогликанов (ГАГ). ГК участвует в создании двояковыпуклых нервных складок, тогда как сульфатированные ГАГ играют решающую роль в придании нервной борозде V-образной формы, а также в закрытии нервной трубки. ЕСМ не играет основной роли в нейруляции спинного мозга из-за плотной упаковки мезодермальных клеток в спинальной области, что обеспечивает мало межклеточного пространства. Кроме того, полагают, что актинсодержащие микрофиламенты необходимы для краниальной нейруляции. Они могут действовать как механизм складывания нервных клеток или стабилизировать уже сформировавшиеся нервные складки; однако их точная роль не определена. Есть некоторые свидетельства того, что Факторы роста , такие как инсулин или трансферрин , также играют роль в нейруляции, но эта связь недостаточно изучена. [6]
Активация гена
[ редактировать ]Было обнаружено, что в эмбрионе на стадии нейрулы экспрессируются различные гены. Различные гены активируются для разных событий нейруляции, например, происходящих в отдельных областях развивающейся нервной трубки. [6] Эти гены необходимы для правильной нейруляции и закрытия нервной трубки. такие как Wnts , FGFs и BMFs, наряду с факторами транскрипции, включающими гены Msx , Snails Сигнальные молекулы , , Sox8/9/10 и Pax3 /7 , играют ключевую роль в формировании нервного гребня. [6]
Транскрипционные факторы Pax играют важную роль в раннем развитии, особенно в отношении ЦНС и нервного гребня. Pax3 и Pax7 способствуют как выживанию клеток нервного гребня, так и повышению устойчивости к стрессу окружающей среды. [7] У эмбрионов мышей Pax3 блокирует ген-супрессор опухоли p53 , который необходим для контролируемой пролиферации и стабильности генома, экспрессируется во всех клетках нейрулы. [6] [7] Во время раннего развития Pax3 экспрессируется в задней и латеральной области нервной пластинки, той же области, из которой возникает нервный гребень. [7] Было обнаружено, что дефекты нервного гребня возникают у мутантов Pax3 мыши и человека, что указывает на важность функциональности. [7] [13] У кур, лягушек и рыб Pax3/Pax7 активируются посредством передачи сигналов Wnt и FGF. [7] Pax3 и Pax7 также необходимы для индукции нервного гребня после того, как истощение этих двух генов привело к отсутствию активации специфических генов нервного гребня Snail2 и Foxd3 , что не позволило дальнейшему развитию или эмиграции нервного гребня. [7] Использование нокаутов помогло понять роль и функции нескольких генов, обнаруженных в нейруле. Например, было обнаружено, что Wnt-1 не играет никакой роли в закрытии нервной пластинки, несмотря на то, что он присутствует на кончике нервных складок во время ее закрытия. Хотя мутанты Wnt-1 действительно приводят к дефектам структуры мозга. Notch1 участвует в формировании сомитов. HNF-3 необходим для развития хорды и узла. [14] Ген аполипопротеина В , который участвует в транспортировке и метаболизме жирорастворимых молекул в крови, экспрессируется в желточном мешке и печени плода. [6] В нейруле Xenopus laevis присутствуют гены развития Xwnt-3 и Xwnt-4. [15]
Ссылки
[ редактировать ]- ^ Jump up to: а б с Стерн, Клаудио Д. (февраль 2001 г.). «Первоначальное формирование структуры центральной нервной системы: сколько организаторов?» (PDF) . Обзоры природы Неврология . 2 (2): 92–98. дои : 10.1038/35053563 . ПМИД 11252999 . S2CID 13652379 . [ постоянная мертвая ссылка ]
- ^ Jump up to: а б с д и Эндрюс, РМ (2004). «Закономерности эмбрионального развития» (PDF) . Инкубация рептилий: окружающая среда, эволюция и поведение : 75–102. S2CID 42246488 . Архивировано из оригинала (PDF) 30 ноября 2018 г.
