О -GlcNAc
O -GlcNAc (сокращение от O -linked GlcNAc или O -linked β- - ацетилглюкозамин ) представляет собой обратимую ферментативную посттрансляционную модификацию , которая обнаруживается на серина и треонина остатках нуклеоцитоплазматических N белков . Модификация характеризуется наличием β-гликозидной связи между гидроксильной группой боковых цепей серина или треонина и N -ацетилглюкозамином (GlcNAc) . O -GlcNAc отличается от других форм гликозилирования белков : (i) O -GlcNAc не удлиняется и не модифицируется с образованием более сложных гликановых структур, (ii) O -GlcNAc почти исключительно обнаруживается на ядерных и цитоплазматических белках, а не на мембранных белках и секреторных белках. белки , и (iii) O -GlcNAc представляет собой высокодинамическую модификацию, которая меняется быстрее, чем белки, которые она модифицирует. O -GlcNAc консервативен у многоклеточных животных . [1]

Благодаря динамической природе O -GlcNAc и его присутствию на остатках серина и треонина, O сходен с фосфорилированием белка -GlcNAcylation в некоторых отношениях . Хотя существует около 500 киназ и 150 фосфатаз , которые регулируют фосфорилирование белков у человека, есть только два фермента, которые регулируют цикл O -GlcNAc: O -GlcNAc-трансфераза (OGT) и O -GlcNAcase (OGA), катализирующие добавление и удаление O- GlcNAc. O -GlcNAc соответственно. [2] OGT использует UDP-GlcNAc в качестве донорного сахара для переноса сахара. [3]
Впервые об этой посттрансляционной модификации было сообщено в 1984 году. С тех пор она была обнаружена более чем в 5000 белках. [4] [5] многочисленных функциональных ролях O Сообщалось о -GlcNAcylation, включая перекрестное взаимодействие с фосфорилированием серина/треонина, регуляцию белок-белковых взаимодействий , изменение структуры белка или активности фермента, изменение субклеточной локализации белка и модуляцию стабильности и деградации белка . [1] [6] Было идентифицировано, что многочисленные компоненты клеточного транскрипционного аппарата модифицируются O -GlcNAc, и во многих исследованиях сообщается о связи между O -GlcNAc, транскрипцией и эпигенетикой . [7] [8] -GlcNAc влияет на многие другие клеточные процессы, O такие как апоптоз , клеточный цикл и реакции на стресс . [9] Поскольку UDP-GlcNAc является конечным продуктом пути биосинтеза гексозамина, который объединяет метаболизм аминокислот , углеводов , жирных кислот и нуклеотидов , было высказано предположение, что O -GlcNAc действует как « сенсор питательных веществ » и реагирует на метаболический статус клетки. . [10] Нарушение регуляции O -GlcNAc вовлечено во многие патологии, включая болезнь Альцгеймера , рак , диабет и нейродегенеративные расстройства . [11]
Открытие
[ редактировать ]
В 1984 году лаборатория Харта исследовала терминальные остатки GlcNAc на поверхности тимоцитов и лимфоцитов . [ Для радиоактивного мечения UDP- 3 Н]галактоза. β-элиминирование остатков серина и треонина показало, что большая часть [ 3 Н]галактоза присоединялась к белкам О -гликозидно; хроматография показала, что основным продуктом β-элиминирования является Galβ1-4GlcNAcitol. Нечувствительность к лечению пептидом N- гликозидазой предоставила дополнительные доказательства существования O -связанного GlcNAc. Пермеабилизация клеток детергентом перед введением радиоактивной метки значительно увеличивала количество [ 3 H]галактоза включена в Galβ1-4GlcNAcitol, что привело авторов к выводу, что большинство O -связанных моносахаридных остатков GlcNAc находятся внутриклеточно. [12]
Механизм
[ редактировать ]
O -GlcNAc обычно представляет собой динамическую модификацию, которую можно включать и выключать из различных белков. Считается, что некоторые остатки конститутивно модифицируются O -GlcNAc. [13] [14] Модификация O -GlcNAc устанавливается с помощью OGT по последовательному механизму bi-bi , где донорный сахар, UDP-GlcNAc, сначала связывается с OGT, а затем с белком-субстратом. [15] Модификация O -GlcNAc удаляется с помощью OGA по механизму гидролиза, включающему анхимерное содействие (субстратный катализ) с получением немодифицированного белка и GlcNAc. [16] Хотя кристаллических структурах как для OGT, так и для OGT. сообщалось о [15] и ОГА, [17] [18] точные механизмы, с помощью которых OGT и OGA распознают субстраты, полностью не выяснены. В отличие от N -связанного гликозилирования , при котором гликозилирование происходит в специфической консенсусной последовательности (Asn-X-Ser/Thr, где X представляет собой любую аминокислоту, кроме Pro), для O -GlcNAc не было идентифицировано никакой окончательной консенсусной последовательности. Следовательно, предсказание сайтов модификации O -GlcNAc является сложной задачей, а идентификация сайтов модификации обычно требует методов масс-спектрометрии . Что касается OGT, исследования показали, что распознавание субстрата регулируется рядом факторов, включая аспартат. [19] и аспарагин [20] лестничные мотивы в просвете суперспирального домена TPR , остатки активного центра, [21] и адаптерные белки. [22] Поскольку кристаллические структуры показали, что OGT требует, чтобы его подложка находилась в вытянутой конформации, было высказано предположение, что OGT отдает предпочтение гибким подложкам. [21] В кинетических экспериментах in vitro по измерению активности OGT и OGA на панели белковых субстратов было показано, что кинетические параметры OGT варьируются между различными белками, в то время как кинетические параметры OGA были относительно постоянными между различными белками. Этот результат предполагает, что OGT является «старшим партнером» в регуляции O -GlcNAc, а OGA в первую очередь распознает субстраты через присутствие O -GlcNAc, а не через идентичность модифицированного белка. [13]

Обнаружение и характеристика
[ редактировать ]Существует несколько методов обнаружения присутствия O -GlcNAc и характеристики конкретных модифицированных остатков.
