ТААР1
| ТААР1 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | TAAR1 , TA1, TAR1, TRAR1, рецептор 1, связанный с следами аминов, рецептор следовых аминов | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | ОМИМ : 609333 ; МГИ : 2148258 ; Гомологен : 24938 ; Генные карты : TAAR1 ; СОБСТВЕННЫЙ : TAAR1 - ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




Рецептор 1, связанный с амином ( TAAR1 ), представляет собой рецептора, связанного с амином (TAAR) белок , который у людей кодируется TAAR1 геном . [5] TAAR1 представляет собой внутриклеточный аминоактивируемый желудке G s -связанный и G q -связанный с G-белком рецептор (GPCR), который в первую очередь экспрессируется в нескольких периферических органах и клетках (например, , тонком кишечнике , двенадцатиперстной кишке и лейкоцитах ). , астроцитах и во внутриклеточной среде внутри пресинаптической плазматической мембраны (т.е. окончаний аксонов ) моноаминных нейронов в центральной нервной системе (ЦНС). [6] [7] [8] [9] TAAR1 был открыт в 2001 году двумя независимыми группами исследователей, Borowski et al. и Бунцов и др. [10] [11] TAAR1 является одним из шести функциональных человеческих рецепторов, связанных с следовыми аминами , которые названы так из-за их способности связывать эндогенные амины, которые встречаются в тканях в следовых концентрациях. [12] [13] TAAR1 играет значительную роль в регуляции нейротрансмиссии в дофаминовых , норадреналиновых и серотониновых нейронах ЦНС; [7] [12] он также влияет на иммунную систему и функцию нейроиммунной системы посредством различных механизмов. [14] [15] [16] [17]
TAAR1 представляет собой рецептор с высоким сродством к амфетамину , метамфетамину , дофамину и следовым аминам , который опосредует некоторые из их клеточных эффектов в моноаминовых нейронах центральной нервной системы . [7] [12]
Основными эндогенными лигандами человеческого рецептора TAAR1 (hTAAR1) в порядке эффективности являются:
тирамин > β-фенэтиламин > дофамин = октопамин . [6]
Открытие
[ редактировать ]TAAR1 был открыт независимо Боровски и др. и Бунцов и др. в 2001 году. Чтобы найти генетические варианты, ответственные за синтез TAAR1, они использовали смеси олигонуклеотидов с последовательностями, связанными с рецепторами, связанными с G-белком (GPCR) серотонина и дофамина , чтобы обнаружить новые последовательности ДНК в крысы геномной ДНК и кДНК , которые затем амплифицировали. и клонирован. Полученная последовательность не была найдена ни в одной базе данных и закодирована для TAAR1. [10] [11] Дальнейшую характеристику функциональной роли TAAR1 и других рецепторов этого семейства провели другие исследователи, в том числе Рауль Гайнетдинов и его коллеги. [18]
Структура
[ редактировать ]TAAR1 имеет структурное сходство с родопсина класса GPCR подсемейством А. [11] Он имеет 7 трансмембранных доменов с короткими N- и C-концевыми отростками. [19] TAAR1 на 62–96% идентичен TAARs2-15, что позволяет предположить, что подсемейство недавно TAAR развилось ; в то же время низкая степень сходства между ортологами TAAR1 позволяет предположить, что они быстро эволюционируют. [10] TAAR1 разделяет предсказательный пептидный мотив со всеми другими TAAR. Этот мотив перекрывается с трансмембранным доменом VII, и его идентичность — NSXXNPXX[Y,H]XXX[Y,F]XWF. TAAR1 и его гомологи имеют векторы лигандного кармана, которые используют наборы из 35 аминокислот, которые, как известно, непосредственно участвуют во взаимодействии рецептор-лиганд. [13]
Ген
[ редактировать ]Все гены TAAR человека расположены на одной хромосоме, охватывающей 109 т.п.н. хромосомы человека 6q23.1, 192 т.п.н. хромосомы мыши 10A4 и 216 т.п.н. хромосомы 1p12 крысы. Каждый TAAR происходит из одного экзона , за исключением TAAR2 , который кодируется двумя экзонами. [13] человека Считается, что ген TAAR1 является безинтронным геном. [20]
Распределение тканей
[ редактировать ]
На сегодняшний день TAAR1 был идентифицирован и клонирован в пяти различных млекопитающих геномах : человека, мыши, крысы, обезьяны и шимпанзе . У крыс мРНК TAAR1 обнаруживается на низких и умеренных уровнях в периферических тканях, таких как желудок , почки , кишечник. [21] и легких , а также на низких уровнях в мозге . [10] Taar1 макаки-резуса и TAAR1 человека имеют высокое сходство последовательностей, а мРНК TAAR1 высоко экспрессируется в одних и тех же важных моноаминергических областях обоих видов . Эти области включают дорсальное и вентральное хвостатое ядро , скорлупу , черную субстанцию , прилежащее ядро , вентральную область покрышки , голубое пятно , миндалевидное тело и ядро шва . [6] [22] hTAAR1 также был идентифицирован в астроцитах человека. [6] [14]
За пределами центральной нервной системы человека hTAAR1 также встречается в качестве внутриклеточного рецептора и преимущественно экспрессируется в желудке , кишечнике , [21] двенадцатиперстная кишка , [21] поджелудочной железы β-клетки и лейкоциты . [9] [21] В двенадцатиперстной кишке активация TAAR1 увеличивает глюкагоноподобного пептида-1 (GLP-1) и пептида YY (PYY); высвобождение [9] В желудке было обнаружено, что активация hTAAR1 увеличивает секрецию соматостатина ( гормона, ингибирующего гормон роста) дельта-клетками . [9]
hTAAR1 — единственный подтип человеческих рецепторов, ассоциированных с следами аминов , который не экспрессируется в обонятельном эпителии человека . [23]
Расположение внутри нейронов
[ редактировать ]TAAR1 представляет собой внутриклеточный рецептор, экспрессируемый в пресинаптическом окончании моноаминных нейронов у людей и других животных. [7] [12] [24] В модельных клеточных системах hTAAR1 имеет крайне низкую мембранную экспрессию. [24] Метод индукции мембранной экспрессии hTAAR1 был использован для изучения его фармакологии с помощью анализа биолюминесцентного резонансного переноса энергии цАМФ. [24]
Поскольку TAAR1 является внутриклеточным рецептором в моноаминовых нейронах, экзогенные лиганды TAAR1 должны проникать в пресинаптический нейрон через мембранный транспортный белок. [примечание 1] или быть способными диффундировать через пресинаптическую мембрану, чтобы достичь рецептора и вызвать ингибирование обратного захвата и отток нейромедиатора . [12] Следовательно, эффективность конкретного лиганда TAAR1 в обеспечении этих эффектов в различных моноаминовых нейронах является функцией как его сродства к связыванию с TAAR1, так и его способности перемещаться через пресинаптическую мембрану в каждом типе нейронов. [12] Вариабельность между сродством к субстрату лиганда TAAR1 к различным переносчикам моноаминов объясняет большую часть различий в его способности вызывать высвобождение нейромедиатора и ингибировать обратный захват в различных типах моноаминовых нейронов. [12] Например, лиганд TAAR1, который может легко проходить через переносчик норадреналина, но не через переносчик серотонина, будет вызывать – при прочих равных условиях – заметно более сильные TAAR1-индуцированные эффекты в норадреналиновых нейронах по сравнению с серотониновыми нейронами.
