Система вознаграждений




Система вознаграждения (мезокортиколимбическая цепь) представляет собой группу нейронных структур, ответственных за значимость стимулов (т. е. «желание»; желание или тяга к вознаграждению и мотивации), ассоциативное обучение (в первую очередь положительное подкрепление и классическое обусловливание ) и положительно-валентное обучение. эмоции , особенно те, в которых является удовольствие основным компонентом (например, радость , эйфория и экстаз ). [2] [3] Награда — это привлекательное и мотивационное свойство стимула, которое вызывает аппетитное поведение, также известное как поведение приближения и завершенное поведение. [2] Поощрительный стимул описывается как «любой стимул, объект, событие, действие или ситуация, которая потенциально может заставить нас приблизиться и употребить его, по определению является наградой». [2] В оперантном обусловливании стимулы вознаграждения действуют как положительное подкрепление ; [1] однако верно и обратное утверждение: положительное подкрепление вознаграждается. [1] [4] Система вознаграждения мотивирует животных приближаться к стимулам или совершать действия, повышающие физическую форму (секс, высокоэнергетическая пища и т. д.). Выживание большинства видов животных зависит от максимального контакта с полезными раздражителями и минимизации контакта с вредными раздражителями. Познание, основанное на вознаграждении, повышает вероятность выживания и воспроизводства, вызывая ассоциативное обучение, вызывая подход и законченное поведение, а также вызывая положительно валентные эмоции. [1] Таким образом, вознаграждение — это механизм, который развился, чтобы помочь повысить адаптивную приспособленность животных. [5] При наркозависимости некоторые вещества чрезмерно активируют цепь вознаграждения, что приводит к компульсивному поведению, связанному с поиском веществ, в результате синаптической пластичности этой цепи. [6]
Первичные вознаграждения представляют собой класс стимулов вознаграждения, которые способствуют выживанию человека и потомства , и они включают гомеостатические (например, вкусная еда ) и репродуктивные (например, сексуальный контакт и родительский вклад ) вознаграждения. [2] [7] Внутренние вознаграждения — это безусловные вознаграждения, которые привлекательны и мотивируют поведение, поскольку по своей сути доставляют удовольствие. [2] Внешние вознаграждения (например, деньги или победа любимой спортивной команды) — это обусловленные награды, которые привлекательны и мотивируют поведение, но не приносят удовольствия по своей сути. [2] [8] Внешние вознаграждения получают свою мотивационную ценность в результате усвоенной ассоциации (т. е. обусловленности) с внутренними вознаграждениями. [2] Внешние вознаграждения также могут вызывать удовольствие (например, эйфорию от выигрыша больших денег в лотерею) после того, как они классически обусловлены внутренними вознаграждениями. [2]
Определение
[ редактировать ]В нейробиологии система вознаграждения представляет собой совокупность структур мозга и нейронных путей, которые отвечают за познание, связанное с вознаграждением, включая ассоциативное обучение (в первую очередь классическое обусловливание и оперантное подкрепление ), значимость стимулов (т. е. мотивацию и «желание», желание или жажда вознаграждения), а также положительной валентности эмоции , особенно эмоции, связанные с удовольствием (т. е. гедонистическое «пристрастие»). [1] [3]
Термины, которые обычно используются для описания поведения, связанного с «желанием» или компонентом желания вознаграждения, включают аппетитное поведение, поведение приближения, подготовительное поведение, инструментальное поведение, упреждающее поведение и поиск. [9] Термины, которые обычно используются для описания поведения, связанного с компонентом вознаграждения «симпатия» или удовольствие, включают в себя законченное поведение и поведение принятия. [9]
Тремя основными функциями вознаграждений являются их способность:
- производить ассоциативное обучение (т.е. классическое обусловливание и оперантное подкрепление ); [1]
- влиять на принятие решений и стимулировать подходящее поведение (через присвоение мотивационной значимости поощряющим стимулам); [1]
- вызывать положительные эмоции, особенно удовольствие. [1]
Нейроанатомия
[ редактировать ]Обзор
[ редактировать ]Структуры мозга, составляющие систему вознаграждения, расположены преимущественно внутри корково-базальных ганглиев-таламо-кортикальной петли ; [10] часть петли базальных ганглиев управляет активностью системы вознаграждения. [10] Большинство путей, которые соединяют структуры внутри системы вознаграждения, представляют собой глутаматергические интернейроны , ГАМКергические средние шипиковые нейроны (MSN) и дофаминергические проекционные нейроны . [10] [11] хотя свой вклад вносят и другие типы проекционных нейронов (например, орексинергические проекционные нейроны). Система вознаграждения включает вентральную покрышку , вентральное полосатое тело (т. е. прилежащее ядро и обонятельный бугорок ), дорсальное полосатое тело (т. е. хвостатое ядро и скорлупу ), черную субстанцию (т. е. компактную и сетчатую части ), префронтальную кору , передняя поясная извилина , островковая кора , гиппокамп , гипоталамус (в частности, орексинергическое ядро в латеральном гипоталамусе ), таламус (множественные ядра), субталамическое ядро , бледный шар (как внешний, так и внутренний ), вентральный бледный шар , парабрахиальное ядро , миндалевидное тело и оставшаяся часть расширенной миндалевидного тела . [3] [10] [12] [13] [14] Ядро дорсального шва и мозжечок , по-видимому, модулируют некоторые формы познания, связанного с вознаграждением (например, ассоциативное обучение , мотивационная значимость и положительные эмоции ), а также поведение. [15] [16] [17] Латеродорсальное тегментальное ядро (LDT) , педункулопонтинное ядро (PPTg) и латеральное хабенула (LHb) (как прямо, так и опосредованно через ростромедиальное тегментальное ядро (RMTg) ) также способны индуцировать аверсивную и стимулирующую значимость через свои проекции на вентральную часть. тегментальная область (ВТА). [18] LDT и PPTg посылают глютаминергические проекции в VTA, которые образуют синапс на дофаминергических нейронах, оба из которых могут вызывать стимулирующую активность. LHb посылает глутаминергические проекции, большинство из которых синапсируются на ГАМКергических нейронах RMTg, что, в свою очередь, вызывает ингибирование дофаминергических нейронов VTA, хотя некоторые проекции LHb заканчиваются на интернейронах VTA. Эти проекции LHb активируются как аверсивными стимулами, так и отсутствием ожидаемого вознаграждения, а возбуждение LHb может вызвать отвращение. [19] [20] [21]
Большинство дофаминовых путей (т.е. нейронов, которые используют нейротрансмиттер дофамин для связи с другими нейронами), выходящих из вентральной области покрышки, являются частью системы вознаграждения; [10] в этих путях дофамин действует на D1-подобные рецепторы или D2-подобные рецепторы , стимулируя (D1-подобно) или ингибируя (D2-подобно) выработку цАМФ . [22] ГАМКергические также являются средние шипиковые нейроны полосатого тела компонентами системы вознаграждения. [10] Ядра глутаматергической проекции в субталамическом ядре, префронтальной коре, гиппокампе, таламусе и миндалевидном теле соединяются с другими частями системы вознаграждения через глутаматные пути. [10] Медиальный пучок переднего мозга , который представляет собой набор множества нервных путей, которые опосредуют вознаграждение за стимуляцию мозга (т. е. вознаграждение, полученное в результате прямой электрохимической стимуляции латерального гипоталамуса ), также является компонентом системы вознаграждения. [23]
Существуют две теории относительно активности прилежащего ядра и поколения симпатий и желаний. Гипотеза торможения (или гиперполяризации) предполагает, что прилежащее ядро оказывает тоническое тормозящее воздействие на нижележащие структуры, такие как вентральный паллидум, гипоталамус или вентральная область покрышки, и что при ингибировании MSN в прилежащем ядре (NAcc) эти структуры возбуждаются». «высвобождение» поведения, связанного с вознаграждением. В то время как рецепторов ГАМК агонисты способны вызывать реакции «приязни» и «желания» в прилежащем ядре, глутаминергические сигналы от базолатеральной миндалины , вентрального гиппокампа и медиальной префронтальной коры могут стимулировать выраженность стимулов. Более того, хотя большинство исследований показывают, что нейроны NAcc уменьшают возбуждение в ответ на вознаграждение, ряд исследований обнаруживает противоположную реакцию. Это привело к выдвижению гипотезы расторможения (или деполяризации), которая предполагает, что возбуждение или нейроны NAcc или, по крайней мере, некоторые их подмножества управляют поведением, связанным с вознаграждением. [3] [24] [25]
После почти 50 лет исследований вознаграждения за стимуляцию мозга эксперты подтвердили, что десятки участков мозга поддерживают внутричерепную самостимуляцию . К регионам относятся латеральный гипоталамус и медиальные пучки переднего мозга, которые особенно эффективны. Стимуляция там активирует волокна, образующие восходящие пути; Восходящие пути включают мезолимбический дофаминовый путь , который проецируется от вентральной покрышки к прилежащему ядру . Существует несколько объяснений того, почему мезолимбический путь дофамина играет центральную роль в цепях, обеспечивающих вознаграждение. Во-первых, наблюдается заметное увеличение высвобождения дофамина по мезолимбическому пути, когда животные участвуют в внутричерепной самостимуляции. [5] Во-вторых, эксперименты постоянно показывают, что вознаграждение за стимуляцию мозга стимулирует усиление путей, которые обычно активируются естественными вознаграждениями , а вознаграждение за лекарства или внутричерепная самостимуляция могут вызвать более мощную активацию центральных механизмов вознаграждения, поскольку они активируют центр вознаграждения напрямую, а не через него. периферические нервы . [5] [26] [27] В-третьих, когда животным вводят наркотики, вызывающие привыкание, или они занимаются естественным поведением, таким как кормление или сексуальная активность, в прилежащем ядре происходит заметный выброс дофамина. [5] Однако дофамин — не единственное соединение, отвечающее за вознаграждение в мозге.
