Гликолиз
Гликолиз – это метаболический путь , который превращает глюкозу ( C 6 H 12 O 6 ) в пируват и у большинства организмов встречается в жидкой части клеток ( цитозоле ). используется Высвобождающаяся при этом свободная энергия для образования высокоэнергетических молекул аденозинтрифосфата (АТФ) и восстановленного никотинамидадениндинуклеотида (НАДН). [1] Гликолиз — это последовательность из десяти реакций, катализируемых ферментами .

Широкое распространение гликолиза у других видов указывает на то, что это древний путь метаболизма. [2] составляющие гликолиз и его параллельный путь, пентозофосфатный путь , могут протекать в бескислородных условиях архейских Действительно, реакции , океанов, а также в отсутствие ферментов, катализируемых ионами металлов, то есть это вероятный пребиотический путь. для абиогенеза . [3]
Наиболее распространенным типом гликолиза является путь Эмбдена-Мейергофа-Парнаса (ЭМП) , который был открыт Густавом Эмбденом , Отто Мейергофом и Якубом Каролем Парнасом . Гликолиз также относится к другим путям, таким как путь Энтнера-Дудорова и различные гетероферментативные и гомоферментативные пути. Однако обсуждение здесь будет ограничено путем Эмбдена–Мейергофа–Парнаса. [4]
Путь гликолиза можно разделить на две фазы: [5]
- Инвестиционная фаза – на которой расходуется АТФ.
- Фаза выхода – когда производится больше АТФ, чем первоначально потребляется.
Обзор
[ редактировать ]Общая реакция гликолиза:


Использование символов в этом уравнении делает его несбалансированным по отношению к атомам кислорода, атомам водорода и зарядам. Атомный баланс поддерживается двумя фосфатными (P i ) группами: [6]
- Каждый существует в форме аниона гидрофосфата ( [ГПО 4 ] 2− ), диссоциируя, чтобы внести свой вклад 2ч + общий
- Каждый из них высвобождает атом кислорода, когда он связывается с молекулой аденозиндифосфата (АДФ), в результате чего в общей сложности образуется 2 О.
Заряды уравновешиваются разницей между ADP и ATP. В клеточной среде все три гидроксильные группы АДФ диссоциируют на -O. − и Х + , давая АДФ 3− , и этот ион стремится существовать в ионной связи с Mg 2+ , давая АДПМг − . АТФ ведет себя идентично, за исключением того, что он имеет четыре гидроксильные группы, что дает АТФМг. 2− . Когда эти различия вместе с истинными зарядами двух фосфатных групп рассматриваются вместе, чистые заряды -4 на каждой стороне уравновешиваются.
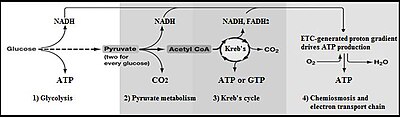
При простой ферментации метаболизм одной молекулы глюкозы в две молекулы пирувата дает чистый выход двух молекул АТФ. Большинство клеток затем проведут дальнейшие реакции, чтобы «погасить» использованный НАД. + и производят конечный продукт из этанола или молочной кислоты . Многие бактерии используют неорганические соединения в качестве акцепторов водорода для регенерации НАД. + .
Клетки, осуществляющие аэробное дыхание, синтезируют гораздо больше АТФ, но не в рамках гликолиза. В этих дальнейших аэробных реакциях используются пируват и НАДН + Н. + от гликолиза. Аэробное дыхание эукариот производит примерно 34 дополнительные молекулы АТФ на каждую молекулу глюкозы, однако большинство из них производятся по механизму, сильно отличающемуся от фосфорилирования на уровне субстрата при гликолизе.
Более низкое производство энергии на глюкозу при анаэробном дыхании по сравнению с аэробным дыханием приводит к большему потоку через путь в условиях гипоксии (с низким содержанием кислорода), если не найдены альтернативные источники анаэробно окисляемых субстратов, таких как жирные кислоты.
| Метаболизм обычных моносахаридов , включая гликолиз, глюконеогенез , гликогенез и гликогенолиз. |
|---|

История
[ редактировать ]Для полного выяснения пути гликолиза, известного сегодня, потребовалось почти 100 лет. [7] Чтобы понять тонкости всего пути, потребовались объединенные результаты многих небольших экспериментов.
Первые шаги в понимании гликолиза начались в девятнадцатом веке с винодельческой промышленности. По экономическим причинам французская винодельческая промышленность стремилась выяснить, почему вино иногда становится неприятным, вместо того, чтобы сбраживаться в алкоголь. Французский ученый Луи Пастер исследовал этот вопрос в 1850-х годах, и результаты его экспериментов положили начало долгому пути к выяснению пути гликолиза. [8] Его эксперименты показали, что брожение происходит под действием живых микроорганизмов , дрожжей, и что потребление глюкозы дрожжами снижается в аэробных условиях брожения по сравнению с анаэробными условиями ( эффект Пастера ). [9]

по неклеточной ферментации Понимание составляющих этапов гликолиза было предоставлено экспериментами Эдуарда Бюхнера в 1890-х годах. [10] [11] Бюхнер продемонстрировал, что превращение глюкозы в этанол возможно с использованием неживого экстракта дрожжей благодаря действию ферментов в экстракте. [12] : 135–148 Этот эксперимент не только произвел революцию в биохимии, но и позволил более поздним ученым проанализировать этот путь в более контролируемых лабораторных условиях. В серии экспериментов (1905–1911) учёные Артур Харден и Уильям Янг открыли ещё фрагменты гликолиза. [13] Они обнаружили регулирующее влияние АТФ на потребление глюкозы во время спиртового брожения. Они также пролили свет на роль одного соединения в качестве промежуточного продукта гликолиза: фруктозо-1,6-бисфосфата. [12] : 151–158
Определение фруктозо-1,6-бисфосфата осуществляли путем измерения Уровни CO 2 при инкубации дрожжевого сока с глюкозой. Производство CO 2 быстро росло, а затем замедлилось. Харден и Янг отметили, что этот процесс возобновится, если в смесь добавить неорганический фосфат (Pi). Харден и Янг пришли к выводу, что в результате этого процесса образуются органические эфиры фосфорной кислоты, а дальнейшие эксперименты позволили им извлечь фруктозодифосфат (F-1,6-DP).
Артур Харден и Уильям Янг вместе с Ником Шеппардом во втором эксперименте определили, что термочувствительная субклеточная фракция с высокой молекулярной массой (ферменты) и термонечувствительная низкомолекулярная фракция цитоплазмы (АДФ, АТФ и НАД) + и другие кофакторы ) необходимы вместе для продолжения ферментации. Этот эксперимент начался с наблюдения, что диализованный (очищенный) дрожжевой сок не может ферментировать или даже создавать фосфат сахара. Эту смесь удалось спасти, добавив кипяченный недиализированный дрожжевой экстракт. Кипячение дрожжевого экстракта делает все белки неактивными (поскольку они денатурируют). Способность кипяченого экстракта и диализированного сока к полному брожению позволяет предположить, что кофакторы имели небелковую природу. [13]

В 1920-х годах Отто Мейерхоф смог связать воедино некоторые из множества отдельных фрагментов гликолиза, открытых Бюхнером, Харденом и Янгом. Мейерхоф и его команда смогли извлечь из мышечной ткани различные гликолитические ферменты и объединить их, чтобы искусственно создать путь от гликогена к молочной кислоте. [14] [15]
В одной статье Мейерхоф и ученый Рената Юнович-Коколати исследовали реакцию, которая расщепляет фруктозо-1,6-дифосфат на два триозофосфата. Предыдущая работа предположила, что расщепление происходит посредством 1,3-дифосфоглицеральдегида плюс окислительного фермента и козимазы. Мейерхофф и Юнович обнаружили, что на константу равновесия реакции изомеразы и альдозы не влияют неорганические фосфаты или любые другие козимазы или окислительные ферменты. Далее они удалили дифосфоглицеральдегид как возможный промежуточный продукт гликолиза. [15]
Имея все эти материалы к 1930-м годам, Густав Эмбден предложил подробную, пошаговую схему того пути, который мы теперь знаем как гликолиз. [16] Наибольшие трудности в определении тонкостей пути были связаны с очень коротким временем жизни и низкими стационарными концентрациями интермедиатов быстрых гликолитических реакций. К 1940-м годам Мейерхоф, Эмбден и многие другие биохимики наконец разгадали загадку гликолиза. [15] В последующие десятилетия понимание изолированного пути расширилось и включило дополнительные детали его регуляции и интеграции с другими метаболическими путями.