Лектины
[ редактировать ]Агглютинин зародышей пшеницы , растительный лектин , способен распознавать концевые остатки GlcNAc и поэтому часто используется для обнаружения O -GlcNAc. Этот лектин применялся в лектиновой аффинной хроматографии для обогащения и обнаружения O -GlcNAc. [23]
Антитела
[ редактировать ]Pan- O -GlcNAc Обычно используются антитела , которые распознают модификацию O -GlcNAc в основном независимо от идентичности модифицированного белка. К ним относятся RL2, [24] антитело IgG , выработанное против белков O -GlcNAcylated комплекса ядерных пор , и CTD110.6, [25] антитело IgM , выработанное против иммуногенного пептида с единственной модификацией серина O -GlcNAc. и другие O -GlcNAc-специфичные антитела имеют некоторую зависимость от идентичности модифицированного белка. Сообщалось, что [26]
Метаболическая маркировка
[ редактировать ]-GlcNAc было разработано множество метаболических химических репортеров Для идентификации O . Метаболические химические репортеры обычно представляют собой аналоги сахаров, которые несут дополнительный химический фрагмент, обеспечивающий дополнительную реакционную способность. Например, перацетилированный GlcNAc (Ac 4 GlcNAz) представляет собой проницаемый для клеток азидосахар , который внутриклеточно деэстерифицируется эстеразами до GlcNAz и превращается в UDP-GlcNAz по пути утилизации гексозамина. UDP-GlcNAz может использоваться в качестве донора сахара с помощью OGT для получения модификации O -GlcNAz. [27] Присутствие азидосахара затем можно визуализировать с помощью биоортогональных алкинсодержащих химических зондов в реакции азид-алкинового циклоприсоединения . Эти зонды могут включать в себя легко идентифицируемые метки, такие как пептид FLAG , биотин и молекулы красителей . [27] [28] Массовые метки на основе полиэтиленгликоля (ПЭГ) также использовались для измерения стехиометрии O -GlcNAc. Конъюгация молекул ПЭГ массой 5 кДа приводит к сдвигу массы модифицированных белков - более сильно O -GlcNAцилированные белки будут иметь несколько молекул ПЭГ и, таким образом, мигрировать медленнее при гель-электрофорезе . [29] Сообщалось о других метаболических химических репортерах, содержащих азиды или алкины (обычно во 2 или 6 положениях). [30] Вместо аналогов GlcNAc можно использовать аналоги GalNAc, поскольку UDP-GalNAc находится в равновесии с UDP-GlcNAc в клетках благодаря действию UDP-галактозо-4'-эпимеразы (GALE) . обработка Ac 4 Обнаружено, что GalNAz приводит к усилению мечения O -GlcNAc по сравнению с Ac 4 GlcNAz, возможно, из-за узкого места в пирофосфорилазе UDP-GlcNAc, обрабатывающей GlcNAz-1-P до UDP-GlcNAz. [31] Было показано, что Ac 3 GlcN-β-Ala-NBD-α-1-P(Ac-SATE) 2 , метаболический химический репортер, который внутриклеточно обрабатывается до меченного флуорофором аналога UDP-GlcNAc, обеспечивает одноэтапное флуоресцентное мечение. - GlcNAc O в живых клетках. [32]
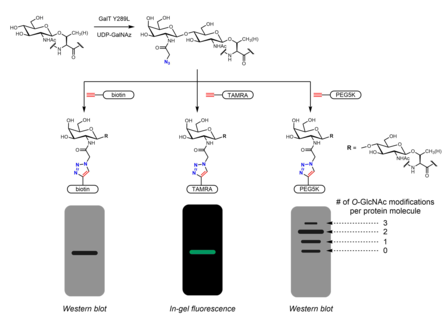
Метаболическое мечение также можно использовать для идентификации партнеров по связыванию O -GlcNAцилированных белков. -ацетильная группа N может быть удлинена с включением диазиринового фрагмента. Обработка клеток перацетилированным, фосфат-защищенным Ac 3 GlcNDAz-1-P(Ac-SATE) 2 приводит к модификации белков O -GlcNDAz. Затем УФ-облучение индуцирует фотосшивку между белками, несущими модификацию O -GlcNDaz, и взаимодействующими белками. [33]
Некоторые проблемы были выявлены с различными метаболическими химическими репортерами, например, их использование может ингибировать путь биосинтеза гексозамина, [30] они могут не распознаваться OGA и, следовательно, не способны улавливать циклический цикл O -GlcNAc, [34] или они могут быть включены в модификации гликозилирования помимо O -GlcNAc, как это наблюдается в секретируемых белках. [35] Метаболические химические репортеры с химическими метками в N -ацетильном положении также могут маркировать ацетилированные белки , поскольку ацетильная группа может гидролизоваться до аналогов ацетата, которые можно использовать для ацетилирования белков. [36] Кроме того, было обнаружено, что пер- O -ацетилированные моносахариды реагируют с цистеинами, что приводит к искусственному S -гликозилированию. [37] Это происходит по механизму исключения-добавления. [38]
Химиоферментная маркировка
[ редактировать ]Хемоферментативное мечение представляет собой альтернативную стратегию включения маркеров для химического анализа щелчков . -GlcNAc Click-IT Система ферментативной маркировки O , разработанная группой Hsieh-Wilson и впоследствии коммерциализированная Invitrogen , использует мутантный фермент GalT Y289L, который способен переносить азидогалактозу (GalNAz) на O -GlcNAc. [28] [39] Присутствие GalNAz (и, следовательно, также O -GlcNAc) можно обнаружить с помощью различных алкинсодержащих зондов с идентифицируемыми метками, такими как биотин, [39] молекулы красителей, [28] и ПЭГ. [29]

Биосенсор резонансной передачи энергии Фёрстера
[ редактировать ]Был разработан инженерный белковый биосенсор, который может обнаруживать изменения уровней O -GlcNAc с использованием резонансной передачи энергии Фёрстера . Этот сенсор состоит из четырех компонентов, соединенных вместе в следующем порядке: голубой флуоресцентный белок (CFP), связывающий домен O -GlcNAc (на основе GafD, лектина, чувствительного к терминальному β - O -GlcNAc), пептид CKII, который является известным Субстрат OGT и желтый флуоресцентный белок (YFP). При O -GlcNAcylation пептида CKII домен GafD связывает фрагмент O -GlcNAc, приводя домены CFP и YFP в непосредственную близость и генерируя сигнал FRET. Генерация этого сигнала обратима и может использоваться для мониторинга динамики O -GlcNAc в ответ на различные виды лечения. Этот сенсор может быть генетически закодирован и использован в клетках. [40] Добавление последовательности локализации позволяет нацелить этот сенсор O -GlcNAc на ядро, цитоплазму или плазматическую мембрану. [41]
Масс-спектрометрия
[ редактировать ]Биохимические подходы, такие как вестерн-блоттинг, могут предоставить подтверждающие доказательства того, что белок модифицируется O -GlcNAc; масс-спектрометрия (МС) способна предоставить окончательные доказательства присутствия O -GlcNAc. Гликопротеомные исследования с применением МС способствовали идентификации белков, модифицированных O -GlcNAc.
Поскольку O -GlcNAc является субстехиометрическим и подавление ионов происходит в присутствии немодифицированных пептидов, этап обогащения обычно выполняется перед масс-спектрометрическим анализом. Этого можно добиться с помощью лектинов, антител или химического мечения. Модификация O -GlcNAc является лабильной при использовании методов фрагментации, индуцированной столкновением, таких как диссоциация, индуцированная столкновением (CID) и столкновительная диссоциация с более высокими энергиями (HCD) , поэтому эти методы по отдельности трудно применимы для O картирования сайтов -GlcNAc. HCD генерирует фрагментированные ионы, характерные для N -ацетилгексозаминов, которые можно использовать для определения статуса O -GlcNAcylation. [42] Чтобы облегчить картирование сайтов с помощью HCD, можно использовать β-элиминирование с последующим Михаэлю добавлением по дитиотреитола (BEMAD) для преобразования лабильной модификации O -GlcNAc в более стабильную массовую метку. Для BEMAD-картирования O -GlcNAc образец необходимо обработать фосфатазой, в противном случае могут быть обнаружены другие посттрансляционные модификации серина/треонина, такие как фосфорилирование. [43] Диссоциация с переносом электронов (ETD) используется для картирования сайтов, поскольку ETD вызывает расщепление пептидного остова, оставляя при этом посттрансляционные модификации, такие как O -GlcNAc, нетронутыми. [44]
Традиционные протеомные исследования проводят тандемную МС на наиболее распространенных видах в полносканированных масс-спектрах, что запрещает полную характеристику видов с меньшей численностью. Одна из современных стратегий целевой протеомики использует изотопные метки, например, дибромид, для маркировки O -GlcNAцилированных белков. Этот метод позволяет алгоритмически обнаруживать виды с низкой численностью, которые затем секвенируются с помощью тандемного МС. [45] Направленная тандемная МС и целевое назначение гликопептидов позволяют идентифицировать последовательности O -GlcNAcylated пептидов. Один из примеров зонда состоит из аффинной метки биотина, расщепляемого кислотой силана, мотива изотопной перекодировки и алкина. [46] [47] [48] Однозначное картирование сайта возможно для пептидов только с одним остатком серина/треонина. [49]
Общая процедура метода изотопно-направленной гликопротеомики (IsoTaG) следующая:

- Метаболически метить O -GlcNAc для установки O -GlcNAz на белки.
- Используйте химию кликов, чтобы связать зонд IsoTaG с O -GlcNAz.
- Используйте стрептавидиновые шарики для обогащения мечеными белками.
- Обработайте шарики трипсином для высвобождения немодифицированных пептидов.
- Расщепляйте изотопно-перекодированные гликопептиды из шариков с помощью мягкой кислоты.
- Получите полный масс-спектр сканирования изотопно перекодированных гликопептидов.
- Применить алгоритм для обнаружения уникальной сигнатуры изотопа с зонда
- Выполните тандемную МС на изотопически перекодированных видах, чтобы получить аминокислотные последовательности гликопептидов.
- Поиск в базе данных белков для идентифицированных последовательностей
Были разработаны другие методологии количественного определения профиля O -GlcNAc с использованием дифференциальной изотопной маркировки. [50] Примеры зондов обычно состоят из аффинной метки биотина, расщепляемого линкера (расщепляемого кислотой или фото), тяжелой или легкой изотопной метки и алкина. [51] [52]
Стратегии манипулирования O -GlcNAc
[ редактировать ]Были разработаны различные химические и генетические стратегии для манипулирования O -GlcNAc как на уровне всего протеома , так и на конкретных белках.