Ключевой путь
[ редактировать ]
Вентральная покрышка
- Вентральная покрышка (VTA) важна для реагирования на стимулы и сигналы, указывающие на наличие награды. Поощрительные стимулы (и все наркотики, вызывающие привыкание) действуют на эту цепь, заставляя ВТА высвобождать сигналы дофамина в прилежащее ядро , прямо или косвенно. [ нужна ссылка ] ВТА имеет два важных пути: мезолимбический путь , проецирующийся в лимбические (стриарные) области и лежащий в основе мотивационного поведения и процессов, и мезокортикальный путь, проецирующийся в префронтальную кору, лежащий в основе когнитивных функций, таких как обучение внешним сигналам и т. д. [28]
- Дофаминергические нейроны в этой области преобразуют аминокислоту тирозин в ДОФА с помощью фермента тирозингидроксилазы , который затем преобразуется в дофамин с помощью фермента дофа-декарбоксилазы . [29]
Полосатое тело (Nucleus Accumbens)
- Полосатое тело широко участвует в приобретении и выявлении заученного поведения в ответ на полезный сигнал. VTA проецируется в полосатое тело и активирует ГАМК-ергические средние шипистые нейроны через рецепторы D1 и D2 в вентральном (прилежащее ядро) и дорсальном полосатом теле. [30]
- Вентральное полосатое тело (прилежащее ядро) широко участвует в приобретении поведения при подаче сигнала от VTA и формировании поведения при подаче от PFC. Оболочка NAc проецируется на паллидум и VTA, регулируя лимбические и вегетативные функции. Это модулирует подкрепляющие свойства стимулов и краткосрочные аспекты вознаграждения. Ядро NAc проецируется на черную субстанцию и участвует в развитии поведения, направленного на поиск вознаграждения, и его выражения. Он участвует в пространственном обучении, условном реагировании и импульсивном выборе; долгосрочные элементы вознаграждения. [28]
- Дорсальное полосатое тело участвует в обучении, дорзальное медиальное полосатое тело — в целенаправленном обучении, а дорсальное латеральное полосатое тело — в обучении на стимул-реакцию, лежащем в основе реакции Павлова. [31] При повторной активации стимулами прилежащее ядро может активировать дорсальное полосатое тело через внутриполосатую петлю. Переход сигналов от NAc к DS позволяет сигналам, связанным с вознаграждением, активировать DS без присутствия самого вознаграждения. Это может активировать тягу и поведение, направленное на вознаграждение (и ответственно за запуск рецидива зависимости во время воздержания). [32]
Префронтальная кора
- Дофаминергические нейроны VTA проецируются на PFC, активируя глутаминергические нейроны, которые проецируются на множество других областей, включая дорсальное полосатое тело и NAc, что в конечном итоге позволяет PFC опосредовать значимость и условное поведение в ответ на стимулы. [32]
- Примечательно, что воздержание от наркотиков, вызывающих зависимость, активирует PFC, глутаматергическую проекцию на NAc, что приводит к сильной тяге к ним и модулирует восстановление поведения зависимости, возникающего в результате воздержания. PFC также взаимодействует с VTA через мезокортикальный путь и помогает связать сигналы окружающей среды с вознаграждением. [32]
Гиппокамп
- Гиппокамп выполняет множество функций, в том числе создание и хранение воспоминаний. В цепи вознаграждения он служит контекстуальным воспоминаниям и связанным с ними сигналам. В конечном итоге это лежит в основе восстановления поведения, направленного на поиск вознаграждения, посредством сигналов и контекстных триггеров. [33]
Миндалевидное тело
- AMY получает входные данные от VTA и выводит их в NAc. Миндалевидное тело играет важную роль в создании мощных эмоциональных воспоминаний и, вероятно, лежит в основе создания сильных воспоминаний, связанных с сигналами. [34] Это также важно для смягчения тревожных последствий абстиненции и увеличения потребления наркотиков при зависимости. [35]
Центры удовольствия
[ редактировать ]Удовольствие является компонентом вознаграждения, но не все вознаграждения доставляют удовольствие (например, деньги не вызывают удовольствия, если эта реакция не обусловлена). [2] Стимулы, которые доставляют удовольствие по природе и, следовательно, привлекательны, известны как внутренние вознаграждения , тогда как стимулы, которые привлекательны и мотивируют поведение, но не доставляют удовольствия по своей сути, называются внешними вознаграждениями . [2] Внешние вознаграждения (например, деньги) вознаграждаются в результате усвоенной ассоциации с внутренним вознаграждением. [2] Другими словами, внешние вознаграждения действуют как мотивационные магниты, которые вызывают реакции «желания», но не «нравятся» после их приобретения. [2]
Система вознаграждения содержит центры удовольствия или гедонистические горячие точки – то есть структуры мозга, которые опосредуют реакции удовольствия или «приязни» от внутренних вознаграждений. По состоянию на октябрь 2017 г. [update] Гедонистические горячие точки были идентифицированы в подотделах оболочки прилежащего ядра , вентральном паллидуме , парабрахиальном ядре , орбитофронтальной коре (ОФК) и островковой коре . [3] [14] [36] Горячая точка внутри оболочки прилежащего ядра расположена в ростродорсальном квадранте медиальной оболочки, тогда как гедоническая холодная точка расположена в более задней области. Задний вентральный паллидум также содержит гедоническую горячую точку, а передний вентральный паллидум содержит гедонистическую холодную точку. У крыс микроинъекции опиоидов , эндоканнабиноидов и орексина способны усиливать реакции симпатии в этих «горячих точках». [3] Было продемонстрировано, что гедонистические горячие точки, расположенные в передней островковой и задней островковой части, реагируют на орексин и опиоиды у крыс, как и перекрывающаяся гедоническая холодная точка в передней островковой и задней островковой доле. [36] С другой стороны, было продемонстрировано, что горячая точка парабрахиального ядра реагирует только на агонисты бензодиазепиновых рецепторов. [3]
Гедонистические «горячие точки» функционально связаны: активация одной «горячей точки» приводит к привлечению других, о чем свидетельствует индуцированная экспрессия c -Fos , непосредственно раннего гена . Более того, подавление одной горячей точки приводит к притуплению эффектов активации другой горячей точки. [3] [36] Таким образом, считается, что одновременная активация всех гедонистических точек системы вознаграждения необходима для создания ощущения сильной эйфории . [37]
Желание и симпатия
[ редактировать ]
Выраженность стимула - это атрибут «желания» или «желания», который включает мотивационный компонент, который присваивается стимулу вознаграждения оболочкой прилежащего ядра (оболочка NAcc). [2] [38] [39] Степень нейротрансмиссии дофамина в оболочку NAcc по мезолимбическому пути тесно коррелирует с величиной значимости стимулов для вознаграждения. [38]
Активация дорсо-ростральной области прилежащего ядра коррелирует с увеличением желания без одновременного увеличения симпатии. [40] Однако дофаминергическая нейротрансмиссия в оболочку прилежащего ядра отвечает не только за аппетитную мотивационную значимость (т. е. стимулирующую значимость) в отношении вознаграждающих стимулов, но также и за аверсивную мотивационную значимость, которая направляет поведение в сторону от нежелательных стимулов. [9] [41] [42] В дорсальном полосатом теле активация MSN, экспрессирующих D1, вызывает выраженность аппетитного стимула, тогда как активация MSN, экспрессирующих D2, вызывает отвращение. В NAcc такая дихотомия не столь очевидна, и активация как D1, так и D2 MSN достаточна для повышения мотивации. [43] [44] вероятно, за счет растормаживания ВТА за счет ингибирования вентрального паллидума. [45] [46]
Теория стимулирования-сенсибилизации Робинсона и Берриджа 1993 года предположила, что вознаграждение содержит отдельные психологические компоненты: желание (стимул) и симпатия (удовольствие). Чтобы объяснить усиление контакта с определенным стимулом, например шоколадом, действуют два независимых фактора – наше желание иметь шоколад (желание) и эффект удовольствия от шоколада (нравится). По мнению Робинсона и Берриджа, желание и симпатия — это два аспекта одного и того же процесса, поэтому вознаграждения обычно желательны и нравятся в одинаковой степени. Однако желания и симпатии также меняются независимо при определенных обстоятельствах. Например, крысы, которые не едят после получения дофамина (испытывая потерю желания есть), ведут себя так, как будто им все еще нравится еда. В другом примере активированные электроды самостимуляции в латеральном гипоталамусе крыс повышают аппетит, но также вызывают более неблагоприятные реакции на такие вкусы, как сахар и соль; по-видимому, стимуляция усиливает желание, но не симпатию. Такие результаты показывают, что система вознаграждения крыс включает независимые процессы желания и симпатии. Считается, что компонент желания контролируется дофаминергических путей , тогда как считается, что компонент симпатии контролируется системами опиат-ГАМК-эндоканнабиноиды. [5]
Система антивознаграждения
[ редактировать ]Кубс и Ле Моал предположили, что существует отдельная цепь, ответственная за ослабление поведения, направленного на получение вознаграждения, которую они назвали цепью анти-награды. Этот компонент действует как тормоз в цепи вознаграждения, тем самым предотвращая чрезмерную погоню за едой, сексом и т. д. В эту цепь входят несколько частей миндалевидного тела (ядро ложа терминальной полоски, центральное ядро), прилежащее ядро и сигнальное ядро. молекулы, включая норадреналин, кортикотропин-высвобождающий фактор и динорфин. [47] Предполагается также, что этот контур опосредует неприятные компоненты стресса и, таким образом, считается, что он участвует в возникновении зависимости и абстиненции. В то время как цепь вознаграждения опосредует первоначальное положительное подкрепление, участвующее в развитии зависимости, именно схема анти-награды позже доминирует через отрицательное подкрепление, которое мотивирует стремление к поощрительным стимулам. [48]
Обучение
[ редактировать ]Поощрительные стимулы могут стимулировать обучение как в форме классического обусловливания (павловское обусловливание), так и в форме оперантного обусловливания (инструментальное обусловливание) . В классическом обусловливании вознаграждение может действовать как безусловный стимул, который, когда он связан с условным стимулом, заставляет условный стимул вызывать как скелетно-мышечные (в форме простого подхода и поведения избегания), так и вегетативные реакции. При оперантном обусловливании вознаграждение может действовать как подкрепление , поскольку оно усиливает или поддерживает действия, которые ведут к нему самому. [1] Приобретенное поведение может быть чувствительным, а может и не быть чувствительным к ценности результатов, к которым оно приводит; поведение, которое чувствительно к непредвиденным обстоятельствам результата выполнения действия, а также к ценности результата, является целенаправленным , в то время как вызываемые действия, которые нечувствительны к непредвиденным обстоятельствам или ценности, называются привычками . [49] Считается, что это различие отражает две формы обучения: свободное от моделей и основанное на моделях. Бесплатное обучение модели предполагает простое кэширование и обновление значений. Напротив, обучение на основе моделей предполагает хранение и построение внутренней модели событий, которая позволяет делать выводы и гибко прогнозировать. Хотя павловское обусловливание обычно считается свободным от моделей, значимость стимула, приписываемая условному стимулу, является гибкой по отношению к изменениям во внутренних мотивационных состояниях. [50]
Различные нейронные системы отвечают за учебные связи между стимулами и результатами, действиями и результатами, а также стимулами и реакциями. Хотя классическое обусловливание не ограничивается системой вознаграждения, для улучшения инструментальной производительности с помощью стимулов (т. е. инструментальной передачи по Павлову ) требуется прилежащее ядро. Привычное и целенаправленное инструментальное обучение зависит от латерального . и медиального полосатого тела соответственно [49]
Во время инструментального обучения противоположные изменения в соотношении AMPA и NMDA- рецепторов и фосфорилированной ERK происходят в MSN D1 - типа и D2 - типа, которые составляют прямой и непрямой пути соответственно. [51] [52] Эти изменения в синаптической пластичности и сопутствующее обучение зависят от активации стриарных рецепторов D1 и NMDA. Внутриклеточный каскад, активируемый рецепторами D1, включает вовлечение протеинкиназы А и, в результате фосфорилирования DARPP-32 , ингибирование фосфатаз, которые деактивируют ERK. Рецепторы NMDA активируют ERK через другой, но взаимосвязанный путь Ras-Raf-MEK-ERK . Сама по себе активация ERK, опосредованная NMDA, является самоограничивающейся, поскольку активация NMDA также ингибирует опосредованное PKA ингибирование фосфатаз, деактивирующих ERK. Однако когда каскады D1 и NMDA совместно активируются, они работают синергетически, и результирующая активация ERK регулирует синаптическую пластичность в форме реструктуризации позвоночника, транспорта рецепторов AMPA, регуляции CREB и повышения клеточной возбудимости посредством ингибирования Kv4.2. [53] [54] [55]
расстройства
[ редактировать ]Зависимость
[ редактировать ]ΔFosB (DeltaFosB) – фактор транскрипции гена – сверхэкспрессия в D1-типа средних шипистых нейронах является прилежащего ядра важнейшим общим фактором практически всех форм зависимости (т. е. поведенческих зависимостей и наркозависимости ), который индуцирует связанное с зависимостью поведение и нейронная пластичность . [56] [57] [58] [59] В частности, ΔFosB способствует самостоятельному приему , сенсибилизации вознаграждения и эффектам перекрестной сенсибилизации между конкретными наркотиками, вызывающими привыкание, и поведением. [56] [57] [58] [60] [61] Известно также, что определенные эпигенетические модификации хвостов гистоновых белков (т.е. модификации гистонов) в определенных областях мозга играют решающую роль в молекулярной основе зависимостей . [59] [62] [63] [64]
Наркотики и поведение, вызывающие привыкание , вознаграждают и подкрепляют (т. е. вызывают привыкание ) из-за их воздействия на путь вознаграждения дофамина . [13] [65]
и Латеральный гипоталамус медиальный пучок переднего мозга были наиболее часто изучаемым местом вознаграждения за стимуляцию мозга, особенно в исследованиях влияния лекарств на вознаграждение за стимуляцию мозга. [66] Нейромедиаторной системой, которая наиболее четко идентифицируется как вызывающее привыкание действие наркотических средств, является мезолимбическая дофаминовая система с ее эфферентными мишенями в прилежащем ядре и локальными ГАМКергическими афферентами . Действие амфетамина и кокаина, связанное с вознаграждением, осуществляется в дофаминергических синапсах прилежащего ядра и, возможно, в медиальной префронтальной коре . Крысы также учатся нажимать на рычаг для инъекции кокаина в медиальную префронтальную кору, которая работает за счет увеличения оборота дофамина в прилежащем ядре. [67] [68] Никотин, введенный непосредственно в прилежащее ядро, также усиливает местное высвобождение дофамина, предположительно за счет пресинаптического воздействия на дофаминергические окончания этой области. Никотиновые рецепторы локализуются в телах дофаминергических клеток, а местные инъекции никотина усиливают активацию дофаминергических клеток, что имеет решающее значение для никотинового вознаграждения. [69] [70] снижать активность средних шипистых нейронов Некоторые дополнительные лекарства, вызывающие привыкание, также могут , как следствие, , несмотря на активацию дофаминергических проекций. Для опиатов участок с самым низким порогом воздействия эффектов вознаграждения включает воздействие на ГАМКергические нейроны в вентральной покрышке , вторичный участок вознаграждающих опиатов воздействий на средние шипистые выходные нейроны прилежащего ядра. Таким образом, ядро ныне охарактеризованной схемы вознаграждения за наркотики составляют следующие элементы: ГАМКергические афференты к мезолимбическим дофаминовым нейронам (первичный субстрат опиатного вознаграждения), сами мезолимбические дофаминовые нейроны (первичный субстрат психомоторного стимулирующего вознаграждения) и ГАМКергические эфференты к мезолимбическим дофаминовым нейронам (вторичный участок опиатного вознаграждения). [66]
Мотивация
[ редактировать ]Дисфункциональная мотивационная значимость проявляется при ряде психиатрических симптомов и расстройств. Ангедония , традиционно определяемая как сниженная способность испытывать удовольствие, была повторно рассмотрена как отражающая притупление выраженности стимулов, поскольку большинство ангедонических популяций демонстрируют неповрежденную «симпатию». [71] [72] На другом конце спектра повышенная выраженность стимулов, суженная для конкретных стимулов, характерна для поведенческих и наркотических зависимостей. В случае страха или паранойи дисфункция может заключаться в повышенном отвращении . [73] В современной литературе ангедония связана с предлагаемыми двумя формами удовольствия: «предвосхищающим» и «завершающим».