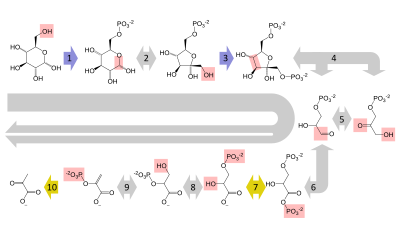
Последовательность реакций
[ редактировать ]Краткое изложение реакций
[ редактировать ]




+


2 × 3-фосфоглицерат

2 × 2-фосфоглицерат

2 × фосфоенолпируват

2 × Пируват
Подготовительный этап
[ редактировать ]Первые пять этапов гликолиза считаются подготовительной (или инвестиционной) фазой, поскольку они потребляют энергию для превращения глюкозы в два трехуглеродных сахарофосфата. [5] ( Г3П ).
| г – Глюкоза ( Glc ) | Гексокиназа глюкокиназа ( ГК ) трансфераза | а- г – Глюкозо-6-фосфат ( G6P ) | |
 | 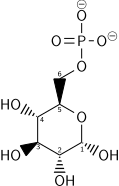 | ||
| СПС | ЧАС + + АДП | ||
Как только глюкоза попадает в клетку, первым шагом является фосфорилирование глюкозы семейством ферментов, называемых гексокиназами, с образованием глюкозо-6-фосфата (G6P). Эта реакция потребляет АТФ, но она поддерживает низкую концентрацию глюкозы внутри клетки, способствуя непрерывному транспорту глюкозы из крови в клетку через транспортеры плазматической мембраны. Кроме того, фосфорилирование блокирует утечку глюкозы – в клетке отсутствуют транспортеры для G6P, и свободная диффузия из клетки предотвращается из-за заряженной природы G6P. Альтернативно, глюкоза может образовываться в результате фосфоролиза или гидролиза внутриклеточного крахмала или гликогена.
У животных в печени используется также изофермент гексокиназы , называемый глюкокиназой , который имеет гораздо меньшее сродство к глюкозе (К м вблизи нормальной гликемии) и отличается регуляторными свойствами. Различное сродство к субстрату и попеременная регуляция этого фермента отражают роль печени в поддержании уровня сахара в крови.
Кофакторы: Mg 2+
| а- г - Глюкозо-6-фосфат ( G6P ) | Фосфоглюкоизомераза ( PGI ) изомераза | б- г - Фруктозо-6-фосфат ( F6P ) | |
|  | ||
G6P затем перегруппировывается во фруктозо-6-фосфат (F6P) под действием глюкозо-фосфат-изомеразы . На этом этапе фруктоза также может вступать в гликолитический путь путем фосфорилирования.
Изменение структуры представляет собой изомеризацию, при которой G6P превращается в F6P. Для проведения реакции требуется фермент фосфоглюкозоизомераза. Эта реакция свободно обратима в нормальных клеточных условиях. Однако он часто продвигается вперед из-за низкой концентрации F6P, который постоянно расходуется на следующем этапе гликолиза. В условиях высокой концентрации F6P эта реакция легко протекает в обратном направлении. Это явление можно объяснить с помощью принципа Ле Шателье . Изомеризация в кетосахар необходима для стабилизации карбаниона на четвертой стадии реакции (ниже).
| б- г - Фруктозо-6-фосфат ( F6P ) | Фосфофруктокиназа ( ПФК-1 ) трансфераза | б- г - Фруктозо-1,6-бисфосфат ( F1,6BP ) | |
|  | ||
| СПС | ЧАС + + АДП | ||
Затраты энергии другой АТФ на этом этапе оправданы двумя способами: Гликолитический процесс (до этого этапа) становится необратимым, и подведенная энергия дестабилизирует молекулу. Поскольку реакция, катализируемая фосфофруктокиназой 1 необходимо использовать другой путь (PFK-1), связана с гидролизом АТФ (энергетически выгодная стадия), она, по сути, необратима, и для обратного преобразования во время глюконеогенеза . Это делает реакцию ключевым моментом регулирования (см. ниже).
Кроме того, второе событие фосфорилирования необходимо для образования двух заряженных групп (а не только одной) на последующей стадии гликолиза, обеспечивая предотвращение свободной диффузии субстратов из клетки.
Эту же реакцию может катализировать пирофосфат-зависимая фосфофруктокиназа ( PFP или PPi-PFK ), которая обнаружена у большинства растений, некоторых бактерий, архей и простейших, но не у животных. Этот фермент использует пирофосфат (PPi) в качестве донора фосфата вместо АТФ. Это обратимая реакция, повышающая гибкость гликолитического метаболизма. [17] У архейных видов был идентифицирован более редкий АДФ-зависимый вариант фермента PFK. [18]
Кофакторы: Mg 2+
| б- г - Фруктозо-1,6-бисфосфат ( F1,6BP ) | Фруктозо-бисфосфатальдолаза ( ALDO ) лиаза | г – глицеральдегид-3-фосфат ( ГАДФ ) | Дигидроксиацетонфосфат ( DHAP ) | ||
|  | + |  | ||
Дестабилизация молекулы в предыдущей реакции позволяет гексозному кольцу расщепиться альдолазой на два триозных сахара: дигидроксиацетонфосфат (кетоза) и глицеральдегид-3-фосфат (альдоза). Существует два класса альдолаз: альдолазы I класса, присутствующие у животных и растений, и альдолазы II класса, присутствующие в грибах и бактериях; эти два класса используют разные механизмы расщепления кетозного кольца.
Электроны, делокализованные при разрыве связи углерод-углерод, присоединяются к спиртовой группе. Образующийся карбанион стабилизируется за счет структуры самого карбаниона за счет резонансного распределения заряда и присутствия простетической группы заряженного иона.
| Дигидроксиацетонфосфат ( DHAP ) | Триозофосфатизомераза ( TPI ) изомераза | г – глицеральдегид-3-фосфат ( ГАДФ ) | |
| | ||
Триозофосфат-изомераза быстро превращает дигидроксиацетонфосфат в глицеральдегид-3-фосфат ( GADP ), который переходит в гликолиз. Это выгодно, поскольку оно направляет дигидроксиацетонфосфат по тому же пути, что и глицеральдегид-3-фосфат, упрощая регуляцию.
Фаза окупаемости
[ редактировать ]Вторая половина гликолиза известна как фаза отдачи, характеризующаяся чистым приростом богатых энергией молекул АТФ и НАДН. [5] Поскольку на подготовительной фазе глюкоза приводит к образованию двух триозных сахаров, каждая реакция на фазе отдачи происходит дважды на молекулу глюкозы. Это дает 2 молекулы НАДН и 4 молекулы АТФ, что приводит к чистому приросту 2 молекул НАДН и 2 молекул АТФ от гликолитического пути на глюкозу.
| Глицеральдегид-3-фосфат ( GADP ) | Глицеральдегидфосфатдегидрогеназа ( ГАФДГ ) оксидоредуктаза | г - 1,3-бисфосфоглицерат ( 1,3BPG ) | |
|  | ||
| ОНИ + + П я | НАДН + Н + | ||
Альдегидные группы триозных сахаров окисляются , и неорганический фосфат к ним присоединяется , образуя 1,3-бисфосфоглицерат .
Водород используется для восстановления двух молекул НАД. + , переносчик водорода, с образованием НАДН + H + за каждую тройку.
Баланс атомов водорода и баланс зарядов сохраняются, поскольку фосфатная группа (P i ) фактически существует в форме аниона гидрофосфата ( ГПО 2- 4 ), [6] который диссоциирует, внося дополнительный H + ион и дает чистый заряд -3 с обеих сторон.
Вот, арсенат ( [АО 4 ] 3− ), анион, родственный неорганическому фосфату, может заменять фосфат в качестве субстрата с образованием 1-арсено-3-фосфоглицерата. Однако он нестабилен и легко гидролизуется с образованием 3-фосфоглицерата , промежуточного продукта на следующем этапе пути. В результате пропуска этого этапа молекула АТФ, образующаяся из 1-3 бисфосфоглицерата в следующей реакции, не будет образована, даже если реакция протекает. В результате арсенат является разобщителем гликолиза. [19]
| 1,3-бисфосфоглицерат ( 1,3BPG ) | Фосфоглицерат киназа ( PGK ) трансфераза | 3-фосфоглицерат ( 3PG ) | |
|  | ||
| АДФ + Ч + | СПС | ||
| Фосфоглицерат киназа ( PGK ) | |||
Этот этап представляет собой ферментативный перенос фосфатной группы от 1,3-бисфосфоглицерата к АДФ с помощью фосфоглицераткиназы с образованием АТФ и 3-фосфоглицерата . На этом этапе гликолиз достиг точки безубыточности: израсходовано 2 молекулы АТФ, и теперь синтезированы 2 новые молекулы. Этот этап, один из двух этапов фосфорилирования на уровне субстрата , требует ADP; таким образом, когда в клетке много АТФ (и мало АДФ), эта реакция не происходит. Поскольку АТФ распадается относительно быстро, когда он не метаболизируется, это важный регуляторный момент гликолитического пути.
ADP на самом деле существует как ADPMg − , а АТФ как ATPMg 2− , уравновешивая заряды на уровне −5 с обеих сторон.
Кофакторы: Mg 2+
| 3-фосфоглицерат ( 3PG ) | Фосфоглицератмутаза ( PGM ) переехал | 2-фосфоглицерат ( 2PG ) | |
|  | ||
Фосфоглицератмутаза изомеризует 3-фосфоглицерат в 2-фосфоглицерат .
| 2-фосфоглицерат ( 2PG ) | Энолаза ( ЭНО ) лиаза | Фосфоенолпируват ( ПЭП ) | |
|  | ||
| Н 2 О | |||
| Энолаза ( ЭНО ) | |||
Энолаза затем превращает 2-фосфоглицерат в фосфоенолпируват . Эта реакция представляет собой реакцию элиминирования с участием механизма E1cB .