Химические методы
[ редактировать ]Сообщалось о низкомолекулярных ингибиторах как для OGT, так и для [53] [54] и ОГА [55] [56] которые функционируют в клетках или in vivo . Ингибиторы OGT приводят к глобальному снижению O -GlcNAc, тогда как ингибиторы OGA приводят к глобальному увеличению O -GlcNAc; эти ингибиторы не способны модулировать O -GlcNAc на специфических белках.
Ингибирование пути биосинтеза гексозамина также способно снизить уровни O -GlcNAc. Например, аналоги глютамина азасерин и 6-диазо-5-оксо-L-норлейцин (ДОН) могут ингибировать GFAT , хотя эти молекулы также могут неспецифически влиять на другие пути. [57]
Синтез белка
[ редактировать ]Лигирование экспрессированных белков было использовано для получения O -GlcNAc-модифицированных белков сайт-специфическим способом. Существуют методы твердофазного пептидного синтеза с включением GlcNAc-модифицированного серина, треонина или цистеина. [58] [59]
Генетические методы
[ редактировать ]Сайт-направленный мутагенез
[ редактировать ]
Сайт-направленный мутагенез O -GlcNAc-модифицированных остатков серина или треонина в аланин можно использовать для оценки функции O -GlcNAc по конкретным остаткам. Поскольку боковая цепь аланина представляет собой метильную группу и, следовательно, не может действовать как сайт O -GlcNAc, эта мутация эффективно навсегда удаляет O -GlcNAc по определенному остатку. Хотя фосфорилирование серина/треонина можно смоделировать путем мутагенеза аспартата или глутамата , которые имеют отрицательно заряженные карбоксилатные боковые цепи, ни одна из 20 канонических аминокислот в достаточной степени не воспроизводит свойства O -GlcNAc. [60] Мутагенез триптофана использовался для имитации стерической массы O -GlcNAc, хотя триптофан гораздо более гидрофобен, чем O -GlcNAc. [61] [62] Мутагенез может также нарушать другие посттрансляционные модификации, например, если серин альтернативно фосфорилируется или O -GlcNAцилируется, мутагенез аланина навсегда устраняет возможности как фосфорилирования, так и O -GlcNAcylation.
S -GlcNAc
[ редактировать ]Масс-спектрометрия идентифицировала S -GlcNAc как посттрансляционную модификацию, обнаруженную на остатках цистеина. Эксперименты in vitro показали, что OGT может катализировать образование S -GlcNAc и что OGA не способен гидролизовать S -GlcNAc. [63] Хотя в предыдущем отчете предполагалось, что OGA способен гидролизовать тиогликозиды, это было продемонстрировано только на арилтиогликозиде пара -нитрофенол- S -GlcNAc; пара -нитротиофенол является более активированной уходящей группой, чем остаток цистеина. [64] Недавние исследования подтвердили использование S -GlcNAc в качестве ферментативно стабильной структурной модели O -GlcNAc, которую можно включать посредством твердофазного пептидного синтеза или сайт-направленного мутагенеза. [65] [60] [58] [66]
Разработанный ОГТ
[ редактировать ]Слитые конструкции нанотела и усеченного TPR OGT позволяют осуществлять индуцированное близостью белок-специфическое O -GlcNAcylation в клетках. Нанотело может быть направлено на белковые метки, например, GFP , которые слиты с целевым белком, или нанотело может быть направлено на эндогенные белки. Например, нанотело, распознающее С-концевую последовательность EPEA, может направлять ферментативную активность OGT на α-синуклеин . [67]
Функции O -GlcNAc
[ редактировать ]Апоптоз
[ редактировать ]Предполагается, что апоптоз, форма контролируемой гибели клеток, регулируется O -GlcNAc. при различных видах рака повышенные уровни O -GlcNAc подавляют апоптоз. Сообщалось, что [68] [69] Сообщалось, что каспаза-3 , каспаза-8 и каспаза-9 модифицируются O -GlcNAc. Каспаза-8 модифицируется вблизи сайтов расщепления/активации; Модификация O -GlcNAc может блокировать расщепление и активацию каспазы-8 за счет стерических препятствий. Фармакологическое снижение O -GlcNAc с помощью 5S - GlcNAc ускоряло активацию каспазы, тогда как фармакологическое повышение O -GlcNAc с помощью тиамет-G ингибировало активацию каспазы. [62]
Эпигенетика
[ редактировать ]Писатели и ластики
[ редактировать ]Белки, регулирующие генетику, часто подразделяют на «писателей», «читателей» и «ластиков», то есть ферменты, которые устанавливают эпигенетические модификации, белки, которые распознают эти модификации, и ферменты, которые удаляют эти модификации. [70] На сегодняшний день O -GlcNAc был идентифицирован на ферментах записи и стирания. O -GlcNAc обнаруживается во многих местах на EZH2 , каталитической метилтрансферазы субъединице PRC2 , и, как полагают, стабилизирует EZH2 до образования комплекса PRC2 и регулирует активность ди- и три-метилтрансферазы. [71] [72] Известно, что все три члена семейства диоксигеназ десять-одиннадцать транслокаций (TET) ( TET1 , TET2 и TET3 ) модифицируются O -GlcNAc. [73] Было высказано предположение, что O -GlcNAc вызывает ядерный экспорт TET3, снижая его ферментативную активность за счет его истощения из ядра. [74] O -GlcNAcylation HDAC1 связано с повышенным активирующим фосфорилированием HDAC1. [75]
Гистон O -GlcNAцилирование
[ редактировать ]Известно, что белки- гистоны , основной белковый компонент хроматина , модифицируются O -GlcNAc. [8] O -GlcNAc был идентифицирован на всех коровых гистонах ( H2A , [8] H2B , [8] Н3 , [76] и H4 [8] ). присутствие O -GlcNAc на гистонах влияет на транскрипцию генов, а также на другие метки гистонов, такие как ацетилирование. Было высказано предположение, что [8] и моноубиквитинирование . [77] TET2 взаимодействует с доменом TPR OGT и способствует привлечению OGT к гистонам. Сообщалось, что [78] Это взаимодействие связано с H2B S112 O -GlcNAc, который, в свою очередь, связан с моноубиквитинированием H2B K120. [77] фосфорилирование OGT T444 через AMPK Было обнаружено, что ингибирует ассоциацию OGT-хроматина и подавляет H2B S112 O -GlcNAc. [79]
Определение питательных веществ
[ редактировать ]Продукт пути биосинтеза гексозамина, UDP-GlcNAc, используется OGT для катализа добавления O -GlcNAc. Этот путь объединяет информацию о концентрациях различных метаболитов, включая аминокислоты, углеводы, жирные кислоты и нуклеотиды. Следовательно, уровни UDP-GlcNAc чувствительны к уровням клеточных метаболитов. Активность OGT частично регулируется концентрацией UDP-GlcNAc, создавая связь между состоянием клеточных питательных веществ и O -GlcNAc. [80]
Депривация глюкозы вызывает снижение уровней UDP-GlcNAc и начальное снижение O -GlcNAc, но, как ни странно, позже уровень O -GlcNAc значительно повышается. Было показано, что это более позднее увеличение зависит от активации AMPK и p38 MAPK , и этот эффект частично обусловлен увеличением уровней мРНК OGT и белка. [81] Было также высказано предположение, что этот эффект зависит от кальция и CaMKII . [82] Активированный р38 способен рекрутировать OGT к специфическим белкам-мишеням, включая нейрофиламент H ; Модификация O -GlcNAc нейрофиламента H повышает его растворимость. [81] Во время депривации глюкозы гликогенсинтаза модифицируется O -GlcNAc, что ингибирует ее активность. [83]
Окислительный стресс
[ редактировать ]Было обнаружено, что NRF2 , фактор транскрипции , связанный с клеточным ответом на окислительный стресс, косвенно регулируется O -GlcNAc. KEAP1 , адаптерный белок для куллина 3- зависимого комплекса E3 убиквитинлигазы , опосредует деградацию NRF2; окислительный стресс приводит к конформационным изменениям KEAP1, которые подавляют деградацию NRF2. Модификация O -GlcNAc KEAP1 по S104 необходима для эффективного убиквитинирования и последующей деградации NRF2, связывая O -GlcNAc с окислительным стрессом. Депривация глюкозы приводит к снижению O -GlcNAc и уменьшению деградации NRF2. Клетки, экспрессирующие мутант KEAP1 S104A, устойчивы к эрастину -индуцированному ферроптозу , что согласуется с более высокими уровнями NRF2 после удаления S104 O -GlcNAc. [84]
Повышенные уровни O -GlcNAc связаны с уменьшением синтеза в печени глутатиона , важного клеточного антиоксиданта . Передозировка ацетаминофена приводит к накоплению в печени сильно окисляющего метаболита NAPQI , который детоксицируется глутатионом. У мышей нокаут OGT оказывает защитное действие против повреждения печени, вызванного ацетаминофеном, тогда как ингибирование OGA тиаметом-G усугубляет повреждение печени, вызванное ацетаминофеном. [85]
Агрегация белков
[ редактировать ]O Было обнаружено, что -GlcNAc замедляет агрегацию белков, хотя общий характер этого явления неизвестен.