- ^ Jump up to: а б Хилл, Массачусетс (30 ноября 2018 г.) Эмбриологическое развитие лягушек . Получено с https://embryology.med.unsw.edu.au/embryology/index.php/Frog_Development .
- ^ Jump up to: а б Хилл, Массачусетс (30 ноября 2018 г.) эмбриологии Подробная хронология мышей . Получено с https://embryology.med.unsw.edu.au/embryology/index.php/Mouse_Timeline_Detailed .
- ^ Jump up to: а б Гилберт, Скотт Ф. (2000). «Параксиальная и промежуточная мезодерма» . Биология развития. 6-е издание .
- ^ Jump up to: а б с д и ж г час я дж Флеминг, А; Джеррелли, Д; Грин, Северная Дакота; Копп, Эй Джей (1 марта 2002 г.). «Механизмы нормальной и аномальной нейруляции: данные исследований культуры эмбрионов» . Международный журнал биологии развития . 41 (2). ISSN 0214-6282 .
- ^ Jump up to: а б с д и ж г Монсоро-Бурк, Энн Х. (1 августа 2015 г.). «Факторы транскрипции PAX в развитии нервного гребня». Семинары по клеточной биологии и биологии развития . 44 : 87–96. дои : 10.1016/j.semcdb.2015.09.015 . ISSN 1084-9521 . ПМИД 26410165 .
- ^ Jump up to: а б с д и ж Слончевски, Джоан. «Глава 14. Гаструляция и нейруляция» . biology.kenyon.edu . Проверено 16 ноября 2018 г.
- ^ Симокита, Эйсуке; Такахаси, Ёсико (апрель 2011 г.). «Вторичная нейруляция: картирование судьбы и манипуляции с генами нервной трубки хвостового зачатка». Развитие, рост и дифференциация . 53 (3): 401–410. дои : 10.1111/j.1440-169x.2011.01260.x . ISSN 0012-1592 . ПМИД 21492152 . S2CID 31342085 .
- ^ Jump up to: а б Шенвульф, Гэри К.; Смит, Джоди Л. (2000), «Механизмы нейруляции» , Протоколы биологии развития , том. 136, Humana Press, стр. 125–134 , номер doi : 10.1385/1-59259-065-9:125 , ISBN. 978-1592590650 , PMID 10840705
- ^ Ларсен, Уильям Дж. (2001). Эмбриология человека (3-е изд.). Филадельфия, Пенсильвания: Черчилль Ливингстон. стр. 53–86. ISBN 0-443-06583-7 .
- ^ Келлер Р.Э., Данилчик М., Гимлих Р. и Ши Дж. (1985). «Функция и механизм конвергентного расширения во время гаструляции Xenopus laevis » (PDF) . Развитие , 89 (Приложение), 185-209.
- ^ Тассабехджи, Маяда; Прочтите, Эндрю П.; Ньютон, Валери Э.; Харрис, Родни; Баллинг, Руди; Грусс, Питер; Страчан, Том (13 февраля 1992 г.). «Пациенты с синдромом Ваарденбурга имеют мутации в человеческом гомологе парного гена Pax-3». Природа . 355 (6361): 635–636. Бибкод : 1992Natur.355..635T . дои : 10.1038/355635a0 . ISSN 0028-0836 . ПМИД 1347148 . S2CID 4330804 .
- ^ Ленкевич, Э.; Ференцова, А.; Шевчикова, Э. (17 марта 2012 г.). «Субконъюнктивальная аутогемотерапия ожогов глаз в наших случаях». Глазная клиника . 94 (4): 113–4. дои : 10.4016/46352.01 . ПМИД 1405409 .
- ^ Кристиан, Ян Л.; Гэвин, Брайан Дж.; МакМахон, Эндрю П.; Мун, Рэндалл Т. (1 февраля 1991 г.). «Выделение кДНК, частично кодирующих четыре белка, родственного XenopusWnt-1int-1, и характеристика их временной экспрессии во время эмбрионального развития». Биология развития . 143 (2): 230–234. дои : 10.1016/0012-1606(91)90073-C . ISSN 0012-1606 . ПМИД 1991549 .