Рецепторные олигомеры
[ редактировать ]TAAR1 образует олигомеры GPCR с моноаминовыми ауторецепторами в нейронах in vivo . [25] [26] Эти и другие зарегистрированные гетероолигомеры TAAR1 включают:
Лиганды
[ редактировать ]| Следовой аминоассоциированный рецептор 1 | |
|---|---|
| Механизмы трансдукции | G s , G q , GIRKs , β-аррестин 2 |
| Первичные эндогенные агонисты | тирамин , β-фенилэтиламин , октопамин , дофамин |
| Агонисты | Эндогенные: следовые амины Экзогенные: RO5166017 , амфетамин , метамфетамин и другие. |
| Нейтральные антагонисты | Ни один не охарактеризован |
| Обратные агонисты | ЭППТБ |
| Положительные аллостерические модуляторы | Н/Д |
| Отрицательные аллостерические модуляторы | Н/Д |
| Внешние ресурсы | |
| ИЮФАР/БПС | 364 |
| Лекарственный Банк | Q96RJ0 |
| ХМДБ | ХМДБП10805 |
Агонисты
[ редактировать ]Следовые амины
[ редактировать ]Следовые амины представляют собой эндогенные амины, которые действуют как агонисты TAAR1 и присутствуют во внеклеточных концентрациях 0,1–10 нМ мозге, составляя менее 1% от общего количества биогенных аминов в нервной системе млекопитающих . [28] Некоторые из следовых аминов человека включают триптамин , фенэтиламин (ПЭА), N -метилфенэтиламин , п -тирамин , м -тирамин , N -метилтирамин , п -октопамин , м -октопамин и синефрин . Они имеют структурное сходство с тремя распространенными моноаминами: серотонином , дофамином и норадреналином . Каждый лиганд обладает разной эффективностью, измеряемой как повышенная концентрация циклического АМФ (цАМФ) после события связывания.
Порядок активности первичных эндогенных лигандов hTAAR1 следующий:
тирамин > β-фенэтиламин > дофамин = октопамин . [6]
Тиронамины
[ редактировать ]Тиронамины являются молекулярными производными гормона железы щитовидной и очень важны для эндокринной системы функции . 3-Йодтиронамин (T 1 AM) является наиболее мощным из когда-либо обнаруженных агонистов TAAR1, хотя ему не хватает сродства к переносчику моноаминов и, следовательно, он мало влияет на моноаминовые нейроны центральной нервной системы . Активация TAAR1 Т 1 AM приводит к продукции больших количеств цАМФ. Этот эффект сочетается со снижением температуры тела и сердечного выброса .
Синтетический
[ редактировать ]- Амфетамин и его замещенные производные метамфетамин и МДМА являются мощными агонистами hTAAR1 . [7] [9] При ассоциации с TAAR1 они вызывают увеличение продукции цАМФ, аналогичное таковому у ПЭА и п-тирамина. [9] Эти соединения структурно подобны ПЭА и п-тирамину. [11] [29]
- Бензофураны : 5-APB , 5-APDB , 6-APB , 6-APDB , 4-APB , 7-APB , 5-EAPB и 5-MAPDB , а также бензодифуран 2C-B-FLY являются hTAAR1 агонистами , которые имеют фармакодинамический профиль, подобный МДМА . [30]
- Метилфенэтиламины hTAAR1 агонистами ; являются к ним относятся α-метилфенэтиламин (амфетамин), β-метилфенэтиламин , N- метилфенэтиламин (следовые количества амина), 2-метилфенэтиламин , 3-метилфенэтиламин и 4-метилфенэтиламин . [31]
- У крыс диэтиламид лизергиновой кислоты (ЛСД) является агонистом rTAAR1 . [11] но у людей он не имеет сродства к hTAAR1 . [31]
- Определенные соединения 2-аминооксазолина ( RO5166017 , RO5256390 , RO5203648 и RO5263397 ) являются биодоступными при пероральном приеме , высокоэффективными и селективными агонистами TAAR1 у лабораторных животных. [32]
- RO5166017 или (S)-4-[(этилфениламино)метил]-4,5-дигидрооксазол-2-иламин представляет собой селективный агонист TAAR1 без значительной активности в отношении других мишеней. [33]
- RO5203648 и RO5263397 являются высокоселективными частичными агонистами TAAR1. [25] RO5203648 продемонстрировал явную антидепрессивную и антипсихотическую активность, кроме того, он ослаблял самостоятельное введение препарата и проявлял свойства, способствующие бодрствованию и улучшению когнитивных функций на моделях мышей и обезьян. [34]
- Улотаронт , исследуемый антипсихотик.
- Гуанфацин , лекарство от СДВГ. [35]
- Солриамфетол , средство, способствующее бодрствованию. [36]
Частичные агонисты
[ редактировать ]- Ралмитаронт , исследуемый антипсихотик.
Обратные агонисты
[ редактировать ]- ЭППТБ или N-(3-этоксифенил)-4-(пирролидин-1-ил)-3-трифторметилбензамид является селективным обратным агонистом hTAAR1 . [6] [37]
Нейтральные антагонисты
[ редактировать ]По состоянию на начало 2018 года [update] нейтральные антагонисты hTAAR1 не охарактеризованы. [6]
Функция
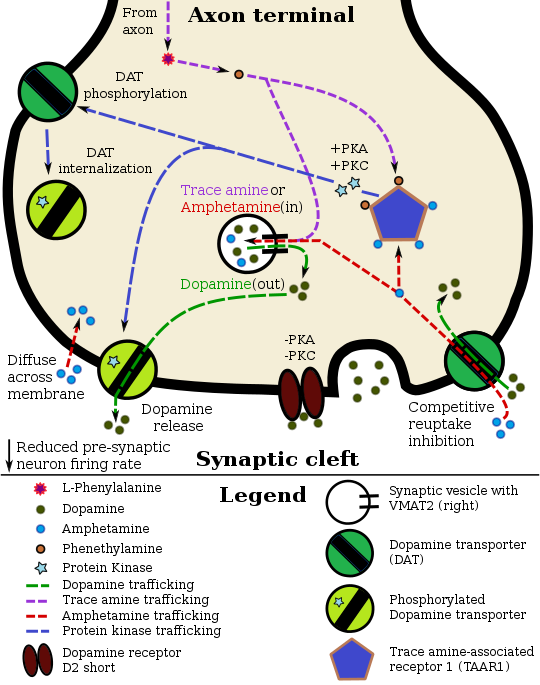
[ редактировать ]Фенэтиламин и амфетамин в дофаминовом нейроне, локализованном в TAAR1 |
Моноаминергические системы
[ редактировать ]До открытия TAAR1 считалось, что следовые амины выполняют очень ограниченные функции. Считалось, что они вызывают высвобождение норадреналина из симпатических нервных окончаний и конкурируют за места связывания катехоламинов или серотонина на родственных рецепторах, транспортерах и местах хранения. [28] Сегодня считается, что они играют гораздо более динамичную роль, регулируя моноаминергические системы мозга.