Твердофазный пептидный синтез использовали для получения полноразмерного α-синуклеина с модификацией O -GlcNAc в положении T72. Анализы агрегации тиофлавина Т и трансмиссионная электронная микроскопия показали, что этот модифицированный α-синуклеин с трудом образует агрегаты. [59]
Было показано, что обработка трансгенных мышей JNPL3 tau ингибитором OGA увеличивает -GlcNAцилирование белка, ассоциированного с микротрубочками тау -O . Иммуногистохимический анализ ствола мозга выявил снижение образования нейрофибриллярных клубков . было показано, что рекомбинантный O -GlcNAцилированный тау агрегирует медленнее, чем немодифицированный тау in vitro В анализе агрегации тиофлавина S . Аналогичные результаты были получены для рекомбинантно полученной O -GlcNAцилированной конструкции TAB1 по сравнению с ее немодифицированной формой. [86]
Фосфорилирование белков
[ редактировать ]Перекрестные помехи
[ редактировать ]Многие известные сайты фосфорилирования и сайты O -GlcNAcylation расположены рядом друг с другом или перекрываются. [49] Поскольку белок O -GlcNAcylation и фосфорилирование происходят как по остаткам серина, так и по треонину, эти посттрансляционные модификации могут регулировать друг друга. Например, в CKIIα , что S347 O -GlcNAc противодействует фосфорилированию T344. было показано [58] Реципрокное ингибирование, т.е. ингибирование фосфорилирования O -GlcNAcylation и O -GlcNAcylation фосфорилирования, наблюдалось на других белках, включая мышиный рецептор эстрогена β , [87] RNA Pol II , [88] да, [89] р53 , [90] КаМКИВ , [91] стр65 , [92] β-катенин , [93] и α-синуклеин. [59] Между этими двумя посттрансляционными модификациями также наблюдалась положительная кооперативность, т.е. фосфорилирование индуцирует O -GlcNAcylation или O -GlcNAcylation индуцирует фосфорилирование. Это было продемонстрировано на MeCP2. [29] и HDAC1. [75] В других белках, например, кофилине , фосфорилирование и O -GlcNAcylation, по-видимому, происходят независимо друг от друга. [94]
В некоторых случаях изучаются терапевтические стратегии для модуляции O -GlcNAcylation, чтобы оказать последующее влияние на фосфорилирование. Например, повышение тау -O -GlcNAcylation может принести терапевтический эффект за счет ингибирования патологического гиперфосфорилирования тау. [95]
Было обнаружено, что помимо фосфорилирования O -GlcNAc влияет на другие посттрансляционные модификации, такие как ацетилирование лизина. [92] и моноубиквитинирование. [77]
Киназы
[ редактировать ]Протеинкиназы — это ферменты, ответственные за фосфорилирование остатков серина и треонина. O -GlcNAc был идентифицирован более чем на 100 (~20% киназ человека ) киназах, и эта модификация часто связана с изменениями активности киназы или количества субстратов киназы. [96]
Первое сообщение о том, что киназа напрямую регулируется с помощью O -GlcNAc, было опубликовано в 2009 году. CaMKIV гликозилируется по нескольким сайтам, хотя было обнаружено, что основным сайтом является S189. Мутант S189A легче активировался фосфорилированием CaMKIV T200, что позволяет предположить, что O -GlcNAc в S189 ингибирует активность CaMKIV. Моделирование гомологии показало, что S189 O -GlcNAc может мешать связыванию АТФ . [91]
Известно, что AMPK и OGT модифицируют друг друга, т.е. AMPK фосфорилирует OGT, а OGT O -GlcNAcylates AMPK. Активация AMPK рибонуклеотидом AICA связана с ядерной локализацией OGT в дифференцированных миотрубках скелетных мышц мышей C2C12, что приводит к увеличению ядерного O -GlcNAc. Этот эффект не наблюдался в пролиферирующих клетках и недифференцированных миобластных клетках. [97] Было обнаружено, что фосфорилирование AMPK OGT T444 блокирует ассоциацию OGT с хроматином и снижает H2B S112 O -GlcNAc. [79] Было обнаружено, что сверхэкспрессия GFAT, фермента, который контролирует поток глюкозы в путь биосинтеза гексозамина, в жировой ткани мышей, приводит к активации AMPK и последующему ингибированию ACC и повышенному окислению жирных кислот . Обработка глюкозамином культивируемых адипоцитов 3T3L1 показала аналогичный эффект. [98] Точная взаимосвязь между O -GlcNAc и AMPK полностью не выяснена, поскольку в различных исследованиях сообщалось, что ингибирование OGA ингибирует активацию AMPK. [97] Ингибирование OGT также ингибирует активацию AMPK, [79] повышение регуляции O -GlcNAc с помощью обработки глюкозамином активирует AMPK, [98] и нокдаун OGT активирует AMPK; [99] эти результаты предполагают дополнительную непрямую связь между путями AMPK и O -GlcNAc или эффектами, специфичными для типа клеток.
