Оптогенетика
Оптогенетика — это биологический метод управления активностью нейронов или других типов клеток с помощью света. Это достигается за счет экспрессии светочувствительных ионных каналов , насосов или ферментов специально в клетках-мишенях. На уровне отдельных активируемые клеток светом ферменты и факторы транскрипции позволяют точно контролировать биохимические сигнальные пути. [1] В системной нейробиологии способность контролировать активность генетически определенного набора нейронов использовалась для понимания их вклада в принятие решений. [2] обучение, [3] память о страхе, [4] спаривание, [5] зависимость, [6] кормление, [7] и локомоция. [8] В ходе первого медицинского применения оптогенетической технологии зрение было частично восстановлено у слепого пациента с пигментным ретинитом . [9]
Оптогенетические методы также были внедрены для картирования функциональных связей мозга . [10] [11] Изменяя активность генетически меченных нейронов светом и используя методы визуализации и электрофизиологии для регистрации активности других клеток, исследователи могут выявить статистические зависимости между клетками и областями мозга. [12] [13]
В более широком смысле область оптогенетики также включает методы регистрации клеточной активности с помощью генетически закодированных показателей .
выбрал оптогенетику «методом года» во всех областях науки и техники В 2010 году междисциплинарный исследовательский журнал Nature Methods . [14] В том же году статья «Прорывы десятилетия» в академическом исследовательском журнале Science посвящена оптогенетике. [15] [16] [17]
История
[ редактировать ]В 1979 году Фрэнсис Крик предположил, что контроль над всеми клетками одного типа мозга, оставляя при этом остальные более или менее неизмененными, является настоящей проблемой для нейробиологии. Крик предположил, что технология, использующая свет, может быть полезна для контроля активности нейронов с временной и пространственной точностью, но в то время не было техники, позволяющей заставить нейроны реагировать на свет.
К началу 1990-х годов Л. К. Кац и Э. Каллауэй показали, что свет способен высвободить глутамат. [18] Хеберле и Бюльдт в 1994 году уже продемонстрировали функциональную гетерологичную экспрессию бактериородопсина для светоактивируемого потока ионов у дрожжей. [19]
В 1995 году Георг Нагель и др. и Эрнст Бамберг попробовал гетерологичную экспрессию микробных родопсинов (также бактериородопсина, а также в не-нейронной системе, ооцитах Xenopus) (Georg Nagel et al., 1995, FEBS Lett.) и продемонстрировал светоиндуцированный ток.
Самый ранний генетически направленный метод, в котором использовался свет для контроля нейронов, сенсибилизированных родопсином, был описан в январе 2002 года Борисом Земельманом и Геро Мизенбеком , которые использовали культивированные родопсин дрозофилы нейроны млекопитающих. [20] В 2003 году Земельман и Мизенбёк разработали второй метод светозависимой активации нейронов, в котором отдельные ионотропные каналы TRPV1, TRPM8 и P2X2 закрывались фотоклеточными лигандами в ответ на свет. [21] Начиная с 2004 года группы Крамера и Исакова в сотрудничестве с группой Траунера разработали органические фотопереключатели или соединения с «обратимо замкнутыми клетками» , которые могли взаимодействовать с генетически введенными ионными каналами. [22] [23] Методика TRPV1, хотя и без триггера освещения, впоследствии использовалась несколькими лабораториями для изменения питания, передвижения и поведенческой устойчивости лабораторных животных. [24] [25] [26] Однако основанные на освещении подходы к изменению активности нейронов не применялись за пределами первоначальных лабораторий, вероятно, потому, что вскоре после этого был клонирован более простой в использовании канальный родопсин. [27]
Питер Хегеманн , изучая световую реакцию зеленых водорослей в Регенсбургском университете, обнаружил фототоки, которые были слишком быстрыми, чтобы их можно было объяснить классическими родопсинами животных, связанными с g-белком . [28] Объединившись с электрофизиологом Георгом Нагелем из Института Макса Планка во Франкфурте, они смогли продемонстрировать, что единственный ген водоросли Chlamydomonas производит большие фототоки при экспрессии в ооците лягушки. [29] Чтобы идентифицировать экспрессирующие клетки, они заменили цитоплазматический хвост белка водоросли флуоресцентным белком YFP , создав первый широко применимый оптогенетический инструмент. [27] В статье 2003 года они заявили, что «экспрессия ChR2 в ооцитах или клетках млекопитающих может использоваться как мощный инструмент для увеличения цитоплазматического Ca». 2+ концентрации или деполяризации клеточной мембраны просто путем освещения».
Карл Дейссерот из факультета биоинженерии Стэнфорда в начале июля 2004 года опубликовал страницы своего первого эксперимента, показывающие световую активацию нейронов, экспрессирующих каналородопсин. [30] В августе 2005 года сотрудники его лаборатории, в том числе аспиранты Эд Бойден и Фэн Чжан , в сотрудничестве с Георгом Нагелем опубликовали первую демонстрацию однокомпонентной оптогенетической системы в нейронах. [31] с использованием мутанта каналородопсин-2(H134R)-eYFP от Георга Нагеля, который является первым мутантом каналородопсина-2 с момента его функциональной характеристики Георгом Нагелем и Хегеманном. [27]
Чжуо-Хуа Пан из Государственного университета Уэйна , исследуя возможность восстановления зрения при слепоте, попробовал направить родопсин в ганглиозные клетки — нейроны в глазах человека, которые напрямую связаны с мозгом. По словам Пана, первое наблюдение Паном оптической активации нейронов сетчатки канальным родопсином было в феврале 2004 года. [32] за пять месяцев до первого наблюдения Дейсерота в июле 2004 года. [33] Действительно, трансфицированные нейроны становились электрически активными в ответ на свет, и в 2005 году Чжуо-Хуа Пан сообщил об успешной трансфекции канального родопсина in vivo в ганглиозных клетках сетчатки мышей и об электрических реакциях на фотостимуляцию в культуре срезов сетчатки. [34] Этот подход в конечном итоге был реализован на пациенте-человеке Ботондом Роской и его коллегами в 2021 году. [9]
В апреле 2005 года Сусана Лима и Мизенбёк сообщили о первом использовании генетически направленной фотостимуляции P2X2 для контроля поведения животного. [35] Они показали, что фотостимуляция генетически ограниченных групп нейронов, например, нейронов дофаминергической системы, вызывает характерные поведенческие изменения у плодовых мух.
В октябре 2005 года Линн Ландмессер и Стефан Херлитце также опубликовали информацию об использовании каналороходпсина-2 для контроля активности нейронов в культивируемых нейронах гиппокампа и цепях спинного мозга кур в интактных развивающихся эмбрионах. [36] Кроме того, они впервые представили родопсин позвоночных, светоактивируемый рецептор, связанный с G-белком, в качестве инструмента для ингибирования активности нейронов посредством рекрутирования внутриклеточных сигнальных путей также в нейронах гиппокампа и интактном развивающемся курином эмбрионе. [36]
Группы Александра Готшалка и Георга Нагеля создали первый мутант ChR2 (H134R) и первыми использовали каналродопсин-2 для контроля активности нейронов у интактных животных, показав, что двигательные паттерны у круглых червей C. elegans могут быть вызваны световой стимуляцией генетически отобранные нейронные цепи (опубликовано в декабре 2005 г.). [37] У мышей контролируемая экспрессия оптогенетических инструментов часто достигается с помощью специфичных для клеточного типа методов Cre/loxP, разработанных для нейробиологии Джо З. Циеном еще в 1990-х годах. [38] активировать или ингибировать определенные области мозга и типы клеток in vivo . [39]
В 2007 году лаборатории Бойдена и Дайссерота (вместе с группами Готшалька и Георга Нагеля) одновременно сообщили об успешном оптогенетическом ингибировании активности нейронов. [40] [41]
В 2007 году группы Георга Нагеля и Хегемана начали оптогенетические манипуляции с цАМФ. [42] В 2014 году Авелар и др. сообщили о первом гене родопсин-гуанилилциклазы гриба. В 2015 году Шейб и др. и Гао и др. охарактеризовали активность гена родопсин-гуанилилциклазы. И Шицян Гао и др. и Георг Нагель, Александр Готшальк определили его как первый 8 ТМ родопсин. [43]
Описание
[ редактировать ]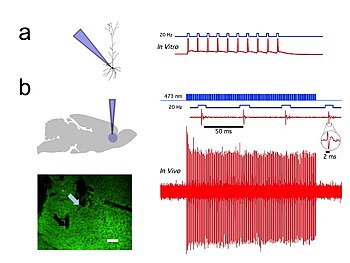
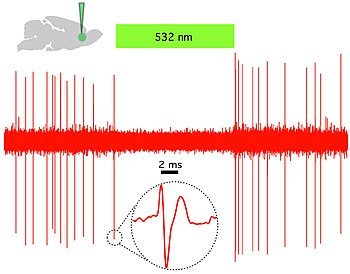
Оптогенетика обеспечивает временную точность в миллисекундном масштабе, что позволяет экспериментатору идти в ногу с быстрой обработкой биологической информации (например, при исследовании причинной роли определенных паттернов потенциала действия в определенных нейронах). Действительно, чтобы исследовать нервный код, оптогенетика по определению должна работать в миллисекундном масштабе времени, чтобы обеспечить добавление или удаление точных паттернов активности внутри конкретных клеток мозга интактных животных, включая млекопитающих (см. Рисунок 1) . Для сравнения, временная точность традиционных генетических манипуляций (используемых для исследования причинной роли конкретных генов внутри клеток посредством изменений «потеря функции» или «приобретение функции» в этих генах) довольно медленная, от нескольких часов до дней. до месяцев. Также важно иметь быстрые показания в оптогенетике, которые могут идти в ногу с оптическим контролем. Это можно сделать с помощью электрических записей («оптродов») или с помощью репортерных белков, которые являются биосенсорами , когда ученые соединяют флуоресцентные белки с белками-детекторами. Кроме того, помимо своего научного воздействия, оптогенетика представляет собой важный пример ценности обоих методов. экологическое сохранение (поскольку многие из ключевых инструментов оптогенетики возникают из микробных организмов, занимающих специализированные ниши в окружающей среде), а также в важности чистой фундаментальной науки, поскольку эти опсины изучались на протяжении десятилетий биофизиками и микробиологами ради них самих, без учета их потенциальная ценность в предоставлении информации о нейробиологии и нейропсихиатрических заболеваниях. [47]
Светоактивируемые белки: каналы, насосы и ферменты
Таким образом, отличительной чертой оптогенетики является внедрение быстрых, активируемых светом каналов, насосов и ферментов, которые позволяют точно во времени манипулировать электрическими и биохимическими событиями, сохраняя при этом разрешение типов клеток за счет использования специфических механизмов нацеливания. Среди микробных опсинов, которые можно использовать для исследования функции нервной системы, есть каналородопсины (ChR2, ChR1, VChR1 и SFO) для возбуждения нейронов и анионпроводящие каналородопсины для светоиндуцированного торможения. контролируемые косвенным светом, для предотвращения генерации потенциала действия в нейронах во время освещения синим светом. калиевые каналы, Недавно были созданы [48] [49] Для ингибирования активности нейронов также используются световые ионные насосы, например галородопсин (NpHR), [50] усиленные галородопсины (eNpHR2.0 и eNpHR3.0, см. рисунок 2), [51] архаэрродопсин (Arch), грибковые опсины (Mac) и усиленный бактериородопсин (eBR). [52]
Оптогенетический контроль четко определенных биохимических событий в поведении млекопитающих теперь также возможен. Опираясь на предыдущую работу по слиянию опсинов позвоночных со специфическими рецепторами, связанными с G-белком. [53] было создано семейство химерных однокомпонентных оптогенетических инструментов, которые позволили исследователям манипулировать поведением млекопитающих концентрацией определенных внутриклеточных мессенджеров, таких как цАМФ и IP3, в клетках-мишенях. [54] Вскоре после этого последовали и другие биохимические подходы к оптогенетике (особенно с инструментами, которые проявляли низкую активность в темноте), когда оптический контроль над малыми ГТФазами и аденилатциклазой был достигнут в культивируемых клетках с использованием новых стратегий, разработанных несколькими различными лабораториями. [55] [56] [57] Фотоактивируемые аденилатциклазы были обнаружены у грибов и успешно использованы для контроля уровня цАМФ в нейронах млекопитающих. [58] [59] Этот новый репертуар оптогенетических актуаторов теперь позволяет специфично для каждого типа клеток и точно во времени контролировать множество осей клеточных функций у интактных животных. [60]