Кофакторы: 2 мг 2+ , один «конформационный» ион, координирующийся с карбоксилатной группой субстрата, и один «каталитический» ион, который участвует в дегидратации.
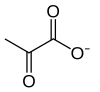
| Фосфоенолпируват ( ПЭП ) | Пируваткиназа ( ПК ) трансфераза | Пируват ( Pyr ) | |
|  | ||
| АДФ + Ч + | СПС | ||
Окончательное фосфорилирование на уровне субстрата теперь образует молекулу пирувата и молекулу АТФ с помощью фермента пируваткиназы . Это служит дополнительным этапом регуляции, аналогичным этапу фосфоглицераткиназы.
Кофакторы: Mg 2+
Биохимическая логика
[ редактировать ]Существование более чем одной точки регуляции указывает на то, что промежуточные соединения между этими точками входят в путь гликолиза и покидают его посредством других процессов. Например, на первом регулируемом этапе гексокиназа превращает глюкозу в глюкозо-6-фосфат. Вместо продолжения пути гликолиза этот промежуточный продукт может превращаться в молекулы хранения глюкозы, такие как гликоген или крахмал . Обратная реакция, при расщеплении, например, гликогена, дает главным образом глюкозо-6-фосфат; В реакции образуется очень мало свободной глюкозы. Полученный таким образом глюкозо-6-фосфат может вступать в гликолиз после первой контрольной точки.
На втором регулируемом этапе (третий этап гликолиза) фосфофруктокиназа превращает фруктозо-6-фосфат во фруктозо-1,6-бисфосфат, который затем превращается в глицеральдегид-3-фосфат и дигидроксиацетонфосфат. Дигидроксиацетонфосфат можно удалить из гликолиза путем превращения в глицерин-3-фосфат, который можно использовать для образования триглицеридов. [20] И наоборот, триглицериды могут расщепляться на жирные кислоты и глицерин; последний, в свою очередь, может превращаться в дигидроксиацетонфосфат, который после второй контрольной точки может вступать в гликолиз.
Бесплатные изменения энергии
[ редактировать ]| Сложный | Концентрация/мМ |
|---|---|
| Глюкоза | 5.0 |
| Глюкозо-6-фосфат | 0.083 |
| Фруктозо-6-фосфат | 0.014 |
| Фруктозо-1,6-бисфосфат | 0.031 |
| Дигидроксиацетонфосфат | 0.14 |
| Глицеральдегид-3-фосфат | 0.019 |
| 1,3-бисфосфоглицерат | 0.001 |
| 2,3-бисфосфоглицерат | 4.0 |
| 3-фосфоглицерат | 0.12 |
| 2-фосфоглицерат | 0.03 |
| фосфоенолпируват | 0.023 |
| Пируват | 0.051 |
| СПС | 1.85 |
| АДП | 0.14 |
| П я | 1.0 |
Изменение свободной энергии ΔG на каждом этапе пути гликолиза можно рассчитать, используя ΔG = ΔG ° ' + RT ln Q , где Q — коэффициент реакции . Для этого необходимо знать концентрации метаболитов . Все эти значения доступны для эритроцитов , за исключением концентрации НАД. + и НАДН. Соотношение НАД + к НАДН в цитоплазме составляет примерно 1000, что делает окисление глицеральдегид-3-фосфата (стадия 6) более благоприятным.
Используя измеренные концентрации на каждом этапе и стандартные изменения свободной энергии, можно рассчитать фактическое изменение свободной энергии. (Игнорирование этого очень распространено: дельта G гидролиза АТФ в клетках не является стандартным изменением свободной энергии при гидролизе АТФ, указанным в учебниках).
| Шаг | Реакция | Δ G °' (кДж/моль) | ΔΔG (кДж/моль) | ||
|---|---|---|---|---|---|
| 1 | Глюкоза + АТФ 4− → Глюкозо-6-фосфат 2− + АДП 3− + Ч + | −16 | .7 | −34 | |
| 2 | Глюкозо-6-фосфат 2− → Фруктозо-6-фосфат 2− | 1 | .67 | −2 | .9 |
| 3 | Фруктозо-6-фосфат 2− + АТФ 4− → Фруктозо-1,6-бисфосфат 4− + АДП 3− + Ч + | −14 | .2 | −19 | |
| 4 | Фруктозо-1,6-бисфосфат 4− → Дигидроксиацетонфосфат 2− + Глицеральдегид-3-фосфат 2− | 23 | .9 | −0 | .23 |
| 5 | Дигидроксиацетонфосфат 2− → Глицеральдегид-3-фосфат 2− | 7 | .56 | 2 | .4 |
| 6 | Глицеральдегид-3-фосфат 2− + П я 2− + ОНИ + → 1,3-бисфосфоглицерат 4− + НАДН + Н + | 6 | .30 | −1 | .29 |
| 7 | 1,3-бисфосфоглицерат 4− + АДП 3− → 3-фосфоглицерат 3− + АТФ 4− | −18 | .9 | 0 | .09 |
| 8 | 3-фосфоглицерат 3− → 2-фосфоглицерат 3− | 4 | .4 | 0 | .83 |
| 9 | 2-фосфоглицерат 3− → Фосфоенолпируват 3− + Н 2 О | 1 | .8 | 1 | .1 |
| 10 | фосфоенолпируват 3− + АДП 3− + Ч + → Пируват − + АТФ 4− | −31 | .7 | −23 | .0 |
Измерение физиологических концентраций метаболитов в эритроцитах показывает, что около семи стадий гликолиза находятся в равновесии для этого типа клеток. Три из стадий — с большими отрицательными изменениями свободной энергии — не находятся в равновесии и называются необратимыми ; такие шаги часто подлежат регулированию.
Шаг 5 на рисунке показан позади других этапов, поскольку этот этап представляет собой побочную реакцию, которая может уменьшить или увеличить концентрацию промежуточного глицеральдегид-3-фосфата. Это соединение превращается в дигидроксиацетонфосфат с помощью фермента триозофосфатизомеразы, который является каталитически совершенным ферментом; ее скорость настолько высока, что можно предположить, что реакция находится в равновесии. Тот факт, что ΔG не равен нулю, указывает на то, что фактические концентрации в эритроцитах точно не известны.
Регулирование
[ редактировать ]Ферменты, катализирующие гликолиз, регулируются с помощью ряда биологических механизмов, чтобы контролировать общий поток гликолиза по этому пути. Это жизненно важно как для гомеостаза в статической среде, так и для метаболической адаптации к изменяющейся среде или потребностям. [22] Детали регуляции некоторых ферментов хорошо консервативны у разных видов, тогда как другие сильно различаются. [23] [24]
- Экспрессия генов. Во-первых, клеточные концентрации гликолитических ферментов модулируются посредством регуляции экспрессии генов с помощью факторов транскрипции . [25] при этом несколько ферментов гликолиза сами действуют как регуляторные протеинкиназы в ядре. [26]
- Аллостерическое ингибирование и активация метаболитами: в частности, ингибирование конечного продукта регулируемых ферментов метаболитами, такими как АТФ, служит регуляцией этого пути по отрицательной обратной связи. [23] [27]
- Аллостерическое ингибирование и активация посредством белок-белковых взаимодействий (PPI). [28] Действительно, некоторые белки взаимодействуют с несколькими гликолитическими ферментами и регулируют их. [29]
- Посттрансляционная модификация (ПТМ) . [30] В частности, фосфорилирование и дефосфорилирование являются ключевым механизмом регуляции пируваткиназы в печени.
- Локализация [27]
Регуляция инсулином у животных
[ редактировать ]У животных регуляция уровня глюкозы в крови поджелудочной железой совместно с печенью является жизненно важной частью гомеостаза . островков Бета-клетки поджелудочной железы чувствительны к концентрации глюкозы в крови. [31] Повышение концентрации глюкозы в крови приводит к выбросу инсулина в кровь, что особенно влияет на печень, а также на жировые и мышечные клетки, заставляя эти ткани удалять глюкозу из крови. Когда уровень сахара в крови падает, бета-клетки поджелудочной железы прекращают выработку инсулина, но вместо этого стимулируют соседние альфа-клетки поджелудочной железы высвобождать глюкагон в кровь. [31] Это, в свою очередь, заставляет печень выделять глюкозу в кровь путем расщепления накопленного гликогена и посредством глюконеогенеза. Если падение уровня глюкозы в крови особенно быстрое или сильное, другие датчики глюкозы вызывают выброс адреналина из надпочечников в кровь. Он оказывает такое же действие, как глюкагон, на метаболизм глюкозы, но его эффект более выражен. [31] В печени глюкагон и адреналин вызывают фосфорилирование ключевых, регулируемых ферментов гликолиза, синтеза жирных кислот , синтеза холестерина , глюконеогенеза и гликогенолиза. Инсулин оказывает противоположное действие на эти ферменты. [32] Фосфорилирование и дефосфорилирование этих ферментов (в конечном итоге в ответ на уровень глюкозы в крови) является доминирующим способом контроля этих путей в клетках печени, жировых и мышечных клетках. Так, фосфорилирование фосфофруктокиназы ингибирует гликолиз, тогда как ее дефосфорилирование под действием инсулина стимулирует гликолиз. [32]
Регулируемые ферменты гликолиза
[ редактировать ]Тремя регуляторными ферментами являются гексокиназа (или глюкокиназа в печени), фосфофруктокиназа и пируваткиназа . Поток . через гликолитический путь регулируется в ответ на условия как внутри, так и снаружи клетки Внутренние факторы, регулирующие гликолиз, действуют главным образом для обеспечения АТФ в количествах, достаточных для нужд клетки. Внешние факторы действуют в первую очередь на печень , жировую ткань и мышцы , которые могут удалять большие количества глюкозы из крови после еды (таким образом предотвращая гипергликемию , сохраняя избыток глюкозы в виде жира или гликогена, в зависимости от типа ткани). Печень также способна выбрасывать глюкозу в кровь между приемами пищи, во время голодания и физических упражнений, тем самым предотвращая гипогликемию посредством гликогенолиза и глюконеогенеза . Эти последние реакции совпадают с остановкой гликолиза в печени.