Было показано, что распознавание субстрата CKIIα изменяется при S347 O -GlcNAcylation. [58]
фосфатазы
[ редактировать ]протеинфосфатазы 1 Было показано, что субъединицы PP1β и PP1γ образуют функциональные комплексы с OGT. Синтетический фосфопептид можно было дефосфорилировать и O -GlcNAцилировать с помощью иммунопреципитата OGT. Этот комплекс был назван «комплексом Инь-Ян», поскольку он заменяет фосфатную модификацию модификацией O -GlcNAc. [100]
MYPT1 представляет собой еще одну субъединицу протеинфосфатазы, которая образует комплексы с OGT и сама является O -GlcNAцилированной. MYPT1, по-видимому, играет роль в направлении OGT к определенным субстратам. [101]
Белково-белковые взаимодействия
[ редактировать ]O- GlcNAcylation белка может изменить его интерактом. Поскольку O -GlcNAc обладает высокой гидрофильностью, его присутствие может нарушать гидрофобные межбелковые взаимодействия. Например, O -GlcNAc нарушает взаимодействие Sp1 с TAF II 110, [102] и O -GlcNAc нарушает CREB взаимодействие с TAF II 130 и CRTC. [103] [104]
Некоторые исследования также выявили случаи, когда межбелковые взаимодействия индуцируются O -GlcNAc. Метаболическое мечение диазиринсодержащим O -GlcNDAz применялось для идентификации белок-белковых взаимодействий, индуцированных O -GlcNAc. [33] Используя гликопептид-приманку, основанный примерно на консенсусной последовательности для O -GlcNAc, α-енолаза , EBP1 и 14-3-3 , были идентифицированы как потенциальные ридеры O -GlcNAc. Рентгеновская кристаллография показала, что 14-3-3 распознает O -GlcNAc через амфипатическую бороздку, которая также связывает фосфорилированные лиганды. [105] Предполагается также, что Hsp70 действует как лектин, распознающий O -GlcNAc. [106] Было высказано предположение, что O -GlcNAc играет роль во взаимодействии α-катенина и β-катенина. [93]
Стабильность и деградация белка
[ редактировать ]Котрансляционный O -GlcNAc был идентифицирован на Sp1 и Nup62 . Эта модификация подавляет котрансляционное убиквитинирование и, таким образом, защищает образующиеся полипептиды от протеасомной деградации. Наблюдались аналогичные защитные эффекты O -GlcNAc на полноразмерный Sp1. Неизвестно, является ли эта закономерность универсальной или применима только к конкретным белкам. [14]
Фосфорилирование белка часто используется как признак последующей деградации. Белок -супрессор опухоли p53 нацелен на протеосомную деградацию посредством COP9 -опосредованного сигналосомой фосфорилирования T155. O -GlcNAcylation p53 S149 было связано со снижением фосфорилирования T155 и защитой p53 от деградации. [90] β-катенин O -GlcNAcylation конкурирует с фосфорилированием T41, которое сигнализирует о деградации β-катенина, стабилизируя белок. [93]
O -GlcNAcylation субъединицы АТФазы Rpt2 26S протеасомы Было показано, что ингибирует активность протеасомы. Тестирование различных пептидных последовательностей показало, что эта модификация замедляет протеасомную деградацию гидрофобных пептидов, при этом деградация гидрофильных пептидов, по-видимому, не затрагивается. [107] Было показано, что эта модификация подавляет другие пути, которые активируют протеасому, такие как Rpt6 фосфорилирование с помощью цАМФ-зависимой протеинкиназы . [108]
OGA-S локализуется в липидных каплях и, как предполагается, локально активирует протеасому, способствуя ремоделированию поверхностных белков липидных капель. [109]
Стрессовая реакция
[ редактировать ]Различные стимулы клеточного стресса были связаны с изменениями O -GlcNAc. Обработка перекисью водорода , хлоридом кобальта(II) , УФ-излучением , этанолом , хлоридом натрия , тепловым шоком и арсенитом натрия приводит к повышению уровня O -GlcNAc. Нокаут OGT повышает чувствительность клеток к тепловому стрессу. Повышенный уровень O -GlcNAc связан с экспрессией Hsp40 и Hsp70. [110]
Терапевтическая значимость
[ редактировать ]болезнь Альцгеймера
[ редактировать ]Многочисленные исследования выявили аберрантное фосфорилирование тау как признак болезни Альцгеймера. [111] O -GlcNAcylation бычьего тау впервые было описано в 1996 году. [112] Последующий отчет 2004 года продемонстрировал, что тау мозга человека также модифицируется O -GlcNAc. Было продемонстрировано, что O -GlcNAcylation тау регулирует фосфорилирование тау, при этом гиперфосфорилирование тау наблюдается в мозге мышей с отсутствием OGT, [113] что связано с образованием нейрофибриллярных клубков. Анализ образцов головного мозга показал, что белок O при болезни Альцгеймера нарушается -GlcNAcylation, а парный спиральный фрагмент-тау не распознается традиционными методами обнаружения O -GlcNAc, что позволяет предположить, что патологический тау нарушает O -GlcNAcylation по сравнению с тау, выделенным из контрольных образцов мозга. Повышение тау -O -GlcNAcylation было предложено в качестве терапевтической стратегии снижения фосфорилирования тау. [89]
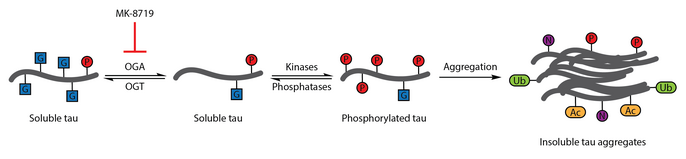
Для проверки этой терапевтической гипотезы был разработан селективный ингибитор OGA, проницаемый для гематоэнцефалического барьера , тиамет-G. Обработка тиаметом-G смогла увеличить тау- O -GlcNAcylation и подавить фосфорилирование тау в культуре клеток и in vivo у здоровых крыс Sprague-Dawley. [56] Последующее исследование показало, что лечение тиаметом-G также увеличивает тау- O -GlcNAcylation в модели трансгенных мышей JNPL3 tau. В этой модели лечение тиаметом-G существенно не влияло на фосфорилирование тау, хотя наблюдалось уменьшение количества нейрофибриллярных клубков и более медленная гибель мотонейронов. Кроме того, было отмечено, что O -GlcNAcylation тау замедляет агрегацию тау in vitro . [86]
Ингибирование OGA с помощью МК -8719 исследуется в клинических испытаниях в качестве потенциальной стратегии лечения болезни Альцгеймера и других таупатий , включая прогрессирующий супрануклеарный паралич . [95] [114] [115]
Рак
[ редактировать ]Нарушение регуляции O -GlcNAc связано с пролиферацией раковых клеток и ростом опухоли.
O -GlcNAцилирование гликолитического фермента PFK1 Было обнаружено, что по S529 ингибирует ферментативную активность PFK1, уменьшая гликолитический поток и перенаправляя глюкозу на пентозофосфатный путь . Структурное моделирование и биохимические эксперименты показали, что O -GlcNAc в положении S529 будет ингибировать аллостерическую активацию PFK1 фруктозо-2,6-бисфосфатом и олигомеризацию в активные формы. В мышиной модели у мышей, которым инъецировали клетки, экспрессирующие мутант PFK1 S529A, наблюдался более низкий рост опухоли, чем у мышей, которым инъецировали клетки, экспрессирующие PFK1 дикого типа. Кроме того, сверхэкспрессия OGT усиливала рост опухоли в последней системе, но не оказывала существенного влияния на систему с мутантным PFK1. Гипоксия индуцирует PFK1 S529 O -GlcNAc и увеличивает поток через пентозофосфатный путь для генерации большего количества НАДФН, который поддерживает уровни глутатиона и детоксицирует активные формы кислорода , обеспечивая преимущество роста раковым клеткам. Было обнаружено, что PFK1 гликозилирован в тканях опухолей молочной железы и легких человека. [116] Сообщалось также, что OGT положительно регулирует HIF-1α . HIF-1α обычно разлагается в нормоксических условиях под действием пролилгидроксилаз , которые используют α-кетоглутарат в качестве ко-субстрата. OGT подавляет уровни α-кетоглутарата, защищая HIF-1α от протеасомной деградации под действием pVHL и способствуя аэробному гликолизу . В отличие от предыдущего исследования PFK1, это исследование показало, что повышение уровня OGT или O -GlcNAc повышает регуляцию PFK1, хотя оба исследования сходятся в выводах о том, что уровни O -GlcNAc положительно связаны с потоком через пентозофосфатный путь. Это исследование также показало, что снижение O -GlcNAc избирательно убивает раковые клетки посредством апоптоза, вызванного стрессом ER . [68]