Одним из последующих эффектов активного TAAR1 является увеличение цАМФ в пресинаптических клетках посредством активации GαsG - белком аденилатциклазы . [10] [11] [13] Одно это может иметь множество клеточных последствий. Основная функция цАМФ может заключаться в повышении экспрессии следовых аминов в цитоплазме клеток . [29] Эти амины затем активируют внутриклеточный TAAR1. Моноаминовые ауторецепторы (например, D 2 короткие , пресинаптические α 2 и пресинаптические 5-HT 1A ) оказывают эффект, противоположный TAAR1, и вместе эти рецепторы обеспечивают систему регуляции моноаминов. [12] Примечательно, что амфетамин и следовые амины обладают высоким сродством связывания с TAAR1, но не с моноаминовыми ауторецепторами. [12] [7] Влияние агонистов TAAR1 на переносчики моноаминов в головном мозге, по-видимому, является сайт-специфичным. [12] Визуализирующие исследования показывают, что ингибирование обратного захвата моноаминов амфетамином и следовыми аминами зависит от присутствия совместной локализации TAAR1 в связанных моноаминовых нейронах. [12] По состоянию на 2010 год совместная локализация TAAR1 и переносчика дофамина (DAT) была визуализирована у макак-резус, но совместная локализация TAAR1 с переносчиком норадреналина (NET) и переносчиком серотонина (SERT) была подтверждена только с помощью информационной РНК. (мРНК) экспрессия. [12]
В нейронах с совместно локализованным TAAR1 агонисты TAAR1 увеличивают концентрации связанных моноаминов в синаптической щели , тем самым увеличивая связывание постсинаптических рецепторов. [12] Посредством прямой активации связанных с G-белком внутренних выпрямляющих калиевых каналов (GIRK) TAAR1 может снизить частоту возбуждения дофаминовых нейронов, в свою очередь предотвращая гипердофаминергическое состояние. [33] [40] [41] Амфетамин и следовые амины могут проникать в пресинаптический нейрон либо через ДАТ , либо путем прямой диффузии через мембрану нейрона. [12] В результате поглощения DAT амфетамин и следовые амины вызывают конкурентное ингибирование обратного захвата транспортера. [12] Попадая в пресинаптический нейрон, эти соединения активируют TAAR1, который посредством передачи сигналов протеинкиназы A (PKA) и протеинкиназы C (PKC) вызывает фосфорилирование DAT . Фосфорилирование любой протеинкиназой может привести к интернализации DAT ( неконкурентное ингибирование обратного захвата), но само фосфорилирование, опосредованное PKC, индуцирует функцию обратного транспортера ( отток дофамина ). [12] [44]
Иммунная система
[ редактировать ]Экспрессия TAAR1 на лимфоцитах связана с активацией иммунохарактеристик лимфоцитов. [16] В иммунной системе TAAR1 передает сигналы через активные фосфорилирования PKA и PKC. каскады [16] В исследовании 2012 года Panas et al. наблюдали, что метамфетамин оказывает такое воздействие, предполагая, что, помимо регуляции моноаминов в мозге, соединения, родственные амфетамину, могут оказывать влияние на иммунную систему. [16] Недавняя статья показала, что, наряду с TAAR1, TAAR2 необходим для полной активности следовых аминов в клетках PMN . [17]
Фитогемагглютинин усиливает hTAAR1 мРНК в циркулирующих лейкоцитах ; [6] в этих клетках активация TAAR1 опосредует хемотаксис лейкоцитов по отношению к агонистам TAAR1. [6] Также было показано, что агонисты TAAR1 (в частности, следовые амины) индуцируют секрецию интерлейкина 4 в Т-клетках и секрецию иммуноглобулина Е (IgE) в В-клетках . [6]
Локализованный в астроцитах TAAR1 регулирует уровни и функции EAAT2 в этих клетках; [14] это связано с патологиями нейроиммунной системы, вызванными метамфетамином . [14]
Клиническое значение
[ редактировать ]Низкая концентрация фенэтиламина (ФЭА) в мозге связана с большим депрессивным расстройством . [10] [28] [45] а высокие концентрации связаны с шизофренией . [45] [46] Низкие уровни PEA и недостаточная активация TAAR1 также, по-видимому, связаны с СДВГ . [45] [46] [47] Предполагается, что недостаточный уровень ПЭА приводит к инактивации TAAR1 и чрезмерному поглощению моноаминов транспортерами, что может привести к депрессии. [10] [28] Некоторые антидепрессанты действуют путем ингибирования моноаминоксидазы (МАО), которая увеличивает концентрацию следовых аминов, что, как предполагается, увеличивает активацию TAAR1 в пресинаптических клетках. [10] [13] ПЭА Снижение метаболизма связано с шизофренией, и это логичный вывод, учитывая, что избыток ПЭА может привести к чрезмерной активации TAAR1 и предотвращению функции переносчика моноаминов. Мутации в регионе q23.1 хромосомы 6 человека – той же хромосомы, которая кодирует TAAR1 – связаны с шизофренией. [13]
Медицинские обзоры за февраль 2015 и 2016 годов отметили, что TAAR1-селективные лиганды обладают значительным терапевтическим потенциалом для лечения психостимулирующих зависимостей (например, кокаина, амфетамина, метамфетамина и т. д.). [7] [8] Несмотря на широкое распространение за пределами ЦНС и ПНС, TAAR1 не влияет на гематологические функции и регуляцию гормонов щитовидной железы на разных стадиях старения. Такие данные показывают, что будущие методы лечения на основе TAAR1 должны оказывать небольшой гематологический эффект и, следовательно, вероятно, будут иметь хороший профиль безопасности. [48]
Исследовать
[ редактировать ]Крупное исследование ассоциаций генов-кандидатов, опубликованное в сентябре 2011 года, выявило значительные различия в частотах аллелей TAAR1 между группой пациентов с фибромиалгией и контрольной группой с хронической болью, что позволяет предположить, что этот ген может играть важную роль в патофизиологии заболевания; возможно, это представляет собой цель терапевтического вмешательства. [49]
В доклинических исследованиях на крысах активация TAAR1 в клетках поджелудочной железы способствует инсулина , пептида YY и GLP-1 ; секреции [50] [ нужен неосновной источник ] следовательно, TAAR1 потенциально является биологической мишенью для лечения ожирения и диабета . [50] [ нужен неосновной источник ]
Отсутствие TAAR1 существенно не влияет на сексуальную мотивацию и обычные липидные и метаболические биохимические параметры крови, что позволяет предположить, что будущие методы лечения на основе TAAR1 должны иметь благоприятный профиль безопасности. [51]
Примечания
[ редактировать ]- ^ В нейронах дофамина , норадреналина и серотонина первичными мембранными переносчиками являются DAT , NET и SERT соответственно. [12]
- ^ TAAR1-D2sh представляет собой пресинаптический гетеродимер , который включает перемещение TAAR1 из внутриклеточного пространства в D2sh на плазматической мембране , повышенное сродство связывания агониста D2sh и передачу сигнала через путь кальций- PKC - NFAT и G-белка независимую от PKB - GSK3. путь. [7] [27]
Ссылки
[ редактировать ]- ^ Jump up to: а б с GRCh38: Версия Ensembl 89: ENSG00000146399 – Ensembl , май 2017 г.
- ^ Jump up to: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000056379 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ген Энтрез: рецептор 1, связанный с следовым амином TAAR1» .