Аппаратное обеспечение для легкого применения
Другим необходимым фактором является аппаратное обеспечение (например, интегрированные оптоволоконные и твердотельные источники света), позволяющее контролировать определенные типы клеток, даже глубоко внутри мозга, у свободно ведущих себя животных. Чаще всего последнее сейчас достигается с использованием диодной технологии с оптоволоконной связью, представленной в 2007 году. [61] [62] [63] хотя, чтобы избежать использования имплантированных электродов, исследователи разработали способы вписать «окно» из диоксида циркония, которое было модифицировано, чтобы стать прозрачным и имплантировано в черепа мышей, чтобы позволить оптическим волнам проникать более глубоко и стимулировать или подавлять отдельные нейроны. [64] Чтобы стимулировать поверхностные области мозга, такие как кора головного мозга, оптические волокна или светодиоды можно прикрепить непосредственно к черепу животного. Для доставки света в более глубокие области мозга используются более глубоко имплантированные оптические волокна. [65] В дополнение к оптоволоконным подходам были разработаны полностью беспроводные методы, использующие беспроводную подачу питания к головным светодиодам для беспрепятственного изучения сложного поведения свободно ведущих себя организмов. [66]
Экспрессия оптогенетических актуаторов
Оптогенетика также обязательно включает разработку стратегий генетического нацеливания, таких как клеточно-специфичные промоторы или другие адаптированные условно-активные вирусы, для доставки светочувствительных зондов к определенным популяциям нейронов в мозгу живых животных (например, червей, плодовых мух, мышей). , крысы и обезьяны). У беспозвоночных, таких как черви и плодовые мухи, некоторое количество полностью транс-ретиналя (ATR) поступает с пищей. Как отмечалось выше, ключевым преимуществом микробных опсинов является то, что они полностью функциональны без добавления экзогенных кофакторов у позвоночных. [63]
Техника
[ редактировать ]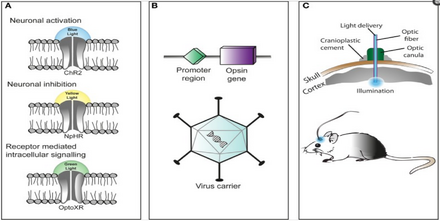
Техника использования оптогенетики гибка и адаптируется к потребностям экспериментатора. Катион-селективные каналородопсины (например, ChR2) используются для возбуждения нейронов, анион-проводящие каналородопсины (например, GtACR2) ингибируют активность нейронов. Объединение этих инструментов в единую конструкцию (например, BiPOLES) позволяет осуществлять как торможение, так и возбуждение в зависимости от длины волны освещения. [68]
Введение микробного опсина в определенную подгруппу клеток является сложной задачей. Популярным подходом является введение сконструированного вирусного вектора, содержащего ген оптогенетического актуатора, прикрепленный к специфическому промотору , такому как CAMKIIα , который активен в возбуждающих нейронах. Это обеспечивает некоторый уровень специфичности, предотвращая, например, экспрессию в глиальных клетках. [69]
Более конкретный подход основан на трансгенных мышах-"драйверах", которые экспрессируют рекомбиназу Cre , фермент, катализирующий рекомбинацию между двумя сайтами lox-P, в специфической подгруппе клеток, например, парвальбумин , экспрессирующих в интернейронах . При введении сконструированного вирусного вектора, содержащего ген оптогенетического актуатора, между двумя сайтами lox-P, только клетки, продуцирующие рекомбиназу Cre, будут экспрессировать микробный опсин. Этот метод позволил использовать несколько модифицированных оптогенетических актуаторов без необходимости создавать целую линию трансгенных животных каждый раз, когда необходим новый микробный опсин. [70]
После введения и экспрессии микробного опсина управляемый компьютером источник света должен быть оптически связан с рассматриваемой областью мозга. светоизлучающие диоды по оптоволокну (LED) или твердотельные лазеры с диодной накачкой Часто используются (DPSS). Недавние достижения включают появление беспроводных устройств, крепящихся на голову, которые применяют светодиоды к целевым областям и, как следствие, дают животным больше свободы передвижения. [71] [72]
Подходы на основе волокон также могут использоваться для объединения оптической стимуляции и визуализации кальция . [65] Это позволяет исследователям визуализировать и манипулировать активностью отдельных нейронов у бодрствующих животных. [73] Также возможна одновременная запись из нескольких глубоких областей мозга с использованием линз GRIN , подключенных через оптическое волокно к внешнему фотодетектору и фотостимулятору. [74] [75]
Технические проблемы
[ редактировать ]Выборочное выражение
[ редактировать ]Одна из основных проблем оптогенетики заключается в том, что не все рассматриваемые клетки могут экспрессировать ген микробного опсина на одном и том же уровне. Таким образом, даже освещение с определенной интенсивностью света будет оказывать различное воздействие на отдельные клетки. Оптогенетическая стимуляция нейронов головного мозга контролируется еще меньше, поскольку интенсивность света от источника света (например, имплантированного оптического волокна) падает экспоненциально.
Остается трудным направить опсин в определенные субклеточные компартменты, например, в плазматическую мембрану, синаптические везикулы или митохондрии. [51] [76] Ограничение опсина определенными областями плазматической мембраны, такими как дендриты , соматы или окончания аксонов, обеспечивает более четкое понимание нейронных цепей. [76]
Математическое моделирование показывает, что избирательная экспрессия опсина в определенных типах клеток может радикально изменить динамическое поведение нейронных цепей. В частности, оптогенетическая стимуляция, преимущественно нацеленная на ингибирующие клетки, может трансформировать возбудимость нервной ткани, влияя также на нетрансфицированные нейроны. [77]
Кинетика и синхронизация
[ редактировать ]Исходный канал родопсин-2 закрывался медленнее, чем типичные катионные каналы корковых нейронов, что приводило к длительной деполяризации и притоку кальция. [78] С тех пор было создано множество вариантов канального родопсина с более благоприятной кинетикой. [55] [56]
Разница между естественными паттернами спайков и оптогенетической активацией заключается в том, что импульсная световая стимуляция вызывает синхронную активацию экспрессирующих нейронов, что исключает возможность последовательной активности в стимулируемой популяции. Следовательно, трудно понять, как клетки в пораженной популяции общаются друг с другом или как их фазовые свойства активации связаны с функцией цепи.
Оптогенетическая активация была объединена с функциональной магнитно-резонансной томографией (МРТ) для выяснения коннектома — подробной карты нейронных связей мозга. [76] [79] Точно рассчитанная по времени оптогенетическая активация используется для калибровки задержанного гемодинамического сигнала ( ЖИРНЫЙ ), на котором основана фМРТ.
Спектр поглощения света
[ редактировать ]Белки опсины, используемые в настоящее время, имеют пики поглощения во всем видимом спектре, но остаются весьма чувствительными к синему свету. [76] Такое спектральное перекрытие очень затрудняет совмещение активации опсина с генетически закодированными индикаторами ( GEVIs , GECIs , GluSnFR , синапто-флуорин ), большинство из которых нуждаются в возбуждении синим светом. Опсины с инфракрасной активацией при стандартном значении освещенности увеличат проникновение света и повысят разрешение за счет уменьшения рассеяния света.
Пространственный отклик
[ редактировать ]Из-за рассеяния узкий луч света, стимулирующий нейроны в участке нервной ткани, может вызвать профиль ответа, который намного шире, чем луч стимуляции. [80] В этом случае нейроны могут активироваться (или тормозиться) непреднамеренно. Инструменты вычислительного моделирования [81] [82] используются для оценки объема стимулируемой ткани для различных длин волн света.
Приложения
[ редактировать ]Область оптогенетики продвинула фундаментальное научное понимание того, как определенные типы клеток способствуют функционированию биологических тканей, таких как нервные цепи, in vivo . С клинической точки зрения исследования, основанные на оптогенетике, привели к пониманию возможности восстановления с помощью света [1] . [83] болезнь Паркинсона [84] [85] и другие неврологические и психиатрические расстройства, такие как аутизм , шизофрения , злоупотребление наркотиками , тревога и депрессия . [52] [86] [87] [88] Экспериментальное лечение слепоты включает в себя канальный родопсин, экспрессируемый в ганглиозных клетках , который стимулируется световыми узорами из специально разработанных очков. [89] [9]
Идентификация конкретных нейронов и сетей
[ редактировать ]Миндалевидное тело
[ редактировать ]Оптогенетические подходы были использованы для картирования нейронных цепей в миндалевидном теле , которые способствуют обусловлению страха . [90] [91] [92] [93] Одним из таких примеров нейронной цепи является соединение базолатеральной миндалевидного тела с дорсо-медиальной префронтальной корой, где нейрональные колебания наблюдались частотой 4 Гц, коррелирующие с поведением замирания, вызванным страхом, у мышей. Трансгенным мышам вводили каналродопозин-2, присоединенный к промотору парвальбумина -Cre, который избирательно инфицировал интернейроны, расположенные как в базолатеральной миндалине, так и в дорсально-медиальной префронтальной коре, ответственной за колебания частотой 4 Гц. Интернейроны подвергались оптической стимуляции, вызывая замирание, и в результате были получены доказательства того, что эти колебания частотой 4 Гц могут быть ответственны за базовую реакцию страха, вызываемую популяциями нейронов дорсально-медиальной префронтальной коры и базолатеральной миндалины. [94]
Обонятельная луковица
[ редактировать ]Оптогенетическая активация обонятельных сенсорных нейронов имела решающее значение для демонстрации времени обработки запаха. [95] и для механизма нейромодуляционно-опосредованного обонятельного поведения (например, агрессия , спаривание ) [96] Кроме того, с помощью оптогенетики были воспроизведены данные, показывающие, что «остаточное изображение» запахов концентрируется более центрально вокруг обонятельной луковицы, а не на периферии, где должны располагаться нейроны обонятельных рецепторов. Трансгенных мышей, инфицированных канальным родопсином Thy1-ChR2, стимулировали лазером с длиной волны 473 нм, транскраниально расположенным над дорсальной частью обонятельной луковицы. Более длительная фотостимуляция митральных клеток в обонятельной луковице привела к наблюдениям более продолжительной активности нейронов в этой области после прекращения фотостимуляции, а это означает, что обонятельная сенсорная система способна претерпевать долгосрочные изменения и распознавать различия между старыми и новыми запахами. [97]
Прилежащее ядро
[ редактировать ]Оптогенетика, свободно перемещающееся поведение млекопитающих, электрофизиология in vivo и физиология срезов были интегрированы для исследования холинергических интернейронов прилежащего ядра путем прямого возбуждения или торможения. Несмотря на то, что эти холинергические клетки составляют менее 1% от общей популяции прилежащих нейронов, они способны контролировать активность дофаминергических терминалей, которые иннервируют средние шипиковые нейроны (MSN) в прилежащем ядре. [98] Известно, что эти прилежащие MSN участвуют в нервных путях, через которые кокаин оказывает свое воздействие, поскольку было показано, что уменьшение вызванных кокаином изменений в активности этих нейронов ингибирует кондиционирование кокаина . Немногочисленные холинергические нейроны, присутствующие в прилежащем ядре, могут оказаться жизнеспособными мишенями для фармакотерапии при лечении кокаиновой зависимости. [52]
Префронтальная кора
[ редактировать ]
Записи in vivo и in vitro из Университета Колорадо, Лаборатории оптофизиологии Боулдера, доктора философии Дональда К. Купера. экспрессирующие CAMKII AAV-ChR2, показаны отдельные пирамидальные нейроны, в префронтальной коре головного мозга, которые продемонстрировали высокую точность вывода потенциала действия с помощью коротких импульсов синего света с частотой 20 Гц ( рис. 1 ). [44]
Моторная кора
Повторная оптогенетическая стимуляция in vivo у здоровых животных могла в конечном итоге вызвать судороги. [99] Эта модель получила название оптокиндлинг.