Нейровизуализационные исследования при диагнозах, связанных с ангедонией, показали снижение активности OFC и вентрального полосатого тела. [74] В одном метаанализе сообщалось, что ангедония связана со снижением нейрональной реакции на ожидание вознаграждения в хвостатом ядре, скорлупе, прилежащем ядре и медиальной префронтальной коре (мПФК). [75]
Расстройства настроения
[ редактировать ]Определенные типы депрессии связаны со снижением мотивации, оцениваемой по готовности прилагать усилия ради вознаграждения. Эти нарушения предположительно связаны со снижением активности в областях полосатого тела, и хотя предполагается, что дофаминергические нарушения играют определенную роль, большинство исследований, изучающих функцию дофамина при депрессии, сообщили о противоречивых результатах. [76] [77] Хотя посмертные и нейровизуализационные исследования выявили отклонения во многих областях системы вознаграждения, лишь немногие результаты последовательно повторяются. В некоторых исследованиях сообщалось о снижении активности NAcc, гиппокампа, медиальной префронтальной коры (mPFC) и орбитофронтальной коры (OFC), а также о повышении активности базолатеральной миндалины и субгенуальной поясной извилины (sgACC) во время задач, связанных с вознаграждением или положительными стимулами. Эти нейровизуализационные аномалии дополняются небольшими посмертными исследованиями, но те небольшие исследования, которые были проведены, позволяют предположить снижение возбуждающих синапсов в mPFC. [78] Снижение активности mPFC во время задач, связанных с вознаграждением, по-видимому, локализуется в более дорсальных областях (т.е. в прегенуальной поясной извилине ), в то время как более вентральная sgACC гиперактивна при депрессии. [79]
Попытки исследовать основные нейронные схемы на животных моделях также дали противоречивые результаты. Для моделирования депрессии, хронического социального поражения (CSDS) и хронического легкого стресса (CMS) обычно используются две парадигмы, хотя их существует много. CSDS вызывает снижение предпочтения сахарозы, снижение социальных взаимодействий и повышенную неподвижность в тесте принудительного плавания. CMS аналогичным образом снижает предпочтение сахарозы и поведенческое отчаяние, что оценивалось с помощью тестов с подвешиванием за хвост и принудительного плавания. Животные, восприимчивые к CSDS, демонстрируют повышенную фазовую активацию VTA, а ингибирование проекций VTA-NAcc ослабляет поведенческие дефициты, вызванные CSDS. [80] Однако ингибирование проекций VTA- mPFC усугубляет социальную изоляцию. С другой стороны, связанное с CMS снижение предпочтения сахарозы и неподвижности ослаблялось и усугублялось возбуждением и торможением VTA соответственно. [81] [82] Хотя эти различия могут быть связаны с разными протоколами стимуляции или плохими парадигмами трансляции, переменные результаты могут также заключаться в гетерогенной функциональности областей, связанных с вознаграждением. [83]
Оптогенетическая стимуляция mPFC в целом оказывает антидепрессивное действие. Этот эффект, по-видимому, локализован в гомологе pgACC у грызунов (прелимбическая кора), поскольку стимуляция гомолога sgACC у грызунов (инфралимбическая кора) не вызывает поведенческих эффектов. Более того, глубокая стимуляция инфралимбической коры головного мозга, которая, как считается, оказывает тормозящее действие, также оказывает антидепрессивное действие. Этот вывод согласуется с наблюдением, что фармакологическое торможение инфралимбической коры ослабляет депрессивное поведение. [83]
Шизофрения
[ редактировать ]Шизофрения связана с дефицитом мотивации, который обычно группируется под другими негативными симптомами, такими как снижение спонтанной речи . Часто сообщается, что переживание «нравится» не повреждено. [84] как на поведенческом, так и на нейронном уровне, хотя результаты могут быть специфичными для определенных стимулов, таких как денежное вознаграждение. [85] Кроме того, при шизофрении не нарушены неявное обучение и простые задачи, связанные с вознаграждением. [86] Скорее, недостатки в системе вознаграждения очевидны при выполнении задач, связанных с вознаграждением, которые являются когнитивно сложными. Эти дефициты связаны как с аномальной активностью полосатого тела, так и с активностью OFC, а также с аномалиями в областях, связанных с когнитивными функциями, таких как дорсолатеральная префронтальная кора (DLPFC). [87]
Синдром дефицита внимания с гиперактивностью
[ редактировать ]У людей с СДВГ основные аспекты системы вознаграждения недостаточно активны, что затрудняет получение вознаграждения от регулярных занятий. Люди с этим расстройством испытывают повышение мотивации после того, как поведение с высокой стимуляцией вызывает выброс дофамина. После этого повышения и вознаграждения возвращение к исходному уровню приводит к немедленному падению мотивации. [88]
Считается, что нарушения дофаминергической и серотонинергической функции являются ключевыми факторами СДВГ. [89] Эти нарушения могут привести к исполнительной дисфункции, такой как нарушение регуляции обработки вознаграждения и мотивационной дисфункции, включая ангедонию. [90]
История
[ редактировать ]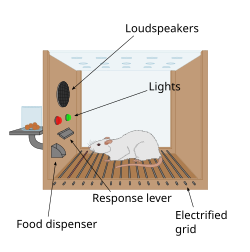
Первым ключом к наличию системы вознаграждения в мозгу стало случайное открытие, сделанное Джеймсом Олдсом и Питером Милнером в 1954 году. Они обнаружили, что крысы могут выполнять такие действия, как нажатие на штангу, чтобы вызвать короткую вспышку электрической стимуляции определенных клеток. сайты в их мозгу. Это явление называется внутричерепной самостимуляцией или вознаграждением за стимуляцию мозга . Обычно крысы нажимают на рычаг сотни или тысячи раз в час, чтобы получить стимуляцию мозга, останавливаясь только тогда, когда они утомлены. Пытаясь научить крыс решать проблемы и проходить лабиринты, стимуляция определенных областей мозга, где была обнаружена стимуляция, по-видимому, доставляла животным удовольствие. Они попробовали то же самое с людьми, и результаты были схожими. Объяснение того, почему животные ведут себя так, что это не имеет никакого значения для выживания ни их самих, ни их вида, заключается в том, что стимуляция мозга активирует систему, лежащую в основе вознаграждения. [91]
В ходе фундаментального открытия, сделанного в 1954 году, исследователи Джеймс Олдс и Питер Милнер обнаружили, что низковольтная электрическая стимуляция определенных областей мозга крыс действует как награда за обучение животных проходить лабиринты и решать задачи. [92] [ не удалось пройти проверку ] [93] Казалось, что стимуляция этих участков мозга доставляла животным удовольствие. [92] а в более поздних работах люди сообщили о приятных ощущениях от такой стимуляции. [ нужна ссылка ] Когда крыс тестировали в ящиках Скиннера , где они могли стимулировать систему вознаграждения нажатием на рычаг, крысы давили часами. [93] Исследования следующих двух десятилетий установили, что дофамин является одним из основных химических веществ, способствующих передаче нервных сигналов в этих регионах, и было высказано предположение, что дофамин является «химическим веществом удовольствия» мозга. [94]
Иван Павлов был психологом, который использовал систему вознаграждения для изучения классической обусловленности . Павлов использовал систему поощрений, награждая собак едой после того, как они услышали звонок или другой стимул. Павлов награждал собак так, чтобы собаки ассоциировали еду, награду, со звонком, стимулом. [95] Эдвард Л. Торндайк использовал систему вознаграждения для изучения оперантного обусловливания. Он начал с того, что поместил кошек в коробку-пазл и разместил еду снаружи коробки, чтобы кошка хотела убежать. Кошки старались выбраться из коробки-головоломки и добраться до еды. Хотя кошки съели еду после того, как сбежали из коробки, Торндайк узнал, что кошки пытались сбежать из коробки, не получив в награду еды. Торндайк использовал вознаграждение в виде еды и свободы, чтобы стимулировать систему вознаграждения кошек. Торндайк использовал это, чтобы увидеть, как кошки научились выбираться из коробки. [96] Совсем недавно Иван Де Араужо и его коллеги использовали питательные вещества внутри кишечника, чтобы стимулировать систему вознаграждения через блуждающий нерв. [97]
Другие виды
[ редактировать ]Животные быстро учатся нажимать на штангу, чтобы получить инъекцию опиатов непосредственно в покрышку среднего мозга или прилежащее ядро . Те же животные не работают на получение опиатов, если дофаминергические нейроны мезолимбического пути инактивированы . С этой точки зрения животные, как и люди, ведут себя так, что увеличивают высвобождение дофамина.
Кент Берридж , исследователь в области аффективной нейробиологии , обнаружил, что сладкий ( нравится ) и горький ( не нравится ) вкусы вызывают различные орофациальные выражения , и эти выражения аналогичным образом проявлялись у новорожденных людей, орангутанов и крыс. Это свидетельствовало о том, что удовольствие (в частности, симпатия ) имеет объективные особенности и по сути одинаково у разных видов животных. Большинство нейробиологических исследований показали, что чем больше дофамина высвобождается в результате вознаграждения, тем эффективнее вознаграждение. Это называется гедонистическим воздействием, которое можно изменить посредством усилий ради вознаграждения и самой награды. Берридж обнаружил, что блокирование дофаминовой системы, похоже, не меняет положительную реакцию на что-то сладкое (судя по выражению лица). Другими словами, гедонистическое воздействие не менялось в зависимости от количества сахара. Это опровергло традиционное предположение о том, что дофамин обеспечивает удовольствие. Даже при более интенсивных изменениях дофамина данные, казалось, оставались постоянными. [98] Тем не менее, клиническое исследование, проведенное в январе 2019 года, в котором оценивалось влияние предшественника дофамина ( леводопы ), антагониста ( рисперидона ) и плацебо на реакцию вознаграждения на музыку, включая степень удовольствия, испытываемого во время музыкального озноба , измеренного по изменениям электродермального активность , а также субъективные оценки — обнаружено, что манипулирование нейротрансмиссией дофамина двунаправленно регулирует познание удовольствия (в частности, гедоническое воздействие музыки ) у людей. [99] [100] Это исследование показало, что повышенная нейротрансмиссия дофамина действует как обязательное условие приятных гедонистических реакций на музыку у людей. [99] [100]
Берридж разработал гипотезу значимости стимулов , чтобы решить проблему недостатка вознаграждений. Это объясняет компульсивное употребление наркотиков наркоманами, даже когда наркотик больше не вызывает эйфории, и тягу к ним, возникающую даже после того, как человек закончил абстиненцию. Некоторые наркоманы реагируют на определенные стимулы, связанные с нервными изменениями, вызванными наркотиками. Эта сенсибилизация в мозге аналогична эффекту дофамина, поскольку возникают реакции желания и симпатии . Мозг и поведение человека и животных претерпевают схожие изменения в отношении систем вознаграждения, поскольку эти системы очень заметны. [98]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я Шульц В. (2015). «Нейронное вознаграждение и сигналы решения: от теорий к данным» . Физиологические обзоры . 95 (3): 853–951. doi : 10.1152/physrev.00023.2014 . ПМЦ 4491543 . ПМИД 26109341 .