Кроме того, гексокиназа и глюкокиназа действуют независимо от гормональных эффектов, контролируя точки входа глюкозы в клетки различных тканей. Гексокиназа реагирует на уровень глюкозо-6-фосфата (G6P) в клетке или, в случае глюкокиназы, на уровень сахара в крови, обеспечивая полностью внутриклеточный контроль гликолитического пути в различных тканях (см. ниже ). [32]
Когда глюкоза превращается в G6P под действием гексокиназы или глюкокиназы, она может либо превращаться в глюкозо-1-фосфат (G1P) для превращения в гликоген , либо, альтернативно, путем гликолиза превращается в пируват , который поступает в митохондрии , где превращается в ацетил-КоА , а затем в цитрат . Избыток цитрата экспортируется из митохондрий обратно в цитозоль, где АТФ-цитратлиаза регенерирует ацетил-КоА и оксалоацетат (ОАА). Ацетил-КоА затем используется для синтеза жирных кислот и синтеза холестерина — двух важных способов использования избытка глюкозы, когда ее концентрация в крови высока. Регулируемые ферменты, катализирующие эти реакции, выполняют эти функции, когда они дефосфорилируются под действием инсулина на клетки печени. Между приемами пищи, во время голодания , физических упражнений или гипогликемии в кровь выбрасываются глюкагон и адреналин. Это приводит к тому, что гликоген печени преобразуется обратно в G6P, а затем превращается в глюкозу с помощью печеночно-специфического фермента. глюкозо-6-фосфатазы и выделяется в кровь. Глюкагон и адреналин также стимулируют глюконеогенез, который превращает неуглеводные субстраты в G6P, который присоединяется к G6P, полученному из гликогена, или заменяет его, когда запасы гликогена в печени истощены. Это имеет решающее значение для функции мозга, поскольку в большинстве случаев мозг использует глюкозу в качестве источника энергии. [33] Одновременное фосфорилирование, в частности, фосфофруктокиназы , а также в определенной степени пируваткиназы предотвращает гликолиз, происходящий одновременно с глюконеогенезом и гликогенолизом.
Гексокиназа и глюкокиназа
[ редактировать ]
Во всех клетках содержится фермент гексокиназа , катализирующий превращение поступившей в клетку глюкозы в глюкозо-6-фосфат (Г6Ф). Поскольку клеточная мембрана непроницаема для G6P, гексокиназа по существу транспортирует глюкозу в клетки, из которых она больше не может выйти. Гексокиназа ингибируется высокими уровнями G6P в клетке. Таким образом, скорость поступления глюкозы в клетки частично зависит от того, насколько быстро G6P может быть утилизирован путем гликолиза и синтеза гликогена (в клетках, запасающих гликоген, а именно в печени и мышцах). [32] [34]
Глюкокиназа , в отличие от гексокиназы, не ингибируется G6P. Он происходит в клетках печени и фосфорилирует глюкозу, поступающую в клетку, с образованием G6P только тогда, когда глюкозы в крови много. Это первый этап гликолитического пути в печени, поэтому он обеспечивает дополнительный уровень контроля гликолитического пути в этом органе. [32]
Фосфофруктокиназа
[ редактировать ]
Фосфофруктокиназа является важной контрольной точкой гликолитического пути, поскольку она является одним из необратимых этапов и имеет ключевые аллостерические эффекторы: АМФ и фруктозо-2,6-бисфосфат (F2,6BP).
F2,6BP является очень мощным активатором фосфофруктокиназы (PFK-1), которая синтезируется при фосфорилировании F6P второй фосфофруктокиназой ( PFK2 ). В печени, когда уровень сахара в крови низкий, а глюкагон повышает цАМФ, PFK2 фосфорилируется протеинкиназой А. Фосфорилирование инактивирует PFK2, и другой домен этого белка становится активным как фруктозобисфосфатаза-2 , которая превращает F2,6BP обратно в F6P. И глюкагон , и адреналин вызывают высокий уровень цАМФ в печени. Результатом более низких уровней F2,6BP в печени является снижение активности фосфофруктокиназы и повышение активности фруктозо-1,6-бисфосфатазы , так что благоприятствует глюконеогенез (по сути, «гликолиз наоборот»). Это согласуется с ролью печени в таких ситуациях, поскольку ответом печени на эти гормоны является выброс глюкозы в кровь.
АТФ конкурирует с АМФ за аллостерический эффекторный участок фермента ПФК. Концентрации АТФ в клетках намного выше, чем у АМФ, обычно в 100 раз выше. [35] но концентрация АТФ в физиологических условиях изменяется не более чем на 10%, тогда как падение АТФ на 10% приводит к 6-кратному увеличению АМФ. [36] Таким образом, значимость АТФ как аллостерического эффектора сомнительна. Увеличение АМФ является следствием уменьшения энергетического заряда в клетке.
Цитрат ингибирует фосфофруктокиназу при тестировании in vitro , усиливая ингибирующий эффект АТФ. Однако сомнительно, что это значимый эффект in vivo , поскольку цитрат в цитозоле используется главным образом для превращения в ацетил-КоА для синтеза жирных кислот и холестерина .
TIGAR , фермент, индуцируемый р53, отвечает за регуляцию фосфофруктокиназы и защищает от окислительного стресса. [37] TIGAR — это единственный фермент с двойной функцией, который регулирует F2,6BP. Он может вести себя как фосфатаза (фруктуозо-2,6-бисфосфатаза), которая расщепляет фосфат по углероду-2 с образованием F6P. Он также может вести себя как киназа (PFK2), добавляя фосфат к углероду-2 F6P, что приводит к образованию F2,6BP. У человека белок TIGAR кодируется геном C12orf5 . Фермент TIGAR будет препятствовать дальнейшему развитию гликолиза, создавая накопление фруктозо-6-фосфата (F6P), который изомеризуется в глюкозо-6-фосфат (G6P). Накопление G6P будет переводить углероды в пентозофосфатный путь. [38] [39]
Пируваткиназа
[ редактировать ]
Последний этап гликолиза катализируется пируваткиназой с образованием пирувата и другого АТФ. Он регулируется рядом различных транскрипционных, ковалентных и нековалентных механизмов регуляции, которые могут широко варьироваться в разных тканях. [40] [41] [42] Например, в печени пируваткиназа регулируется в зависимости от доступности глюкозы. Во время голодания (глюкоза отсутствует) глюкагон активирует протеинкиназу А , которая фосфорилирует пируваткиназу, ингибируя ее. [43] Повышение уровня сахара в крови приводит к секреции инсулина , который активирует протеинфосфатазу 1 , что приводит к дефосфорилированию и повторной активации пируваткиназы. [43] Эти средства контроля предотвращают активность пируваткиназы одновременно с ферментами, катализирующими обратную реакцию ( пируваткарбоксилаза и фосфоенолпируваткарбоксикиназа ), предотвращая бесполезный цикл . [43] И наоборот, изоформа пируваткиназы, обнаруженная в мышцах, не подвергается воздействию протеинкиназы А (которая активируется адреналином в этой ткани), поэтому гликолиз остается активным в мышцах даже во время голодания. [43]
Постгликолизные процессы
[ редактировать ]Общий процесс гликолиза:
- Глюкоза + 2 НАД + + 2 АДФ + 2 П i → 2 Пируват + 2 НАДН + 2 Н + + 2 АТФ + 2 Н 2 О
Если бы гликолиз продолжался бесконечно, весь НАД + будет израсходовано, и гликолиз прекратится. Чтобы гликолиз мог продолжаться, организмы должны быть способны окислять НАДН обратно в НАД. + . Как это осуществляется, зависит от того, какой внешний акцептор электронов доступен.