Грушевидная кора
Повторная оптогенетическая стимуляция пирамидных клеток грушевидной коры in vivo у здоровых животных могла в конечном итоге вызвать судороги. [100] Исследования in vitro выявили потерю торможения по обратной связи в грушевидной цепи из-за нарушения синтеза ГАМК. [100]
Сердце
[ редактировать ]Оптогенетика была применена к предсердным кардиомиоцитам, чтобы остановить спирально-волновые аритмии , возникающие при фибрилляции предсердий , с помощью света. [101] Этот метод все еще находится на стадии разработки. Недавнее исследование изучило возможности оптогенетики как метода коррекции аритмий и ресинхронизации кардиостимуляции. В ходе исследования каналродопсин-2 был введен в кардиомиоциты желудочковых областей сердца трансгенных мышей и проведены in vitro исследования фотостимуляции на мышах как с открытой, так и с закрытой полостью. Фотостимуляция привела к усилению активации клеток и, таким образом, к усилению сокращений желудочков, что привело к увеличению частоты сердечных сокращений. Кроме того, этот подход был применен в сердечной ресинхронизирующей терапии ( СРТ ) в качестве нового биологического водителя ритма вместо электродной ЭЛТ. [102] В последнее время оптогенетика стала применяться в сердце для дефибрилляции желудочковых аритмий с локальным эпикардиальным освещением. [103] генерализованное освещение всего сердца [104] или с индивидуальными схемами стимуляции, основанными на аритмогенных механизмах, для снижения энергии дефибрилляции. [105]
Спиральный ганглий
[ редактировать ]Оптогенетическая стимуляция спирального ганглия у глухих мышей восстановила слуховую активность. [106] Оптогенетическое применение в области улитки позволяет стимулировать или ингибировать клетки спиральных ганглиев (SGN). Кроме того, из-за особенностей потенциалов покоя SGN использовались различные варианты белка каналородопсин-2, такие как Chronos, [107] CatCh и f-Chrimson. [108] Варианты Chronos и CatCh особенно полезны тем, что они проводят меньше времени в деактивированном состоянии, что позволяет осуществлять большую активность с меньшим количеством излучаемых вспышек синего света. Кроме того, использование специально разработанных каналов с красным смещением, таких как f-Chrimson, позволяет проводить стимуляцию с использованием более длинных волн, что снижает потенциальный риск фототоксичности в долгосрочной перспективе без ущерба для скорости стробирования. [109] В результате светодиод, излучающий свет, потребует меньше энергии, и идея кохлеарного протезирования в сочетании с фотостимуляцией станет более осуществимой. [110]
Мозговой ствол
[ редактировать ]Оптогенетическая стимуляция модифицированного родопсина, возбудимого красным светом (ReaChR), экспрессируемого в двигательном ядре лица, позволила минимально инвазивно активировать мотонейроны , эффективные в управлении движениями усов у мышей. [111] В одном новом исследовании оптогенетика использовалась на ядре дорсального шва для активации и ингибирования высвобождения дофаминергического вещества в вентральной области покрышки. Для активации трансгенных мышей инфицировали каналродопсином-2 с промотором TH-Cre, а для осуществления ингибирования гиперполяризующий к промотору TH-Cre добавляли опсин NpHR. Результаты показали, что оптическая активация дофаминергических нейронов приводила к увеличению социальных взаимодействий, а их торможение уменьшало потребность в общении только после периода изоляции. [112]
Визуальная система
[ редактировать ]Изучение зрительной системы с помощью оптогенетики может оказаться сложной задачей. Действительно, свет, используемый для оптогенетического контроля, может приводить к активации фоторецепторов в результате близости между первичными зрительными цепями и этими фоторецепторами. В этом случае трудно добиться пространственной избирательности (особенно в случае оптической доли мухи). Таким образом, исследование зрительной системы требует спектрального разделения с использованием каналов , которые активируются длинами волн света, отличными от родопсинов, внутри фоторецепторов (пиковая активация при 480 нм для родопсина 1 у дрозофилы ). Красносмещенный CsChrimson [113] или бистабильный каналродопсин [114] используются для оптогенетической активации нейронов (т.е. деполяризации ), поскольку оба позволяют спектральное разделение. Чтобы добиться молчания нейронов (т.е. гиперполяризации был обнаружен анионный канал родопсин . ), в криптофитных водорослях вида Guillardia theta (названных GtACR1) [115] можно использовать. GtACR1 более чувствителен к свету, чем другие ингибирующие каналы, такие как хлоридные насосы класса галородопсина, и придает сильную проводимость. Поскольку пик его активации (515 нм) близок к пику активации Родопсина 1, необходимо тщательно калибровать оптогенетическое освещение, а также зрительный стимул. Факторами, которые следует учитывать, являются длина волны оптогенетического освещения (возможно, выше, чем пик активации GtACR1), размер стимула (во избежание активации каналов светом стимула) и интенсивность оптогенетического освещения. освещение. Было показано, что GtACR1 может быть полезным ингибирующим инструментом в оптогенетическом исследовании зрительной системы дрозофилы путем подавления экспрессии нейронов T4/T5. [116] Эти исследования также можно проводить на интактных животных с хорошим поведением, например, для изучения оптомоторной реакции .
Сенсомоторная система
[ редактировать ]Оптогенетическое торможение или активация нейронов проверяет их необходимость и достаточность соответственно для формирования поведения. [117] Используя этот подход, исследователи могут проанализировать нейронную схему, контролирующую двигательную активность. Возмущая нейроны в различных местах сенсомоторной системы, исследователи узнали о роли нисходящих нейронов в возникновении стереотипного поведения. [118] как локализован тактильный сенсорный ввод [119] и активность интернейронов [120] изменяет локомоцию и роль клеток Пуркинье в генерации и модуляции движения. [121] Это мощный метод для более широкого понимания нейронных основ передвижения и движения животных .
Точный временной контроль вмешательств
[ редактировать ]Доступные в настоящее время оптогенетические приводы позволяют осуществлять точный временной контроль необходимого вмешательства (т.е. торможения или возбуждения целевых нейронов) с точностью, обычно доходящей до миллисекундного уровня. [122] Однако временная точность варьируется в зависимости от оптогенетических актуаторов. [123] и зависит от частоты и интенсивности стимуляции. [80]
Теперь можно разработать эксперименты, в которых свет, используемый для вмешательства, инициируется определенным элементом поведения (чтобы подавить поведение), определенным безусловным стимулом (чтобы связать что-то с этим стимулом) или конкретным колебательным событием в мозге (чтобы подавить событие). [124] [125] Подобный подход уже использовался в нескольких областях мозга:
Гиппокамп
[ редактировать ]Острые волны и комплексы пульсаций (КСВ) представляют собой отдельные высокочастотные колебательные явления в гиппокампе, которые, как полагают, играют роль в формировании и консолидации памяти. Эти события можно легко обнаружить, проследив за колебательными циклами записанного в режиме онлайн локального потенциала поля . Таким образом, начало события можно использовать в качестве триггерного сигнала для световой вспышки, которая направляется обратно в гиппокамп для торможения нейронов, особенно во время КСВ, а также для оптогенетического подавления самих колебаний. [126] Подобные эксперименты с «замкнутым контуром» полезны для изучения комплексов SWR и их роли в памяти.
Клеточная биология/клеточные сигнальные пути
[ редактировать ]Аналогично тому, как естественные светозависимые ионные каналы, такие как каналродопсин-2, обеспечивают оптический контроль потока ионов, что особенно полезно в нейронауке, естественные светорегулируемые белки передачи сигналов также обеспечивают оптический контроль биохимических путей, включая как генерацию вторых посланников, так и белок-белковые взаимодействия, что особенно полезно при изучении клеточной биологии и биологии развития. [128] В 2002 году был продемонстрирован первый пример использования фотобелков другого организма для контроля биохимического пути с использованием светоиндуцированного взаимодействия между растительным фитохромом и фактором, взаимодействующим с фитохромом (PIF), для контроля транскрипции генов у дрожжей. [1] Путем слияния фитохрома с ДНК-связывающим доменом и PIF с доменом активации транскрипции, активация транскрипции генов, распознаваемых ДНК-связывающим доменом, может быть индуцирована светом. [1] Это исследование предвосхитило аспекты более позднего развития оптогенетики в мозге, например, предположив, что «направленная доставка света с помощью оптоволокна потенциально может воздействовать на выбранные клетки или ткани даже внутри более крупных и непрозрачных организмов». [1] В литературе противоречивы относительно того, следует ли включать контроль клеточной биохимии с помощью фотопротеинов в определение оптогенетики, поскольку в обычном использовании оптогенетика относится конкретно к контролю возбуждения нейронов с помощью опсинов. [129] [130] [17] [131] и как контроль возбуждения нейронов с помощью опсинов, постдатируется и использует механизмы, отличные от контроля клеточной биохимии с помощью фотопротеинов. [128]
Фоточувствительные белки, используемые в различных сигнальных путях клеток
[ редактировать ]В дополнение к фитохромам, которые обнаружены в растениях и цианобактериях, домены LOV ( домен свето-кислородного напряжения ) растений и дрожжей и криптохромные домены растений представляют собой другие природные фотосенсорные домены, которые используются для оптического контроля биохимических путей в организме. клетки. [132] [128] Кроме того, на основе флуоресцентного белка Dronpa был создан синтетический фотосенсорный домен для оптического контроля биохимических путей. [128] В фотосенсорных доменах поглощение света связано либо с изменением белок-белковых взаимодействий (в случае фитохромов, некоторых LOV-доменов, криптохромов и мутантов Dronpa), либо с конформационным изменением, которое обнажает связанный сегмент белка или изменяет активность связанный белковый домен (в случае фитохромов и некоторых LOV-доменов). [128] Затем белок-белковые взаимодействия, регулируемые светом, можно использовать для рекрутирования белков в ДНК, например, для индукции транскрипции генов или модификаций ДНК, или в плазматическую мембрану, например, для активации резидентных сигнальных белков. [127] [133] [134] [135] [136] [137] CRY2 также кластеризуется, когда активен, поэтому он был слит с сигнальными доменами и впоследствии фотоактивирован, чтобы обеспечить активацию на основе кластеризации. [138] Домен LOV2 Avena sativa (овсяный обыкновенный) использовался для светозависимого воздействия на короткие пептиды или активный белковый домен. [139] [140] [141] Введение этого домена LOV в другой белок может регулировать функцию посредством пептидного нарушения, индуцированного светом. [142] Белок asLOV2, который оптогенетически экспонирует пептид, также использовался в качестве каркаса для нескольких синтетических систем светоиндуцированной димеризации и светоиндуцированной диссоциации (iLID и LOVTRAP соответственно). [143] [144] Системы можно использовать для контроля белков с помощью стратегии расщепления белков. [145] Фотодиссоциируемые домены Dronpa также использовались для захвата активного сайта белка в темноте, освобождения его после освещения голубым светом и повторного захвата после освещения фиолетовым светом. [146]
Временной контроль передачи сигнала светом
[ редактировать ]Способность оптически управлять сигналами в течение различной продолжительности времени исследуется, чтобы выяснить, как клеточные сигнальные пути преобразуют длительность сигнала и реакцию на различные выходные сигналы. [147] Естественные сигнальные каскады способны реагировать разными выходами на различия во времени и динамике стимула. [148] Например, обработка клеток PC12 эпидермальным фактором роста (EGF, индуцирующим временный профиль активности ERK) приводит к клеточной пролиферации, тогда как введение фактора роста нервов (NGF, индуцирующего устойчивый профиль активности ERK) приводит к дифференцировке в нейроноподобные клетки. . [149] Первоначально такое поведение было охарактеризовано с использованием приложений EGF и NGF, но результаты были частично воспроизведены с помощью оптических входов. [150] Кроме того, с помощью пульсирующей активации фотопереключаемого RAF, сконструированного с помощью фотодиссоциируемых доменов Dronpa, была обнаружена быстрая петля отрицательной обратной связи в пути RAF-MEK-ERK. [146]
Оптогенетическая шумо-фотостимуляция
[ редактировать ]Исследовательская группа профессора Элиаса Манхарреса представила оптогенетическую шумовую фотостимуляцию. [151] [152] [153] Это метод, использующий случайный шумный свет для активации нейронов, экспрессирующих ChR2. Оптимальный уровень опто-шумовой фотостимуляции головного мозга позволяет повысить потенциалы соматосенсорных вызванных полей, частотную характеристику импульсных нейронов пирамидных нейронов на соматосенсорную стимуляцию и амплитуду натриевого тока.
Награды
[ редактировать ]Мощное влияние оптогенетических технологий на исследования мозга было отмечено многочисленными наградами, присужденными ключевым игрокам в этой области.
В 2010 году Георг Нагель, Петер Хегеманн и Эрнст Бамберг были удостоены премии Уайли в области биомедицинских наук. [154] и они также были среди награжденных премией Карла Хайнца Беккурта в 2010 году. [155] В том же году Карл Дейссерот был награжден первой премией HFSP Nakasone за «его новаторскую работу по разработке оптогенетических методов изучения функции нейронных сетей, лежащих в основе поведения». [156]
В 2012 году Бамберг, Дайссерот, Хегеманн и Георг Нагель были удостоены премии Цульха от Общества Макса Планка . [157] а Мизенбёк был награжден премией Байе Латура в области здравоохранения за «новаторство в оптогенетических подходах к манипулированию активностью нейронов и контролю над поведением животных». [158]
В 2013 году Георг Нагель и Хегеманн были среди награжденных премией Луи-Жанте в области медицины . [159] В том же году Бамберг, Бойден, Дейссерот, Хегеманн, Мизенбёк и Георг Нагель были совместно удостоены премии «Мозг» за «изобретение и усовершенствование оптогенетики». [160] [161]
В 2017 году Дейссерот был награжден исследовательской премией Эльзы Крёнер Фрезениус «за открытия в оптогенетике и химии гидрогелевых тканей, а также за исследования нейронных цепей, лежащих в основе депрессии». [162]
В 2018 году Фонд Инамори вручил Дейссероту Киотскую премию за «лидерство в оптогенетике» и «революционные исследования в области системной нейробиологии». [163]
В 2019 году Бамберг, Бойден, Дайссерот, Хегеманн, Мизенбёк и Георг Нагель были награждены премией Румфорда Американской академии искусств и наук в знак признания «их выдающегося вклада, связанного с изобретением и усовершенствованием оптогенетики». [164]
В 2020 году Дейссерот был награжден премией Heineken в области медицины от Королевской Нидерландской академии искусств и наук за разработку оптогенетики и химии гидрогелевых тканей. [165]
В 2020 году Мизенбёк, Хегеманн и Георг Нагель совместно получили премию Шоу в области наук о жизни и медицины. [166]
В 2021 году Хегеманн, Дейсерот и Дитер Остерхельт получили премию Альберта Ласкера за фундаментальные медицинские исследования .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Симидзу-Сато С., Хук Э., Тепперман Дж.М., Куэйл П.Х. (октябрь 2002 г.). «Система промотора генов с переключателем света». Природная биотехнология . 20 (10): 1041–1044. дои : 10.1038/nbt734 . ПМИД 12219076 . S2CID 24914960 .