Награды в оперантном обусловливании являются положительным подкреплением. ... Оперантное поведение дает хорошее определение вознаграждения. Все, что заставляет человека возвращаться и требовать большего, является положительным подкреплением и, следовательно, наградой. Хотя положительное подкрепление дает хорошее определение, оно является лишь одной из нескольких функций вознаграждения. ... Награды привлекательны. Они мотивируют и заставляют нас прилагать усилия. ... Награды вызывают поведение приближения, также называемое аппетитным или подготовительным поведением, сексуальным поведением и конверсионным поведением. ... Таким образом, любой стимул, объект, событие, деятельность или ситуация, которая потенциально может заставить нас приблизиться и поглотить его, по определению является наградой. ... Поощрительные стимулы, объекты, события, ситуации и действия состоят из нескольких основных компонентов. Во-первых, награды имеют основные сенсорные компоненты (зрительные, слуховые, соматосенсорные, вкусовые и обонятельные). Во-вторых, награды заметны и, таким образом, вызывают внимание, которое проявляется в виде ориентировочных реакций. Выраженность вознаграждений обусловлена тремя основными факторами, а именно их физической интенсивностью и воздействием (физическая значимость), их новизной и неожиданностью (заметность новизны/сюрприза) и их общим мотивационным воздействием, разделяемым с карателями (мотивационная значимость). Отдельная форма, не включенная в эту схему, — побудительная значимость — касается в первую очередь дофаминовой функции при аддикции и относится только к подходу к поведению (в отличие от обучения). не содержится в сенсорных компонентах и компонентах внимания и не объясняется ими. Этот компонент отражает поведенческие предпочтения и поэтому является субъективным и лишь частично определяется физическими параметрами. Только этот компонент и представляет собой то, что мы понимаем под наградой. Он опосредует специфическое поведенческое подкрепление, создание подходов и эмоциональные эффекты вознаграждений, которые имеют решающее значение для выживания и воспроизводства организма, тогда как все остальные компоненты лишь поддерживают эти функции. ... Награды также могут быть неотъемлемой частью поведения. Они контрастируют с внешними вознаграждениями, которые обеспечивают мотивацию поведения и составляют суть оперантного поведения в лабораторных тестах. Внутренние вознаграждения — это действия, которые доставляют удовольствие сами по себе и предпринимаются ради самих себя, не являясь средством получения внешних вознаграждений. ... Внутренние награды сами по себе являются настоящими наградами, поскольку они побуждают к обучению, подходу и удовольствию, например, к совершенствованию, игре на фортепиано и получению удовольствия. Хотя они могут служить для обусловления вознаграждений более высокого порядка, они не являются обусловленными вознаграждениями более высокого порядка, поскольку для достижения их свойств вознаграждения не требуется соединение с безусловным вознаграждением. ... Эти эмоции также называются симпатией (к удовольствию) и желанием (к желанию) в исследованиях зависимостей и решительно поддерживают функции вознаграждения, генерирующие обучение и подход.
- ^ Jump up to: а б с д и ж г час я дж к л м Шульц, Вольфрам (июль 2015 г.). «Нейрональное вознаграждение и сигналы решения: от теорий к данным» . Физиологические обзоры . 95 (3): 853–951. doi : 10.1152/physrev.00023.2014 . ПМЦ 4491543 . ПМИД 26109341 .
- ^ Jump up to: а б с д и ж г час Берридж К.К., Крингельбах М.Л. (май 2015 г.). «Системы удовольствия в мозге» . Нейрон . 86 (3): 646–664. дои : 10.1016/j.neuron.2015.02.018 . ПМЦ 4425246 . ПМИД 25950633 .
Последние данные указывают на то, что в префронтальной коре [орбитофронтальная кора] OFC и островковая кора могут содержать свои собственные дополнительные горячие точки (DC Castro et al., Soc. Neurosci., реферат). В определенных субрегионах каждой области микроинъекции, стимулирующие опиоиды или стимулирующие орексин, увеличивают количество реакций симпатии , вызываемых сладким, подобно «горячим точкам» [прилежащего ядра] NAc и [вентрального паллидума] VP. Успешное подтверждение гедонистических горячих точек в OFC или островковой части было бы важным и, возможно, актуальным для орбитофронтального среднепереднего участка, упомянутого ранее, который особенно отслеживает субъективное удовольствие от еды у людей (Georgiadis et al., 2012; Kringelbach, 2005; Kringelbach et al., 2012; Kringelbach, 2005; Kringelbach et др., 2003; Смолл и др., 2001; Наконец, в стволе мозга участок заднего мозга рядом с парабрахиальным ядром дорсального моста также, по-видимому, способен способствовать гедоническому улучшению функций (Söderpalm and Berridge, 2000). Стволовой механизм получения удовольствия может показаться более удивительным, чем «горячие точки» переднего мозга, для любого, кто рассматривает ствол мозга как просто рефлекторный механизм, но парабрахиальное ядро моста способствует вкусовым, болевым и многим висцеральным ощущениям тела, а также, как предполагается, играет важную роль. роль в мотивации (Wu et al., 2012) и человеческих эмоциях (особенно в связи с гипотезой соматических маркеров) (Damasio, 2010).
- ^ Го, Ронг; Бёмер, Венделин; Хебарт, Мартин; Чиен, Самсон; Соммер, Тобиас; Обермайер, Клаус; Глешер, январь (14 декабря 2016 г.). «Взаимодействие инструментального и целенаправленного обучения модулирует представления ошибок прогнозирования в вентральном полосатом теле» . Журнал неврологии . 36 (50). Общество нейронаук: 12650–12660. doi : 10.1523/jneurosci.1677-16.2016 . ISSN 0270-6474 . ПМК 6705659 . ПМИД 27974615 .
- ^ Jump up to: а б с д и Колб, Брайан; Уишоу, Ян К. (2001). Введение в мозг и поведение (1-е изд.). Нью-Йорк: Стоит. стр. 438–441 . ISBN 9780716751694 .
- ^ Фонд исследований мозга и поведения (13 марта 2019 г.). «Биология зависимости» . Ютуб .
- ^ «Дофамин, участвующий в агрессии» . Медицинские новости сегодня . 15 января 2008 г. Архивировано из оригинала 23 сентября 2010 г. . Проверено 14 ноября 2010 г.
- ^ Дуарте, Изабель К.; Афонсу, Соня; Хорхе, Хелена; Кайолла, Рикардо; Феррейра, Карлос; Кастело-Бранко, Мигель (1 мая 2017 г.). «Племенная любовь: нейронные корреляты страстной вовлеченности футбольных фанатов» . Социальная когнитивная и аффективная нейронаука . 12 (5): 718–728. doi : 10.1093/scan/nsx003 . ПМК 5460049 . ПМИД 28338882 .
- ^ Jump up to: а б с Саламоне, Джон Д.; Корреа, Мерсе (ноябрь 2012 г.). «Таинственные мотивационные функции мезолимбического дофамина» . Нейрон . 76 (3): 470–485. дои : 10.1016/j.neuron.2012.10.021 . ПМК 4450094 . ПМИД 23141060 .
- ^ Jump up to: а б с д и ж г Ягер Л.М., Гарсия А.Ф., Вунш А.М., Фергюсон С.М. (август 2015 г.). «Все тонкости полосатого тела: роль в наркозависимости» . Нейронаука . 301 : 529–541. doi : 10.1016/j.neuroscience.2015.06.033 . ПМЦ 4523218 . ПМИД 26116518 .
[Стриатум] получает дофаминергические сигналы из вентральной покрышки (VTA) и черной субстанции (SNr), а также глутаматергические сигналы из нескольких областей, включая кору, гиппокамп, миндалевидное тело и таламус (Swanson, 1982; Phillipson and Griffiths, 1985; Финч, 1996; Гроеневеген и др., 1999; Бритт и др., 2012). Эти глутаматергические входы вступают в контакт с головками дендритных отростков стриарных ГАМКергических средних шипиковых проекционных нейронов (MSN), тогда как дофаминергические входы синапсируются на шейке позвоночника, обеспечивая важное и сложное взаимодействие между этими двумя входами при модуляции активности MSN... Следует также отметить, что существует небольшая популяция нейронов в [прилежащем ядре] NAc, которые коэкспрессируют как рецепторы D1, так и D2, хотя это в значительной степени ограничено оболочкой NAc (Bertran-Gonzalez et al., 2008). ... Нейроны в подразделениях ядра NAc и оболочки NAc также различаются функционально. Ядро NAc участвует в обработке условных раздражителей, тогда как оболочка NAc более важна в обработке безусловных раздражителей; Классически считается, что эти две популяции полосатого MSN оказывают противоположные эффекты на продукцию базальных ганглиев. Активация dMSN вызывает чистое возбуждение таламуса, что приводит к возникновению положительной кортикальной обратной связи; тем самым действуя как сигнал «инициировать» поведение. Однако активация iMSN вызывает общее торможение таламической активности, что приводит к образованию отрицательной кортикальной обратной связи и, следовательно, служит «тормозом» для подавления поведения... появляется также все больше свидетельств того, что iMSN играют роль в мотивации и зависимости ( Лобо и Нестлер, 2011; Грютер и др., 2013). Например, оптогенетическая активация iMSN ядра и оболочки NAc подавляла развитие кокаинового CPP, тогда как селективная абляция iMSN ядра и оболочки NAc... усиливала развитие и устойчивость амфетаминового CPP (Durieux et al., 2009; Lobo et др., 2010). Эти результаты показывают, что iMSN могут двунаправленно модулировать вознаграждение за прием лекарств. ... В совокупности эти данные позволяют предположить, что iMSN обычно сдерживают поведение, связанное с употреблением наркотиков, и рекрутирование этих нейронов на самом деле может защищать от развития компульсивного употребления наркотиков.
- ^ Тейлор С.Б., Льюис С.Р., Олив М.Ф. (2013). «Нейросхема незаконной психостимулирующей зависимости: острые и хронические последствия у человека» . Реабилитация после злоупотребления наркотиками . 4 : 29–43. дои : 10.2147/SAR.S39684 . ПМЦ 3931688 . ПМИД 24648786 .
Области базальных ганглиев, включающие дорсальное и вентральное полосатое тело, внутренние и наружные сегменты бледного шара, субталамическое ядро и тела дофаминергических клеток черной субстанции, активно участвуют не только в регуляции мелкой моторики, но и в префронтальной коре. ] Функция PFC.43 Из этих областей на предмет зависимости чаще всего исследуются [прилежащее ядро] NAc (описанное выше) и [дорсальное полосатое тело] DS (описанное ниже). Таким образом, здесь будет упомянуто лишь краткое описание модулирующей роли базальных ганглиев в цепях, связанных с зависимостью. Общий вывод базальных ганглиев осуществляется преимущественно через таламус, который затем возвращается в префронтальную кору, образуя кортико-стриарно-таламо-кортикальные (CSTC) петли. Предлагаются три петли CSTC для модуляции исполнительной функции, выбора действия и поведенческого торможения. В дорсолатеральном префронтальном контуре базальные ганглии в первую очередь модулируют идентификацию и выбор целей, включая вознаграждение.44 Схема OFC [орбитофронтальной коры] модулирует принятие решений и импульсивность, а передняя поясная извилина модулирует оценку последствий.44 Эти цепи модулируются дофаминергическими входами из [вентральной покрышки] VTA, чтобы в конечном итоге направлять поведение, связанное с зависимостью, включая сохранение и сужение поведенческого репертуара в сторону поиска наркотиков и продолжения употребления наркотиков, несмотря на негативные последствия.43–45
- ^ Гралль-Броннек М., Соваже А (2014). «Использование повторяющейся транскраниальной магнитной стимуляции для модуляции тяги и аддиктивного поведения: критический обзор литературы по эффективности, техническим и методологическим соображениям» . Неврология. Биоповедение. Преподобный . 47 : 592–613. doi : 10.1016/j.neubiorev.2014.10.013 . ПМИД 25454360 .
Исследования показали, что тяга подкрепляется активацией цепей вознаграждения и мотивации (McBride et al., 2006, Wang et al., 2007, Wing et al., 2012, Goldman et al., 2013, Jansen et al., 2013). и Волков и др., 2013). По мнению этих авторов, основными задействованными нервными структурами являются: прилежащее ядро, дорсальное полосатое тело, орбитофронтальная кора, передняя поясная извилина, дорсолатеральная префронтальная кора (ДЛПФК), миндалевидное тело, гиппокамп и островковая доля.
- ^ Jump up to: а б Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). Сидор А., Браун Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 365–366, 376. ISBN. 978-0-07-148127-4 .