Аноксическая регенерация НАД +
[ редактировать ]Один из способов сделать это — просто заставить пируват совершить окисление; в этом процессе пируват превращается в лактат ( основание, сопряженное с молочной кислотой) в процессе, называемом молочнокислым брожением :
- Пируват + НАДН + Н + → Молочные продукты + НАД +
Этот процесс происходит в бактериях , участвующих в приготовлении йогурта (молочная кислота вызывает свертывание молока). Этот процесс происходит и у животных в гипоксических (или частично анаэробных) условиях, например, в переутомленных мышцах, которым не хватает кислорода. Во многих тканях это последнее средство получения энергии для клеток; большинство тканей животных не могут переносить анаэробные условия в течение длительного периода времени.
Некоторые организмы, например дрожжи, превращают НАДН обратно в НАД. + в процессе, называемом ферментацией этанола . В этом процессе пируват сначала превращается в ацетальдегид и углекислый газ, а затем в этанол.
Молочнокислое брожение и этаноловое брожение могут происходить в отсутствие кислорода. Эта анаэробная ферментация позволяет многим одноклеточным организмам использовать гликолиз в качестве единственного источника энергии.
Аноксическая регенерация НАД + является лишь эффективным средством производства энергии во время коротких интенсивных упражнений у позвоночных в течение периода от 10 секунд до 2 минут при максимальном усилии у человека. (При более низкой интенсивности упражнений он может поддерживать мышечную активность у ныряющих животных , таких как тюлени, киты и другие водные позвоночные, в течение гораздо более длительных периодов времени.) В этих условиях НАД + пополняется НАДН, отдавая свои электроны пирувату с образованием лактата. При этом на одну молекулу глюкозы приходится 2 молекулы АТФ, или около 5% энергетического потенциала глюкозы (38 молекул АТФ у бактерий). Но скорость, с которой производится таким образом АТФ, примерно в 100 раз превышает скорость окислительного фосфорилирования. Уровень pH в цитоплазме быстро падает, когда ионы водорода накапливаются в мышцах, что в конечном итоге ингибирует ферменты, участвующие в гликолизе.
Ощущение жжения в мышцах во время тяжелых упражнений можно объяснить высвобождением ионов водорода во время перехода к ферментации глюкозы от окисления глюкозы к углекислому газу и воде, когда аэробный метаболизм больше не может идти в ногу с энергетическими потребностями мышц. Эти ионы водорода входят в состав молочной кислоты. Организм прибегает к этому менее эффективному, но более быстрому методу производства АТФ в условиях низкого содержания кислорода. Считается, что это было основным средством производства энергии у более ранних организмов до того, как кислород достиг высоких концентраций в атмосфере между 2000 и 2500 миллионами лет назад, и, таким образом, представляет собой более древнюю форму производства энергии, чем аэробное пополнение НАД. + в клетках.
Печень млекопитающих избавляется от избытка лактата, превращая его обратно в пируват в аэробных условиях; см. цикл Кори .
Ферментацию пирувата до лактата иногда еще называют «анаэробным гликолизом», однако гликолиз заканчивается образованием пирувата независимо от присутствия или отсутствия кислорода.
В двух приведенных выше примерах ферментации НАДН окисляется путем передачи двух электронов пирувату. Однако анаэробные бактерии используют широкий спектр соединений в качестве терминальных акцепторов электронов при клеточном дыхании : азотистые соединения, такие как нитраты и нитриты; соединения серы, такие как сульфаты, сульфиты, диоксид серы и элементарная сера; углекислый газ; соединения железа; соединения марганца; соединения кобальта; и соединения урана.
Аэробная регенерация НАД + и дальнейший катаболизм пирувата
[ редактировать ]У аэробных эукариот развился сложный механизм использования кислорода воздуха в качестве конечного акцептора электронов в процессе, называемом окислительным фосфорилированием . Аэробные прокариоты , у которых отсутствуют митохондрии, используют множество более простых механизмов .
- Во-первых, НАДН + Н + Образующийся в результате гликолиза НАД должен быть передан в митохондрии для окисления и, таким образом, для регенерации НАД. + необходимо для продолжения гликолиза. Однако внутренняя мембрана митохондрий непроницаема для НАДН и НАД. + . [44] Поэтому для транспортировки электронов от НАДН через митохондриальную мембрану используются два «челнока». Это малат-аспартатный челнок и глицеринфосфатный челнок . В первом случае электроны НАДН передаются на цитозольный оксалоацетат с образованием малата . Затем малат проникает через внутреннюю митохондриальную мембрану в митохондриальный матрикс, где повторно окисляется НАД. + образуя внутримитохондриальный оксалоацетат и НАДН. Затем оксалоацетат возвращается в цитозоль путем превращения в аспартат, который легко транспортируется из митохондрии. В глицеринфосфатном челноке электроны от цитозольного НАДН передаются на дигидроксиацетон с образованием глицерин-3-фосфата , который легко проникает через внешнюю митохондриальную мембрану. Затем глицерин-3-фосфат повторно окисляется до дигидроксиацетона, отдавая свои электроны ФАД вместо НАД. + . [44] Эта реакция происходит на внутренней митохондриальной мембране, позволяя FADH 2 отдавать свои электроны непосредственно коферменту Q ( убихинон ), который является частью цепи переноса электронов , которая в конечном итоге передает электроны молекулярному кислороду. O 2 с образованием воды и выделением энергии в конечном итоге захватывается в виде АТФ .
- Конечный продукт гликолиза — пируват (плюс НАД + ) преобразуется в ацетил-КоА , СО 2 и НАДН + Н + внутри митохондрий в процессе, называемом декарбоксилированием пирувата .
- Образующийся ацетил-КоА вступает в цикл лимонной кислоты (или цикл Кребса), где ацетильная группа ацетил-КоА превращается в диоксид углерода в результате двух реакций декарбоксилирования с образованием еще большего количества внутримитохондриального НАДН + Н. + .
- Внутримитохондриальный НАДН + Н + окисляется до НАД + цепью переноса электронов , используя кислород в качестве конечного акцептора электронов для образования воды. Энергия, выделяемая во время этого процесса, используется для создания градиента ионов водорода (или протонов) через внутреннюю мембрану митохондрии .
- Наконец, протонный градиент используется для производства примерно 2,5 АТФ на каждую НАДН + Н. + окисляются в процессе, называемом окислительным фосфорилированием . [44]
Превращение углеводов в жирные кислоты и холестерин
[ редактировать ]Пируват, образующийся в результате гликолиза, является важным посредником в превращении углеводов в жирные кислоты и холестерин . [45] Это происходит путем превращения пирувата в ацетил-КоА в митохондриях . Однако этот ацетил-КоА необходимо транспортировать в цитозоль, где происходит синтез жирных кислот и холестерина. Это не может произойти напрямую. Чтобы получить цитозольный ацетил-КоА, цитрат (полученный при конденсации ацетил-КоА с оксалоацетатом ) удаляется из цикла лимонной кислоты и переносится через внутреннюю митохондриальную мембрану в цитозоль . [45] Там он расщепляется АТФ-цитратлиазой на ацетил-КоА и оксалоацетат. Оксалоацетат возвращается в митохондрии в виде малата (а затем обратно в оксалоацетат для переноса большего количества ацетил-КоА из митохондрии). Цитозольный ацетил-КоА может карбоксилироваться ацетил-КоА-карбоксилазой с образованием малонил-КоА , что является первой стадией синтеза жирных кислот , или он может объединяться с ацетоацетил-КоА с образованием 3-гидрокси-3-метилглутарил-КоА ( HMG). -КоА ), который является лимитирующей стадией, контролирующей синтез холестерина . [46] Холестерин можно использовать как есть, как структурный компонент клеточных мембран, или его можно использовать для синтеза стероидных гормонов , желчных солей и витамина D. [34] [45] [46]
Превращение пирувата в оксалоацетат для цикла лимонной кислоты
[ редактировать ]Молекулы пирувата, образующиеся в результате гликолиза, активно транспортируются через внутреннюю мембрану митохондрий в матрикс, где они могут либо окисляться , либо соединяться с коферментом А с образованием CO 2 , ацетил-КоА и НАДН, [34] или они могут быть карбоксилированы ( пируваткарбоксилазой ) с образованием оксалоацетата . Эта последняя реакция «восполняет» количество оксалоацетата в цикле лимонной кислоты и, следовательно, является анаплеротической реакцией (от греческого слова «заполнять»), увеличивая способность цикла метаболизировать ацетил-КоА, когда ткани нуждаются в энергии ( например, в сердце и скелетных мышцах ) внезапно увеличиваются при активности. [47] В цикле лимонной кислоты все промежуточные соединения (например, цитрат, изоцитрат, альфа-кетоглутарат, сукцинат, фумарат, малат и оксалоацетат) регенерируются во время каждого цикла цикла. Таким образом, добавление большего количества любого из этих промежуточных продуктов в митохондрии означает, что это дополнительное количество сохраняется в цикле, увеличивая количество всех других промежуточных продуктов по мере превращения одного в другое. Следовательно, добавление оксалоацетата значительно увеличивает количество всех промежуточных продуктов лимонной кислоты, тем самым увеличивая способность цикла метаболизировать ацетил-КоА, превращая его ацетатный компонент в CO 2 и воды, с выделением энергии, достаточной для образования 11 молекулы АТФ и 1 молекулы ГТФ на каждую дополнительную молекулу ацетил-КоА, соединяющуюся в цикле с оксалоацетатом. [47]
Чтобы катаплеротически удалить оксалоацетат из цикла лимонных кислот, малат может транспортироваться из митохондрий в цитоплазму, уменьшая количество оксалоацетата, которое может быть регенерировано. [47] Кроме того, промежуточные соединения лимонной кислоты постоянно используются для образования различных веществ, таких как пурины, пиримидины и порфирины . [47]
Промежуточные соединения для других путей
[ редактировать ]Эта статья концентрируется на катаболической роли гликолиза в отношении преобразования потенциальной химической энергии в полезную химическую энергию во время окисления глюкозы до пирувата. Многие из метаболитов гликолитического пути также используются анаболическими путями, и, как следствие, поток через этот путь имеет решающее значение для поддержания запаса углеродных скелетов для биосинтеза. [48]
Следующие метаболические пути в значительной степени зависят от гликолиза как источника метаболитов: и многие другие.