человека Клеточные линии аденокарциномы протоков поджелудочной железы (PDAC) имеют более высокие O уровни эпителия -GlcNAc, чем клетки протоков поджелудочной железы человека (HPDE). Выживание клеток PDAC в некоторой степени зависит от O -GlcNAc, поскольку нокдаун OGT избирательно ингибирует пролиферацию клеток PDAC (нокдаун OGT не оказывает существенного влияния на пролиферацию клеток HPDE), а ингибирование OGT с помощью 5 S -GlcNAc показало тот же результат. Гипер -O -GlcNAcylation в клетках PDAC оказался антиапоптотическим, ингибируя расщепление и активацию каспазы-3 и каспазы-9 . Было обнаружено, что многочисленные сайты субъединицы p65 NF-κB модифицируются O динамически -GlcNAc; O -GlcNAc в p65 T305 и S319, в свою очередь, положительно регулирует другие модификации, связанные с активацией NF-κB, такие как p300 -опосредованное ацетилирование K310 и IKK -опосредованное фосфорилирование S536. Эти результаты позволяют предположить, что NF-κB конститутивно активируется O -GlcNAc при раке поджелудочной железы. [69] [92]
Было обнаружено, что OGT-стабилизация EZH2 в различных клеточных линиях рака молочной железы ингибирует экспрессию генов-супрессоров опухоли. [71] В гепатоцеллюлярной карциномы моделях O -GlcNAc связан с активацией фосфорилирования HDAC1, который, в свою очередь, регулирует экспрессию регулятора клеточного цикла p21. Ваф1/Cip1 и регулятор подвижности клеток Е-кадгерин . [75]
Было обнаружено, что OGT стабилизирует SREBP-1 и активирует липогенез в клеточных линиях рака молочной железы. Эта стабилизация зависела от протеасомы и AMPK. Нокдаун OGT приводил к снижению ядерного SREBP-1, но протеасомное ингибирование с помощью MG132 блокировало этот эффект. Нокдаун OGT также усиливал взаимодействие между SREBP-1 и убиквитинлигазой E3 FBW7. AMPK активируется фосфорилированием T172 при нокдауне OGT, а AMPK фосфорилирует SREBP-1 S372, чтобы ингибировать его расщепление и созревание. Нокдаун OGT оказывал меньшее влияние на уровни SREBP-1 в клеточных линиях с нулевыми AMPK. На мышиной модели нокдаун OGT ингибировал рост опухоли, но сверхэкспрессия SREBP-1 частично спасала этот эффект. [99] Эти результаты контрастируют с результатами предыдущего исследования, в котором было обнаружено, что нокдаун/ингибирование OGT ингибирует фосфорилирование AMPK T172 и увеличивает липогенез. [79]
В клеточных линиях рака молочной железы и простаты высокие уровни OGT и O -GlcNAc были связаны как in vitro , так и in vivo с процессами, связанными с прогрессированием заболевания, например, ангиогенезом , инвазией и метастазированием . Было обнаружено, что нокдаун или ингибирование OGT подавляет транскрипционный фактор FoxM1 и активирует ингибитор клеточного цикла p27. Кип1 (который регулируется FoxM1-зависимой экспрессией компонента убиквитинлигазы Е3 Skp2 ), вызывая остановку клеточного цикла G1. Это, по-видимому, зависит от протеасомной деградации FoxM1, поскольку экспрессия мутанта FoxM1, лишенного дегрона, устраняет эффекты нокдауна OGT. Было обнаружено, что FoxM1 не подвергается прямой модификации с помощью O -GlcNAc, что указывает на то, что гипер- O -GlcNAcylation регуляторов FoxM1 нарушает деградацию FoxM1. Нацеливание на OGT также снизило уровни белков, регулируемых FoxM1, связанных с инвазией и метастазированием рака ( MMP-2 и MMP-9 ), а также ангиогенезом ( VEGF ). [117] [118] S108 O модификация кофилина Сообщалось также, что -GlcNAc важна для инвазии клеток рака молочной железы путем регуляции субклеточной локализации кофилина в инвадоподиях . [94]
Диабет
[ редактировать ]Повышенный уровень O -GlcNAc связан с диабетом.
поджелудочной железы β-клетки синтезируют и секретируют инсулин для регулирования уровня глюкозы в крови. Одно исследование показало, что ингибирование OGA стрептозотоцином с последующим лечением глюкозамином приводит к накоплению O -GlcNAc и апоптозу в β-клетках; [119] последующее исследование показало, что аналог стрептозотоцина на основе галактозы не способен ингибировать OGA, но все же приводит к апоптозу, что позволяет предположить, что апоптотические эффекты стрептозотоцина не связаны напрямую с ингибированием OGA. [120]
O Было высказано предположение, что -GlcNAc ослабляет передачу сигналов инсулина . 3T3-L1 В адипоцитах ингибирование OGA с помощью PUGNAc ингибировало опосредованное инсулином поглощение глюкозы. Лечение PUGNAc также ингибировало стимулируемое инсулином фосфорилирование Akt T308 и последующее фосфорилирование GSK3β S9. [121] В более позднем исследовании стимуляция инсулином клеток COS-7 привела к локализации OGT на плазматической мембране. Ингибирование PI3K вортманнином -трифосфата обращало этот эффект вспять, что указывает на зависимость от фосфатидилинозитол(3,4,5) . Увеличение уровней O -GlcNAc путем воздействия на клетки условий с высоким содержанием глюкозы или обработки PUGNAc ингибировало стимулируемое инсулином фосфорилирование Akt T308 и активность Akt. Фосфорилирование IRS1 на S307 и S632/S635, которое связано с ослаблением передачи сигналов инсулина, было усилено. Последующие эксперименты на мышах с аденовирусной доставкой OGT показали, что сверхэкспрессия OGT отрицательно регулирует передачу сигналов инсулина in vivo . Многие компоненты сигнального пути инсулина, включая β-катенин , [121] IR-β , IRS1, Akt, PDK1 Было обнаружено, что и субъединица p110α PI3K непосредственно модифицируются O -GlcNAc. [122] Сообщалось также, что передача сигналов инсулина приводит к фосфорилированию тирозина OGT и активации OGT, что приводит к увеличению уровней O -GlcNAc. [123]
Поскольку PUGNAc также ингибирует лизосомальные β-гексозаминидазы , был разработан селективный ингибитор OGA NButGT для дальнейшего исследования взаимосвязи между O -GlcNAc и передачей сигналов инсулина в адипоцитах 3T3-L1. Это исследование также показало, что PUGNAc приводил к нарушению передачи сигналов инсулина, а NButGT - нет, что было измерено по изменениям фосфорилирования Akt T308, что позволяет предположить, что эффекты, наблюдаемые с PUGNAc, могут быть обусловлены нецелевыми эффектами, помимо ингибирования OGA. [124]
болезнь Паркинсона
[ редактировать ]Болезнь Паркинсона связана с агрегацией α-синуклеина. [125] Поскольку было обнаружено, что модификация α-синуклеина O -GlcNAc ингибирует его агрегацию, повышение уровня α-синуклеина O -GlcNAc исследуется в качестве терапевтической стратегии лечения болезни Паркинсона. [59] [126]
Инфекционное заболевание
[ редактировать ]Бактериальный
[ редактировать ]Обработка макрофагов липополисахаридом (ЛПС) , основным компонентом внешней мембраны грамотрицательных бактерий , приводит к повышению уровня O -GlcNAc на клеточных и мышиных моделях. Во время инфекции цитозольный OGT де -S -нитрозилировался и активировался. Подавление O -GlcNAc с помощью DON ингибировало O -GlcNAcylation и ядерную транслокацию NF-κB, а также последующую индукцию индуцибельной синтазы оксида азота и продукцию IL-1β . Лечение ДОН также улучшало выживаемость клеток во время лечения ЛПС. [127]
Популярный
[ редактировать ]O -GlcNAc участвует в вирусом гриппа А (IAV) индуцированном цитокиновом шторме, . В частности, O -GlcNAcylation S430 на регуляторном факторе-5 интерферона (IRF5) было показано, что способствует его взаимодействию с фактором 6, ассоциированным с рецептором TNF (TRAF6), на клеточных и мышиных моделях. TRAF6 опосредует K63-связанное убиквитинирование IRF5, которое необходимо для активности IRF5 и последующей продукции цитокинов. Анализ клинических образцов показал, что уровень глюкозы в крови у пациентов, инфицированных IAV, был повышен по сравнению со здоровыми людьми. У пациентов, инфицированных IAV, уровень глюкозы в крови положительно коррелировал с уровнями IL-6 и IL-8 . O -GlcNAcylation IRF5 также был относительно выше в мононуклеарных клетках периферической крови пациентов, инфицированных IAV. [128]
Другие приложения
[ редактировать ]Пептидные терапевтические средства, такие как привлекательны своей высокой специфичностью и эффективностью, но они часто имеют плохие фармакокинетические профили из-за их разложения под действием сывороточных протеаз . [129] Хотя O -GlcNAc обычно связан с внутриклеточными белками, было обнаружено, что сконструированные пептидные терапевтические средства, модифицированные O -GlcNAc, обладают повышенной стабильностью в сыворотке на мышиной модели и имеют аналогичную структуру и активность по сравнению с соответствующими немодифицированными пептидами. Этот метод был применен для создания пептидов GLP-1 и PTH. [130]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Зейдан, Кира; Харт, Джеральд В. (1 января 2010 г.). «Пересечение между O-GlcNAcylation и фосфорилированием: значение для нескольких сигнальных путей» . Журнал клеточной науки . 123 (1): 13–22. дои : 10.1242/jcs.053678 . ISSN 0021-9533 . ПМЦ 2794709 . ПМИД 20016062 .