Нейронные субстраты, лежащие в основе восприятия вознаграждения и феномена положительного подкрепления, представляют собой набор взаимосвязанных структур переднего мозга, называемых путями вознаграждения мозга; к ним относятся прилежащее ядро (NAc; основной компонент вентрального полосатого тела), базальный отдел переднего мозга (компоненты которого получили название расширенной миндалины, как обсуждается далее в этой главе), гиппокамп, гипоталамус и лобные области коры головного мозга. Эти структуры получают богатую дофаминергическую иннервацию из вентральной покрышки (ВТА) среднего мозга. Наркотики, вызывающие привыкание, вознаграждают и подкрепляют, потому что они действуют на пути вознаграждения мозга, усиливая либо высвобождение дофамина, либо эффекты дофамина в NAc или родственных структурах, либо потому, что они производят эффекты, аналогичные дофамину. ... Макроструктура, которая, как предполагается, объединяет многие функции этой цепи, описывается некоторыми исследователями как расширенная миндалина. Считается, что расширенная миндалина состоит из нескольких базальных структур переднего мозга, которые имеют схожую морфологию, иммуноцитохимические особенности и связи и хорошо подходят для опосредования аспектов функции вознаграждения; к ним относятся ядро ложа терминальной полоски, центральная медиальная миндалина, оболочка NAc и сублентикулярная безымянная субстанция.
- ^ Jump up to: а б Ричард Дж.М., Кастро, округ Колумбия, Дифеликантонио АГ, Робинсон М.Дж., Берридж К.К. (ноябрь 2013 г.). «Картирование мозговых цепей вознаграждения и мотивации: по стопам Энн Келли» . Неврология. Биоповедение. Преподобный . 37 (9 ч. А): 1919–1931. doi : 10.1016/j.neubiorev.2012.12.008 . ПМК 3706488 . ПМИД 23261404 .
Рисунок 3: Нейронные цепи, лежащие в основе мотивированного «желания» и гедонического «нравится». - ^ Ло М., Чжоу Дж., Лю З. (август 2015 г.). «Обработка вознаграждения ядром дорсального шва: 5-HT и выше» . Учиться. Мем . 22 (9): 452–460. дои : 10.1101/lm.037317.114 . ПМК 4561406 . ПМИД 26286655 .
- ^ Моултон Э.А., Элман И., Бесерра Л.Р., Гольдштейн Р.З., Борсук Д. (май 2014 г.). «Мозжечок и зависимость: выводы, полученные в результате исследований нейровизуализации» . Биология наркомании . 19 (3): 317–331. дои : 10.1111/adb.12101 . ПМК 4031616 . ПМИД 24851284 .
- ^ Калиджоре Д, Пеццуло Г, Бальдассарре Г, Бостан А.С., Стрик П.Л., Дойя К., Хельмих Р.С., Дирккс М., Хоук Дж., Йорнтелл Х., Лаго-Родригес А., Галеа Х.М., Миалл Р.К., Попа Т., Кишоре А., Вершуре П.Ф., Зукка Р., Эррерос I (февраль 2017 г.). «Консенсусный документ: К взгляду на функции мозжечка на системном уровне: взаимодействие между мозжечком, базальными ганглиями и корой» . Мозжечок . 16 (1): 203–229. дои : 10.1007/s12311-016-0763-3 . ПМК 5243918 . ПМИД 26873754 .
- ^ Огава, СК; Ватабе-Учида, М (2018). «Организация системы дофамина и серотонина: Анатомическое и функциональное картирование моносинаптических входов с использованием вируса бешенства». Фармакология Биохимия и поведение . 174 : 9–22. дои : 10.1016/j.pbb.2017.05.001 . ПМИД 28476484 . S2CID 5089422 .
- ^ Моралес, М; Марголис, Е.Б. (февраль 2017 г.). «Вентральная область покрышки: клеточная гетерогенность, связность и поведение». Обзоры природы. Нейронаука . 18 (2): 73–85. дои : 10.1038/nrn.2016.165 . ПМИД 28053327 . S2CID 10311562 .
- ^ Ламмель, С; Лим, БК; Маленка, РЦ (январь 2014 г.). «Награда и отвращение в гетерогенной дофаминовой системе среднего мозга» . Нейрофармакология . 76 Часть Б: 351–9. doi : 10.1016/j.neuropharm.2013.03.019 . ПМЦ 3778102 . ПМИД 23578393 .
- ^ Ние, ЭХ; Ким, С.Ю.; Намбури, П; Тай, К.М. (20 мая 2013 г.). «Оптогенетическое вскрытие нейронных цепей, лежащих в основе эмоциональной валентности и мотивированного поведения» . Исследования мозга . 1511 : 73–92. дои : 10.1016/j.brainres.2012.11.001 . hdl : 1721.1/92890 . ПМК 4099056 . ПМИД 23142759 .
- ^ Трэнтэм-Дэвидсон Х., Нили Л.К., Лавин А., Симанс Дж.К. (2004). «Механизмы, лежащие в основе дифференциальной регуляции торможения дофаминовых рецепторов D1 и D2 в префронтальной коре» . Журнал неврологии . 24 (47): 10652–10659. doi : 10.1523/jneurosci.3179-04.2004 . ПМК 5509068 . ПМИД 15564581 .
- ^ Ю ЗБ, Чен ЮК, Мудрый РА (2001). «Высвобождение дофамина и глутамата в прилежащем ядре и вентральной покрышке крысы после латеральной самостимуляции гипоталамуса». Нейронаука . 107 (4): 629–639. дои : 10.1016/s0306-4522(01)00379-7 . ПМИД 11720786 . S2CID 33615497 .
- ^ Jump up to: а б Кастро, округ Колумбия; Коул, СЛ; Берридж, КЦ (2015). «Роль латерального гипоталамуса, прилежащего ядра и вентрального паллидума в еде и голоде: взаимодействие между гомеостатическими схемами и схемами вознаграждения» . Границы системной нейронауки . 9 : 90. дои : 10.3389/fnsys.2015.00090 . ПМЦ 4466441 . ПМИД 26124708 .
- ^ Карлесон, Вашингтон, младший; Томас, MJ (2009). «Биологические субстраты вознаграждения и отвращения: гипотеза активности прилежащего ядра» . Нейрофармакология . 56 (Приложение 1): 122–32. doi : 10.1016/j.neuropharm.2008.06.075 . ПМЦ 2635333 . ПМИД 18675281 .
- ^ Мудрый Р.А., Ромпре П.П. (1989). «Мозговой дофамин и вознаграждение». Ежегодный обзор психологии . 40 : 191–225. дои : 10.1146/annurev.ps.40.020189.001203 . ПМИД 2648975 .
- ^ Мудрый РА (октябрь 2002 г.). «Схема вознаграждения мозга: выводы из неощутимых стимулов» . Нейрон . 36 (2): 229–240. дои : 10.1016/S0896-6273(02)00965-0 . ПМИД 12383779 . S2CID 16547037 .
- ^ Jump up to: а б Кокане, СС, и Перротти, Л.И. (2020). Половые различия и роль эстрадиола в мезолимбических цепях вознаграждения и уязвимости к кокаиновой и опиатной зависимости. Границы поведенческой нейронауки, 14.
- ^ Беккер, Дж. Б., и Чартофф, Э. (2019). Половые различия в нейронных механизмах, опосредующих вознаграждение и зависимость. Нейропсихофармакология, 44(1), 166-183.
- ^ Стуф, JC, и Кебабиан, JW (1984). Два дофаминовых рецептора: биохимия, физиология и фармакология. Науки о жизни, 35(23), 2281-2296.
- ^ Инь, Х.Х., Ноултон, Б.Дж., и Баллейн, Б.В. (2005). Блокада NMDA-рецепторов в дорсомедиальном полосатом теле предотвращает обучение действию-результату при инструментальном обусловливании. Европейский журнал неврологии, 22(2), 505-512.
- ^ Jump up to: а б с Кооб, Г.Ф., и Волков, Н.Д. (2016). Нейробиология зависимости: анализ нейросхем. The Lancet Psychiatry, 3 (8), 760–773.
- ^ Кутлу, М.Г., и Гулд, Т.Дж. (2016). Влияние злоупотребления наркотиками на пластичность гиппокампа и зависящее от гиппокампа обучение и память: вклад в развитие и поддержание зависимости. Обучение и память, 23 (10), 515–533.
- ^ Макгоф, JL (июль 2004 г.). «Миндалина модулирует консолидацию воспоминаний об эмоционально возбуждающих переживаниях». Ежегодный обзор неврологии. 27 (1): 1–28.
- ^ Кооб Г.Ф., Ле Моал М. (2008). Зависимость и система антивознаграждения мозга. Анну. Преподобный Психол. 59 29–53. 10.1146/annurev.psych.59.103006.093548 [PubMed] [CrossRef] [Google Scholar]Кооб Г.Ф., Санна П.П., Блум Ф.Э. (1998). Нейронаука зависимости. Нейрон 21 467–476.
- ^ Jump up to: а б с Кастро, округ Колумбия; Берридж, Канзас (24 октября 2017 г.). «Горячие точки гедонического гедонизма опиоидов и орексина в орбитофронтальной коре и островке крысы» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (43): Е9125–Е9134. Бибкод : 2017PNAS..114E9125C . дои : 10.1073/pnas.1705753114 . ПМЦ 5664503 . ПМИД 29073109 .
Здесь мы показываем, что стимуляция опиоидами или орексином в орбитофронтальной коре и островковой зоне причинно усиливает гедонистические реакции «приязни» к сладкому и находим третий участок коры, где те же самые нейрохимические стимуляции снижают положительное гедонистическое воздействие.
- ^ Крингельбах М.Л., Берридж К.К. (2012). «Радостный разум» (PDF) . Научный американец . 307 (2): 44–45. Бибкод : 2012SciAm.307b..40K . doi : 10.1038/scientificamerican0812-40 . ПМИД 22844850 . Архивировано из оригинала (PDF) 29 марта 2017 года . Проверено 17 января 2017 г.
Поэтому вполне логично, что настоящие центры удовольствия в мозгу – те, которые непосредственно отвечают за создание приятных ощущений – оказались внутри некоторых структур, ранее идентифицированных как часть схемы вознаграждения. Одна из этих так называемых гедонистических горячих точек находится в субобласти прилежащего ядра, называемой медиальной оболочкой. Второй находится внутри вентрального паллидума, глубоко расположенной структуры у основания переднего мозга, которая получает большую часть сигналов от прилежащего ядра. ...
С другой стороны, сильную эйфорию достичь труднее, чем повседневные удовольствия. Причина может заключаться в том, что сильное усиление удовольствия (например, химически вызванный всплеск удовольствия, который мы получили у лабораторных животных) требует одновременной активации всей сети. Отказ от любого отдельного компонента ослабляет кайф.
Неясно, работает ли схема удовольствия – и в частности вентральный паллидум – у людей таким же образом. - ^ Jump up to: а б Берридж К.К. (апрель 2012 г.). «От ошибки прогнозирования к значимости стимулов: мезолимбические вычисления мотивации вознаграждения» . Евро. Дж. Нейроски . 35 (7): 1124–1143. дои : 10.1111/j.1460-9568.2012.07990.x . ПМК 3325516 . ПМИД 22487042 .
Здесь я обсуждаю, как мезокортиколимбические механизмы генерируют мотивационный компонент значимости стимулов. Стимулирующая значимость рассматривает обучение и память по Павлову как одни из входных данных, а в качестве не менее важных входных данных принимают факторы нейробиологического состояния (например, состояние приема препарата, состояние аппетита, состояние насыщения), которые могут меняться независимо от обучения. Изменения нейробиологического состояния могут вызывать необученные колебания или даже изменение способности ранее усвоенного сигнала вознаграждения вызывать мотивацию. Такие колебания мотивации, запускаемой сигналами, могут резко отклоняться от всех ранее усвоенных значений относительно соответствующего результата вознаграждения. ... Ассоциативное обучение и прогнозирование играют важную роль в мотивации к вознаграждению. Обучение придает стимулирующую ценность произвольным сигналам, таким как условный стимул Павлова (УС), который связан с вознаграждением (безусловный стимул или УКС). Заученные сигналы о вознаграждении часто являются мощными триггерами желаний. Например, заученные сигналы могут вызвать нормальный аппетит у каждого, а иногда могут вызвать компульсивные побуждения и рецидивы у наркоманов.
Вызванное сигналом «желание» для UCS
Кратковременная встреча с CS (или короткая встреча с UCS) часто вызывает импульс повышенной мотивации для получения и использования большего количества вознаграждений UCS. Это характерная черта значимости стимулов.
Сигналы как привлекательные мотивационные магниты
Когда Павловскому CS+ приписывается стимулирующая значимость, это не только вызывает «желание» его UCS, но часто сам сигнал становится очень привлекательным – даже до иррациональной степени. Это привлечение сигнала является еще одной характерной особенностью значимости стимулов... Часто можно увидеть две узнаваемые особенности значимости стимулов, которые можно использовать в нейробиологических экспериментах: (i) «желание», направленное на UCS – запускаемые CS импульсы усиленного «желания» вознаграждение UCS; и (ii) «желание», направленное на CS – мотивированное влечение к сигналу Павлова, которое превращает произвольный стимул CS в мотивационный магнит. - ^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). Сидор А., Браун Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 147–148, 367, 376. ISBN. 978-0-07-148127-4 .