- Пентозофосфатный путь , который начинается с дегидрирования глюкозо-6-фосфата , первого промежуточного продукта, образующегося в результате гликолиза, приводит к образованию различных пентозных сахаров и НАДФН для синтеза жирных кислот и холестерина .
- Синтез гликогена также начинается с глюкозо-6-фосфата в начале гликолитического пути.
- Глицерин для образования триглицеридов и фосфолипидов производится из гликолитического промежуточного продукта глицеральдегид-3-фосфата .
- Различные постгликолитические пути:
- Синтез жирных кислот
- Синтез холестерина
- Цикл лимонной кислоты , который, в свою очередь, приводит к:
Хотя глюконеогенез и гликолиз имеют много общих промежуточных продуктов, один из них функционально не является ветвью или притоком другого. В обоих путях есть два регуляторных этапа, которые, будучи активными в одном пути, автоматически неактивны в другом. Поэтому два процесса не могут быть активными одновременно. [49] Действительно, если бы оба набора реакций были одновременно высокоактивными, конечным результатом был бы гидролиз четырех высокоэнергетических фосфатных связей (двух АТФ и двух ГТФ) за один реакционный цикл. [49]
ОНИ + является окислителем при гликолизе, как и в большинстве других метаболических реакций, дающих энергию (например, бета-окисление жирных кислот и во время цикла лимонной кислоты ). Образующийся таким образом НАДН в основном используется для окончательного переноса электронов на O 2 для получения воды или, когда O 2 недоступен для производства таких соединений, как лактат или этанол (см. Аноксическая регенерация НАД). + выше). НАДН редко используется в синтетических процессах, за исключением глюконеогенеза. В процессе жирных кислот и синтеза холестерина восстановителем является НАДФН . Это различие иллюстрирует общий принцип, согласно которому НАДФН расходуется во время реакций биосинтеза, тогда как НАДН генерируется в реакциях с выделением энергии. [49] Источник НАДФН двоякий. Когда малат окислительно декарбоксилируется «НАДФ + -связанный яблочный фермент пируват , CO 2 Образуются и НАДФН. НАДФН также образуется по пентозофосфатному пути , который превращает глюкозу в рибозу, которая может использоваться в синтезе нуклеотидов и нуклеиновых кислот , или может катаболизироваться до пирувата. [49]
Гликолиз при заболеваниях
[ редактировать ]Диабет
[ редактировать ]Поглощение глюкозы клетками происходит в ответ на сигналы инсулина, а затем глюкоза расщепляется посредством гликолиза, снижая уровень сахара в крови. Однако резистентность к инсулину или низкие уровни инсулина, наблюдаемые при диабете, приводят к гипергликемии, когда уровень глюкозы в крови повышается, и глюкоза не усваивается клетками должным образом. Гепатоциты также способствуют этой гипергликемии посредством глюконеогенеза . Гликолиз в гепатоцитах контролирует выработку глюкозы в печени, и когда глюкоза перепроизводится печенью, не имея возможности расщепляться организмом, возникает гипергликемия. [50]
Генетические заболевания
[ редактировать ]Гликолитические мутации обычно встречаются редко из-за важности метаболического пути; большинство возникающих мутаций приводят к неспособности клетки дышать и, следовательно, вызывают гибель клетки на ранней стадии. Однако некоторые мутации ( болезни накопления гликогена и другие врожденные нарушения углеводного обмена наблюдаются ), одним из ярких примеров которых является дефицит пируваткиназы , приводящий к хронической гемолитической анемии. [ нужна ссылка ]
Рак
[ редактировать ]Клетки злокачественных опухолей осуществляют гликолиз со скоростью, которая в десять раз быстрее, чем их аналоги из нераковых тканей. [51] Во время их возникновения ограниченная капиллярная поддержка часто приводит к гипоксии (снижению снабжения O2) внутри опухолевых клеток. Таким образом, эти клетки полагаются на анаэробные метаболические процессы, такие как гликолиз АТФ (аденозинтрифосфат). Некоторые опухолевые клетки сверхэкспрессируют специфические гликолитические ферменты, что приводит к более высокой скорости гликолиза. [52] Часто эти ферменты представляют собой изоферменты традиционных ферментов гликолиза, которые различаются по своей восприимчивости к традиционному ингибированию по принципу обратной связи. Увеличение гликолитической активности в конечном итоге нейтрализует последствия гипоксии за счет генерации достаточного количества АТФ по этому анаэробному пути. [53] Это явление было впервые описано в 1930 году Отто Варбургом и называется эффектом Варбурга . Гипотеза Варбурга утверждает, что рак в первую очередь вызван нарушением митохондриального метаболизма, а не неконтролируемым ростом клеток.Для объяснения эффекта Варбурга был выдвинут ряд теорий. Одна из таких теорий предполагает, что повышенный гликолиз является нормальным защитным процессом организма и что злокачественные изменения могут быть в первую очередь вызваны энергетическим метаболизмом. [54]
Эта высокая скорость гликолиза имеет важные медицинские применения, поскольку высокий аэробный гликолиз злокачественными опухолями используется клинически для диагностики и мониторинга реакции на лечение рака путем визуализации поглощения 2- 18 F-2-дезоксиглюкоза (ФДГ) ( радиоактивно гексокиназы модифицированный субстрат ) с помощью позитронно-эмиссионной томографии (ПЭТ). [55] [56]
Продолжаются исследования по влиянию на митохондриальный метаболизм и лечению рака путем уменьшения гликолиза и, таким образом, голодания раковых клеток различными новыми способами, включая кетогенную диету . [57] [58] [59]
Интерактивная карта маршрутов
[ редактировать ]На диаграмме ниже показаны названия белков человека. Названия у других организмов могут быть другими, и количество изоферментов (например, HK1, HK2, ...), вероятно, тоже будет другим.
Нажмите на гены, белки и метаболиты ниже, чтобы перейти к соответствующим статьям. [§ 1]

- ^ Интерактивную карту маршрутов можно редактировать на WikiPathways: «Гликолиз-Глюконеогенез_WP534» .
Альтернативная номенклатура
[ редактировать ]Некоторые метаболиты гликолиза имеют альтернативные названия и номенклатуру. Частично это связано с тем, что некоторые из них являются общими для других путей, таких как цикл Кальвина .
| Эта статья | Альтернатива | |||
|---|---|---|---|---|
| 1 | Глюкоза | Глк | Декстроза | |
| 2 | Глюкозо-6-фосфат | G6P | ||
| 3 | Фруктозо-6-фосфат | F6P | ||
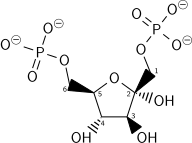
| 4 | Фруктозо-1,6-бисфосфат | Ф1,6БП | Фруктозо-1,6-дифосфат | ФБП; СвДП; Ф1,6ДП |
| 5 | Дигидроксиацетонфосфат | ДГАП | Глицеронфосфат | |
| 6 | Глицеральдегид-3-фосфат | ГАДП | 3-фосфоглицеральдегид | ПГАЛ; Г3П; ГАЛП; зазор; тп |
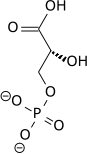
| 7 | 1,3-бисфосфоглицерат | 1,3 БПГ | Глицерат-1,3-бисфосфат, глицерат-1,3-дифосфат, 1,3-дифосфоглицерат | ПГАП; БПГ; ДПГ |
| 8 | 3-фосфоглицерат | 3PG | Глицерат-3-фосфат | ПГА; врач общей практики |
| 9 | 2-фосфоглицерат | 2PG | Глицерат-2-фосфат | |
| 10 | фосфоенолпируват | ПКП | ||
| 11 | Пируват | Пир | Основание, сопряженное с пировиноградной кислотой | |
Структура компонентов гликолиза в проекциях Фишера и полигональной модели
[ редактировать ]Промежуточные продукты гликолиза, изображенные в проекциях Фишера, демонстрируют постепенное химическое изменение. Такое изображение можно сравнить с представлением полигональной модели. [60]

См. также
[ редактировать ]- Катаболизм углеводов
- Цикл лимонной кислоты
- Цикл Кори
- Ферментация (биохимия)
- глюконеогенез
- Гликолитические колебания
- Гликогенозы (болезни накопления гликогена)
- Врожденные нарушения углеводного обмена
- Пентозофосфатный путь
- Декарбоксилирование пирувата
- Триозокиназа
Ссылки
[ редактировать ]- ^ Альфарук К.О., Вердуско Д., Раух С., Муддатир А.К., Адиль Х.Х., Эльхассан Г.О. и др. (18 декабря 2014 г.). «Гликолиз, метаболизм опухоли, рост и распространение рака. Новая этиопатогенетическая перспектива на основе pH и терапевтический подход к старому вопросу рака» . Онсознание . 1 (12): 777–802. doi : 10.18632/oncoscience.109 . ПМК 4303887 . ПМИД 25621294 .