- ^ Jump up to: а б с д и ж г час я дж Магуайр Джей Джей, Davenport AP (20 февраля 2018 г.). «Рецептор следовых аминов: рецептор ТА 1 » . Руководство IUPHAR/BPS по ФАРМАКОЛОГИИ . Международный союз фундаментальной и клинической фармакологии . Проверено 16 июля 2018 г.
Распределение тканей
ЦНС (зависит от региона) и некоторые периферические ткани:
Желудок > миндалевидное тело, почки, легкие, тонкий кишечник > мозжечок, дорсальный ганглий, гиппокамп, гипоталамус, печень, продолговатый мозг, поджелудочная железа, гипофиз, ретикулярная формация моста, простата, скелетные мышцы, селезенка. ...
Лейкоциты ... β-клетки островков поджелудочной железы ... Первичные тонзиллярные В-клетки ... Циркулирующие лейкоциты здоровых людей (усиление регуляции происходит при добавлении фитогемагглютинина).
Вид: Человек...
В головном мозге (мыши, макак-резус) рецептор ТА1 локализуется в нейронах момаминергических путей, и появляются доказательства модулирующей роли ТА1 в функционировании этих систем. Совместная экспрессия ТА1 с переносчиком дофамина (либо внутри того же нейрона, либо в соседних нейронах) предполагает прямую/непрямую модуляцию дофаминергической функции ЦНС. В клетках, экспрессирующих как человеческий TA1, так и переносчик моноаминов (DAT, SERT или NET), передача сигналов через TA1 усиливается [26,48,50–51]. ...
Функциональные анализы ...
Мобилизация внутреннего кальция в клетках RD-HGA16, трансфицированных немодифицированным человеческим ТА1
Измеренный ответ: Увеличение цитопазматического кальция...
Измерение уровней цАМФ в культивируемых астроцитах человека.
Измеренный ответ: накопление цАМФ...
Активация лейкоцитов
Вид: Человек
Ткань: ПМН, Т- и В-клетки.
Измеряемый ответ: хемотаксическая миграция к лигандам ТА1 (β-фенилэтиламин, тирамин и 3-йодтиронамин), индуцированная следами аминов секреция IL-4 (Т-клетки) и индуцированная следами аминов регуляция экспрессии РНК-маркера Т-клеток, следы аминов, индуцированные секрецией IgE в В-клетки. - ^ Jump up to: а б с д и ж г час Гранди Д.К., Миллер ГМ, Ли Дж.С. (февраль 2016 г.). « TAARgeting наркомания» - Аламо является свидетелем еще одной революции: обзор пленарного симпозиума Конференции по поведению, биологии и химии 2015 года» . Зависимость от алкоголя и наркотиков . 159 : 9–16. doi : 10.1016/j.drugalcdep.2015.11.014 . ПМЦ 4724540 . ПМИД 26644139 .
TAAR1 является рецептором с высоким сродством к METH/AMPH и DA... Это оригинальное наблюдение взаимодействия TAAR1 и DA-D2R впоследствии было подтверждено и дополнено наблюдениями о том, что оба рецептора могут гетеродимеризоваться друг с другом при определенных условиях... Дополнительные DA Взаимодействия D2R/TAAR1 с функциональными последствиями выявляются результатами экспериментов, демонстрирующих, что помимо пути цАМФ/PKA (Panas et al., 2012) стимуляция TAAR1-опосредованной передачи сигналов связана с активацией пути Ca++/PKC/NFAT. (Panas et al., 2012) и DA-D2R-связанный, независимый от G-белка сигнальный путь AKT/GSK3 (Espinoza et al., 2015; Harmeier et al., 2015), так что одновременная активация TAAR1 и DA DR2R может привести к при снижении передачи сигналов по одному пути (например, цАМФ/PKA), но сохранении передачи сигналов по другому пути (например, Ca++/PKC/NFA)
- ^ Jump up to: а б Цзин Л., Ли JX (август 2015 г.). «Рецептор 1, связанный с следами аминов: многообещающая мишень для лечения зависимости от психостимуляторов» . Евро. Дж. Фармакол . 761 : 345–352. дои : 10.1016/j.ejphar.2015.06.019 . ПМЦ 4532615 . ПМИД 26092759 .
TAAR1 в основном локализован во внутриклеточных компартментах как в нейронах (Miller, 2011), так и в глиальных клетках (Cisneros and Ghorpade, 2014) и в периферических тканях (Grandy, 2007). Существующие данные предоставили надежные доклинические доказательства, подтверждающие развитие TAAR1. агонисты как потенциальное лечение злоупотребления психостимуляторами и зависимости. ... Учитывая, что TAAR1 в основном расположен во внутриклеточных компартментах, и предполагается, что существующие агонисты TAAR1 получают доступ к рецепторам путем транслокации внутрь клетки (Miller, 2011), в будущем усилия по разработке и разработке лекарств, возможно, потребуют принятия стратегий лекарственного лечения. доставка во внимание (Раджендран и др., 2010).
- ^ Jump up to: а б с д и ж г час я Берри М.Д., Гайнетдинов Р.Р., Хонер М.К., Шахид М. (декабрь 2017 г.). «Фармакология человеческих рецепторов, связанных с следами аминов: терапевтические возможности и проблемы» . Фармакология и терапия . 180 : 161–180. doi : 10.1016/j.pharmthera.2017.07.002 . ПМИД 28723415 .
- ^ Jump up to: а б с д и ж г час Боровский Б., Адхам Н., Джонс К.А., Раддац Р., Артымишин Р., Огозалек К.Л. и др. (июль 2001 г.). «Следовые амины: идентификация семейства рецепторов млекопитающих, связанных с G-белком» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (16): 8966–8971. Бибкод : 2001PNAS...98.8966B . дои : 10.1073/pnas.151105198 . ПМЦ 55357 . ПМИД 11459929 .
- ^ Jump up to: а б с д и ж Бунцов Дж.Р., Сондерс М.С., Арттамангкул С., Харрисон Л.М., Чжан Г., Куигли Д.И. и др. (декабрь 2001 г.). «Амфетамин, 3,4-метилендиоксиметамфетамин, диэтиламид лизергиновой кислоты и метаболиты нейротрансмиттеров катехоламинов являются агонистами аминных рецепторов крысы». Молекулярная фармакология . 60 (6): 1181–1188. дои : 10.1124/моль.60.6.1181 . ПМИД 11723224 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v Миллер GM (январь 2011 г.). «Новая роль рецептора 1, связанного с следами аминов, в функциональной регуляции переносчиков моноаминов и дофаминергической активности» . Журнал нейрохимии . 116 (2): 164–176. дои : 10.1111/j.1471-4159.2010.07109.x . ПМК 3005101 . ПМИД 21073468 .
- ^ Jump up to: а б с д и ж Линдеманн Л., Эбелинг М., Краточвил Н.А., Бунцов Дж.Р., Гранди Д.К., Хёнер М.К. (март 2005 г.). «Рецепторы, связанные с следовыми аминами, образуют структурно и функционально различные подсемейства новых рецепторов, связанных с G-белком». Геномика . 85 (3): 372–385. дои : 10.1016/j.ygeno.2004.11.010 . ПМИД 15718104 .
- ^ Jump up to: а б с д Сиснерос И.Е., Горпаде А. (октябрь 2014 г.). «Метамфетамин и нейротоксичность, вызванная ВИЧ-1: роль передачи сигналов цАМФ, связанного с следами амина, в астроцитах» . Нейрофармакология . 85 : 499–507. doi : 10.1016/j.neuropharm.2014.06.011 . ПМК 4315503 . ПМИД 24950453 .