Нейроны VTA DA играют решающую роль в мотивации, поведении, связанном с вознаграждением (глава 15), внимании и различных формах памяти. Такая организация системы DA, широкая проекция ограниченного числа клеточных тел, обеспечивает скоординированные ответы на новые мощные вознаграждения. Таким образом, действуя в различных терминальных полях, дофамин придает мотивационную значимость («желание») самому вознаграждению или связанным с ним сигналам (область оболочки прилежащего ядра), обновляет ценность, придаваемую различным целям в свете этого нового опыта (орбитальная префронтальная кора), помогает консолидировать несколько форм памяти (миндалевидное тело и гиппокамп) и кодирует новые двигательные программы, которые облегчат получение этого вознаграждения в будущем (область ядра прилежащего ядра и дорсальное полосатое тело). В этом примере дофамин модулирует обработку сенсомоторной информации в различных нейронных цепях, чтобы максимизировать способность организма получать будущие вознаграждения. ...
Схема вознаграждения мозга, на которую воздействуют наркотики, вызывающие привыкание, обычно обеспечивает удовольствие и усиление поведения, связанного с естественными подкреплениями, такими как еда, вода и сексуальный контакт. Дофаминовые нейроны в VTA активируются пищей и водой, а высвобождение дофамина в NAc стимулируется присутствием естественных подкреплений, таких как еда, вода или сексуальный партнер. ...
NAc и VTA являются центральными компонентами схемы, лежащей в основе вознаграждения и памяти о вознаграждении. Как упоминалось ранее, активность дофаминергических нейронов в ВТА, по-видимому, связана с предсказанием вознаграждения. NAc участвует в обучении, связанном с подкреплением и модуляцией двигательных ответов на стимулы, удовлетворяющие внутренние гомеостатические потребности. Оболочка NAc, по-видимому, особенно важна для начального действия препарата в схеме вознаграждения; наркотики, вызывающие привыкание, по-видимому, оказывают большее влияние на высвобождение дофамина в оболочке, чем в ядре NAc. - ^ Берридж К.К., Крингельбах М.Л. (1 июня 2013 г.). «Нейронаука аффекта: мозговые механизмы удовольствия и неудовольствия» . Современное мнение в нейробиологии . 23 (3): 294–303. дои : 10.1016/j.conb.2013.01.017 . ПМЦ 3644539 . ПМИД 23375169 .
Например, мезолимбический дофамин, вероятно, самый популярный нейромедиатор мозга, отвечающий за удовольствие два десятилетия назад, оказывается, вообще не вызывает удовольствия или симпатии. Скорее, дофамин более избирательно опосредует мотивационный процесс значимости стимулов, который является механизмом желания вознаграждений, а не симпатии к ним... Скорее, опиоидная стимуляция обладает особой способностью усиливать симпатию, только если стимуляция происходит в анатомической «горячей точке».
- ^ Калипари, Эрин С.; Багот, Розмари К.; Пурушотаман, Иммануил; Дэвидсон, Томас Дж.; Йоргасон, Джордан Т.; Пенья, Кэтрин Дж.; Уокер, Дина М.; Пирпиниас, Стивен Т.; Гиз, Кевин Г.; Рамакришнан, Чару; Дейсерот, Карл; Нестлер, Эрик Дж. (8 марта 2016 г.). «Визуализация in vivo идентифицирует временную подпись средних шипистых нейронов D1 и D2 в вознаграждении за кокаин» . Труды Национальной академии наук . 113 (10): 2726–2731. Бибкод : 2016PNAS..113.2726C . дои : 10.1073/pnas.1521238113 . ПМК 4791010 . ПМИД 26831103 .
- ^ Балики, Миннесота; Мансур, А.; Бария, АТ; Хуанг, Л.; Бергер, SE; Филдс, HL; Апкарян А.В. (9 октября 2013 г.). «Разделение прилежащего человека на предполагаемое ядро и оболочку диссоциирует кодирование ценностей вознаграждения и боли» . Журнал неврологии . 33 (41): 16383–16393. doi : 10.1523/JNEUROSCI.1731-13.2013 . ПМЦ 3792469 . ПМИД 24107968 .
- ^ Соарес-Кунья, Карина; Коимбра, Барбара; Соуза, Нуно; Родригес, Ана Дж. (сентябрь 2016 г.). «Переоценка полосатых D1- и D2-нейронов при вознаграждении и отвращении». Неврологические и биоповеденческие обзоры . 68 : 370–386. doi : 10.1016/j.neubiorev.2016.05.021 . hdl : 1822/47044 . ПМИД 27235078 . S2CID 207092810 .
- ^ Бэмфорд, Найджел С.; Вайтман, Р. Марк; Зульцер, Дэвид (февраль 2018 г.). «Влияние дофамина на кортикостриатальные синапсы во время поведения, основанного на вознаграждении» . Нейрон . 97 (3): 494–510. дои : 10.1016/j.neuron.2018.01.006 . ПМЦ 5808590 . ПМИД 29420932 .
- ^ Соарес-Кунья, Карина; Коимбра, Барбара; Давид-Перейра, Ана; Борхес, Соня; Пинто, Луиза; Коста, Патрисио; Соуза, Нуно; Родригес, Ана Дж. (сентябрь 2016 г.). «Активация нейронов, экспрессирующих дофаминовые рецепторы D2, в прилежащем ядре повышает мотивацию» . Природные коммуникации . 7 (1): 11829. Бибкод : 2016NatCo...711829S . дои : 10.1038/ncomms11829 . ПМК 4931006 . ПМИД 27337658 .
- ^ Соарес-Кунья, Карина; Коимбра, Барбара; Домингес, Ана Вероника; Васконселос, Нивалдо; Соуза, Нуно; Родригес, Ана Жуан (март 2018 г.). «Микросхема прилежащего ядра, лежащая в основе увеличения мотивации, вызванного D2-MSN» . эНейро . 5 (2): ЭНЕВРО.0386–18.2018. дои : 10.1523/ENEURO.0386-18.2018 . ПМЦ 5957524 . ПМИД 29780881 .
- ^ Кооб Г.Ф., Ле Моал М. (2008). Зависимость и система антивознаграждения мозга. Анну. Преподобный Психол. 59 29–53. 10.1146/annurev.psych.59.103006.093548 [PubMed] [CrossRef] [Google Scholar]Кооб Г.Ф., Санна П.П., Блум Ф.Э. (1998). Нейронаука зависимости. Нейрон 21 467–476
- ^ Мейер, Дж. С., и Квенцер, Л. Ф. (2013). Психофармакология: Наркотики, мозг и поведение. Синауэр Ассошиэйтс.
- ^ Jump up to: а б Инь, ХХ; Остлунд, SB; Баллейн, BW (октябрь 2008 г.). «Обучение, ориентированное на вознаграждение, помимо дофамина в прилежащем ядре: интегративные функции сетей кортико-базальных ганглиев» . Европейский журнал неврологии . 28 (8): 1437–48. дои : 10.1111/j.1460-9568.2008.06422.x . ПМК 2756656 . ПМИД 18793321 .
- ^ Даян, П; Берридж, КК (июнь 2014 г.). «Павловское обучение с вознаграждением на основе моделей и без моделей: переоценка, пересмотр и раскрытие» . Когнитивная, аффективная и поведенческая нейронаука . 14 (2): 473–92. дои : 10.3758/s13415-014-0277-8 . ПМК 4074442 . ПМИД 24647659 .
- ^ Баллейн, BW; Моррис, RW; Люнг, БК (2 декабря 2015 г.). «Таламокортикальная интеграция инструментального обучения и производительности и их распад при зависимости». Исследования мозга . 1628 (Часть А): 104–16. дои : 10.1016/j.brainres.2014.12.023 . ПМИД 25514336 . S2CID 11776683 .
Важно отметить, что мы обнаружили доказательства повышенной активности прямого пути; В нейронах D1-прямого пути повышались внутриклеточные изменения экспрессии маркера пластичности pERK и соотношения AMPA/NMDA, вызванные стимуляцией корковых афферентов. Напротив, нейроны D2 показали противоположное изменение пластичности; стимуляция корковых афферентов снижает соотношение AMPA/NMDA в этих нейронах (Shan et al., 2014).
- ^ Наканиши, С; Хикида, Т; Явата, С. (12 декабря 2014 г.). «Четкий дофаминергический контроль прямых и косвенных путей обучения, основанного на вознаграждении и избегании». Нейронаука . 282 : 49–59. doi : 10.1016/j.neuroscience.2014.04.026 . ПМИД 24769227 . S2CID 21652525 .
- ^ Шифлетт, Миссури; Баллейн, BW (15 сентября 2011 г.). «Молекулярные субстраты контроля действия в кортико-стриатальных цепях» . Прогресс нейробиологии . 95 (1): 1–13. doi : 10.1016/j.pneurobio.2011.05.007 . ПМК 3175490 . ПМИД 21704115 .
- ^ Шульц, В. (апрель 2013 г.). «Обновление сигналов вознаграждения дофамина» . Современное мнение в нейробиологии . 23 (2): 229–38. дои : 10.1016/j.conb.2012.11.012 . ПМЦ 3866681 . ПМИД 23267662 .
- ^ Шифлетт, Миссури; Баллейн, BW (17 марта 2011 г.). «Вклад передачи сигналов ERK в полосатом теле в инструментальное обучение и производительность» . Поведенческие исследования мозга . 218 (1): 240–7. дои : 10.1016/j.bbr.2010.12.010 . ПМК 3022085 . ПМИД 21147168 .
- ^ Jump up to: а б Нестлер Э.Дж. (декабрь 2013 г.). «Клеточная основа памяти при наркомании» . Диалоги Клин. Нейроски . 15 (4): 431–443. ПМЦ 3898681 . ПМИД 24459410 .
- ^ Jump up to: а б Раффл Дж.К. (ноябрь 2014 г.). «Молекулярная нейробиология зависимости: о чем вообще (Δ)FosB?». Являюсь. J. Злоупотребление наркотиками и алкоголем . 40 (6): 428–437. дои : 10.3109/00952990.2014.933840 . ПМИД 25083822 . S2CID 19157711 .
Сильная корреляция между хроническим воздействием наркотиков и ΔFosB открывает новые возможности для таргетной терапии зависимости (118) и предлагает методы анализа их эффективности (119). За последние два десятилетия исследования продвинулись от выявления индукции ΔFosB к изучению его последующего действия (38). Вполне вероятно, что исследования ΔFosB теперь перейдут в новую эру – использование ΔFosB в качестве биомаркера. ...
Выводы
ΔFosB является важным фактором транскрипции, участвующим в молекулярных и поведенческих путях развития зависимости после неоднократного воздействия наркотиков. Образование ΔFosB во многих областях мозга и молекулярный путь, ведущий к образованию комплексов AP-1, хорошо изучены. Установление функционального назначения ΔFosB позволило продолжить определение некоторых ключевых аспектов его молекулярных каскадов, включая такие эффекторы, как GluR2 (87,88), Cdk5 (93) и NFkB (100). Более того, многие из этих выявленных молекулярных изменений теперь напрямую связаны со структурными, физиологическими и поведенческими изменениями, наблюдаемыми после хронического воздействия наркотиков (60,95,97,102). Новые горизонты исследований по изучению молекулярной роли ΔFosB были открыты эпигенетическими исследованиями, а недавние достижения проиллюстрировали роль ΔFosB, действующего на ДНК и гистоны, действительно как молекулярный переключатель (34). Благодаря нашему лучшему пониманию ΔFosB при зависимости, стало возможным оценить потенциал привыкания современных лекарств (119), а также использовать его в качестве биомаркера для оценки эффективности терапевтических вмешательств (121,122,124). Некоторые из этих предлагаемых мер имеют ограничения (125) или находятся на начальной стадии разработки (75). Однако есть надежда, что некоторые из этих предварительных результатов могут привести к созданию инновационных методов лечения, которые крайне необходимы при зависимости. - ^ Jump up to: а б Олсен CM (декабрь 2011 г.). «Естественные награды, нейропластичность и ненаркотическая зависимость» . Нейрофармакология . 61 (7): 1109–1122. doi : 10.1016/j.neuropharm.2011.03.010 . ПМК 3139704 . ПМИД 21459101 .