- ^ Го З.В., Ли Н., Хубер Д., Офир Э., Гутниски Д., Тинг Дж.Т. и др. (январь 2014 г.). «Поток корковой активности, лежащий в основе тактильного решения у мышей» . Нейрон . 81 (1): 179–194. дои : 10.1016/j.neuron.2013.10.020 . ПМЦ 3984938 . ПМИД 24361077 .
- ^ Лак А., Окунь М., Мосс М.М., Гурнани Х., Фаррелл К., Уэллс М.Дж. и др. (февраль 2020 г.). «Дофаминергическая и префронтальная основа обучения на основе сенсорной уверенности и ценности вознаграждения» . Нейрон . 105 (4): 700–711.e6. дои : 10.1016/j.neuron.2019.11.018 . ПМК 7031700 . ПМИД 31859030 .
- ^ Лю X, Рамирес С., Панг П.Т., Пурье CB, Говиндараджан А., Дейссерот К., Тонегава С. (март 2012 г.). «Оптогенетическая стимуляция энграммы гиппокампа активирует вызов воспоминаний о страхе» . Природа . 484 (7394): 381–385. Бибкод : 2012Natur.484..381L . дои : 10.1038/nature11028 . ПМК 3331914 . ПМИД 22441246 .
- ^ Танака Р., Хигучи Т., Кохацу С., Сато К., Ямамото Д. (ноябрь 2017 г.). «Оптогенетическая активация бесплодной меченной цепи у самцов Drosophila subobscura вызывает моторные акты спаривания» . Журнал неврологии . 37 (48): 11662–11674. doi : 10.1523/JNEUROSCI.1943-17.2017 . ПМК 6705751 . ПМИД 29109241 .
- ^ Стаматакис А.М., Стубер Г.Д. (ноябрь 2012 г.). «Оптогенетические стратегии анализа нейронных цепей, лежащих в основе вознаграждения и зависимости» . Перспективы Колд-Спринг-Харбора в медицине . 2 (11): а011924. doi : 10.1101/cshperspect.a011924 . ПМК 3543095 . ПМИД 23043156 .
- ^ Муссо, Пьер-Ив; Жунка, Пьер; Джелен, Меган; Фельдман-Кисс, Дамиан; Чжан, Хан; Чан, Рэйчел CW; Гордон, Майкл Д. (19 июля 2019 г.). Рамасвами, Мани; Дюлак, Кэтрин (ред.). «Замкнутая оптогенетическая активация периферических или центральных нейронов модулирует питание свободно движущихся дрозофил» . электронная жизнь . 8 : е45636. дои : 10.7554/eLife.45636 . ISSN 2050-084X . ПМК 6668987 . ПМИД 31322499 .
- ^ Фэн, Кай; Сен, Раджьяшри; Минэгиси, Ре; Дюбберт, Майкл; Бокемюль, Тилль; Бюшгес, Ансгар; Диксон, Барри Дж. (2 декабря 2020 г.). «Распределенное управление двигательными цепями при ходьбе назад у дрозофилы» . Природные коммуникации . 11 (1): 6166. Бибкод : 2020NatCo..11.6166F . дои : 10.1038/s41467-020-19936-x . ISSN 2041-1723 . ПМК 7710706 . PMID 33268800 . S2CID 227255627 .
- ^ Перейти обратно: а б с Сахель Дж.А., Буланже-Сцемама Е., Пагот С., Арлео А., Галлуппи Ф., Мартель Дж.Н. и др. (июль 2021 г.). «Частичное восстановление зрительных функций у слепого пациента после оптогенетической терапии» . Природная медицина . 27 (7): 1223–1229. дои : 10.1038/s41591-021-01351-4 . ПМИД 34031601 .
- ^ Лим, Диана; Ледью, Джеффри; Мохаджерани, Маджид; Ванни, Матье; Мерфи, Тимоти (2013). «Оптогенетические подходы к функциональному картированию мозга мышей» . Границы в неврологии . 7:54 . дои : 10.3389/fnins.2013.00054 . ISSN 1662-453X . ПМК 3622058 . ПМИД 23596383 .
- ^ Ли, Кэндис; Лавуа, Андреанна; Лю, Цзяшу; Чен, Саймон X.; Лю, Бао-хуа (2020). «Осветите мозг: применение оптогенетики при специфическом вскрытии цепей мозга мышей» . Границы в нейронных цепях . 14:18 . doi : 10.3389/fncir.2020.00018 . ISSN 1662-5110 . ПМЦ 7193678 . ПМИД 32390806 .
- ^ Франконвиль, Ромен; Берон, Селия; Джаяраман, Вивек (20 августа 2018 г.). Виджай Рагхаван, К; Скотт, Кристин; Хайнце, Стэнли (ред.). «Построение функционального коннектома центрального комплекса дрозофилы» . электронная жизнь . 7 : e37017. doi : 10.7554/eLife.37017 . ISSN 2050-084X . ПМК 6150698 . ПМИД 30124430 .
- ^ Чен, Чэнхао; Агравал, Света; Марк, Брэндон; Мамия, Акира; Сустар, Энн; Фелпс, Джаспер С.; Ли, Вэй-Чунг Аллен; Диксон, Барри Дж.; Кард, Гвинет М.; Тутхилл, Джон К. (06 декабря 2021 г.). «Функциональная архитектура нейронных цепей проприоцепции ног у дрозофилы» . Современная биология . 31 (23): 5163–5175.e7. дои : 10.1016/j.cub.2021.09.035 . ISSN 0960-9822 . ПМЦ 8665017 . ПМИД 34637749 .
- ^ Букварь по оптогенетике: Пастрана Э (2010). «Оптогенетика: управление функцией клеток с помощью света». Природные методы . 8 (1): 24–25. дои : 10.1038/nmeth.f.323 . S2CID 5808517 .
Редакция: «Метод года 2010» . Природные методы . 8 (1): 1. 2010. doi : 10.1038/nmeth.f.321 .
Комментарий: Дейсерот К. (январь 2011 г.). «Оптогенетика» . Природные методы . 8 (1): 26–29. дои : 10.1038/nmeth.f.324 . ПМК 6814250 . ПМИД 21191368 . - ^ Сотрудники новостей (декабрь 2010 г.). «Идеи десятилетия. Отойдя от деревьев, чтобы взглянуть на лес. Введение». Наука . 330 (6011): 1612–1613. Бибкод : 2010Sci...330.1612. . дои : 10.1126/science.330.6011.1612 . ПМИД 21163985 . S2CID 206593135 .
- ^ «Метод года 2010: Оптогенетика» . Видео о природе . 17 декабря 2010 г.
- ^ Перейти обратно: а б Дейсерот К. (20 октября 2010 г.). «Оптогенетика: управление мозгом с помощью света» . Научный американец . Спрингер Натура Америка, Инк.
- ^ Крик Ф (декабрь 1999 г.). «Влияние молекулярной биологии на нейробиологию» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 354 (1392): 2021–2025 гг. дои : 10.1098/rstb.1999.0541 . ПМК 1692710 . ПМИД 10670022 .
- ^ Хоффманн А., Хильдебрандт В., Хеберле Дж., Бюльдт Г. (сентябрь 1994 г.). «Фотоактивные митохондрии: перенос светового протонного насоса in vivo во внутреннюю митохондриальную мембрану Schizosaccharomyces pombe» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (20): 9367–9371. Бибкод : 1994PNAS...91.9367H . дои : 10.1073/pnas.91.20.9367 . ПМК 44813 . ПМИД 7937771 .
- ^ Земельман Б.В., Ли Г.А., Нг М., Мизенбёк Г. (январь 2002 г.). «Селективная фотостимуляция генетически заряженных нейронов» . Нейрон . 33 (1): 15–22. дои : 10.1016/S0896-6273(01)00574-8 . ПМИД 11779476 . S2CID 16391269 .
- ^ Земельман Б.В., Неснас Н., Ли Г.А., Мизенбок Г. (февраль 2003 г.). «Фотохимическое ворота гетерологичных ионных каналов: дистанционный контроль над генетически обусловленными популяциями нейронов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (3): 1352–1357. Бибкод : 2003PNAS..100.1352Z . дои : 10.1073/pnas.242738899 . ПМК 298776 . ПМИД 12540832 .
- ^ Бангхарт М., Борхес К., Исакофф Э., Траунер Д., Крамер Р.Х. (декабрь 2004 г.). «Светоактивируемые ионные каналы для дистанционного управления возбуждением нейронов» . Природная неврология . 7 (12): 1381–1386. дои : 10.1038/nn1356 . ПМЦ 1447674 . ПМИД 15558062 .
- ^ Волграф М., Горостиза П., Нумано Р., Крамер Р.Х., Исакофф Е.Ю., Траунер Д. (январь 2006 г.). «Аллостерический контроль ионотропного рецептора глутамата с помощью оптического переключателя» . Химическая биология природы . 2 (1): 47–52. дои : 10.1038/nchembio756 . ПМЦ 1447676 . ПМИД 16408092 .
- ^ Аренкель Б.Р., Кляйн М.Е., Дэвисон И.Г., Кац Л.К., Элерс, доктор медицинских наук (апрель 2008 г.). «Генетический контроль активности нейронов у мышей, условно экспрессирующих TRPV1» . Природные методы . 5 (4): 299–302. дои : 10.1038/nmeth.1190 . ПМК 3127246 . ПМИД 18327266 .
- ^ Гюлер А.Д., Рейнуотер А., Паркер Дж.Г., Джонс Г.Л., Аргилли Э., Аренкиель Б.Р. и др. (март 2012 г.). «Транзиторная активация специфических нейронов у мышей путем избирательной экспрессии рецептора капсаицина» . Природные коммуникации . 3 : 746. Бибкод : 2012NatCo...3..746G . дои : 10.1038/ncomms1749 . ПМК 3592340 . ПМИД 22434189 .
- ^ Ван М., Перова З., Аренкиель Б.Р., Ли Б (май 2014 г.). «Синаптические модификации медиальной префронтальной коры восприимчивости и устойчивости к стрессу» . Журнал неврологии . 34 (22): 7485–7492. doi : 10.1523/JNEUROSCI.5294-13.2014 . ПМК 4035514 . ПМИД 24872553 .
- ^ Перейти обратно: а б с Нагель Г., Селлас Т., Хун В., Катерия С., Адеишвили Н., Бертольд П. и др. (ноябрь 2003 г.). «Канал родопсин-2, катион-селективный мембранный канал с прямым светорегулированием» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (24): 13940–13945. Бибкод : 2003PNAS..10013940N . дои : 10.1073/pnas.1936192100 . ПМЦ 283525 . ПМИД 14615590 .
- ^ Гарц Х., Хегеманн П. (6 июня 1991 г.). «Токи кальция, регулируемые родопсином, у хламидомонады». Природа . 351 (6326): 489–491. Бибкод : 1991Natur.351..489H . дои : 10.1038/351489a0 . S2CID 4309593 .
- ^ Нагель Г., Оллиг Д., Фурманн М., Катерия С., Мусти А.М., Бамберг Э., Хегеманн П. (июнь 2002 г.). «Канал родопсин-1: светозапираемый протонный канал в зеленых водорослях». Наука . 296 (5577): 2395–2398. Бибкод : 2002Sci...296.2395N . дои : 10.1126/science.1072068 . ПМИД 12089443 . S2CID 206506942 .
- ^ Дейссерот К. (сентябрь 2015 г.). «Оптогенетика: 10 лет микробных опсинов в нейробиологии» . Природная неврология . 18 (9): 1213–1225. дои : 10.1038/nn.4091 . ПМЦ 4790845 . ПМИД 26308982 .
- ^ Бойден Э.С., Чжан Ф., Бамберг Э., Нагель Г., Дейссерот К. (сентябрь 2005 г.). «Генетически целенаправленный оптический контроль нейронной активности в миллисекундном масштабе». Природная неврология . 8 (9): 1263–1268. дои : 10.1038/nn1525 . ПМИД 16116447 . S2CID 6809511 .
- ^ «Возможно, он является законным изобретателем крупнейшего прорыва в нейробиологии за последние десятилетия. Но вы никогда о нем не слышали» . СТАТ . 1 сентября 2016 года . Проверено 9 февраля 2020 г.
- ^ Дейсерот, Карл (26 августа 2015 г.). «Оптогенетика: 10 лет микробных опсинов в нейробиологии» . Природная неврология . 18 (9): 1213–1225. дои : 10.1038/nn.4091 . ПМЦ 4790845 . ПМИД 26308982 .
- ^ Би А, Цуй Дж, Ма Ю.П., Ольшевская Е, Пу М, Дижоор А.М., Пан Ж. (апрель 2006 г.). «Эктопическая экспрессия родопсина микробного типа восстанавливает зрительные реакции у мышей с дегенерацией фоторецепторов» . Нейрон . 50 (1): 23–33. дои : 10.1016/j.neuron.2006.02.026 . ПМК 1459045 . ПМИД 16600853 .
