Фосфофруктокиназа 1
| 6-фосфофруктокиназа | |||
|---|---|---|---|
 | |||
| Идентификаторы | |||
| Номер ЕС. | 2.7.1.11 | ||
| Номер CAS. | 9001-80-3 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||
| Фосфофруктокиназа | |||
|---|---|---|---|
| Идентификаторы | |||
| Символ | ПФК | ||
| Пфам | PF00365 | ||
| Пфам Клан | CL0240 | ||
| ИнтерПро | IPR000023 | ||
| PROSITE | PDOC00336 | ||
| СКОП2 | 5пфк / СКОПе / СУПФАМ | ||
| |||
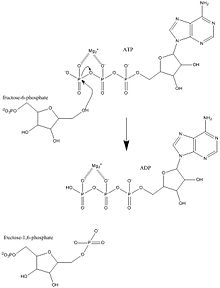
Фосфофруктокиназа-1 ( ПФК-1 ) — один из важнейших регуляторных ферментов ( EC 2.7.1.11 ) гликолиза . Это аллостерический фермент, состоящий из 4 субъединиц и контролируемый множеством активаторов и ингибиторов . ПФК-1 катализирует важную «совершенную» стадию гликолиза, превращение фруктозо-6-фосфата и АТФ во фруктозо-1,6-бисфосфат и АДФ . Гликолиз является основой дыхания, как анаэробного, так и аэробного. Поскольку фосфофруктокиназа (ФФК) катализирует АТФ-зависимое фосфорилирование с превращением фруктозо-6-фосфата во фруктозо-1,6-бисфосфат и АДФ, это один из ключевых регуляторных этапов гликолиза. [1] ПФК способна регулировать гликолиз посредством аллостерического ингибирования, и таким образом клетка может увеличивать или уменьшать скорость гликолиза в ответ на энергетические потребности клетки. Например, высокое соотношение АТФ и АДФ будет ингибировать ПФК и гликолиз. Ключевое различие между регуляцией ПФК у эукариот и прокариот заключается в том, что у эукариот ПФК активируется фруктозо-2,6-бисфосфатом. Целью фруктозо-2,6-бисфосфата является преодоление ингибирования АТФ, что позволяет эукариотам иметь большую чувствительность к регуляции с помощью таких гормонов, как глюкагон и инсулин. [2]
| β- D - фруктозо-6-фосфат | Фосфофруктокиназа 1 | β- D - фруктозо-1,6-бисфосфат | |
 |  | ||
| СПС | АДП | ||
| П я | Н 2 О | ||
| Фруктозобисфосфатаза | |||
Структура
[ редактировать ]У млекопитающих PFK1 весит 340 кг. [3] тетрамер состоит из различных комбинаций трех типов субъединиц: мышечной (М), печени (L) и тромбоцитов (Р). Состав тетрамера PFK1 различается в зависимости от типа ткани, в которой он присутствует. Например, зрелая мышца экспрессирует только изозим M , следовательно, мышечная PFK1 состоит исключительно из гомотетрамеров M4. Печень и почки экспрессируют преимущественно изоформу L. В эритроцитах субъединицы M и L случайным образом тетрамеризуются с образованием M4, L4 и трех гибридных форм фермента (ML3, M2L2, M3L). В результате кинетические и регуляторные свойства различных пулов изоферментов зависят от состава субъединиц. Тканеспецифические изменения активности ПФК и содержания изоферментов в значительной степени способствуют различиям скоростей гликолитического и глюконеогенного действия , которые наблюдаются для разных тканей. [4]
PFK1 является аллостерическим ферментом и имеет структуру, аналогичную структуре гемоглобина , поскольку он представляет собой димер димера. [5] Одна половина каждого димера содержит сайт связывания АТФ, тогда как другая половина — сайт связывания субстрата (фруктозо-6-фосфат или (F6P)), а также отдельный аллостерический сайт связывания. [6]
Каждая субъединица тетрамера состоит из 319 аминокислот и состоит из двух доменов: один связывает субстрат АТФ, а другой связывает фруктозо-6-фосфат. Каждый домен представляет собой бета-цилиндр и имеет цилиндрический бета-лист, окруженный альфа-спиралями.