Исследования функциональной нейровизуализации на людях показали, что азартные игры (Breiter et al, 2001), шоппинг (Knutson et al, 2007), оргазм ( Komisaruk et al, 2004), видеоигры (Koepp et al, 1998; Hoeft et al, 2008) ) и вид аппетитной еды (Wang et al, 2004a) активируют многие из тех же областей мозга (т.е. мезокортиколимбическую систему и расширенную миндалевидное тело), что и наркотики (Volkow et al, 2004). ... Перекрестная сенсибилизация также является двунаправленной, поскольку прием амфетамина в анамнезе облегчает сексуальное поведение и усиливает связанное с этим увеличение NAc DA ... Как описано в отношении пищевого вознаграждения, сексуальный опыт также может привести к активации сигнальных каскадов, связанных с пластичностью. Транскрипционный фактор delta FosB увеличивается в NAc, PFC, дорсальном полосатом теле и VTA после повторного сексуального поведения (Wallace et al., 2008; Pitchers et al., 2010b). Это естественное увеличение дельта-FosB или вирусная сверхэкспрессия дельта-FosB в NAc модулирует сексуальную активность, а блокада дельта-FosB NAc ослабляет это поведение (Hedges et al, 2009; Pitchers et al., 2010b). Кроме того, вирусная сверхэкспрессия дельта-FosB усиливает обусловленное предпочтение места в сочетании с сексуальным опытом (Hedges et al., 2009). ... У некоторых людей происходит переход от «нормального» к компульсивному занятию естественными вознаграждениями (такими как еда или секс), состояние, которое некоторые называют поведенческой или ненаркотической зависимостью (Holden, 2001; Grant et al. , 2006а). ... У людей роль передачи сигналов дофамина в процессах стимулирующей сенсибилизации недавно была подчеркнута наблюдением синдрома нарушения регуляции дофамина у некоторых пациентов, принимающих дофаминергические препараты. Этот синдром характеризуется вызванным приемом лекарств увеличением (или компульсивным) вовлечением в ненаркотические вознаграждения, такие как азартные игры, покупки или секс (Evans et al, 2006; Aiken, 2007; Lader, 2008)».
Таблица 1. Сводная информация о пластичности, наблюдаемой после воздействия лекарственного средства или натуральных подкреплений . - ^ Jump up to: а б Билински П., Войтыла А., Капка-Скшипчак Л., Хведорович Р., Циранка М., Студзински Т. (2012). «Эпигенетическая регуляция при наркомании». Энн. Сельское хозяйство. Окружающая среда. Мед . 19 (3): 491–496. ПМИД 23020045 .
По этим причинам ΔFosB считается основным и причинным фактором транскрипции в создании новых нейронных связей в центре вознаграждения, префронтальной коре и других областях лимбической системы. Это отражается на повышенном, стабильном и продолжительном уровне чувствительности к кокаину и другим наркотикам, а также на склонности к рецидивам даже после длительных периодов воздержания. Эти вновь построенные сети функционируют очень эффективно по новым путям, как только злоупотребляют наркотиками... Таким образом, индукция экспрессии гена CDK5 происходит вместе с подавлением гена G9A, кодирующего диметилтрансферазу, действующего на гистон H3. Механизм обратной связи можно наблюдать в регуляции этих двух важнейших факторов, определяющих адаптивный эпигенетический ответ на кокаин. Это зависит от ингибирования ΔFosB экспрессии гена G9a, т.е. синтеза H3K9me2, который, в свою очередь, ингибирует факторы транскрипции ΔFosB. По этой причине наблюдаемая гиперэкспрессия G9a, которая обеспечивает высокие уровни диметилированной формы гистона H3, устраняет эффекты нейрональной структуры и пластичности, вызванные кокаином, посредством этой обратной связи, которая блокирует транскрипцию ΔFosB.
- ^ Питчерс К.К., Виалу В., Нестлер Э.Дж., Лавиолетт С.Р., Леман М.Н., Кулен Л.М. (февраль 2013 г.). «Естественные и лекарственные вознаграждения действуют на общие механизмы нейронной пластичности, при этом ΔFosB является ключевым медиатором» . Журнал неврологии . 33 (8): 3434–3442. doi : 10.1523/JNEUROSCI.4881-12.2013 . ПМЦ 3865508 . ПМИД 23426671 .
Злоупотребление наркотиками вызывает нейропластичность в естественном пути вознаграждения, в частности в прилежащем ядре (NAc), тем самым вызывая развитие и проявление аддиктивного поведения. ... В совокупности эти результаты показывают, что злоупотребление наркотиками и естественное поведение вознаграждения действуют на общие молекулярные и клеточные механизмы пластичности, которые контролируют уязвимость к наркозависимости, и что эта повышенная уязвимость опосредована ΔFosB и его нижестоящими мишенями транскрипции. ... Сексуальное поведение приносит большое удовлетворение (Tenk et al., 2009), а сексуальный опыт вызывает сенсибилизированное поведение, связанное с наркотиками, включая перекрестную сенсибилизацию к двигательной активности, индуцированной амфетамином (Amph) (Bradley and Meisel, 2001; Pitchers et al. ., 2010a) и повышенное вознаграждение Amph (Pitchers et al., 2010a). Более того, сексуальный опыт вызывает нейронную пластичность NAc, аналогичную той, которая вызывается воздействием психостимуляторов, включая увеличение плотности дендритных отростков (Meisel and Mullins, 2006; Pitchers et al., 2010a), изменение трафика глутаматных рецепторов и снижение синаптической силы в префронтальной коре. -отвечающие нейроны оболочки NAc (Pitchers et al., 2012). Наконец, было обнаружено, что периоды воздержания от сексуального опыта имеют решающее значение для усиления вознаграждения Amph, спиногенеза NAc (Pitchers et al., 2010a) и торговли глутаматными рецепторами (Pitchers et al., 2012). Эти результаты показывают, что естественное и лекарственное вознаграждение имеют общие механизмы нейронной пластичности.
- ^ Белоате Л.Н., Уимс П.В., Кейси Г.Р., Уэбб И.К., Кулен Л.М. (февраль 2016 г.). «Активация рецептора NMDA прилежащего ядра регулирует перекрестную сенсибилизацию к амфетамину и экспрессию deltaFosB после сексуального опыта у самцов крыс». Нейрофармакология . 101 : 154–164. doi : 10.1016/j.neuropharm.2015.09.023 . ПМИД 26391065 . S2CID 25317397 .
- ^ Робисон А.Дж., Нестлер Э.Дж. (ноябрь 2011 г.). «Транскрипционные и эпигенетические механизмы зависимости» . Нат. Преподобный Нейроски . 12 (11): 623–637. дои : 10.1038/nrn3111 . ПМЦ 3272277 . ПМИД 21989194 .
ΔFosB служит одним из главных контролирующих белков, управляющих этой структурной пластичностью. ... ΔFosB также подавляет экспрессию G9a, что приводит к снижению репрессивного метилирования гистонов в гене cdk5. Конечным результатом является активация генов и увеличение экспрессии CDK5. ... Напротив, ΔFosB связывается с геном c-fos и рекрутирует несколько ко-репрессоров, включая HDAC1 (деацетилаза гистонов 1) и SIRT 1 (сиртуин 1). ... Конечным результатом является репрессия гена c-fos .
Рисунок 4: Эпигенетические основы лекарственной регуляции экспрессии генов. - ^ Хичкок Л.Н., Латтал К.М. (2014). «Гистон-опосредованная эпигенетика при зависимости». Эпигенетика и нейропластичность — данные и дебаты . Прогресс молекулярной биологии и трансляционной науки. Том. 128. стр. 51–87. дои : 10.1016/B978-0-12-800977-2.00003-6 . ISBN 9780128009772 . ПМЦ 5914502 . ПМИД 25410541 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Уокер Д.М., Нестлер Э.Дж. (2018). «Нейроэпигенетика и зависимость». Нейрогенетика, Часть II . Справочник по клинической неврологии. Том. 148. стр. 747–765. дои : 10.1016/B978-0-444-64076-5.00048-X . ISBN 9780444640765 . ПМЦ 5868351 . ПМИД 29478612 .
- ^ Позвонил HP (2003). Фармакология . Эдинбург: Черчилль Ливингстон. п. 596. ИСБН 978-0-443-07145-4 .
- ^ Jump up to: а б Рой А. Уайз, Активация лекарственными средствами путей вознаграждения мозга, Зависимость от наркотиков и алкоголя , 1998; 51 13–22.
- ^ Гедерс Н.Е., Смит Дж.Е. (1983). «Корковое дофаминергическое участие в подкреплении кокаина». Наука . 221 (4612): 773–775. Бибкод : 1983Sci...221..773G . дои : 10.1126/science.6879176 . ПМИД 6879176 .
- ^ Гедерс Н.Е., Смит Дж.Е. (1993). «Внутричерепное самостоятельное введение кокаина в медиальную префронтальную кору увеличивает обмен дофамина в прилежащем ядре». Дж. Фармакол. Эксп. Там . 265 (2): 592–600. PMID 8496810 .
- ^ Кларк, Хоммер Д.В.; Перт А.; Скирболл Л.Р. (1985). «Электрофизиологическое действие никотина на отдельные единицы черной субстанции» . Бр. Дж. Фармакол . 85 (4): 827–835. дои : 10.1111/j.1476-5381.1985.tb11081.x . ЧВК 1916681 . ПМИД 4041681 .
- ^ Вестфолл, Томас К.; Грант, Хизер; Перри, Холли (январь 1983 г.). «Высвобождение дофамина и 5-гидрокситриптамина из срезов полосатого тела крысы после активации никотиновых холинергических рецепторов». Общая фармакология: Сосудистая система . 14 (3): 321–325. дои : 10.1016/0306-3623(83)90037-x . ПМИД 6135645 .
- ^ Рёмер Томсен, К.; Уайброу, ПК; Крингельбах, МЛ (2015). «Реконцептуализация ангедонии: новые взгляды на балансировку сетей удовольствия в человеческом мозге» . Границы поведенческой нейронауки . 9 : 49. дои : 10.3389/fnbeh.2015.00049 . ПМЦ 4356228 . ПМИД 25814941 .
- ^ Томсен, КР (2015). «Измерение ангедонии: нарушение способности добиваться, испытывать и узнавать о вознаграждении» . Границы в психологии . 6 : 1409. doi : 10.3389/fpsyg.2015.01409 . ПМК 4585007 . ПМИД 26441781 .
- ^ Олни, Джей-Джей; Варлоу, С.М.; Нафцигер, Э.Э.; Берридж, КК (август 2018 г.). «Современные взгляды на значимость стимулов и их применение к клиническим расстройствам» . Современное мнение в области поведенческих наук . 22 : 59–69. дои : 10.1016/j.cobeha.2018.01.007 . ПМЦ 5831552 . ПМИД 29503841 .
- ^ Преда, Адриан (2014). «Мозговые изображения, коррелирующие ангедонии». В Ритснер, Майкл (ред.). Ангедония: всеобъемлющий справочник . Дордрехт: Springer Нидерланды. ISBN 978-94-017-8590-7 .
- ^ Чжан, Б; Лин, П; Ши, Х; Онгур, Д; Ауэрбах, Р.П.; Ван, X; Яо, С; Ван, X (сентябрь 2016 г.). «Картирование дисфункции, специфичной для ангедонии, в трансдиагностическом подходе: метаанализ ALE» . Мозговые изображения и поведение . 10 (3): 920–39. дои : 10.1007/s11682-015-9457-6 . ПМЦ 4838562 . ПМИД 26487590 .
- ^ Тредуэй, Майкл, Т. (2016). «Нейробиология мотивационного дефицита при депрессии - обновленная информация о потенциальных патомеханизмах». В Симпсоне, Элеонора Х.; Бальзам, Питер Д. (ред.). Поведенческая нейронаука мотивации (актуальные темы поведенческой нейронауки) (1-е изд.). Швейцария: Международное издательство Springer. п. 343. ИСБН 978-3-319-26933-7 .
В сравнительно недавней литературе исследования мотивации и подкрепления при депрессии в значительной степени показали различия в выявлении различий по сравнению со здоровыми людьми (Whitton et al. 2015). В нескольких исследованиях с использованием затрат усилий на задание вознаграждения (EEfRT) пациенты с БДР затрачивали меньше усилий на вознаграждение по сравнению с контрольной группой (Treadway et al. 2012; Yang et al. 2014).
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Саламоне, доктор юридических наук; Йон, ЮВ; Лопес-Крус, Л; Сан-Мигель, Север; Корреа, М. (май 2016 г.). «Активные и связанные с усилием аспекты мотивации: нейронные механизмы и последствия для психопатологии» . Мозг: журнал неврологии . 139 (Часть 5): 1325–47. дои : 10.1093/brain/aww050 . ПМЦ 5839596 . ПМИД 27189581 .