- ^ Диас, Вагнер Б.; Чунг, Вин Д.; Харт, Джеральд В. (1 июня 2012 г.). «O-GlcNAcylation киназ» . Связь с биохимическими и биофизическими исследованиями . 422 (2): 224–228. дои : 10.1016/j.bbrc.2012.04.124 . ISSN 0006-291X . ПМЦ 3387735 . ПМИД 22564745 .
- ^ Халтивангер, РС; Холт, Джорджия; Харт, GW (15 февраля 1990 г.). «Ферментативное добавление O-GlcNAc к ядерным и цитоплазматическим белкам. Идентификация уридиндифосфо-N-ацетилглюкозамина:пептида бета-N-ацетилглюкозаминилтрансферазы» . Журнал биологической химии . 265 (5): 2563–8. дои : 10.1016/S0021-9258(19)39838-2 . ПМИД 2137449 .
- ^ Вульф-Фуэнтес Э., Берендт Р.Р., Массман Л., Даннер Л., Малард Ф., Вора Дж., Кахсай Р., Оливье-Ван Стихелен С. (январь 2021 г.). -GlcNAcome человека «База данных O и метаанализ» . Научные данные . 8 (1): 25. Бибкод : 2021NatSD...8...25W . дои : 10.1038/s41597-021-00810-4 . ПМЦ 7820439 . ПМИД 33479245 .
- ^ Ма, Цзюньфэн; Харт, Джеральд В. (5 марта 2014 г.). «Профилирование O-GlcNAc: от белков к протеомам» . Клиническая протеомика . 11 (1): 8. дои : 10.1186/1559-0275-11-8 . ISSN 1542-6416 . ПМК 4015695 . ПМИД 24593906 .
- ^ Кинг, Дастин Т.; Серрано-Негрон, Хесус Э.; Чжу, Яньпин; Мур, Кристофер Л.; Плечи, Мэтью Д.; Фостер, Леонард Дж.; Вокадло, Дэвид Дж. (9 марта 2022 г.). «Термическое профилирование протеома выявляет O-GlcNAc-зависимый мельтом» . Журнал Американского химического общества . 144 (9): 3833–3842. дои : 10.1021/jacs.1c10621 . ISSN 1520-5126 . ПМЦ 8969899 . ПМИД 35230102 .
- ^ Келли, WG; Дамус, Мэн; Харт, GW (15 мая 1993 г.). «РНК-полимераза II представляет собой гликопротеин. Модификация COOH-концевого домена с помощью O-GlcNAc» . Журнал биологической химии . 268 (14): 10416–24. дои : 10.1016/S0021-9258(18)82216-5 . ПМИД 8486697 .
- ^ Перейти обратно: а б с д и ж Сакабе, К; Ван, З; Харт, GW (16 ноября 2010 г.). «Бета-N-ацетилглюкозамин (O-GlcNAc) является частью гистонового кода» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (46): 19915–20. Бибкод : 2010PNAS..10719915S . дои : 10.1073/pnas.1009023107 . ПМЦ 2993388 . ПМИД 21045127 .
- ^ Левин, З; Уокер, С. (2 июня 2016 г.). «Биохимия трансферазы O-GlcNAc: какие функции делают ее незаменимой в клетках млекопитающих?» . Ежегодный обзор биохимии . 85 : 631–57. doi : 10.1146/annurev-biochem-060713-035344 . ПМИД 27294441 .
- ^ Онг, Цюньсян; Хан, Вэйпин; Ян, Сяоюн (16 октября 2018 г.). «O-GlcNAc как интегратор сигнальных путей» . Границы эндокринологии . 9 : 599. дои : 10.3389/fendo.2018.00599 . ISSN 1664-2392 . ПМК 6234912 . PMID 30464755 .
- ^ Харт, Джеральд В.; Слоусон, Чад; Рамирес-Корреа, Хенаро; Лагерлёф, Олоф (7 июня 2011 г.). «Перекрестный разговор между O-GlcNAcylation и фосфорилированием: роль в передаче сигналов, транскрипции и хронических заболеваниях» . Ежегодный обзор биохимии . 80 : 825–858. doi : 10.1146/annurev-biochem-060608-102511 . ISSN 0066-4154 . ПМК 3294376 . ПМИД 21391816 .
- ^ Торрес, ЧР; Харт, GW (10 марта 1984 г.). «Топография и полипептидное распределение терминальных остатков N-ацетилглюкозамина на поверхности интактных лимфоцитов. Доказательства существования O-связанного GlcNAc» . Журнал биологической химии . 259 (5): 3308–17. дои : 10.1016/S0021-9258(17)43295-9 . ПМИД 6421821 .
- ^ Перейти обратно: а б Шен, Дэвид Л.; Глостер, Трейси М.; Юзва, Скотт А.; Вокадло, Дэвид Дж. (4 мая 2012 г.). «Понимание процесса обработки и динамики O-связанного N-ацетилглюкозамина (O-GlcNAc) посредством кинетического анализа трансферазы O-GlcNAc и активности O-GlcNAcase на белковых субстратах» . Журнал биологической химии . 287 (19): 15395–15408. дои : 10.1074/jbc.M111.310664 . ISSN 0021-9258 . ПМК 3346082 . ПМИД 22311971 .
- ^ Перейти обратно: а б Чжу, Ю; Лю, ТВ; Чечони, С; Эскандари, Р; Зандберг, ВФ; Вокадло, диджей (май 2015 г.). «O-GlcNAc происходит котрансляционно, стабилизируя возникающие полипептидные цепи». Химическая биология природы . 11 (5): 319–25. дои : 10.1038/nchembio.1774 . ПМИД 25774941 .
- ^ Перейти обратно: а б с Лазарус, МБ; Нам, Ю; Цзян, Дж; Слиз, П; Уокер, С. (27 января 2011 г.). «Структура трансферазы O-GlcNAc человека и ее комплекса с пептидным субстратом» . Природа . 469 (7331): 564–7. Бибкод : 2011Natur.469..564L . дои : 10.1038/nature09638 . ПМК 3064491 . ПМИД 21240259 .
- ^ Маколи, Миссисипи; Уитворт, GE; Дебовски, AW; Чин, Д; Вокадло, диджей (8 июля 2005 г.). «O-GlcNAcase использует катализ с участием субстрата: кинетический анализ и разработка высокоселективных ингибиторов, основанных на механизме действия» . Журнал биологической химии . 280 (27): 25313–22. дои : 10.1074/jbc.M413819200 . ПМИД 15795231 .
- ^ Рот, Кристиан; Чан, Шерри; Оффен, Венди А; Хемсворт, Глин Р.; Виллемс, Лианна I; Кинг, Дастин Т; Варгезе, Вимал; Бриттон, Роберт; Вокадло, Дэвид Дж; Дэвис, Гидеон Дж. (июнь 2017 г.). «Структурное и функциональное понимание человеческой O-GlcNAcase» . Химическая биология природы . 13 (6): 610–612. дои : 10.1038/nchembio.2358 . ISSN 1552-4450 . ПМЦ 5438047 . ПМИД 28346405 .
- ^ Элсен, Нидерланды; Патель, С.Б.; Форд, RE; Холл, ДЛ; Хесс, Ф; Кандула, Х; Корниенко М; Рид, Дж; Селник, Х. (июнь 2017 г.). «Понимание активности и ингибирования кристаллической структуры человеческой O-GlcNAcase». Химическая биология природы . 13 (6): 613–615. дои : 10.1038/nchembio.2357 . ПМИД 28346407 .
- ^ Столяр, CM; Левин, З.Г.; Аонбангхен, К; Ву, СМ; Уокер, С. (21 августа 2019 г.). «Остатки аспартата вдали от активного сайта управляют выбором субстрата трансферазы O-GlcNAc» . Журнал Американского химического общества . 141 (33): 12974–12978. дои : 10.1021/jacs.9b06061 . ПМК 6849375 . ПМИД 31373491 .