- ^ Лима SQ, Мизенбёк Г (апрель 2005 г.). «Дистанционное управление поведением посредством генетически направленной фотостимуляции нейронов» . Клетка . 121 (1): 141–152. дои : 10.1016/j.cell.2005.02.004 . ПМИД 15820685 . S2CID 14608546 .
- ^ Перейти обратно: а б Ли Х, Гутьеррес Д.В., Хансон М.Г., Хан Дж., Марк М.Д., Чил Х. и др. (декабрь 2005 г.). «Быстрая неинвазивная активация и ингибирование нейронной и сетевой активности родопсином позвоночных и родопсином зеленых водорослей» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (49): 17816–17821. Бибкод : 2005PNAS..10217816L . дои : 10.1073/pnas.0509030102 . ПМК 1292990 . ПМИД 16306259 .
- ^ Нагель Г., Браунер М., Ливальд Дж. Ф., Адеишвили Н., Бамберг Э., Готшалк А. (декабрь 2005 г.). «Световая активация канала родопсина-2 в возбудимых клетках Caenorhabditis elegans вызывает быстрые поведенческие реакции» . Современная биология . 15 (24): 2279–2284. дои : 10.1016/j.cub.2005.11.032 . ПМИД 16360690 . S2CID 7036529 .
- ^ Цянь Дж.З., Чен Д.Ф., Гербер Д., Том С., Мерсер Э.Х., Андерсон Дж.Д. и др. (декабрь 1996 г.). «Нокаут генов, ограниченных по субрегионам и типам клеток, в мозгу мыши» . Клетка . 87 (7): 1317–1326. дои : 10.1016/S0092-8674(00)81826-7 . ПМИД 8980237 . S2CID 863399 .
- ^ Цянь Дж.З. (2016). «Нейрогенетика Cre-Lox: 20 лет универсального применения в исследованиях мозга и подсчете...» Границы генетики . 7:19 . дои : 10.3389/fgene.2016.00019 . ПМЦ 4759636 . ПМИД 26925095 .
- ^ Хан X, Бойден ES (март 2007 г.). «Многоцветная оптическая активация, подавление и десинхронизация нейронной активности с временным разрешением в один пик» . ПЛОС ОДИН . 2 (3). Публичная научная библиотека: e299. Бибкод : 2007PLoSO...2..299H . дои : 10.1371/journal.pone.0000299 . OCLC 678618519 . ПМК 1808431 . ПМИД 17375185 .
- ^ Чжан Ф., Ван Л.П., Браунер М., Лиевальд Дж.Ф., Кей К., Вацке Н. и др. (апрель 2007 г.). «Мультимодальный быстрый оптический опрос нейронных цепей». Природа . 446 (7136): 633–639. Бибкод : 2007Natur.446..633Z . дои : 10.1038/nature05744 . ПМИД 17410168 . S2CID 4415339 .
- ^ Шредер-Ланг С., Шварцель М., Зайферт Р., Стрюнкер Т., Катерия С., Лузер Дж. и др. (январь 2007 г.). «Быстрое манипулирование уровнем клеточного цАМФ с помощью света in vivo» . Природные методы . 4 (1): 39–42. дои : 10.1038/nmeth975 . ПМИД 17128267 . S2CID 10616442 .
- ^ Гао С., Нагпал Дж., Шнайдер М.В., Козьяк-Павлович В., Нагель Г., Готшалк А. (сентябрь 2015 г.). «Оптогенетическое манипулирование цГМФ в клетках и животных с помощью жестко регулируемого светом гуанилилциклазного опсина CyclOp» . Природные коммуникации . 6 (1): 8046. Бибкод : 2015NatCo...6.8046G . дои : 10.1038/ncomms9046 . ПМЦ 4569695 . ПМИД 26345128 .
- ^ Перейти обратно: а б с Баратта М.В., Накамура С., Добелис П., Помренце М.Б., Долзани С.Д., Купер, округ Колумбия (2 апреля 2012 г.). «Оптогенетический контроль активности генетически направленных пирамидных нейронов в префронтальной коре» (PDF) . Предшественники природы . arXiv : 1204.0710 . Бибкод : 2012arXiv1204.0710B . дои : 10.1038/npre.2012.7102.1 . S2CID 31641314 .
- ^ Хассон С.Дж., Ливальд Дж.Ф., Шультайс С., Стирман Дж.Н., Лу Х., Готшалк А. (2012). Сэмюэл А. (ред.). «Микробные протонные насосы, активируемые светом, как ингибиторы нейронов для функционального разделения нейронных сетей у C. elegans» . ПЛОС ОДИН . 7 (7): е40937. Бибкод : 2012PLoSO...740937H . дои : 10.1371/journal.pone.0040937 . ПМК 3397962 . ПМИД 22815873 .

- ^ Лю Ю, ЛеБаф Б, Го X, Корреа П.А., Гуальберто Д.Г., Линтс Р., Гарсия ЛР (март 2011 г.). Гудман М.Б. (ред.). «Холинергическая регулируемая схема координирует поддержание и бистабильные состояния сенсомоторного поведения во время совокупления самцов Caenorhabditis elegans» . ПЛОС Генетика . 7 (3): e1001326. дои : 10.1371/journal.pgen.1001326 . ПМК 3053324 . ПМИД 21423722 .
- ^ Дейссерот К. «Оптогенетика: управление мозгом с помощью света [Расширенная версия]» . Научный американец . Проверено 28 ноября 2016 г.
- ^ Бек С., Ю-Стшельчик Дж., Паулс Д., Константин О.М., Джи К.Э., Эманн Н. и др. (2018). «Синтетические светоактивируемые ионные каналы для оптогенетической активации и ингибирования» . Границы в неврологии . 12 : 643. дои : 10.3389/fnins.2018.00643 . ПМК 6176052 . ПМИД 30333716 .
- ^ Сьерра Я., Рост Б., Олдани С., Шнайдер-Варме Ф., Зайферт Р., Шмитц Д., Хегеманн П. (ноябрь 2018 г.). «Двухкомпонентный оптогенетический инструмент на основе калиевых каналов для подавления возбудимых клеток» . Биофизический журнал . 114 (3): 668а. Бибкод : 2018BpJ...114..668A . дои : 10.1016/j.bpj.2017.11.3607 . hdl : 21.11116/0000-0003-4AEF-E .
- ^ Чжао С., Кунья С., Чжан Ф., Лю К., Глосс Б., Дейссерот К. и др. (август 2008 г.). «Улучшенная экспрессия галородопсина для светоиндуцированного подавления активности нейронов» . Биология клеток мозга . 36 (1–4): 141–154. дои : 10.1007/s11068-008-9034-7 . ПМК 3057022 . ПМИД 18931914 .
- ^ Перейти обратно: а б Градинару В., Томпсон К.Р., Дейссерот К. (август 2008 г.). «eNpHR: галородопсин Natronomonas, улучшенный для оптогенетических применений» . Биология клеток мозга . 36 (1–4): 129–139. дои : 10.1007/s11068-008-9027-6 . ПМК 2588488 . ПМИД 18677566 .
- ^ Перейти обратно: а б с Виттен И.Б., Лин С.К., Бродский М., Пракаш Р., Дистер И., Аникеева П. и др. (декабрь 2010 г.). «Холинергические интернейроны контролируют активность локальных цепей и кондиционирование кокаина» . Наука . 330 (6011): 1677–1681. Бибкод : 2010Sci...330.1677W . дои : 10.1126/science.1193771 . ПМК 3142356 . ПМИД 21164015 .
- ^ Ким Дж.М., Хва Дж., Гаррига П., Ривз П.Дж., РаджБхандари У.Л., Корана Х.Г. (февраль 2005 г.). «Световая активация передачи сигналов бета-2-адренергического рецептора с помощью химерного родопсина, содержащего цитоплазматические петли бета-2-адренергического рецептора». Биохимия . 44 (7): 2284–2292. дои : 10.1021/bi048328i . ПМИД 15709741 .
- ^ Айран Р.Д., Томпсон К.Р., Фенно Л.Е., Бернштейн Х., Дейссерот К. (апрель 2009 г.). «Временно точный контроль внутриклеточной передачи сигналов in vivo». Природа . 458 (7241): 1025–1029. Бибкод : 2009Natur.458.1025A . дои : 10.1038/nature07926 . ПМИД 19295515 . S2CID 4401796 .
- ^ Левская А., Вайнер О.Д., Лим В.А., Фойгт К.А. (октябрь 2009 г.). «Пространственно-временной контроль клеточной передачи сигналов с использованием светопереключаемого взаимодействия белков» . Природа . 461 (7266): 997–1001. Бибкод : 2009Natur.461..997L . дои : 10.1038/nature08446 . ПМЦ 2989900 . ПМИД 19749742 .
- ^ Ву Й.И., Фрей Д., Лунгу О.И., Джариг А., Шлихтинг И. , Кульман Б., Хан К.М. (сентябрь 2009 г.). «Генетически закодированный фотоактивируемый Rac контролирует подвижность живых клеток» . Природа . 461 (7260): 104–108. Бибкод : 2009Natur.461..104W . дои : 10.1038/nature08241 . ПМЦ 2766670 . ПМИД 19693014 .
- ^ Ядзава М., Садагиани А.М., Сюэ Б., Долмеч Р.Э. (октябрь 2009 г.). «Индукция белок-белковых взаимодействий в живых клетках с помощью света». Природная биотехнология . 27 (10): 941–945. дои : 10.1038/nbt.1569 . ПМИД 19801976 . S2CID 205274357 .
- ^ Стирл М., Штумпф П., Удвари Д., Гета Р., Хагедорн Р., Лоси А. и др. (январь 2011 г.). «Световая модуляция клеточного цАМФ небольшой бактериальной фотоактивируемой аденилатциклазой, bPAC, почвенной бактерии Beggiatoa» . Журнал биологической химии . 286 (2): 1181–1188. дои : 10.1074/jbc.M110.185496 . ПМК 3020725 . ПМИД 21030594 .
- ^ Рю М.Х., Москвин О.В., Силтберг-Либерлес Ю., Гомельский М. (декабрь 2010 г.). «Природные и сконструированные фотоактивируемые нуклеотидилциклазы для оптогенетических применений» . Журнал биологической химии . 285 (53): 41501–41508. дои : 10.1074/jbc.M110.177600 . ПМК 3009876 . ПМИД 21030591 .
- ^ Лернер Т.Н., Й.Л., Дейссерот К. (март 2016 г.). «Коммуникация в нейронных цепях: инструменты, возможности и проблемы» . Клетка . 164 (6): 1136–1150. дои : 10.1016/j.cell.2016.02.027 . ПМЦ 5725393 . ПМИД 26967281 .
- ^ Араванис А.М., Ван Л.П., Чжан Ф., Мельцер Л.А., Могри М.З., Шнайдер М.Б., Дейссерот К. (сентябрь 2007 г.). «Оптический нейронный интерфейс: контроль моторной коры головного мозга грызунов in vivo с помощью интегрированной оптоволоконной и оптогенетической технологии». Журнал нейронной инженерии . 4 (3): С143–С156. Бибкод : 2007JNEng...4S.143A . дои : 10.1088/1741-2560/4/3/S02 . ПМИД 17873414 . S2CID 1488394 .
- ^ Адамантидис А.Р., Чжан Ф., Араванис А.М., Дейссерот К., де Лесеа Л. (ноябрь 2007 г.). «Нейронные субстраты пробуждения, исследованные при оптогенетическом контроле гипокретиновых нейронов» . Природа . 450 (7168): 420–424. Бибкод : 2007Natur.450..420A . дои : 10.1038/nature06310 . ПМЦ 6744371 . ПМИД 17943086 .
- ^ Перейти обратно: а б Градинару В., Томпсон К.Р., Чжан Ф., Могри М., Кей К., Шнайдер М.Б., Дейссерот К. (декабрь 2007 г.). «Стратегии нацеливания и считывания для быстрого оптического нейронного контроля in vitro и in vivo» . Журнал неврологии . 27 (52): 14231–14238. doi : 10.1523/JNEUROSCI.3578-07.2007 . ПМК 6673457 . ПМИД 18160630 .
- ^ Даместани Ю., Рейнольдс К.Л., Сзу Дж., Сюй М.С., Кодера Ю., Биндер Д.К. и др. (ноябрь 2013 г.). «Прозрачный нанокристаллический протез черепа, стабилизированный иттрием и цирконием» . Наномедицина . 9 (8): 1135–1138. дои : 10.1016/j.nano.2013.08.002 . ПМИД 23969102 . S2CID 14212180 . • Объяснено Мохан Дж. (4 сентября 2013 г.). «Окно в мозг? Оно здесь», — говорит команда Калифорнийского университета в Риверсайде . Лос-Анджелес Таймс .