На противоположной стороне каждой субъединицы от каждого активного сайта находится аллостерический сайт, на границе раздела субъединиц в димере. ATP и AMP конкурируют за этот сайт. N-концевой домен играет каталитическую роль, связывая АТФ, а C-концевой — регуляторную роль. [7]
Механизм
[ редактировать ]PFK1 — аллостерический фермент, активность которого можно описать с помощью симметричной модели аллостеризма. [8] при этом происходит согласованный переход из ферментативно неактивного Т-состояния в активное R-состояние. F6P с высоким сродством связывается с ферментом в состоянии R, но не с ферментом в состоянии T. Для каждой молекулы F6P, которая связывается с PFK1, фермент постепенно переходит из состояния T в состояние R. Таким образом, график, отображающий активность PFK1 в зависимости от увеличения концентрации F6P, будет иметь форму сигмоидальной кривой, традиционно связанной с аллостерическими ферментами.
PFK1 принадлежит к семейству фосфотрансфераз и катализирует перенос γ-фосфата из АТФ во фруктозо-6-фосфат. PFK1 Активный сайт включает сайты связывания ATP-Mg2+ и F6P. Некоторые предполагаемые остатки, участвующие в связывании субстрата в PFK1 E. coli , включают Asp127 и Arg171 . [9] У B. stearothermophilus PFK1 положительно заряженная боковая цепь остатка Arg162 образует солевой мостик с водородными связями с отрицательно заряженной фосфатной группой F6P, взаимодействие, которое стабилизирует состояние R относительно состояния T и частично отвечает за гомотропный эффект. связывания F6P. В состоянии Т конформация фермента слегка смещается, так что пространство, ранее занимаемое Arg162, заменяется Glu161 . Эта замена позиций между соседними аминокислотными остатками ингибирует способность F6P связываться с ферментом.
Аллостерические активаторы, такие как АМФ и АДФ, связываются с аллостерическим сайтом, чтобы облегчить формирование состояния R, вызывая структурные изменения в ферменте. Аналогично, ингибиторы, такие как АТФ и ПЭП, связываются с одним и тем же аллостерическим сайтом и способствуют образованию Т-состояния, тем самым ингибируя активность фермента.
Гидроксильный кислород углерода 1 осуществляет нуклеофильную атаку на бета-фосфат АТФ. Эти электроны отодвигаются к ангидридному кислороду между бета- и гамма-фосфатами АТФ. [10] [11]
Регулирование
[ редактировать ]PFK1 является наиболее важным центром контроля гликолитического пути млекопитающих. Этот этап подлежит тщательному регулированию, поскольку он не только высоко экзергоничен в физиологических условиях , но и потому, что это обязательный этап – первая необратимая реакция, уникальная для гликолитического пути. Это приводит к точному контролю глюкозы и других моносахаридов, галактозы и фруктозы, идущих по гликолитическому пути. До реакции этого фермента глюкозо-6-фосфат потенциально может перемещаться по пентозофосфатному пути или превращаться в глюкозо-1-фосфат для гликогенеза .
PFK1 аллостерически ингибируется высокими уровнями АТФ , но AMP обращает ингибирующее действие АТФ. Следовательно, активность фермента увеличивается при снижении клеточного соотношения АТФ/АМФ. Таким образом, гликолиз стимулируется, когда энергетический заряд падает. PFK1 имеет два сайта с разным сродством к АТФ, который является одновременно субстратом и ингибитором. [3]
PFK1 также ингибируется низким уровнем pH, что усиливает ингибирующий эффект АТФ. Уровень pH падает, когда мышцы функционируют анаэробно и производят избыточное количество молочной кислоты (хотя молочная кислота сама по себе не является причиной снижения pH). [12] ). Этот ингибирующий эффект служит для защиты мышц от повреждений, которые могут возникнуть в результате накопления слишком большого количества кислоты. [3]
Наконец, PFK1 аллостерически ингибируется PEP , цитратом и АТФ. Фосфоенолпировиноградная кислота является продуктом, находящимся дальше по гликолитическому пути. Хотя цитрат действительно накапливается, когда ферменты цикла Кребса достигают максимальной скорости, сомнительно, накапливается ли цитрат до концентрации, достаточной для ингибирования PFK-1 в нормальных физиологических условиях. [ нужна ссылка ] . Увеличение концентрации АТФ указывает на избыток энергии и имеет место аллостерической модуляции на PFK1, где снижается сродство PFK1 к ее субстрату.