- ^ Руссо, С.Дж.; Нестлер, Э.Дж. (сентябрь 2013 г.). «Схема вознаграждения мозга при расстройствах настроения» . Обзоры природы. Нейронаука . 14 (9): 609–25. дои : 10.1038/nrn3381 . ПМЦ 3867253 . ПМИД 23942470 .
- ^ Тредуэй, Монтана; Зальд, Д.Х. (январь 2011 г.). «Переосмысление ангедонии при депрессии: уроки трансляционной нейробиологии» . Неврологические и биоповеденческие обзоры . 35 (3): 537–55. doi : 10.1016/j.neubiorev.2010.06.006 . ПМК 3005986 . ПМИД 20603146 .
- ^ Уолш, Джей-Джей; Хан, Миннесота (12 декабря 2014 г.). «Неоднородность нейронов вентральной покрышки: проекционные функции в контексте, связанном с настроением» . Нейронаука . 282 : 101–8. doi : 10.1016/j.neuroscience.2014.06.006 . ПМЦ 4339667 . ПМИД 24931766 .
- ^ Ламмель, С; Лим, БК; Маленка, РЦ (январь 2014 г.). «Награда и отвращение в гетерогенной дофаминовой системе среднего мозга» . Нейрофармакология . 76 Часть Б: 351–9. doi : 10.1016/j.neuropharm.2013.03.019 . ПМЦ 3778102 . ПМИД 23578393 .
- ^ Ноулэнд, Д; Лим, БК (5 января 2018 г.). «Схемы депрессивного поведения: роль схемы вознаграждения и не только» . Фармакология Биохимия и поведение . 174 : 42–52. дои : 10.1016/j.pbb.2017.12.010 . ПМК 6340396 . ПМИД 29309799 .
- ^ Jump up to: а б Ламмель, С; Тай, К.М.; Уорден, MR (январь 2014 г.). «Прогресс в понимании расстройств настроения: оптогенетическое рассечение нейронных цепей» . Гены, мозг и поведение . 13 (1): 38–51. дои : 10.1111/gbb.12049 . ПМИД 23682971 . S2CID 18542868 .
- ^ Буччи, П; Галдериси, С. (май 2017 г.). «Классификация и оценка негативных симптомов». Современное мнение в психиатрии . 30 (3): 201–208. дои : 10.1097/YCO.0000000000000322 . ПМИД 28212174 . S2CID 9923114 .
Они также дают отдельную оценку консумматорной ангедонии (снижение ощущения удовольствия, получаемого от продолжающихся приятных занятий) и предвосхищающей ангедонии (снижение способности предвидеть будущее удовольствие). Фактически, первый при шизофрении кажется относительно сохранным, тогда как второй, по-видимому, нарушается [32 – 34]. Однако имеются и противоречивые данные [35].
- ^ Янг, Антицевич и Барч, 2018 , стр. 215a: «Несколько недавних обзоров (например, Cohen and Minor, 2010) показали, что люди, страдающие шизофренией, демонстрируют относительно сохранные эмоциональные реакции на вызывающие аффект стимулы, а также другие показатели сохранной реакции... Возникает более смешанная картина. из функциональных исследований нейровизуализации, изучающих реакцию мозга на другие типы приятных стимулов при шизофрении (Paradiso et al., 2003)»
- ^ Янг, Антицевич и Барч, 2018 , стр. 215b: «Поэтому удивительно, что поведенческие исследования показали, что обучение с подкреплением сохраняется при шизофрении, когда обучение является относительно неявным (хотя см. Siegert et al., 2008 о доказательствах нарушения обучения задачам с последовательным временем реакции), но более ухудшается, когда необходимы явные представления о непредвиденных обстоятельствах стимул-вознаграждение (см. Gold et al., 2008). Эта закономерность привела к появлению теории о том, что система обучения с постепенным подкреплением, опосредованная полосатым телом, может оставаться неизменной при шизофрении, хотя и более быстрой, в режиме онлайн, кортикально. системы опосредованного обучения нарушены».
- ^ Янг, Антицевич и Барч, 2018 , стр. 216: «Недавно мы показали, что люди с шизофренией могут демонстрировать улучшение когнитивного контроля, когда информация о вознаграждениях представлена извне, а не тогда, когда ее необходимо поддерживать внутри (Mann et al., 2013), с некоторыми доказательствами нарушений в DLPFC и полосатом теле. активация во время внутреннего поддержания информации о вознаграждении связана с различиями в мотивации людей (Чунг и Барч, 2016)».
- ^ Литтман, доктор философии, Эллен (февраль 2017 г.). «Никогда не достаточно? Почему мозг с СДВГ жаждет стимуляции» . Журнал ADDditude . ООО «Нью Хоуп Медиа» . Проверено 27 мая 2021 г.
- ^ Чендлер DJ, Waterhouse BD, Гао WJ (май 2014 г.). «Новые взгляды на катехоламинергическую регуляцию исполнительных цепей: доказательства независимой модуляции префронтальных функций дофаминергическими и норадренергическими нейронами среднего мозга» . Границы в нейронных цепях . 8 : 53. doi : 10.3389/fncir.2014.00053 . ПМК 4033238 . ПМИД 24904299 .
- ^ Блюм, Кеннет; Чен, Аманда Ли-Чуан; Браверман, Эрик Р.; Комингс, Дэвид Э; Чен, Томас Дж. Х.; Аркури, Ванесса; Блюм, Сет Х; Даунс, Бернард В.; Уэйт, Роджер Л.; Нотаро, Элисон; Любар, Джоэл; Уильямс, Лонна; Прихода, Томас Дж; Паломо, Томас; Оскар-Берман, Марлен (октябрь 2008 г.). «Синдром дефицита внимания и гиперактивности и синдром дефицита вознаграждения» . Нервно-психические заболевания и лечение . 4 (5): 893–918. дои : 10.2147/ndt.s2627 . ISSN 1176-6328 . ПМК 2626918 . ПМИД 19183781 .
- ^ Мудрый РА (1996). «Наркотики, вызывающие привыкание, и награда за стимуляцию мозга». Анну. Преподобный Нейроски . 19 : 319–340. дои : 10.1146/annurev.ne.19.030196.001535 . ПМИД 8833446 .
- ^ Jump up to: а б «Нервная система человека | Описание, развитие, анатомия и функции» . Британская энциклопедия . 23 января 2024 г.
- ^ Jump up to: а б Джеймс Олдс и Питер Милнер (декабрь 1954 г.). «Положительное подкрепление, вызываемое электрической стимуляцией области перегородки и других областей мозга крысы» . Журнал сравнительной и физиологической психологии . 47 (6): 419–427. дои : 10.1037/h0058775 . ПМИД 13233369 . Архивировано из оригинала 5 февраля 2012 года . Проверено 26 апреля 2011 г.
- ^ Крингельбах, Мортен Л.; Берридж, Кент К. (25 июня 2010 г.). «Функциональная нейроанатомия удовольствия и счастья» . Медицина открытий . 9 (49): 579–587. ПМК 3008353 . PMID 20587348 – через www.discoverymedicine.com.
- ^ Иван Петрович Павлов; Г. В. Анреп (2003). Условные рефлексы . Курьерская корпорация. стр. 1–. ISBN 978-0-486-43093-5 .
- ^ Фридлунд, Алан и Джеймс Калат. Разум и мозг, наука психология. Калифорния: Cengage Learning, 2014. Печать.
- ^ Хан В., Теллес Л.А., Перкинс М.Х., Перес И.О., Ку Т., Феррейра Дж.; и др. (2018). «Нейронная цепь для вознаграждения, вызываемого кишечником» . Клетка . 175 (3): 665–678.e23. дои : 10.1016/j.cell.2018.08.049 . ПМК 6195474 . ПМИД 30245012 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Берридж, Кент К.; Крингельбах, Мортен Л. (август 2008 г.). «Аффективная нейробиология удовольствия: вознаграждение у людей и животных» . Психофармакология . 199 (3): 457–480. дои : 10.1007/s00213-008-1099-6 . ПМК 3004012 . ПМИД 18311558 .
- ^ Jump up to: а б Феррери Л., Мас-Эрреро Э., Заторре Р.Х., Риполлес П., Гомес-Андрес А., Аликарт Х., Оливе Г., Марко-Пальярес Х., Антонихоан Р.М., Валле М., Риба Х., Родригес-Форнеллс А. (январь 2019 г.). «Дофамин модулирует вознаграждение, вызываемое музыкой» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (9): 3793–3798. Бибкод : 2019PNAS..116.3793F . дои : 10.1073/pnas.1811878116 . ПМК 6397525 . ПМИД 30670642 .
Прослушивание приятной музыки часто сопровождается измеримыми телесными реакциями, такими как мурашки по коже или озноб, обычно называемые «ознобом» или «дрожью». ... В целом, наши результаты прямо показали, что фармакологические вмешательства двунаправленно модулируют реакции вознаграждения, вызываемые музыкой. В частности, мы обнаружили, что рисперидон ухудшал способность участников испытывать музыкальное удовольствие, тогда как леводопа усиливала ее. ... Здесь, напротив, изучая реакции на абстрактные вознаграждения у людей, мы показываем, что манипуляция дофаминергической передачей влияет как на удовольствие (т.е. количество времени, в течение которого сообщается о ознобе, и эмоциональное возбуждение, измеряемое EDA), так и на мотивационные компоненты музыкального вознаграждения. (деньги готовы потратить). Эти данные позволяют предположить, что дофаминергическая передача сигналов является непременным условием не только для мотивационных реакций, как было показано на примере первичных и вторичных вознаграждений, но также и для гедонистических реакций на музыку. Этот результат подтверждает недавние открытия, показывающие, что дофамин также опосредует восприятие приятности, достигаемой от других типов абстрактных вознаграждений, и бросает вызов предыдущим выводам на животных моделях о первичных вознаграждениях, таких как еда.
- ^ Jump up to: а б Гупиль Л., Окутюрье Ж.Ж. (февраль 2019 г.). «Музыкальное удовольствие и музыкальные эмоции» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (9): 3364–3366. Бибкод : 2019PNAS..116.3364G . дои : 10.1073/pnas.1900369116 . ПМК 6397567 . ПМИД 30770455 .
В фармакологическом исследовании, опубликованном в PNAS, Ferreri et al. (1) представили доказательства того, что усиление или ингибирование передачи сигналов дофамина с помощью леводопы или рисперидона модулирует удовольствие, испытываемое при прослушивании музыки. ... В последней попытке установить не только корреляционную, но и причинно-следственную связь дофамина с музыкальным удовольствием, авторы обратились к прямому манипулированию дофаминергической передачей сигналов в полосатом теле, сначала применяя возбуждающую и тормозящую транскраниальную магнитную стимуляцию к мозгу участников. левая дорсолатеральная префронтальная кора, область, которая, как известно, модулирует функцию полосатого тела (5), и, наконец, в текущем исследовании, путем введения фармацевтических агентов, способных изменить доступность синапсов дофамина (1), оба из которых влияют на восприятие удовольствия, физиологические показатели возбуждения, и денежная стоимость, присвоенная музыке в предсказанном направлении. ... Хотя вопрос о музыкальном выражении эмоций имеет долгую историю исследований, в том числе в PNAS (6), а психофизиологическое направление исследований 1990-х годов уже установило, что музыкальное удовольствие может активировать вегетативную нервную систему (7), Демонстрация авторами влияния системы вознаграждения на музыкальные эмоции была воспринята как первое доказательство того, что это настоящие эмоции, исследование которых имеет полную законность для информирования нейробиологии о наших повседневных когнитивных, социальных и аффективных функциях (8). Кстати, это направление работы, кульминацией которого стала статья Феррери и др. (1), вероятно, сделал больше для привлечения финансирования исследований в области музыкальных наук, чем кто-либо другой в этом сообществе. Доказательства Феррери и др. (1) обеспечивает новейшую поддержку убедительной нейробиологической модели, в которой музыкальное удовольствие возникает в результате взаимодействия древних систем вознаграждения/оценки (полосатого, лимбического и паралимбического) с более филогенетически развитыми системами восприятия/предсказания (темпорофронтальными).
- Янг, Джаред В.; Античевич, Алан; Барч, Дина М. (2018). «Когнитивная и мотивационная нейронаука психотических расстройств». В Чарни, Деннис С.; Склар, Памела; Буксбаум, Джозеф Д.; Нестлер, Эрик Дж. (ред.). Нейробиология психических заболеваний Чарни и Нестлера (5-е изд.). Нью-Йорк: Издательство Оксфордского университета. ISBN 9780190681425 .
Внешние ссылки
[ редактировать ]| Зависимость |
| ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Зависимость |
| ||||||||||
| Уход и управление |
| ||||||||||
| См. также | |||||||||||