- ^ Перейти обратно: а б Легария А.А., Матикайнен-Анкни Б.А., Ян Б., Аханону Б, Лихолаи Дж.А., Паркер Дж.Г., Кравиц А.В. (30 августа 2022 г.). «Волоконная фотометрия в полосатом теле отражает прежде всего несоматические изменения кальция» . Природная неврология . 25 (9): 1124–1128. дои : 10.1038/s41593-022-01152-z .
- ^ Венц К.Т., Бернштейн Дж.Г., Монахан П., Герра А., Родригес А., Бойден Э.С. (август 2011 г.). «Управляемое устройство с беспроводным питанием для оптического нейронного контроля свободно ведущих себя животных» . Журнал нейронной инженерии . 8 (4): 046021. Бибкод : 2011JNEng...8d6021W . дои : 10.1088/1741-2560/8/4/046021 . ПМК 3151576 . ПМИД 21701058 .
- ^ Пама Э.А., Колзато Л.С., Хоммель Б. (1 января 2013 г.). «Оптогенетика как инструмент нейромодуляции в когнитивной нейробиологии» . Границы в психологии . 4 : 610. doi : 10.3389/fpsyg.2013.00610 . ПМЦ 3764402 . ПМИД 24046763 .
- ^ Вирок, Йоханнес; Родригес-Розада, Сильвия; Дитер, Александр; Пипер, Флориан; Симс, Рут; Тенедини, Федерико; Бергс, Амели CF; Бендифаллах, Имане; Чжоу, Фанминь; Цейтшель, Надя; Альбек, Иоахим (26 июля 2021 г.). «BiPOLES — это оптогенетический инструмент, разработанный для двунаправленного двухцветного контроля нейронов» . Природные коммуникации . 12 (1): 4527. Бибкод : 2021NatCo..12.4527V . дои : 10.1038/s41467-021-24759-5 . ISSN 2041-1723 . ПМЦ 8313717 . ПМИД 34312384 .
- ^ Чжан Ф., Градинару В., Адамантидис А.Р., Дюран Р., Айран Р.Д., де Лесеа Л., Дейссерот К. (март 2010 г.). «Оптогенетический опрос нейронных цепей: технология исследования структур мозга млекопитающих» . Протоколы природы . 5 (3): 439–456. дои : 10.1038/nprot.2009.226 . ПМК 4503465 . ПМИД 20203662 .
- ^ Цзэн Х, Мэдисен Л (5 сентября 2012 г.). «Мышиные трансгенные подходы в оптогенетике». Оптогенетика: инструменты контроля и мониторинга активности нейронов . Прогресс в исследованиях мозга. Том. 196. стр. 193–213. дои : 10.1016/B978-0-444-59426-6.00010-0 . ISBN 9780444594266 . ПМЦ 3433654 . ПМИД 22341327 .
- ^ Надзиратель М.Р., Кардин Дж.А., Дейссерот К. (июль 2014 г.). «Оптические нейронные интерфейсы» . Ежегодный обзор биомедицинской инженерии . 16 : 103–129. doi : 10.1146/annurev-bioeng-071813-104733 . ПМК 4163158 . ПМИД 25014785 .
- ^ Гуру А, Пост Р.Дж., Хо Ю.Й., Надзиратель М.Р. (июль 2015 г.). «Понимание оптогенетики» . Международный журнал нейропсихофармакологии . 18 (11): pyv079. дои : 10.1093/ijnp/pyv079 . ПМЦ 4756725 . ПМИД 26209858 .
- ^ «Эволюция свободно действующих технологий визуализации и оптогенетики» . Имплантат ОАЗИС . Майтекс . Проверено 3 июня 2021 г.
- ^ Цуй Дж., Джун С.Б., Джин Икс, Луо Дж., Фам М.Д., Ловинджер Д.М. и др. (апрель 2016 г.). «Глубокие оптические измерения мозговой активности нейронной активности конкретного типа клеток у ведущих мышей» . Протоколы природы . 9 (6): 1213–1228. дои : 10.1038/nmeth.3770 . ПМК 4100551 . ПМИД 24784819 .
- ^ Цуй Дж., Джун С.Б., Джин Икс, Луо Дж., Фам М.Д., Ловинджер Д.М. и др. (июнь 2014 г.). «Глубокие оптические измерения мозговой активности нейронной активности конкретного типа клеток у ведущих мышей» . Протоколы природы . 9 (6): 1213–1228. дои : 10.1038/nprot.2014.080 . ПМК 4100551 . ПМИД 24784819 .
- ^ Перейти обратно: а б с д Залокусский К.А., Фенно Л.Е., Дейсерот К. (2013). «Актуальные проблемы оптогенетики» . Общество нейробиологии .
- ^ Хайтманн С., Рул М., Трукколо В., Эрментраут Б. (январь 2017 г.). «Оптогенетическая стимуляция сдвигает возбудимость коры головного мозга с типа I на тип II: возникновение колебаний и распространение волн» . PLOS Вычислительная биология . 13 (1): e1005349. Бибкод : 2017PLSCB..13E5349H . дои : 10.1371/journal.pcbi.1005349 . ПМЦ 5295702 . ПМИД 28118355 .
- ^ Чжан, Ян-Пин; Эртнер, Томас Г. (2007). «Оптическая индукция синаптической пластичности с помощью светочувствительного канала» . Природные методы . 4 (2): 139–141. дои : 10.1038/nmeth988 . ISSN 1548-7091 . ПМИД 17195846 . S2CID 17721823 .
- ^ Леергаард ТБ, Хильгетаг CC, Спорнс О (01 мая 2012 г.). «Картирование коннектома: многоуровневый анализ связей мозга» . Границы нейроинформатики . 6:14 . дои : 10.3389/fninf.2012.00014 . ПМК 3340894 . ПМИД 22557964 .
- ^ Перейти обратно: а б Любоеинский Ю., Чуматченко Т. (сентябрь 2020 г.). «Характеристики нелинейного ответа нейронных сетей и отдельных нейронов, подвергающихся оптогенетическому возбуждению» . Сетевая нейронаука . 4 (3): 852–870. дои : 10.1162/netn_a_00154 . ПМЦ 7888483 . ПМИД 33615093 .
- ^ «PyRhO: виртуальная лаборатория оптогенетики» . Гитхаб .
- ^ «Инструмент моделирования нейронных сетей и одиночных нейронов со светочувствительными каналами» . Гитхаб .
- ^ Азеес, Аджмал (декабрь 2023 г.). «Распространение активации и взаимодействие между каналами при многоканальной оптогенетической стимуляции в улитке мыши» . Исследование слуха . 440 . дои : 10.1016/j.heares.2023.108911 . ПМИД 37977051 .
- ^ Кравиц А.В., Фриз Б.С., Паркер П.Р., Кей К., Твин М.Т., Дейссерот К., Крейцер А.С. (июль 2010 г.). «Регуляция паркинсонического двигательного поведения посредством оптогенетического контроля цепей базальных ганглиев» . Природа . 466 (7306): 622–626. Бибкод : 2010Natur.466..622K . дои : 10.1038/nature09159 . ПМЦ 3552484 . ПМИД 20613723 .
- ^ Градинару В., Могри М., Томпсон К.Р., Хендерсон Дж.М., Дейссерот К. (апрель 2009 г.). «Оптическая деконструкция паркинсонических нейронных цепей» . Наука . 324 (5925): 354–359. Бибкод : 2009Sci...324..354G . CiteSeerX 10.1.1.368.668 . дои : 10.1126/science.1167093 . ПМК 6744370 . ПМИД 19299587 .
- ^ Карден Дж.А., Карлен М., Мелетис К., Кноблих У., Чжан Ф., Дейссерот К. и др. (июнь 2009 г.). «Управление быстродействующими клетками индуцирует гамма-ритм и контролирует сенсорные реакции» . Природа . 459 (7247): 663–667. Бибкод : 2009Natur.459..663C . дои : 10.1038/nature08002 . ПМЦ 3655711 . ПМИД 19396156 .
- ^ Сохал В.С., Чжан Ф., Ижар О., Дейссерот К. (июнь 2009 г.). «Нейроны парвальбумина и гамма-ритмы улучшают работу корковых цепей» . Природа . 459 (7247): 698–702. Бибкод : 2009Natur.459..698S . дои : 10.1038/nature07991 . ПМЦ 3969859 . ПМИД 19396159 .
- ^ Цай Х.К., Чжан Ф., Адамантидис А., Стубер Г.Д., Бончи А., де Лесеа Л., Дейссерот К. (май 2009 г.). «Фазовая активация дофаминергических нейронов достаточна для поведенческого кондиционирования» . Наука . 324 (5930): 1080–1084. Бибкод : 2009Sci...324.1080T . дои : 10.1126/science.1168878 . ПМК 5262197 . ПМИД 19389999 .
- ^ Циммер С (24 мая 2021 г.). «Ученые частично восстановили зрение слепому с помощью новой генной терапии» . Нью-Йорк Таймс . Проверено 25 мая 2021 г.
- ^ Хаубенсак В., Кунвар П.С., Кай Х., Чокки С., Уолл Н.Р., Поннусами Р. и др. (ноябрь 2010 г.). «Генетическое вскрытие микросхемы миндалевидного тела, управляющей обусловленным страхом» . Природа . 468 (7321): 270–276. Бибкод : 2010Natur.468..270H . дои : 10.1038/nature09553 . ПМК 3597095 . ПМИД 21068836 .
- ^ Йохансен Дж.П., Хаманака Х., Монфис М.Х., Бениа Р., Дейсерот К., Блэр Х.Т., Леду Дж.Е. (июль 2010 г.). «Оптическая активация латеральных пирамидных клеток миндалевидного тела стимулирует ассоциативное обучение страху» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (28): 12692–12697. Бибкод : 2010PNAS..10712692J . дои : 10.1073/pnas.1002418107 . ПМК 2906568 . ПМИД 20615999 .
- ^ Яснов А.М., Эрлих Д.Э., Чой Д.С., Дабровска Дж., Бауэрс М.Е., Маккалоу К.М. и др. (июнь 2013 г.). «Нейроны, экспрессирующие Thy1 в базолатеральной миндалине, могут опосредовать торможение страха» . Журнал неврологии . 33 (25): 10396–10404. doi : 10.1523/JNEUROSCI.5539-12.2013 . ПМЦ 3685835 . ПМИД 23785152 .
- ^ Диас Б.Г., Банерджи С.Б., Гудман СП, Ресслер К.Дж. (июнь 2013 г.). «К новым подходам к расстройствам страха и тревоги» . Современное мнение в нейробиологии . 23 (3): 346–352. дои : 10.1016/j.conb.2013.01.013 . ПМЦ 3672317 . ПМИД 23402950 .
- ^ Каралис Н., Дежан С., Чаудун Ф., Ходер С., Розеске Р.Р., Вурц Х. и др. (апрель 2016 г.). «Колебания 4 Гц синхронизируют префронтальные цепи миндалевидного тела во время поведения, вызванного страхом» . Природная неврология . 19 (4): 605–612. дои : 10.1038/nn.4251 . ПМЦ 4843971 . ПМИД 26878674 .
- ^ Шустерман Р., Смир М.С., Кулаков А.А., Ринберг Д. (июль 2011 г.). «Точные обонятельные реакции составляют цикл обнюхивания». Природная неврология . 14 (8): 1039–1044. дои : 10.1038/nn.2877 . ПМИД 21765422 . S2CID 5194595 .
- ^ Смит Р.С., Ху Р., ДеСоуза А., Эберли С.Л., Крахе К., Чан В., Аранеда Р.К. (июль 2015 г.). «Дифференциальная мускариновая модуляция в обонятельной луковице» . Журнал неврологии . 35 (30): 10773–10785. doi : 10.1523/JNEUROSCI.0099-15.2015 . ПМЦ 4518052 . ПМИД 26224860 .
- ^ Паттерсон М.А., Лагьер С., Карлтон А. (август 2013 г.). «Представления запаха в обонятельной луковице развиваются после первого вдоха и сохраняются как остаточное изображение запаха» . Труды Национальной академии наук Соединенных Штатов Америки . 110 (35): Е3340–Е3349. Бибкод : 2013PNAS..110E3340P . дои : 10.1073/pnas.1303873110 . ПМЦ 3761593 . ПМИД 23918364 .
- ^ Текуапетла Ф., Патель Дж.К., Ксенияс Х., Инглиш Д., Тадрос И., Шах Ф. и др. (май 2010 г.). «Глутаматергическая передача сигналов мезолимбическими дофаминовыми нейронами в прилежащем ядре» . Журнал неврологии . 30 (20): 7105–7110. doi : 10.1523/JNEUROSCI.0265-10.2010 . ПМЦ 3842465 . ПМИД 20484653 .
- ^ Села Э., Макфарлан А.Р., Чунг А.Дж., Ван Т., Кьерзи С., Мурай К.К., Сьёстрем П.Дж. (март 2019 г.). «Оптогенетическая модель Киндлинга неокортикальной эпилепсии» . Научные отчеты . 9 (1): 5236. Бибкод : 2019НатСР...9.5236С . дои : 10.1038/s41598-019-41533-2 . ПМК 6437216 . ПМИД 30918286 .