PFK1 аллостерически активируется высокой концентрацией AMP , но наиболее мощным активатором является фруктозо-2,6-бисфосфат , который также вырабатывается из фруктозо-6-фосфата с помощью PFK2 . Следовательно, обилие F6P приводит к более высокой концентрации фруктозо-2,6-бисфосфата (F-2,6-BP). Связывание F-2,6-BP увеличивает сродство PFK1 к F6P и уменьшает ингибирующий эффект АТФ. Это пример прямой стимуляции, поскольку гликолиз ускоряется при избытке глюкозы. [3]
Активность ПФК снижается за счет подавления синтеза глюкагоном . Глюкагон активирует протеинкиназу А, которая, в свою очередь, отключает киназную активность PFK2 . Это обращает вспять любой синтез F-2,6-BP из F6P и, таким образом, деактивирует PFK1.
Точная регуляция PFK1 предотвращает гликолиза и глюконеогенеза одновременное протекание происходит циклическое изменение субстратов . Однако между F6P и F-1,6-BP . Фруктозо-1,6-бисфосфатаза (FBPase) катализирует гидролиз F-1,6-BP обратно до F6P, обратную реакцию, катализируемую PFK1. Существует небольшая активность FBPase во время гликолиза и некоторая активность PFK1 во время глюконеогенеза. Этот цикл позволяет усиливать метаболические сигналы, а также выделять тепло за счет гидролиза АТФ.
Серотонин (5-HT) увеличивает PFK путем связывания с рецептором 5-HT(2A), вызывая фосфорилирование тирозинового остатка PFK с помощью фосфолипазы C. Это, в свою очередь, перераспределяет PFK внутри клеток скелетных мышц. Поскольку PFK регулирует гликолитический поток, серотонин играет регулирующую роль в гликолизе. [13]
Гены
[ редактировать ]У человека есть три гена фосфофруктокиназы:
Клиническое значение
[ редактировать ]Генетическая мутация в гене PFKM приводит к болезни Таруи , болезни накопления гликогена, при которой способность определенных типов клеток использовать углеводы в качестве источника энергии. нарушается [14]
Болезнь Таруи представляет собой болезнь накопления гликогена, симптомы которой включают мышечную слабость (миопатию), судороги и спазмы, вызванные физической нагрузкой, миоглобинурию (наличие миоглобина в моче, что указывает на разрушение мышц) и компенсированный гемолиз. АТФ является естественным аллостерическим ингибитором ПФК, предотвращающим ненужное производство АТФ посредством гликолиза. Однако мутация в Asp(543)Ala может привести к более сильному ингибирующему эффекту АТФ (из-за повышенного связывания с ингибирующим аллостерическим сайтом связывания PFK). [15] [16]
Мутация фосфофруктокиназы и рак. Чтобы раковые клетки могли удовлетворить свои энергетические потребности благодаря быстрому росту и делению клеток, они выживают более эффективно, если у них есть гиперактивный фермент фосфофруктокиназа 1. [17] [18] Когда раковые клетки растут и делятся быстро, они изначально не имеют достаточного кровоснабжения и, таким образом, могут испытывать гипоксию (недостаток кислорода), и это запускает O-GlcNAcylation по серину 529 PFK. Эта модификация ингибирует активность PFK1 и поддерживает пролиферацию рака, в отличие от мнения, что для рака необходима высокая активность PFK1. Это может быть связано с перенаправлением потока глюкозы на пентозофосфатный путь для генерации НАДФН для детоксикации активных форм кислорода. [19]
Простой герпес типа 1 и фосфофруктокиназа: некоторые вирусы, включая ВИЧ, ЦМВ и Майаро, влияют на клеточные метаболические пути, такие как гликолиз, путем MOI-зависимого увеличения активности ПФК. Механизм повышения активности PFK при герпесе заключается в фосфорилировании фермента по остаткам серина. Гликолиз, индуцированный ВПГ-1, увеличивает содержание АТФ, что имеет решающее значение для репликации вируса. [20]
См. также
[ редактировать ]- PFK2 (превращает фруктозо-6-фосфат во фруктозо-2,6-бисфосфат на одном сайте или, наоборот, на другом сайте)
- PFP (обратимо преобразует фруктозо-6-фосфат и фруктозо-1,6-бисфосфат с использованием неорганического пирофосфата , а не АТФ)
- Фруктозобисфосфатаза (гидролизует фруктозо-1,6-бисфосфат до фруктозо-6-фосфата)
Ссылки
[ редактировать ]- ^ Уокер, ЛР; Симкок, округ Колумбия; Педли, КК; Симпсон, HV ; Браун, С. (апрель 2012 г.). «Кинетика и регуляция фосфофруктокиназы из Teladorsagiacircumcincta» . Экспериментальная паразитология . 130 (4): 348–353. дои : 10.1016/j.exppara.2012.02.011 . ISSN 0014-4894 .