Сверхэкспрессия TAAR1 значительно снизила уровни EAAT-2 и клиренс глутамата... Лечение METH активировало TAAR1, что привело к внутриклеточному цАМФ в астроцитах человека и модулировало способность к клиренсу глутамата. Более того, молекулярные изменения уровней TAAR1 астроцитов соответствуют изменениям уровней и функции EAAT-2 астроцитов.
- ^ Роджерс Т.Дж. (2012). «Молекулярная основа передачи сигналов нейроиммунных рецепторов» . J Нейроиммунный Фармакол . 7 (4): 722–724. дои : 10.1007/s11481-012-9398-4 . ПМК 4011130 . ПМИД 22935971 .
- ^ Jump up to: а б с д Панас М.В., Се З., Панас Х.Н., Хонер М.К., Валлендер Э.Дж., Миллер Г.М. (декабрь 2012 г.). «Отслеживание передачи сигналов рецептора 1, связанного с амином, в активированных лимфоцитах» . Журнал нейроиммунной фармакологии . 7 (4): 866–876. дои : 10.1007/s11481-011-9321-4 . ПМЦ 3593117 . ПМИД 22038157 .
- ^ Jump up to: а б Бабушите А., Коттхофф М., Фидлер Дж., Краутвурст Д. (март 2013 г.). «Биогенные амины активируют лейкоциты крови через следовые аминоассоциированные рецепторы TAAR1 и TAAR2». Журнал биологии лейкоцитов . 93 (3): 387–394. дои : 10.1189/jlb.0912433 . ПМИД 23315425 . S2CID 206996784 .
- ^ Гайнетдинов Р.Р., Хонер MC, Берри MD (июль 2018 г.). «Микроамины и их рецепторы» . Фармакологические обзоры . 70 (3): 549–620. дои : 10.1124/пр.117.015305 . ПМИД 29941461 .
- ^ Се Зи, генеральный директор Miller (ноябрь 2009 г.). «Отследить аминоассоциированный рецептор 1 как моноаминергический модулятор в мозге» . Биохимическая фармакология . 78 (9): 1095–1104. дои : 10.1016/j.bcp.2009.05.031 . ПМЦ 2748138 . ПМИД 19482011 .
- ^ «ТААР1» . Атлас белков человека . Проверено 24 августа 2017 г.
- ^ Jump up to: а б с д Бугда Гвилт К., Гонсалес Д.П., Оллифф Н., Оллер Х., Хоффинг Р., Пузан М. и др. (декабрь 2019 г.). «Действие следовых аминов на ось мозг-кишечник-микробиом через рецептор-1, связанный с микроэлементами (TAAR1)» (PDF) . Клеточная и молекулярная нейробиология . 40 (2): 191–201. дои : 10.1007/s10571-019-00772-7 . ПМИД 31836967 . S2CID 209339614 .
- ^ Се З., Уэстморленд С.В., Бан М.Э., Чен Г.Л., Ян Х., Валлендер Э.Дж. и др. (апрель 2007 г.). «Передача сигнала рецептора 1, связанного с следами амина у макаки-резуса: усиление переносчиками моноаминов и ослабление ауторецептором D2 in vitro». Журнал фармакологии и экспериментальной терапии . 321 (1): 116–127. дои : 10.1124/jpet.106.116863 . ПМИД 17234900 . S2CID 578835 .
- ^ Либерлес С.Д., Бак Л.Б. (август 2006 г.). «Второй класс хемосенсорных рецепторов обонятельного эпителия». Природа . 442 (7103): 645–650. Бибкод : 2006Natur.442..645L . дои : 10.1038/nature05066 . ПМИД 16878137 . S2CID 2864195 .
- ^ Jump up to: а б с Барак Л.С., Салахпур А., Чжан Х., Масри Б., Сотникова Т.Д., Рэмси А.Дж. и др. (сентябрь 2008 г.). «Фармакологическая характеристика экспрессируемого на мембране человеческого следового аминоассоциированного рецептора 1 (TAAR1) с помощью биолюминесцентного биосенсора цАМФ для резонансной передачи энергии» . Молекулярная фармакология . 74 (3): 585–594. дои : 10.1124/моль.108.048884 . ПМЦ 3766527 . ПМИД 18524885 .
- ^ Jump up to: а б с Лам В.М., Эспиноза С., Герасимов А.С., Гайнетдинов Р.Р., Салахпур А. (июнь 2015 г.). «Фармакология рецептора 1, связанного с трассировочным амином, in vivo». Евро. Дж. Фармакол . 763 (Часть Б): 136–42. дои : 10.1016/j.ejphar.2015.06.026 . ПМИД 26093041 .
Периферическая и иммунная локализация/функции TAAR1. Важно отметить, что помимо головного мозга TAAR1 также экспрессируется в спинном мозге (Gozal et al., 2014) и периферии (Revel et al., 2012c). Показано, что TAAR1 экспрессируется и регулирует иммунную функцию в лейкоцитах макак-резус (Babusyte et al., 2013; Nelson et al., 2007; Panas et al., 2012). В гранулоцитах TAAR1 необходим для хемотаксической миграции клеток к агонистам TAAR1. Кроме того, передача сигналов TAAR1 в B- и T-клетках может запускать высвобождение иммуноглобулина и цитокинов соответственно (Babusyte et al., 2013). TAAR1 также экспрессируется в островках Лангерганса, желудке и кишечнике, судя по картинам окрашивания LacZ, проведенным на мышах TAAR1-KO LacZ (Revel et al., 2012c). Интересно, что введение селективного частичного агониста TAAR1 RO5263397 обращает вспять побочный эффект увеличения веса, наблюдаемый при приеме антипсихотика оланзапина, указывая на то, что периферическая передача сигналов TAAR1 может регулировать метаболический гомеостаз (Revel et al., 2012b). ...
Транспортеры моноаминов и подсемейство носителей SLC22A транспортируют лиганд TAAR1: исследования с использованием TAAR1 макаки-резуса показали, что этот рецептор взаимодействует с переносчиками моноаминов DAT, SERT и NET в клетках HEK (Miller et al., 2005; Xie and Miller, 2007; Xie). и др., 2007). Было высказано предположение, что взаимодействие TAAR1 с этими транспортерами может обеспечить механизм, с помощью которого лиганды TAAR1 могут проникать в цитоплазму и связываться с TAAR1 во внутриклеточных компартментах. Недавнее исследование показало, что в мотонейронах новорожденных крыс для специфической передачи сигналов следовых аминов требуется наличие и функция трансмембранного растворенного переносчика SLC22A, но не транспортеров моноаминов (DAT, SERT и NET) (Gozal et al., 2014). . В частности, было показано, что добавление β-PEA, тирамина или триптамина индуцирует двигательно-подобную активность (LLA) этих нейронов в присутствии N-метил-D-аспартата. Во временном отношении было обнаружено, что индукция LLA следами аминов задерживается по сравнению с LLA, индуцируемой серотонином и норэпинефрином, что указывает на то, что целевой сайт для следовых аминов не расположен на плазматической мембране и, возможно, может быть внутриклеточным. Важно отметить, что блокирование SLC22A пентамидином устраняло LLA, индуцированную следами аминов, что указывает на то, что LLA, индуцированная следами аминов, не действует на рецепторы, обнаруженные на плазматической мембране, но требует их транспорта в цитозоль с помощью SLC22A для индукции LLA. - ^ Jump up to: а б Динтер Дж., Мюльхаус Дж., Якоби С.Ф., Винчоль К.Л., Кёстер М., Мейстер Дж. и др. (июнь 2015 г.). «3-йодтиронамин дифференциально модулирует передачу сигналов, опосредованную α-2A-адренергическими рецепторами» . Дж. Мол. Эндокринол . 54 (3): 205–216. дои : 10.1530/JME-15-0003 . ПМИД 25878061 .