- ^ Перейти обратно: а б Рю Б., Нагаппан С., Сантос-Валенсия Ф., Ли П., Родригес Э., Лэки М. и др. (апрель 2021 г.). «Хроническая потеря торможения в грушевидной коре после кратковременной ежедневной оптогенетической стимуляции» . Отчеты по ячейкам . 35 (3): 109001. doi : 10.1016/j.celrep.2021.109001 . ПМК 8102022 . ПМИД 33882304 .
- ^ Бинген Б.О., Энгельс М.К., Шалидж М.Дж., Джангсангтонг В., Нешати З., Феола I и др. (октябрь 2014 г.). «Светоиндуцированное прекращение спирально-волновых аритмий методом оптогенетической инженерии предсердных кардиомиоцитов» . Сердечно-сосудистые исследования . 104 (1): 194–205. дои : 10.1093/cvr/cvu179 . ПМИД 25082848 .
- ^ Нусинович У, Гепштейн Л (июль 2015 г.). «Оптогенетика для кардиостимуляции и ресинхронизирующей терапии in vivo». Природная биотехнология . 33 (7): 750–754. дои : 10.1038/nbt.3268 . ПМИД 26098449 . S2CID 1794556 .
- ^ Нинс Э.К., Кип А., Барт С.И., Пломп Дж.Дж., Цеппенфельд К., Шалидж М.Дж. и др. (июль 2017 г.). «Оптогенетическое прекращение желудочковых аритмий во всем сердце: на пути к биологическому управлению сердечным ритмом» . Европейский кардиологический журнал . 38 (27): 2132–2136. doi : 10.1093/eurheartj/ehw574 . ПМЦ 5837774 . ПМИД 28011703 .
- ^ Брюгманн Т., Бойл П.М., Фогт CC, Каратанос Т.В., Аревало Х.Дж., Флейшманн Б.К. и др. (октябрь 2016 г.). «Оптогенетическая дефибрилляция прекращает желудочковую аритмию в сердцах мышей и на человеческих симуляциях» . Журнал клинических исследований . 126 (10): 3894–3904. дои : 10.1172/JCI88950 . ПМК 5096832 . ПМИД 27617859 .
- ^ Крочини С., Феррантини С., Коппини Р., Скардигли М., Ян П., Лоу Л.М. и др. (октябрь 2016 г.). «Оптогенетический дизайн паттернов механистической стимуляции для дефибрилляции сердца» . Научные отчеты . 6 : 35628. Бибкод : 2016NatSR...635628C . дои : 10.1038/srep35628 . ПМК 5066272 . ПМИД 27748433 .
- ^ Эрнандес В.Х., Герт А., Рейтер К., Цзин З., Йешке М., Мендоса Шульц А. и др. (март 2014 г.). «Оптогенетическая стимуляция слухового пути» . Журнал клинических исследований . 124 (3): 1114–1129. дои : 10.1172/JCI69050 . ПМЦ 3934189 . ПМИД 24509078 .
- ^ Кеппелер Д., Мерино Р.М., Лопес де ла Морена Д., Бали Б., Уэт А.Т., Герт А. и др. (декабрь 2018 г.). «Сверхбыстрая оптогенетическая стимуляция слухового пути с помощью оптимизированного по нацеливанию Chronos» . Журнал ЭМБО . 37 (24): e99649. дои : 10.15252/embj.201899649 . ПМК 6293277 . ПМИД 30396994 .
- ^ Магер Т., Лопес де ла Морена Д., Сенн В., Шлотте Дж., Эррико Д., Фельдбауэр К. и др. (май 2018 г.). «Высокочастотные нейронные импульсы и слуховые сигналы с помощью сверхбыстрой оптогенетики с красным смещением» . Природные коммуникации . 9 (1): 1750. Бибкод : 2018NatCo...9.1750M . дои : 10.1038/s41467-018-04146-3 . ПМЦ 5931537 . ПМИД 29717130 .
- ^ «Разработка длинноволновых ионных каналов, управляемых светом, чтобы слышать свет. Атлас науки» . Проверено 7 ноября 2019 г.
- ^ Мозер Т. (октябрь 2015 г.). «Оптогенетическая стимуляция слухового пути для исследований и будущего протезирования». Современное мнение в нейробиологии . 34 : 29–36. дои : 10.1016/j.conb.2015.01.004 . ПМИД 25637880 . S2CID 35199775 .
- ^ Линь Ю., Кнутсен П.М., Мюллер А., Кляйнфельд Д., Цянь Р.Ю. (октябрь 2013 г.). «ReaChR: вариант канального родопсина со смещением в красную сторону обеспечивает глубокое транскраниальное оптогенетическое возбуждение» . Природная неврология . 16 (10): 1499–1508. дои : 10.1038/nn.3502 . ПМЦ 3793847 . ПМИД 23995068 .
- ^ Мэтьюз Г.А., Них Э.Х., Вандер Вил К.М., Халберт С.А., Прадхан Р.В., Джосафат А.С. и др. (февраль 2016 г.). «Дофаминовые нейроны дорсального шва представляют собой опыт социальной изоляции» . Ячейка 164 (4): 617–631. дои : 10.1016/j.cell.2015.12.040 . ПМЦ 4752823 . ПМИД 26871628 .
- ^ Клапоетке Н.К., Мурата Ю., Ким С.С., Пулвер С.Р., Бердси-Бенсон А., Чо Ю.К. и др. (март 2014 г.). «Независимое оптическое возбуждение отдельных нейронных популяций» . Природные методы . 11 (3): 338–346. дои : 10.1038/nmeth.2836 . ПМЦ 3943671 . ПМИД 24509633 .
- ^ Берндт А., Ижар О., Гюнайдин Л.А., Хегеманн П., Дейссерот К. (февраль 2009 г.). «Бистабильные переключатели состояний нейронов». Природная неврология . 12 (2): 229–234. дои : 10.1038/nn.2247 . ПМИД 19079251 . S2CID 15125498 .
- ^ Говорунова Е.Г., Синещеков О.А., Янц Р., Лю Х, Спудич Ю.Л. (август 2015 г.). «НЕЙРОНАУКА. Естественные светозависимые анионные каналы: семейство микробных родопсинов для передовой оптогенетики» . Наука . 349 (6248): 647–650. дои : 10.1126/science.aaa7484 . ПМЦ 4764398 . ПМИД 26113638 .
- ^ Мосс А.С., Буш С., Борст А. (октябрь 2017 г.). «Оптогенетическое замалчивание нейронов у дрозофилы во время обработки изображений» . Научные отчеты . 7 (1): 13823. Бибкод : 2017НатСР...713823М . дои : 10.1038/s41598-017-14076-7 . ПМЦ 5653863 . ПМИД 29061981 .
- ^ Португальский, Рубен; Севери, Кристен Э; Вайарт, Клэр; Аренс, Миша Б (01 февраля 2013 г.). «Оптогенетика прозрачного животного: функция контура личинки данио» . Современное мнение в нейробиологии . Нейрогенетика. 23 (1): 119–126. дои : 10.1016/j.conb.2012.11.001 . ISSN 0959-4388 . ПМИД 23246238 . S2CID 19906279 .
- ^ Канде, Джессика; Намики, Сигэхиро; Цю, Джируй; Корфф, Вятт; Кард, Гвинет М; Шаевиц, Джошуа В; Стерн, Дэвид Л; Берман, Гордон Дж (26 июня 2018 г.). Скотт, Кристин (ред.). «Оптогенетическое исследование нисходящего поведенческого контроля у дрозофилы» . электронная жизнь . 7 : е34275. дои : 10.7554/eLife.34275 . ISSN 2050-084X . ПМК 6031430 . ПМИД 29943729 .
- ^ ДеАнджелис, Брайан Д.; Заватоне-Вет, Джейкоб А; Гонсалес-Суарес, Анейсис Д; Кларк, Дэймон А. (22 апреля 2020 г.). Калабрезе, Рональд Л. (ред.). «Пространственно-временно точная оптогенетическая активация сенсорных нейронов свободно передвигающейся дрозофилы» . электронная жизнь . 9 : е54183. doi : 10.7554/eLife.54183 . ISSN 2050-084X . ПМЦ 7198233 . ПМИД 32319425 .
- ^ Бидай, Салил С.; Латурни, Меган; Чанг, Эми К.; Лю, Юэцзян; Бокемюль, Тилль; Бюшгес, Ансгар; Скотт, Кристин (11 ноября 2020 г.). «Два мозговых пути инициируют различные программы ходьбы вперед у дрозофилы» . Нейрон . 108 (3): 469–485.e8. дои : 10.1016/j.neuron.2020.07.032 . ISSN 0896-6273 . ПМЦ 9435592 . ПМИД 32822613 . S2CID 221198570 .
- ^ Хейни, Шейн А.; Ким, Джинсук; Августин, Джордж Дж.; Медина, Хавьер Ф. (5 февраля 2014 г.). «Точный контроль кинематики движения путем оптогенетического торможения активности клеток Пуркинье» . Журнал неврологии . 34 (6): 2321–2330. doi : 10.1523/JNEUROSCI.4547-13.2014 . ISSN 0270-6474 . ПМЦ 3913874 . ПМИД 24501371 .
- ^ Солари Н., Святко К., Ласловский Т., Хегедюс П., Ханья Б. (май 2018 г.). «Инструменты с открытым исходным кодом для временного контроля поведения грызунов, подходящие для электрофизиологии и оптогенетических манипуляций» . Границы системной нейронауки . 12:18 . дои : 10.3389/fnsys.2018.00018 . ПМЦ 5962774 . ПМИД 29867383 .
- ^ Лин Дж. Ю. (январь 2011 г.). «Руководство пользователя по вариантам канального родопсина: особенности, ограничения и будущие разработки» . Экспериментальная физиология . 96 (1): 19–25. doi : 10.1113/expphysicalol.2009.051961 . ПМЦ 2995811 . ПМИД 20621963 .
- ^ Гросеник, Логан; Маршел, Джеймс Х.; Дейссерот, Карл (08 апреля 2015 г.). «Замкнутый контур и оптогенетический контроль, управляемый активностью» . Нейрон . 86 (1): 106–139. дои : 10.1016/j.neuron.2015.03.034 . ISSN 0896-6273 . ПМЦ 4775736 . ПМИД 25856490 .
- ^ Армстронг, Карен; Крук-Магнусон, Эстер; Ойала, Микко; Солтеш, Иван (2013). «Замкнутое оптогенетическое вмешательство у мышей» . Протоколы природы . 8 (8): 1475–1493. дои : 10.1038/нпрот.2013.080 . ISSN 1750-2799 . ПМЦ 3988315 . ПМИД 23845961 .
- ^ Ковач К.А., О'Нил Дж., Шененбергер П., Пенттонен М., Рангель Герреро Д.К., Чиксвари Дж. (19 ноября 2016 г.). «Оптогенетическое блокирование резких волновых пульсаций во сне не мешает формированию стабильного пространственного представления в области CA1 гиппокампа» . ПЛОС ОДИН . 11 (10): e0164675. Бибкод : 2016PLoSO..1164675K . дои : 10.1371/journal.pone.0164675 . ПМК 5070819 . ПМИД 27760158 .
- ^ Перейти обратно: а б Валон Л., Марин-Ллаурадо А., Вятт Т., Чаррас Г., Трепат Икс (февраль 2017 г.). «Оптогенетический контроль клеточных сил и механотрансдукции» . Природные коммуникации . 8 : 14396. Бибкод : 2017NatCo...814396V . дои : 10.1038/ncomms14396 . ПМК 5309899 . ПМИД 28186127 .
- ^ Перейти обратно: а б с д и Хамо Дж.С., Кришнамурти В.В., Шарум С.Р., Мондал П., Чжан К. (октябрь 2017 г.). «Применение оптобиологии в интактных клетках и многоклеточных организмах». Журнал молекулярной биологии . 429 (20): 2999–3017. дои : 10.1016/j.jmb.2017.08.015 . ПМИД 28882542 .
- ^ Фенно Л., Ижар О., Дейсерот К. (2011). «Развитие и применение оптогенетики» . Ежегодный обзор неврологии . 34 : 389–412. doi : 10.1146/annurev-neuro-061010-113817 . ПМК 6699620 . ПМИД 21692661 .
- ^ «Метод года 2010: Оптогенетика» . Видео о природе . 17 декабря 2010 г.
- ^ "оптогенетика - Результаты поиска" . ПабМед . Проверено 29 февраля 2020 г.
- ^ Виттманн Т., Дема А., ван Харен Дж. (октябрь 2020 г.). «Свет, цитоскелет, действие: Оптогенетический контроль динамики клеток» . Современное мнение в области клеточной биологии . 66 . ООО «Эльзевир»: 1–10. дои : 10.1016/j.ceb.2020.03.003 . ПМЦ 7577957 . ПМИД 32371345 .
- ^ Конерманн С., Бригам М.Д., Тревино А., Сюй П.Д., Хайденрайх М., Конг Л. и др. (август 2013 г.). «Оптический контроль эндогенной транскрипции и эпигенетических состояний млекопитающих» . Природа . 500 (7463): 472–476. Бибкод : 2013Natur.500..472K . дои : 10.1038/nature12466 . ПМЦ 3856241 . ПМИД 23877069 .