- ^ Усеник А, Легиша М (ноябрь 2010 г.). Кобе Б. (ред.). «Эволюция аллостерических сайтов связывания цитрата на 6-фосфофрукто-1-киназы» . ПЛОС ОДИН . 5 (11): 677–683. дои : 10.1371/journal.pone.0015447 . ПМК 2990764 . ПМИД 21124851 .
- ^ Jump up to: а б с д Страйер Л., Берг Дж.М., Тимочко Дж.Л. (2007). Биохимия (Шестое изд.). Сан-Франциско: WH Freeman. ISBN 978-0-7167-8724-2 .
- ^ Данауэй Г.А., Кастен Т.П., Себо Т., Трапп Р. (май 1988 г.). «Анализ субъединиц и изоферментов фосфофруктокиназы в тканях человека» . Биохим. Дж . 251 (3): 677–83. дои : 10.1042/bj2510677 . ПМК 1149058 . ПМИД 2970843 .
- ^ PDB : 4pfk ; Эванс П.Р., Фаррантс Г.В., Хадсон П.Дж. (июнь 1981 г.). «Фосфофруктокиназа: структура и контроль» . Философские труды Королевского общества Б. 293 (1063): 53–62. дои : 10.1098/rstb.1981.0059 . ПМИД 6115424 .
- «Гликолитические ферменты» . Банк данных по белкам . Архивировано из оригинала 12 декабря 2010 г.
- ^ Ширакихара Ю., Эванс П.Р. (декабрь 1988 г.). «Кристаллическая структура комплекса фосфофруктокиназы Escherichia coli с продуктами его реакции». Дж. Мол. Биол . 204 (4): 973–94. дои : 10.1016/0022-2836(88)90056-3 . ПМИД 2975709 .
- ^ Банашак К., Мечин И., Обмолова Г., Олдхэм М., Чанг Ш., Руис Т., Радермахер М., Коппершлегер Г., Рыпневски В. (март 2011 г.). «Кристаллические структуры эукариотических фосфофруктокиназ пекарских дрожжей и скелетных мышц кролика». Дж Мол Биол . 407 (7): 284–97. дои : 10.1016/j.jmb.2011.01.019 . ПМИД 21241708 .
- ^ Песков К., Горянин И., Демин О. (август 2008 г.). «Кинетическая модель фосфофруктокиназы-1 из Escherichia coli». Журнал Биоинформ Компьютерная Биол . 6 (4): 843–67. дои : 10.1142/S0219720008003643 . ПМИД 18763746 .
- ^ Хеллинга HW, Эванс PR (1987). «Мутации в активном центре фосфофруктокиназы Escherichia coli». Природа . 327 (6121): 437–9. дои : 10.1038/327437a0 . ПМИД 2953977 . S2CID 4357039 .
- ^ Фонг Вайоминг, Лин В., Рао С.П., Дик Т., Алонсо С., Пете К. (август 2013 г.). Пэриш Т. (ред.). «Характеристика активности фосфофруктокиназы в микобактериях туберкулеза показывает, что функциональный гликолитический поток углерода необходим для ограничения накопления токсичных метаболических промежуточных продуктов в условиях гипоксии» . ПЛОС ОДИН . 8 (2): 1198–206. дои : 10.1371/journal.pone.0056037 . ПМК 3567006 . ПМИД 23409118 .
- ^ Папаянни М., Аврамидис Н. (май 2012 г.). «Разработка центральных путей в Lactococcus Lactis: функциональная экспрессия генов фосфофруктокиназы (pfk) и альтернативной оксидазы (aox1) из Aspergillus niger в Lactococcus Lactis способствует повышению скорости конверсии углерода в окислительных условиях». Ферментные и микробные технологии . 51 (113): 125–30. doi : 10.1016/j.enzmictec.2012.04.007 . ПМИД 22759530 .