Более того, в гетероолигомерах ADRA2A/TAAR1 способность NorEpi стимулировать передачу сигналов Gi/o снижается при совместной стимуляции с 3-T1AM. Таким образом, настоящее исследование указывает на сложный спектр модификаций передачи сигналов, опосредованных 3-T1AM на различных рецепторах, связанных с G-белком.
- ^ Хармейер А., Обермюллер С., Мейер К.А., Ревель Ф.Г., Буши Д., Шабоз С. и др. (2015). «Активация рецептора 1, связанного с следовым амином, подавляет передачу сигналов GSK3β от гетеромеров TAAR1 и D2R». Eur Нейропсихофармакол . 25 (11): 2049–2061. дои : 10.1016/j.euroneuro.2015.08.011 . ПМИД 26372541 . S2CID 41667764 .
Взаимодействие TAAR1 с D2R изменяло субклеточную локализацию TAAR1 и повышало аффинность связывания агониста D2R.
- ^ Jump up to: а б с д Зукки Р., Кьеллини Дж., Сканлан Т.С., Гранди Д.К. (декабрь 2006 г.). «Отследить аминоассоциированные рецепторы и их лиганды» . Британский журнал фармакологии . 149 (8): 967–978. дои : 10.1038/sj.bjp.0706948 . ПМК 2014643 . ПМИД 17088868 .
Другие биогенные амины присутствуют в центральной нервной системе в очень низких концентрациях, порядка 0,1–10 нм, что составляет <1% от общего количества биогенных аминов (Berry, 2004). Для этих соединений был введен термин «следовые амины». Молекулы, которые обычно считаются следовыми аминами, хотя и несколько нечетко определены, включают паратирамин, метатирамин, триптамин, β-фенилэтиламин, параоктопамин и метаоктопамин (Berry, 2004) (рис. 2).
- ^ Jump up to: а б Се З, генеральный директор Miller (июль 2009 г.). «Рецепторный механизм действия метамфетамина в регуляции транспортера дофамина в мозге» . Журнал фармакологии и экспериментальной терапии . 330 (1): 316–325. дои : 10.1124/jpet.109.153775 . ПМК 2700171 . ПМИД 19364908 .
- ^ Рикли А., Копф С., Хонер М.К., Лихти М.Э. (июль 2015 г.). «Фармакологический профиль новых психоактивных бензофуранов» . Британский журнал фармакологии . 172 (13): 3412–3425. дои : 10.1111/bph.13128 . ПМК 4500375 . ПМИД 25765500 .
- ^ Jump up to: а б Уэйнскотт Д.Б., Литтл С.П., Инь Т., Ту Ю., Рокко В.П., Хе JX и др. (январь 2007 г.). «Фармакологическая характеристика клонированного человеческого следового аминоассоциированного рецептора 1 (TAAR1) и доказательства видовых различий с крысиным TAAR1». Журнал фармакологии и экспериментальной терапии . 320 (1): 475–485. дои : 10.1124/jpet.106.112532 . ПМИД 17038507 . S2CID 10829497 .
Несколько серий замещенных фенилэтиламинов исследовали на активность в отношении TAAR1 человека (табл. 2). Неожиданным открытием стала эффективность фенилэтиламинов с заместителями в положении фенила C2 по сравнению с их соответствующими C4-замещенными аналогами. В каждом случае, за исключением гидроксильного заместителя, C2-замещенное соединение имело в 8-27 раз более высокую эффективность, чем C4-замещенное соединение. С3-замещенное соединение в каждом гомологическом ряду обычно было в 2-5 раз менее эффективным, чем 2-замещенное соединение, за исключением гидроксильного заместителя. Самым сильным из 2-замещенных фенилэтиламинов был 2-хлор-β-ПЭА, за ним следовали 2-фтор-β-ПЭА, 2-бром-β-ПЭА, 2-метокси-β-ПЭА, 2-метил-β-. ПЭА, а затем 2-гидрокси-β-ПЭА.
Также было исследовано влияние β-углеродного замещения на боковую цепь фенилэтиламина (табл. 3). β-метиловый заместитель хорошо переносился по сравнению с β-ПЭА. Фактически, S-(–)-β-метил-β-PEA был столь же эффективен, как и β-PEA, в отношении TAAR1 человека. Однако замена β-гидроксила не переносилась по сравнению с β-PEA. В обоих случаях β-замещения была продемонстрирована энантиомерная селективность.
В отличие от замены метила на β-углероде, замена α-метила снижает эффективность примерно в 10 раз для d-амфетамина и в 16 раз для l-амфетамина по сравнению с β-ПЭА (таблица 4). Замещение N-метилом переносилось довольно хорошо; однако N,N-диметилзамещения не было. - ^ Галлей Г., Бьюрье А., Декоре Г., Герглер А., Хаттер Р., Мор С. и др. (2016). «Открытие и характеристика 2-аминооксазолинов как высокоэффективных, селективных и перорально активных агонистов TAAR1» . ACS Med Chem Lett . 7 (2): 192–197. doi : 10.1021/acsmedchemlett.5b00449 . ПМЦ 4753552 . ПМИД 26985297 .
- ^ Jump up to: а б Ревель Ф.Г., Моро Дж.Л., Гайнетдинов Р.Р., Брадая А., Сотникова Т.Д., Мори Р. и др. (май 2011 г.). «Активация TAAR1 модулирует моноаминергическую нейротрансмиссию, предотвращая гипердофаминергическую и гипоглутаматергическую активность» . Учеб. Натл. акад. наук. США . 108 (20): 8485–8490. Бибкод : 2011PNAS..108.8485R . дои : 10.1073/pnas.1103029108 . ПМК 3101002 . ПМИД 21525407 .
- ^ Ревель Ф.Г., Моро Дж.Л., Гайнетдинов Р.Р., Феррагуд А., Веласкес-Санчес С., Сотникова Т.Д. и др. (июнь 2012 г.). «Частичный агонизм рецептора 1, связанного с следовым амином, открывает новую парадигму нейропсихиатрической терапии». Биологическая психиатрия . 72 (11): 934–942. doi : 10.1016/j.biopsych.2012.05.014 . ПМИД 22705041 . S2CID 27334223 .
- ^ Чичеро Э., Франческони В., Казини Б., Казале М., Канов Е., Герасимов А.С. и др. (ноябрь 2023 г.). «Открытие гуанфацина как нового агониста TAAR1: стратегия комбинирования посредством исследований молекулярного моделирования и биологических анализов» . Фармацевтика . 16 (11): 1632. doi : 10.3390/ph16111632 . ПМЦ 10674299 . ПМИД 38004497 .