- ^ Люнг Д.В., Отомо С., Чори Дж., Розен М.К. (сентябрь 2008 г.). «Генетически закодированное фотопереключение сборки актина посредством сложного пути Cdc42-WASP-Arp2/3» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (35): 12797–12802. Бибкод : 2008PNAS..10512797L . дои : 10.1073/pnas.0801232105 . ПМК 2525560 . ПМИД 18728185 .
- ^ Тетчер Дж. Э., Гонг Д., Лим В.А., Вайнер О.Д. (сентябрь 2011 г.). «Световая обратная связь для управления динамикой внутриклеточной передачи сигналов» . Природные методы . 8 (10): 837–839. дои : 10.1038/nmeth.1700 . ПМК 3184382 . ПМИД 21909100 .
- ^ Стрикленд Д., Лин Ю., Вагнер Э., Хоуп С.М., Зайнер Дж., Антониу С. и др. (март 2012 г.). «ТЮЛЬПЫ: настраиваемые, управляемые светом взаимодействующие белковые метки для клеточной биологии» . Природные методы . 9 (4): 379–384. дои : 10.1038/nmeth.1904 . ПМЦ 3444151 . ПМИД 22388287 .
- ^ Идевалл-Хагрен О., Диксон Э.Дж., Хилле Б., Тоомре Д.К., Де Камилли П. (август 2012 г.). «Оптогенетический контроль метаболизма фосфоинозитидов» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (35): Е2316–Е2323. Бибкод : 2012PNAS..109E2316I . дои : 10.1073/pnas.1211305109 . ПМЦ 3435206 . ПМИД 22847441 .
- ^ Бугай Л.Дж., Чокси А.Т., Месуда К.К., Кейн Р.С., Шаффер Д.В. (март 2013 г.). «Оптогенетическая кластеризация белков и активация передачи сигналов в клетках млекопитающих». Природные методы . 10 (3): 249–252. дои : 10.1038/nmeth.2360 . ПМИД 23377377 . S2CID 8737019 .
- ^ Лунгу О.И., Халлетт Р.А., Чой Э.Дж., Айкен М.Дж., Хан К.М., Кульман Б. (апрель 2012 г.). «Проектирование фотопереключаемых пептидов с использованием домена AsLOV2» . Химия и биология . 19 (4): 507–517. doi : 10.1016/j.chembiol.2012.02.006 . ПМЦ 3334866 . ПМИД 22520757 .
- ^ Ву Й.И., Фрей Д., Лунгу О.И., Джариг А., Шлихтинг И., Кульман Б., Хан К.М. (сентябрь 2009 г.). «Генетически закодированный фотоактивируемый Rac контролирует подвижность живых клеток» . Природа . 461 (7260): 104–108. Бибкод : 2009Natur.461..104W . дои : 10.1038/nature08241 . ПМЦ 2766670 . ПМИД 19693014 .
- ^ Смарт А.Д., Паче Р.А., Томсен Н.Д., Кортемме Т. , Дэвис Г.В., Уэллс Дж.А. (сентябрь 2017 г.). «Разработка активируемой светом каспазы-3 для точной абляции нейронов in vivo» . Труды Национальной академии наук Соединенных Штатов Америки . 114 (39): Е8174–Е8183. Бибкод : 2017PNAS..114E8174S . дои : 10.1073/pnas.1705064114 . ПМК 5625904 . ПМИД 28893998 .
- ^ Даглиян О, Тарнавский М, Чу П.Х., Ширванянц Д, Шлихтинг И, Дохолян Н.В., Хан К.М. (декабрь 2016 г.). «Инженерия внешнего расстройства для контроля активности белка в живых клетках» . Наука . 354 (6318): 1441–1444. Бибкод : 2016Sci...354.1441D . дои : 10.1126/science.aah3404 . ПМК 5362825 . ПМИД 27980211 .
- ^ Гунтас Г., Халлетт Р.А., Циммерман С.П., Уильямс Т., Юмерефенди Х., Беар Дж.Э., Кульман Б. (январь 2015 г.). «Разработка улучшенного светоиндуцируемого димера (iLID) для контроля локализации и активности сигнальных белков» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (1): 112–117. Бибкод : 2015PNAS..112..112G . дои : 10.1073/pnas.1417910112 . ПМК 4291625 . ПМИД 25535392 .
- ^ Ван Х., Вилела М., Винклер А., Тарнавски М., Шлихтинг И., Юмерефенди Х. и др. (сентябрь 2016 г.). «LOVTRAP: оптогенетическая система фотоиндуцированной диссоциации белков» . Природные методы . 13 (9): 755–758. дои : 10.1038/nmeth.3926 . ПМК 5137947 . ПМИД 27427858 .
- ^ Ван Харен Дж., Чарафеддин Р.А., Эттингер А., Ван Х., Хан К.М., Виттманн Т. (март 2018 г.). «Локальный контроль внутриклеточной динамики микротрубочек посредством фотодиссоциации EB1» . Природная клеточная биология . 20 (3). Исследования природы.: 252–261. дои : 10.1038/s41556-017-0028-5 . ПМЦ 5826794 . ПМИД 29379139 .
- ^ Перейти обратно: а б Чжоу XX, Чунг Х.К., Лам А.Дж., Линь М.З. (ноябрь 2012 г.). «Оптический контроль активности белка с помощью флуоресцентных белковых доменов» . Наука . 338 (6108): 810–814. Бибкод : 2012Sci...338..810Z . дои : 10.1126/science.1226854 . ПМК 3702057 . ПМИД 23139335 .
- ^ Тишер Д., Вайнер О.Д. (август 2014 г.). «Освещение клеточной сигнализации с помощью оптогенетических инструментов» . Обзоры природы. Молекулярно-клеточная биология . 15 (8): 551–558. дои : 10.1038/nrm3837 . ПМК 4145075 . ПМИД 25027655 .
- ^ Первис Дж. Э., Лахав Г. (февраль 2013 г.). «Кодирование и декодирование клеточной информации посредством динамики сигналов» . Клетка . 152 (5): 945–956. дои : 10.1016/j.cell.2013.02.005 . ПМК 3707615 . ПМИД 23452846 .
- ^ Сантос С.Д., Вервир П.Дж., Бастианс П.И. (март 2007 г.). «Топология сети MAPK, индуцированная фактором роста, формирует ответ Erk, определяющий судьбу клеток PC-12». Природная клеточная биология . 9 (3): 324–330. дои : 10.1038/ncb1543 . ПМИД 17310240 . S2CID 31709706 .
- ^ Тоттчер Дж. Э., Вайнер О. Д., Лим В. А. (декабрь 2013 г.). «Использование оптогенетики для исследования динамического контроля передачи сигналов модулем Ras/Erk» . Клетка . 155 (6): 1422–1434. дои : 10.1016/j.cell.2013.11.004 . ПМЦ 3925772 . ПМИД 24315106 .
- ^ Уидобро Н., Мендес-Фернандес А., Мендес-Бальбуэна И., Гутьеррес Р., Кристева Р., Манхаррес Э. (2017). «Брауновская оптогенетико-шумовая фотостимуляция мозга усиливает соматосенсорно-вызванные полевые потенциалы» . Границы в неврологии . 11 : 464. дои : 10.3389/fnins.2017.00464 . ПМЦ 5583167 . ПМИД 28912671 .
- ^ Уидобро Н., Де ла Торре-Вальдовинос Б., Мендес А., Тревиньо М., Ариас-Каррион О., Чавес Ф. и др. (январь 2018 г.). «Оптогенетическая шумовая фотостимуляция мозга усиливает реакцию соматосенсорных импульсов». Письма по неврологии . 664 : 51–57. дои : 10.1016/j.neulet.2017.11.004 . ПМИД 29128628 . S2CID 3370851 .
- ^ Мабил П., Уидобро Н., Торрес-Рамирес О., Флорес-Эрнандес Х., Флорес А., Гутьеррес Р., Манхаррес Э. (2020). «Шумный свет усиливает Na» + оптогенетических трансгенных мышей» . Frontiers in Neuroscience . 14 : 490. doi : 10.3389/fnins.2020.00490 . PMC 7263390. Ток в соматосенсорных пирамидальных нейронах PMID 32528244 .
- ^ Девятая ежегодная премия Wiley в области биомедицинских наук присуждена доктору Питеру Хегеманну, доктору Георгу Нагелю и доктору Эрнсту Бамбергу (wiley.com)
- ^ «Карл Хайнц Бекуртс-Прейс 2010» . Фонд Карла Хайнца Беккурта .
- ^ «Премия HFSP Накасоне 2010» . Программа человеческих передовых наук .
- ^ «Международная премия в области трансляционной нейронауки Фонда Гертруды Реемтсма (Премия К. Я. Цульха до 2019 г.)» . Общество Макса Планка .
- ^ «Международная премия в области здравоохранения InBev-Baillet Latour» (PDF) . Фонд научных исследований — ФНРС .
- ^ Премия Луи-Жанте
- ^ «Премия Мозга 2013» . Архивировано из оригинала 4 октября 2013 года . Проверено 3 октября 2013 г.
- ^ Райнер А., Исакофф Е.Ю. (октябрь 2013 г.). «Премия мозга 2013: революция в оптогенетике». Тенденции в нейронауках . 36 (10): 557–560. дои : 10.1016/j.tins.2013.08.005 . ПМИД 24054067 . S2CID 205404606 .
- ^ «Премия Эльзы Крёнер Фрезениус за медицинские исследования 2017» . Фонд Эльзы Крёнер-Фрезениус .
- ^ «Лауреат Киотской премии 2018 года Карл Дейссерот» . Киотская премия .
- ^ «Премия Рамфорда присуждена за изобретение и усовершенствование оптогенетики» . Американская академия искусств и наук . 30 января 2019 года . Проверено 12 марта 2019 г.
- ^ «Лауреат премии Heineken 2020 года Карл Дейссерот» . Премии Хайнекен .
- ^ «Лауреаты премии Шоу 2020 года Мизенбёк, Хегеманн и Георг Нагель» . Премия Шоу .
Дальнейшее чтение
[ редактировать ]- Апасани К. (2017). Оптогенетика: от функции нейронов к картированию и биологии заболеваний . Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 978-1-107-05301-4 .
- Банерджи С., Митра Д. (январь 2020 г.). «Структурные основы проектирования и разработки перспективной оптогенетики растений». Тенденции в науке о растениях . 25 (1): 35–65. doi : 10.1016/j.tplants.2019.10.002 . ПМИД 31699521 . S2CID 207942668 .
- Ху В, Ли Ц, Ли Б, Ма К, Чжан С, Фу X (январь 2020 г.). «Оптогенетика проливает новый свет на тканевую инженерию и регенеративную медицину». Биоматериалы . 227 : 119546. doi : 10.1016/j.bimaterials.2019.119546 . PMID 31655444 . S2CID 204918731 .
- Джаррин С., Финн Д.П. (октябрь 2019 г.). «Оптогенетика и ее применение в исследованиях боли и тревоги». Неврологические и биоповеденческие обзоры . 105 : 200–211. doi : 10.1016/j.neubiorev.2019.08.007 . ПМИД 31421140 . S2CID 199577276 .
- Джонсон Х.Э., Тетчер Дж.Э. (август 2018 г.). «Освещение биологии развития с помощью клеточной оптогенетики» . Современное мнение в области биотехнологии . 52 : 42–48. дои : 10.1016/j.copbio.2018.02.003 . ПМК 6082700 . ПМИД 29505976 .
- Крюгер Д., Искьердо Э., Вишванатан Р., Хартманн Дж., Палларес Картес К., Де Ренцис С. (октябрь 2019 г.). «Принципы и применение оптогенетики в биологии развития» . Разработка . 146 (20): dev175067. дои : 10.1242/dev.175067 . ПМК 6914371 . ПМИД 31641044 .
- Лоси А., Гарднер К.Х., Мёглич А. (ноябрь 2018 г.). «Рецепторы синего света для оптогенетики» . Химические обзоры . 118 (21): 10659–10709. doi : 10.1021/acs.chemrev.8b00163 . ПМК 6500593 . ПМИД 29984995 .
- Вриз С., Одзава Т. (сентябрь 2018 г.). Оптогенетика: световые приводы и светоизлучающие датчики в клеточной биологии . Комплексная серия по фотохимии и фотобиологии. Том. 18. Лондон: Королевское химическое общество. ISBN 978-1-78801-237-9 .
- Виттманн Т., Дема А., ван Харен Дж. (октябрь 2020 г.). «Свет, цитоскелет, действие: Оптогенетический контроль динамики клеток» . Современное мнение в области клеточной биологии . 66 . ООО «Эльзевир»: 1–10. дои : 10.1016/j.ceb.2020.03.003 . ПМЦ 7577957 . ПМИД 32371345 .
Внешние ссылки
[ редактировать ]- «Оптогенетика: проливает свет на тайны мозга» . Научка .
- «Оптогенетика: интегрированная визуализация кальция и оптогенетика» . Инскопикс . 6 апреля 2020 г.