- ^ Линдингер, Майкл И.; Ковальчук, Джон М.; Хайгенхаузер, Джордж Дж. Ф. (1 сентября 2005 г.). «Применение физико-химических принципов к кислотно-щелочному состоянию скелетных мышц» . Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 289 (3): R891–R894. дои : 10.1152/ajpregu.00225.2005 . ISSN 0363-6119 . ПМИД 16105823 .
- ^ Коэльо В.С., Сола-Пенна М. (январь 2013 г.). «Серотонин регулирует активность 6-фосфофрукто-1-киназы в сигнальном пути, зависимом от PLC-PKC-CaMK II и Янус-киназы». Мол. Клетка. Биохим . 372 (1–2): 211–20. дои : 10.1007/s11010-012-1462-0 . ПМИД 23010892 . S2CID 14570273 .
- ^ Накадзима Х., Рабен Н., Хамагути Т., Ямасаки Т. (март 2002 г.). «Дефицит фосфофруктокиназы; прошлое, настоящее и будущее». Курс. Мол. Мед . 2 (2): 197–212. дои : 10.2174/1566524024605734 . ПМИД 11949936 .
- ^ Брюзер А., Кирхбергер Дж., Шенеберг Т. (октябрь 2012 г.). «Измененная аллостерическая регуляция мышечной 6-фосфофруктокиназы вызывает болезнь Таруи». Биохимия Биофиз Рес Коммьюнити . 427 (1): 133–7. дои : 10.1016/j.bbrc.2012.09.024 . ПМИД 22995305 .
- ^ Брюзер А., Кирхбергер Дж., Шенеберг Т. (октябрь 2012 г.). «Измененная аллостерическая регуляция мышечной 6-фосфофруктокиназы вызывает болезнь Таруи». Биохим. Биофиз. Рез. Коммун . 427 (1): 133–7. дои : 10.1016/j.bbrc.2012.09.024 . ПМИД 22995305 .
- ^ Гомес Л.С., Занкан П., Маркондес М.К., Рамос-Сантос Л., Мейер-Фернандес-младший, Сола-Пенна М., Да Силва Д. (февраль 2013 г.). «Ресвератрол снижает жизнеспособность клеток рака молочной железы и метаболизм глюкозы, ингибируя 6-фосфофрукто-1-киназу» . Биохимия . 95 (6): 1336–43. дои : 10.1016/j.biochi.2013.02.013 . ПМИД 23454376 .
- ^ Ваз К.В., Алвес М.Г., Маркес Р., Морейра П.И., Оливейра П.Ф., Майя К.Дж., Сокорро С. (февраль 2013 г.). «Клетки рака предстательной железы, реагирующие и не реагирующие на андрогены, имеют особый профиль гликолитического метаболизма». Int J Biochem Cell Biol . 44 (11): 2077–84. дои : 10.1016/j.biocel.2012.08.013 . ПМИД 22964025 .
- ^ Йи В., Кларк П.М., Мейсон Д.Е., Кинан М.К., Хилл С., Годдард В.А., Питерс Э.К., Дриггерс Э.М., Се-Уилсон Л.С. (август 2012 г.). «Гликозилирование фосфофруктокиназы 1 регулирует рост и метаболизм клеток» . Наука . 337 (6097): 975–80. дои : 10.1126/science.1222278 . ПМЦ 3534962 . ПМИД 22923583 .
- ^ Абрантес Х.Л., Алвес ЦМ, Коста Х., Алмейда Ф.К., Сола-Пенна М., Фонтес К.Ф., Соуза Т.М. (август 2012 г.). «Простой герпес 1 типа активирует гликолиз посредством участия фермента 6-фосфофрукто-1-киназы (PFK-1)» . Биохим Биофиз Акта . 1822 (8): 1198–206. дои : 10.1016/j.bbadis.2012.04.011 . ПМИД 22542512 .
Внешние ссылки
[ редактировать ]- Фосфофруктокиназа-1 в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- Proteopedia.org Фосфофруктокиназа
- Обзор всей структурной информации, доступной в PDB для UniProt : P08237 (АТФ-зависимая 6-фосфофруктокиназа, мышечный тип) в PDBe-KB .
- Обзор всей структурной информации, доступной в PDB для UniProt : Q01813 (АТФ-зависимая 6-фосфофруктокиназа, тромбоцитарный тип) на PDBe-KB .