- ^ Романо А.Х., Конвей Т. (1996). «Эволюция путей метаболизма углеводов» . Исследования в области микробиологии . 147 (6–7): 448–455. дои : 10.1016/0923-2508(96)83998-2 . ПМИД 9084754 .
- ^ Келлер М.А., Турчин А.В., Ралсер М. (апрель 2014 г.). «Неферментативный гликолиз и реакции, подобные пентозофосфатному пути, в вероятном архейском океане» . Молекулярная системная биология . 10 (4): 725. doi : 10.1002/msb.20145228 . ПМК 4023395 . ПМИД 24771084 .
- ^ Ким Б.Х., Гэдд GM . (2011) Бактериальная физиология и метаболизм, 3-е издание.
- ^ Jump up to: а б с Мехта С (20 сентября 2011 г.). «Гликолиз – Анимация и примечания» . ФармаXchange . Архивировано из оригинала 25 марта 2012 года . Проверено 22 сентября 2011 г.
- ^ Jump up to: а б Лейн А.Н., Фан Т.В., Хигаши Р.М. (2009). «Метаболический ацидоз и важность сбалансированных уравнений». Метаболомика . 5 (2): 163–165. дои : 10.1007/s11306-008-0142-2 . S2CID 35500999 .
- ^ Барнетт Дж. А. (апрель 2003 г.). «История исследований дрожжей 5: путь ферментации» . Дрожжи . 20 (6): 509–543. дои : 10.1002/да.986 . ПМИД 12722184 . S2CID 26805351 .
- ^ «Луи Пастер и алкогольное брожение» . www.pasteurbrewing.com . Архивировано из оригинала 13 января 2011 г. Проверено 23 февраля 2016 г.
- ^ Альба-Луа Л., Сигал-Кищиневский С (январь 2010 г.). «Дрожжевое брожение и изготовление пива и вина» . Природное образование . 3 (9): 17.
- ^ Колер Р. (1 марта 1971 г.). «Предыстория открытия Эдуардом Бюхнером бесклеточной ферментации». Журнал истории биологии . 4 (1): 35–61. дои : 10.1007/BF00356976 . ПМИД 11609437 . S2CID 46573308 .
- ^ «Эдуард Бюхнер — Биографический» . www.nobelprize.org . Проверено 23 февраля 2016 г.
- ^ Jump up to: а б Корниш-Боуден А, изд. (1997). «Открытие Харденом и Янгом фруктозо-1,6-бисфосфата». Новое пиво в старой бутылке: Эдуард Бюхнер и рост биохимических знаний . Валенсия, Испания: Публикации Университета Валенсии.
- ^ Jump up to: а б Палмер Г. «Глава 3: История гликолиза: пример линейного метаболического пути». Биос 302 (PDF) . Архивировано из оригинала (PDF) 18 ноября 2017 года.
- ^ «Отто Мейергоф — Биографическое» . www.nobelprize.org . Проверено 23 февраля 2016 г.
- ^ Jump up to: а б с Кресге Н., Симони Р.Д., Хилл Р.Л. (январь 2005 г.). «Отто Фриц Мейерхоф и выяснение гликолитического пути» . Журнал биологической химии . 280 (4): e3. дои : 10.1016/S0021-9258(20)76366-0 . PMID 15665335 .
- ^ «Эмбден, Густав - Словарное определение Эмбдена, Густав | Энциклопедия.com: БЕСПЛАТНЫЙ онлайн-словарь» . www.энциклопедия.com . Проверено 23 февраля 2016 г.
- ^ Ривз Р.Э., Южный ди-джей, Блитт Х.Дж., Уоррен Л.Г. (декабрь 1974 г.). «Пирофосфат:D-фруктозо-6-фосфат-1-фосфотрансфераза. Новый фермент с гликолитической функцией 6-фосфофруктокиназы» . Журнал биологической химии . 249 (24): 7737–7741. дои : 10.1016/S0021-9258(19)42029-2 . ПМИД 4372217 .
- ^ Селиг М., Ксавье К.Б., Сантос Х., Шёнхайт П. (апрель 1997 г.). «Сравнительный анализ гликолитических путей Эмбдена-Мейергофа и Энтнера-Дудорова у гипертермофильных архей и бактерии Thermotoga». Архив микробиологии . 167 (4): 217–232. Бибкод : 1997ArMic.167..217S . дои : 10.1007/BF03356097 . ПМИД 9075622 . S2CID 19489719 .
- ^ Гарретт Р.Х., Гришэм К.М. (2012). Биохимия (5-е изд.). Cengage Обучение. ISBN 978-1-133-10629-6 .
- ^ Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2007). Биохимия (6-е изд.). Нью-Йорк: Фриман. п. 622. ИСБН 978-0-7167-8724-2 .
- ^ Jump up to: а б Гарретт Р., Гришэм К.М. (2005). Биохимия (3-е изд.). Бельмонт, Калифорния: Томсон Брукс/Коул. п. 584. ИСБН 978-0-534-49033-1 .
- ^ Симидзу К., Мацуока Ю. (март 2019 г.). «Регуляция гликолитического потока и избыточного метаболизма в зависимости от источника выработки энергии для удовлетворения потребностей в энергии». Достижения биотехнологии . 37 (2): 284–305. doi : 10.1016/j.biotechadv.2018.12.007 . ПМИД 30576718 . S2CID 58591361 .
- ^ Jump up to: а б Чубуков В., Героса Л., Кочановский К., Зауэр У. (май 2014 г.). «Координация микробного метаболизма». Обзоры природы. Микробиология . 12 (5): 327–340. дои : 10.1038/nrmicro3238 . ПМИД 24658329 . S2CID 28413431 .
- ^ Хочачка П.В. (1999). «Межвидовые исследования гликолитической функции». В Roach RC, Wagner PD, Hackett PH (ред.). Гипоксия . Достижения экспериментальной медицины и биологии. Том. 474. Бостон, Массачусетс: Springer US. стр. 219–229. дои : 10.1007/978-1-4615-4711-2_18 . ISBN 978-1-4613-7134-2 . ПМИД 10635004 .
- ^ Лемигр Ф.П., Руссо Г.Г. (октябрь 1994 г.). «Транскрипционный контроль генов, регулирующих гликолиз и глюконеогенез в печени взрослых» . Биохимический журнал . 303 (1): 1–14. дои : 10.1042/bj3030001 . ПМЦ 1137548 . ПМИД 7945228 .
- ^ Бянь Х, Цзян Х, Мэн Ю, Ли ЮП, Фан Дж, Лу Цзи (март 2022 г.). «Регуляция экспрессии генов гликолитическими и глюконеогенными ферментами». Тенденции в клеточной биологии . 32 (9): 786–799. дои : 10.1016/j.tcb.2022.02.003 . ПМИД 35300892 . S2CID 247459973 .
- ^ Jump up to: а б Героза Л., Зауэр У. (август 2011 г.). «Регуляция и контроль метаболических потоков у микробов». Современное мнение в области биотехнологии . 22 (4): 566–575. дои : 10.1016/j.copbio.2011.04.016 . ПМИД 21600757 .
- ^ Чоудхури С., Хеппер С., Лоди М.К., Сайер М.Х., Утц П. (апрель 2021 г.). «Белковый интерактом гликолиза в Escherichia coli » . Протеомы . 9 (2): 16. doi : 10.3390/proteomes9020016 . ПМК 8167557 . ПМИД 33917325 .
- ^ Родионова И.А., Чжан З., Мехла Дж., Гудакр Н., Бабу М., Эмили А. и др. (август 2017 г.). «Белок-фосфоноситель HPr бактериальной фосфотрансферазной системы глобально регулирует энергетический обмен, напрямую взаимодействуя с множеством ферментов Escherichia coli » . Журнал биологической химии . 292 (34): 14250–14257. дои : 10.1074/jbc.M117.795294 . ПМК 5572926 . ПМИД 28634232 .
- ^ Писиткул Т., Патель Н.М., Амадор-Ногез Д. (апрель 2015 г.). «Посттрансляционные модификации как ключевые регуляторы бактериальных метаболических потоков». Современное мнение в микробиологии . 24 : 29–37. дои : 10.1016/j.mib.2014.12.006 . ПМИД 25597444 .