- ^ Гуршахани Х., Джолас Т., Мартин М., Котье С., Хьюз С., Макфадден В. и др. (2023). «Доклиническая фармакология солриамфетола: потенциальные механизмы стимулирования пробуждения». Спектры ЦНС . 28 (2): 222. дои : 10.1017/S1092852923001396 . ISSN 1092-8529 .
Функциональные исследования in vitro показали агонистическую активность солриамфетола в отношении рецепторов TAAR1 человека, мышей и крыс. Значения EC50 hTAAR1 (10–16 мкМ) находились в пределах клинически наблюдаемого диапазона терапевтических концентраций солриамфетола в плазме и перекрывались с наблюдаемой ингибирующей активностью солриамфетола в отношении DAT/NET in vitro. Агонистическая активность TAAR1 была уникальной для солриамфетола; ни модафинил WPA, ни ингибитор DAT/NET бупропион не обладали агонистической активностью TAAR1.
- ^ Брадая А., Трубе Г., Сталдер Х., Норкросс Р.Д., Озмен Л., Веттштейн Дж.Г. и др. (ноябрь 2009 г.). «Селективный антагонист EPPTB выявляет TAAR1-опосредованные регуляторные механизмы в дофаминергических нейронах мезолимбической системы» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (47): 20081–20086. Бибкод : 2009PNAS..10620081B . дои : 10.1073/pnas.0906522106 . ПМЦ 2785295 . ПМИД 19892733 .
- ^ Jump up to: а б Эйден Л.Е., Вэйхэ Э. (январь 2011 г.). «VMAT2: динамический регулятор функции моноаминергических нейронов мозга, взаимодействующий с наркотиками» . Энн. Н-Й Акад. Наука . 1216 (1): 86–98. Бибкод : 2011NYASA1216...86E . дои : 10.1111/j.1749-6632.2010.05906.x . ПМЦ 4183197 . ПМИД 21272013 .
VMAT2 является везикулярным транспортером ЦНС не только для биогенных аминов DA, NE, EPI, 5-HT и HIS, но, вероятно, также для следовых аминов TYR, PEA и тиронамина (THYR). ЦНС млекопитающих можно было бы идентифицировать как нейроны, экспрессирующие VMAT2 для хранения и биосинтетический фермент декарбоксилазу ароматических аминокислот (AADC). ... Высвобождение DA из синапсов AMPH требует как действия на VMAT2 для высвобождения DA в цитоплазму, так и согласованного высвобождения DA из цитоплазмы посредством «обратного транспорта» через DAT.
- ^ Зульцер Д., Крэгг С.Дж., Райс М.Э. (август 2016 г.). «Нейротрансмиссия стриарного дофамина: регуляция высвобождения и поглощения» . Базальные ганглии . 6 (3): 123–148. дои : 10.1016/j.baga.2016.02.001 . ПМК 4850498 . ПМИД 27141430 .
Несмотря на трудности определения pH синаптических везикул, градиент протонов через мембрану везикул имеет фундаментальное значение для ее функции. Воздействие протонофоров на изолированные везикулы катехоламинов разрушает градиент pH и быстро перераспределяет медиатор изнутри пузырька наружу. ... Амфетамин и его производные, такие как метамфетамин, представляют собой слабоосновные соединения и являются единственным широко используемым классом наркотиков, которые, как известно, вызывают высвобождение медиатора по неэкзоцитическому механизму. В качестве субстратов как для DAT, так и для VMAT амфетамины могут переноситься в цитозоль, а затем изолироваться в везикулах, где они действуют, разрушая везикулярный градиент pH.
- ^ Jump up to: а б Ледонн А., Берретта Н., Даволи А., Риццо Г.Р., Бернарди Дж., Меркури Н.Б. (2011). «Электрофизиологические эффекты следовых аминов на мезэнцефальные дофаминергические нейроны» . Границы системной нейронауки . 5 : 56. дои : 10.3389/fnsys.2011.00056 . ПМК 3131148 . ПМИД 21772817 .
торможение стрельбы из-за увеличения выброса дофамина; (б) снижение тормозных реакций, опосредованных рецепторами D2 и GABAB (возбуждающие эффекты вследствие растормаживания); и (c) прямая активация GIRK-каналов, опосредованная рецептором TA1, которая вызывает гиперполяризацию клеточной мембраны.
- ^ Jump up to: а б mct (28 января 2012 г.). «ТААР1» . ГенАтлас . Парижский университет . Проверено 29 мая 2014 г.
« • тонически активирует внутренние выпрямляющие K(+) каналы, что снижает базальную частоту импульсов дофаминовых (DA) нейронов вентральной покрышки (VTA)» - ^ Андерхилл С.М., Уилер Д.С., Ли М., Уоттс С.Д., Ингрэм С.Л., Амара С.Г. (июль 2014 г.). «Амфетамин модулирует возбуждающую нейротрансмиссию посредством эндоцитоза транспортера глутамата EAAT3 в дофаминовых нейронах» . Нейрон . 83 (2): 404–416. дои : 10.1016/j.neuron.2014.05.043 . ПМК 4159050 . ПМИД 25033183 .
AMPH также увеличивает внутриклеточный кальций (Gnegy et al., 2004), что связано с активацией кальмодулина/CamKII (Wei et al., 2007), а также модуляцией и транспортировкой DAT (Fog et al., 2006; Sakrikar et al., 2012). ). ... Например, AMPH увеличивает внеклеточный глутамат в различных областях мозга, включая полосатое тело, VTA и NAc (Del Arco et al., 1999; Kim et al., 1981; Mora and Porras, 1993; Xue et al., 1996). , но не установлено, можно ли объяснить это изменение увеличением синаптического высвобождения или снижением клиренса глутамата. ... DHK-чувствительное поглощение EAAT2 не изменялось под действием AMPH (рис. 1А). Оставшийся транспорт глутамата в этих культурах среднего мозга, вероятно, опосредован EAAT3, и этот компонент был значительно уменьшен AMPH.
- ^ Воган Р.А., Фостер Дж.Д. (сентябрь 2013 г.). «Механизмы регуляции транспортера дофамина в норме и при заболеваниях» . Тренды Фармакол. Наука . 34 (9): 489–496. дои : 10.1016/j.tips.2013.07.005 . ПМЦ 3831354 . ПМИД 23968642 .
АМФ и МЕТН также стимулируют отток ДА, который считается решающим элементом их аддиктивных свойств [80], хотя механизмы, по-видимому, не идентичны для каждого препарата [81]. Эти процессы являются PKCβ- и CaMK-зависимыми [72, 82], а у мышей с нокаутом PKCβ наблюдается снижение индуцированного AMPH оттока, что коррелирует со снижением индуцированной AMPH локомоции [72].
- ^ Магуайр Дж.Дж., Паркер В.А., Фурд С.М., Боннер Т.И., Нойбиг Р.Р., Давенпорт А.П. (март 2009 г.). «Международный союз фармакологии. LXXII. Рекомендации по номенклатуре следовых аминных рецепторов» . Фармакологические обзоры . 61 (1): 1–8. дои : 10.1124/пр.109.001107 . ПМК 2830119 . ПМИД 19325074 .
- ^ Jump up to: а б с Линдеманн Л., Хонер MC (май 2005 г.). «Ренессанс следовых аминов, вдохновленный новым семейством GPCR». Тренды Фармакол. Наука . 26 (5): 274–281. дои : 10.1016/j.tips.2005.03.007 . ПМИД 15860375 .