- ^ Jump up to: а б с Коэслаг Дж.Х., Сондерс П.Т., Тербланш Э. (июнь 2003 г.). «Переоценка гомеостата глюкозы в крови, которая всесторонне объясняет комплекс сахарный диабет 2 типа и синдром X» . Журнал физиологии . 549 (Часть 2) (опубликовано в 2003 г.): 333–346. дои : 10.1113/jphysicalol.2002.037895 . ПМК 2342944 . ПМИД 12717005 .
- ^ Jump up to: а б с д и Страйер Л. (1995). «Гликолиз». Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 483–508. ISBN 0-7167-2009-4 .
- ^ Страйер Л. (1995). Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. п. 773. ИСБН 0-7167-2009-4 .
- ^ Jump up to: а б с Воет Д., Воет Дж.Г., Пратт К.В. (2006). Основы биохимии (2-е изд.). John Wiley and Sons, Inc., стр. 547, 556 . ISBN 978-0-471-21495-3 .
- ^ Бейс I, Ньюсхолм Э.А. (октябрь 1975 г.). «Содержание адениновых нуклеотидов, фосфагенов и некоторых гликолитических промежуточных продуктов в покоящихся мышцах позвоночных и беспозвоночных» . Биохимический журнал . 152 (1): 23–32. дои : 10.1042/bj1520023 . ПМЦ 1172435 . ПМИД 1212224 .
- ^ Воет Д., Воет Дж.Г. (2004). Биохимия (3-е изд.). Нью-Йорк: John Wiley & Sons, Inc.
- ^ Лэки Дж (2010). ТИГАР . Оксфордский справочник в Интернете: Издательство Оксфордского университета. ISBN 978-0-19-954935-1 .
- ^ Бенсаад К., Цурута А., Селак М.А., Видал М.Н., Накано К., Бартронс Р. и др. (июль 2006 г.). «TIGAR, p53-индуцируемый регулятор гликолиза и апоптоза» . Клетка . 126 (1): 107–120. дои : 10.1016/j.cell.2006.05.036 . ПМИД 16839880 . S2CID 15006256 .
- ^ «TIGAR TP53 индуцирует регуляторную фосфатазу гликолиза [Homo sapiens (человек)] - Ген - NCBI» . www.ncbi.nlm.nih.gov . Проверено 17 мая 2018 г.
- ^ Карбонелл Дж., Фелиу Дж.Е., Марко Р., Солс А. (август 1973 г.). «Пируваткиназа. Классы регуляторных изоферментов в тканях млекопитающих». Европейский журнал биохимии . 37 (1): 148–156. дои : 10.1111/j.1432-1033.1973.tb02969.x . hdl : 10261/78345 . ПМИД 4729424 .
- ^ Валентини Дж., Кьярелли Л., Фортин Р., Сперанца М.Л., Галицци А., Маттеви А. (июнь 2000 г.). «Аллостерическая регуляция пируваткиназы» . Журнал биологической химии . 275 (24): 18145–18152. дои : 10.1074/jbc.m001870200 . ПМИД 10751408 .
- ^ Исраэльсен В.Дж., Вандер Хайден М.Г. (июль 2015 г.). «Пируваткиназа: функция, регуляция и роль при раке» . Семинары по клеточной биологии и биологии развития . 43 : 43–51. дои : 10.1016/j.semcdb.2015.08.004 . ПМК 4662905 . ПМИД 26277545 .
- ^ Jump up to: а б с д Энгстрем Л. (1978). «Регуляция пируваткиназы печени путем фосфорилирования-дефосфорилирования». Актуальные темы клеточной регуляции . 13 . Эльзевир: 28–51. дои : 10.1016/b978-0-12-152813-3.50006-9 . ISBN 978-0-12-152813-3 . ПМИД 208818 .
- ^ Jump up to: а б с Страйер Л. (1995). «Окислительное фосфорилирование». Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 537–549. ISBN 0-7167-2009-4 .
- ^ Jump up to: а б с Страйер Л. (1995). «Обмен жирных кислот». Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 603–628. ISBN 0-7167-2009-4 .
- ^ Jump up to: а б Страйер Л. (1995). «Биосинтез мембранных липидов и стероидов». Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 691–707. ISBN 0-7167-2009-4 .
- ^ Jump up to: а б с д Страйер Л. (1995). «Цикл лимонной кислоты». Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 509–527, 569–579, 614–616, 638–641, 732–735, 739–748, 770–773. ISBN 0-7167-2009-4 .
- ^ Судья А, Додд М.С. (08 октября 2020 г.). «Метаболизм» . Очерки по биохимии . 64 (4): 607–647. дои : 10.1042/EBC20190041 . ISSN 0071-1365 . ПМК 7545035 . ПМИД 32830223 .
- ^ Jump up to: а б с д Страйер Л. (1995). Биохимия (Четвертое изд.). Нью-Йорк: WH Freeman and Company. стр. 559–565, 574–576, 614–623. ISBN 0-7167-2009-4 .
- ^ Го X, Ли Х, Сюй Х, Ву С, Донг Х, Лу Ф и др. (01 августа 2012 г.). «Гликолиз в контроле гомеостаза глюкозы в крови» . Акта Фармацевтика Синика Б. 2 (4): 358–367. дои : 10.1016/j.apsb.2012.06.002 . ISSN 2211-3835 .
- ^ Альфарук К.О., Вердуско Д., Раух С., Муддатир А.К., Адиль Х.Х., Эльхассан Г.О. и др. (2014). «Гликолиз, метаболизм опухоли, рост и распространение рака. Новая этиопатогенетическая перспектива на основе pH и терапевтический подход к старому вопросу рака» . Онсознание . 1 (12): 777–802. doi : 10.18632/oncoscience.109 . ПМК 4303887 . ПМИД 25621294 .
- ^ Альфарук КО, Шаюб М.Э., Муддатир АК, Эльхассан ГО, Башир А.Х. (июль 2011 г.). «Эволюция опухолевого метаболизма может отражать канцерогенез как процесс обратной эволюции (демонтаж многоклеточности)» . Раки . 3 (3): 3002–3017. дои : 10.3390/cancers3033002 . ПМЦ 3759183 . ПМИД 24310356 .
- ^ Нельсон Д.Л., Кокс М.М. (2005). Ленингерские принципы биохимии (4-е изд.). Нью-Йорк: WH Freeman. ISBN 978-0-7167-4339-2 .
- ^ Голд J (октябрь 2011 г.). «Что такое рак?» . Архивировано из оригинала 19 мая 2018 года . Проверено 8 сентября 2012 г.
- ^ Паувелс Э.К., Штурм Э.Дж., Бомбардьери Э., Клетон Ф.Дж., член парламента Стоккеля (октябрь 2000 г.). «Позитронно-эмиссионная томография с [18F]фтордезоксиглюкозой. Часть I. Механизм биохимического поглощения и его значение для клинических исследований». Журнал исследований рака и клинической онкологии . 126 (10): 549–59. дои : 10.1007/pl00008465 . ПМИД 11043392 . S2CID 2725555 .
- ^ «ПЭТ-сканирование: информация о ПЭТ-сканировании раскрывает ...» Проверено 5 декабря 2005 г. .
- ^ Шварц Л., Сейфрид Т., Альфарук КО, Да Вейга Морейра Дж., Фейс С. (апрель 2017 г.). «Эффект Варбурга: эффективное лечение рака, направленное на специфический метаболизм опухоли и нарушение регуляции pH». Семинары по биологии рака . 43 : 134–138. doi : 10.1016/j.semcancer.2017.01.005 . ПМИД 28122260 .
- ^ Шварц Л., Супуран КТ, Альфарук КО (2017). «Эффект Варбурга и признаки рака». Противораковые агенты в медицинской химии . 17 (2): 164–170. дои : 10.2174/1871520616666161031143301 . ПМИД 27804847 .
- ^ Марун Дж., Бост Дж., Амос А., Зукколи Дж. (август 2013 г.). «Кетогенная диета с ограничением калорий для лечения мультиформной глиобластомы». Журнал детской неврологии . 28 (8): 1002–1008. дои : 10.1177/0883073813488670 . ПМИД 23670248 . S2CID 1994087 .
- ^ Бонафе CF, Биспо ХА, де Хесус МБ (январь 2018 г.). «Многоугольная модель: простое представление биомолекул как инструмент обучения метаболизму» . Образование в области биохимии и молекулярной биологии . 46 (1): 66–75. дои : 10.1002/bmb.21093 . ПМИД 29131491 . S2CID 31317102 .
Внешние ссылки
[ редактировать ]- Подробная анимация гликолиза, предоставленная IUBMB ( требуется Adobe Flash )
- Гликолитические ферменты гликолиза в RCSB PDB
- Гликолитический цикл с анимацией на wdv.com
- Метаболизм, клеточное дыхание и фотосинтез - Виртуальная библиотека биохимии, молекулярной биологии и клеточной биологии
- Химическая логика гликолиза на ufp.pt
- Плакат о биохимических путях Expasy на выставке ExPASy
- Medical Mnemonics .com : 317 5468
- metpath : Интерактивное представление гликолиза
метаболизма Карта |
|---|

| Базы данных органов управления : Национальные |
|---|