Нарушение регуляции уровней ТА связано с несколькими заболеваниями, что делает соответствующих членов семейства TAAR потенциальными мишенями для разработки лекарств. В этой статье мы фокусируемся на значении ТА и их рецепторов для расстройств, связанных с нервной системой, а именно шизофрении и депрессии; однако ТА также связаны с другими заболеваниями, такими как мигрень, синдром дефицита внимания и гиперактивности, злоупотребление психоактивными веществами и расстройства пищевого поведения [7,8,36]. Клинические исследования сообщают о повышенных уровнях β-ПЭА в плазме у пациентов, страдающих острой шизофренией [37] и повышенной экскреции β-ПЭА с мочой у параноидальных шизофреников [38], что подтверждает роль ТА при шизофрении. В результате этих исследований β-ПЭА был назван «эндогенным амфетамином» организма [39].
- ^ Jump up to: а б Сотникова Т.Д., Карон М.Г., Гайнетдинов Р.Р. (август 2009 г.). «Отследить аминоассоциированные рецепторы как новые терапевтические мишени» . Мол. Фармакол . 76 (2): 229–235. дои : 10.1124/моль.109.055970 . ПМЦ 2713119 . ПМИД 19389919 .
Хотя функциональная роль следовых аминов у млекопитающих остается в значительной степени загадочной, было отмечено, что уровни следовых аминов могут быть изменены при различных заболеваниях человека, включая шизофрению, болезнь Паркинсона, синдром дефицита внимания и гиперактивности (СДВГ), синдром Туретта и фенилкетонурию (Бултон). Сэндлер и др., 1980). Обычно считалось, что следовые количества аминов влияют на моноаминовую систему опосредованно через взаимодействие с переносчиками плазматической мембраны (такими как переносчик дофамина на плазматической мембране (DAT)) и везикулярным хранилищем (Premont et al., 2001; Branchek and Blackburn, 2003; Berry, 2004; Сотникова и др., 2004). ...
Кроме того, мыши с дефицитом DAT представляют собой модель для изучения ингибирующего действия амфетаминов на гиперактивность, особенность амфетаминов, которая считается важной для их терапевтического действия при СДВГ (Гаинетдинов и др., 1999; Гаинетдинов и Карон, 2003). Следует также отметить, что наиболее известный агонист TAAR1, β-PEA, обладает способностью амфетамина индуцировать ингибирование дофаминзависимой гиперактивности мышей DAT-KO (Гаинетдинов и др., 1999; Сотникова и др., 2004). ).
Кроме того, если можно будет доказать, что TAAR1 является медиатором некоторых действий амфетамина in vivo, разработка новых селективных агонистов и антагонистов TAAR1 может обеспечить новый подход к лечению состояний, связанных с амфетамином, таких как зависимость и/или расстройства, при которых амфетамин используется в терапевтических целях. В частности, поскольку амфетамин на протяжении многих лет остается наиболее эффективным фармакологическим средством лечения СДВГ, необходимо изучить потенциальную роль TAAR1 в механизме «парадоксальной» эффективности амфетамина при этом расстройстве. - ^ Берри, доктор медицины (январь 2007 г.). «Потенциал следовых аминов и их рецепторов для лечения неврологических и психиатрических заболеваний». Преподобный Недавние клинические испытания . 2 (1): 3–19. CiteSeerX 10.1.1.329.563 . дои : 10.2174/157488707779318107 . ПМИД 18473983 .
изменения в следовых аминах, в частности PE, были идентифицированы как возможный фактор возникновения синдрома дефицита внимания/гиперактивности (СДВГ) [5, 27, 43, 78]. Было показано, что ПЭ вызывает у экспериментальных животных гиперактивность и агрессию, две основные клинические особенности СДВГ [100]. Гиперактивность также является симптомом фенилкетонурии, которая, как обсуждалось выше, связана с заметно повышенным обменом ПЭ [44]. Кроме того, амфетамины, которые имеют клиническое применение при СДВГ, являются хорошими лигандами следовых аминорецепторов [2]. Возможное значение в этом аспекте имеет модафанил, который показал положительные эффекты у пациентов с СДВГ [101] и, как сообщается, усиливает активность PE в TAAR1 [102]. И наоборот, метилфенидат, который также клинически полезен при СДВГ, показал низкую эффективность в отношении рецептора TAAR1 [2]. В этом отношении стоит отметить, что улучшение функционирования TAAR1, наблюдаемое при приеме модафанила, не было результатом прямого взаимодействия с TAAR1 [102].
Недавно были получены более прямые доказательства роли следовых аминов в развитии СДВГ. Сообщалось, что уровни ПЭ в моче снижаются у пациентов с СДВГ по сравнению как с контрольной группой, так и с пациентами с аутизмом [103-105]. Недавно также были получены данные о снижении уровня ПЭ в мозге пациентов с СДВГ [4]. Кроме того, сообщалось о снижении уровня метаболита ФЭ фенилуксусной кислоты и предшественников фенилаланина и тирозина в моче и плазме наряду со снижением уровня тирамина в плазме [103]. После лечения метилфенидатом у пациентов, которые ответили положительно, наблюдалась нормализация ПЭ в моче, тогда как у пациентов, не ответивших на лечение, не наблюдалось никаких изменений по сравнению с исходными значениями [105]. - ^ Жуков И.С., Кубарская Л.Г., Тиссен И.Ю., Козлова А.А., Дагаев С.Г., Кашуро В.А. и др. (март 2020 г.). «Минимальные возрастные изменения в поведенческих и гематологических параметрах у мышей, нокаутных по рецептору 1, связанному с амином (TAAR1)». Клеточная и молекулярная нейробиология . 40 (2): 273–282. дои : 10.1007/s10571-019-00721-4 . ПМИД 31399838 . S2CID 199511689 .
- ^ Смит С.Б., Майкснер Д.В., Филлингим Р.Б., Слэйд Г., Грейсли Р.Х., Эмброуз К. и др. (февраль 2012 г.). «Крупное исследование ассоциаций генов-кандидатов выявляет генетические факторы риска и терапевтические цели фибромиалгии» . Артрит и ревматизм . 64 (2): 584–593. дои : 10.1002/арт.33338 . ПМЦ 3237946 . ПМИД 21905019 .
- ^ Jump up to: а б Рааб С., Ван Х., Улес С., Коул Н., Альварес-Санчес Р., Кюннеке Б. и др. (2016). «Инкретиноподобные эффекты низкомолекулярных агонистов рецептора 1, ассоциированных с амином» . Мол Метаб . 5 (1): 47–56. doi : 10.1016/j.molmet.2015.09.015 . ПМК 4703809 . ПМИД 26844206 .
- ^ Жуков И.С., Птуха М.А., Золотоверхая Е.А., Синица Е.Л., Тиссен И.Ю., Карпова И.В. и др. (май 2022 г.). «Оценка подхода к конспецифическим и биохимическим параметрам крови у мышей с нокаутом TAAR1» . Науки о мозге . 12 (5): 614. doi : 10.3390/brainsci12050614 . ПМЦ 9139149 . ПМИД 35625001 .
Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .
Внешние ссылки
[ редактировать ] СМИ, связанные с TAAR1 , на Викискладе?
СМИ, связанные с TAAR1 , на Викискладе?