- ^ Левин, З.Г.; Фан, С; Мелишер, М.С.; Орман, М; Бенджамин, Т; Уокер, С. (14 марта 2018 г.). «O-GlcNAc Трансфераза распознает белковые субстраты с использованием аспарагиновой лестницы в суперспирали тетратрикопептидного повтора (TPR)» . Журнал Американского химического общества . 140 (10): 3510–3513. дои : 10.1021/jacs.7b13546 . ПМЦ 5937710 . ПМИД 29485866 .
- ^ Перейти обратно: а б С, Патхак; Дж, Алонсо; М, Шимпл; К, Рафи; Де, Блэр; Вс, Бородкин; О, Альбарбарави; Dmf, ван Алтен (сентябрь 2015 г.). «Активный сайт трансферазы O-GlcNAc накладывает ограничения на последовательность субстрата» . Структурная и молекулярная биология природы . 22 (9): 744–750. дои : 10.1038/nsmb.3063 . ПМЦ 4979681 . ПМИД 26237509 .
- ^ Чунг, штат Вирджиния; Сакабе, К; Хаусли, член парламента; Диас, ВБ; Харт, GW (05 декабря 2008 г.). «Специфичность субстрата O-связанной бета-N-ацетилглюкозаминилтрансферазы регулируется путем нацеливания на миозинфосфатазу и другие взаимодействующие белки» . Журнал биологической химии . 283 (49): 33935–41. дои : 10.1074/jbc.M806199200 . ПМК 2590692 . ПМИД 18840611 .
- ^ Захара, Наташа Е.; Восселлер, Кейт; Харт, Джеральд В. (ноябрь 2011 г.). «Обнаружение и анализ белков, модифицированных О-связанным N-ацетилглюкозамином» . Современные протоколы в науке о белках . ГЛАВА: 12.8.1–12.8.33. дои : 10.1002/0471140864.ps1208s66 . ISSN 1934-3655 . ПМЦ 3349994 . ПМИД 22045558 .
- ^ Сноу, СМ; Старший, А.; Джераче, Л. (1 мая 1987 г.). «Моноклональные антитела идентифицируют группу гликопротеинов комплекса ядерных пор» . Журнал клеточной биологии . 104 (5): 1143–1156. дои : 10.1083/jcb.104.5.1143 . ISSN 0021-9525 . ПМК 2114474 . ПМИД 2437126 .
- ^ Комер, Финляндия; Восселер, К; Уэллс, Л; Аккавитти, Массачусетс; Харт, GW (15 июня 2001 г.). «Характеристика мышиного моноклонального антитела, специфичного к O-связанному N-ацетилглюкозамину». Аналитическая биохимия . 293 (2): 169–77. дои : 10.1006/abio.2001.5132 . ПМИД 11399029 .
- ^ Тео, CF; Ингале, С; Вольферт, Массачусетс; Эльсайед, Джорджия; Нот, Л.Г.; Чатем, Джей Си; Уэллс, Л; Бунс, GJ (май 2010 г.). «Гликопептид-специфические моноклональные антитела открывают новую роль O-GlcNAc» . Химическая биология природы . 6 (5): 338–43. дои : 10.1038/nchembio.338 . ПМЦ 2857662 . ПМИД 20305658 .
- ^ Перейти обратно: а б диджей, Вокадло; ХК, повесить; Эй, Ким; Да, Ганновер; Кр, Бертоцци (05 августа 2003 г.). «Химический подход к идентификации O-GlcNAc-модифицированных белков в клетках» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (16): 9116–21. Бибкод : 2003PNAS..100.9116V . дои : 10.1073/pnas.1632821100 . ПМК 171382 . ПМИД 12874386 .
- ^ Перейти обратно: а б с Кларк, премьер-министр; Двек, Дж. Ф.; Мейсон, Делавэр; Харт, ЧР; Бак, С.Б.; Петерс, ЕС; Агнью, Би Джей; Се-Уилсон, LC (3 сентября 2008 г.). «Прямое обнаружение флуоресценции в геле и клеточная визуализация O-GlcNAc-модифицированных белков» . Журнал Американского химического общества . 130 (35): 11576–7. дои : 10.1021/ja8030467 . ПМЦ 2649877 . ПМИД 18683930 .
- ^ Перейти обратно: а б с Рексач, Дж. Э.; Роджерс, CJ; Ю, Ш; Тао, Дж; Солнце, Ю; Се-Уилсон, LC (сентябрь 2010 г.). «Количественная оценка стехиометрии и динамики O-гликозилирования с использованием разрешимых массовых меток» . Химическая биология природы . 6 (9): 645–51. дои : 10.1038/nchembio.412 . ПМЦ 2924450 . ПМИД 20657584 .
- ^ Перейти обратно: а б Уолтер, Луизиана; Батт, Арканзас; Дарабедян, Н.; Заро, BW; Пратт, MR (17 сентября 2018 г.). «Метаболические химические репортеры гликозилирования, содержащие азид и алкин, демонстрируют структурно-зависимое ингибирование пути биосинтеза гексозамина по обратной связи» . ХимБиоХим . 19 (18): 1918–1921. дои : 10.1002/cbic.201800280 . ПМК 6261355 . ПМИД 29979493 .
- ^ Бойс, М; Каррико, Исландия; Гангули, А.С.; Ю, Ш; Хангауэр, MJ; Хаббард, Южная Каролина; Колер, Джей-Джей; Бертоцци, ЧР (22 февраля 2011 г.). «Метаболические перекрестные помехи позволяют маркировать O-связанные белки, модифицированные бета-N-ацетилглюкозамином, через путь спасения N-ацетилгалактозамина» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (8): 3141–6. Бибкод : 2011PNAS..108.3141B . дои : 10.1073/pnas.1010045108 . ПМК 3044403 . ПМИД 21300897 .
- ^ Тан, Хай; Эскандари, Р; Шен, Д; Чжу, Ю; Лю, ТВ; Виллемс, Л.И.; Алтин, МГ; Мэдден, З; Вокадло, диджей (14 ноября 2018 г.). «Прямое одноэтапное флуоресцентное мечение O-GlcNAc-модифицированных белков в живых клетках с использованием метаболических промежуточных продуктов» . Журнал Американского химического общества . 140 (45): 15300–15308. дои : 10.1021/jacs.8b08260 . ПМИД 30296064 . S2CID 207194442 .
- ^ Перейти обратно: а б Ю, Ш; Бойс, М; Вандс, AM; Бонд, MR; Бертоцци, ЧР; Колер, Джей-Джей (27 марта 2012 г.). «Метаболическая маркировка обеспечивает селективное фотосшивание O-GlcNAc-модифицированных белков с их партнерами по связыванию» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (13): 4834–9. Бибкод : 2012PNAS..109.4834Y . дои : 10.1073/pnas.1114356109 . ПМК 3323966 . ПМИД 22411826 .
- ^ Родригес, AC; Колер, Джей-Джей (1 августа 2014 г.). «Распознавание модифицированного диазирином O-GlcNAc человеческой O-GlcNAcase» . МедХимКомм . 5 (8): 1227–1234. дои : 10.1039/C4MD00164H . ПМК 4109824 . ПМИД 25068034 .
- ^ Заро, BW; Ян, ГГ; Ханг, ХК; Пратт, MR (17 мая 2011 г.). «Химические репортеры для флуоресцентного обнаружения и идентификации O-GlcNAc-модифицированных белков обнаруживают гликозилирование убиквитинлигазы NEDD4-1» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (20): 8146–51. Бибкод : 2011PNAS..108.8146Z . дои : 10.1073/pnas.1102458108 . ПМК 3100932 . ПМИД 21540332 .
- ^ Заро, Балин В.; Чу, Келли Н.; Пратт, Мэтью Р. (19 сентября 2014 г.). «Химический репортер для визуализации метаболических взаимодействий между метаболизмом углеводов и модификацией белка» . АКС Химическая биология . 9 (9): 1991–1996. дои : 10.1021/cb5005564 . ISSN 1554-8929 . ПМЦ 4168799 . ПМИД 25062036 .
- ^ Цинь, Кэ; Пэн, Линхан; Чжу, Юньтао; Ду, Жунбин; Чэн, Лю, Чжоу; Вэнь, Чу; Чен, Син (12 февраля 2018 г.). «Искусственное S-гликозилирование цистеина, индуцированное пер-O-ацетилированными неприродными моносахаридами во время метаболического маркирования гликанов» . Angewandte Chemie International Edition : 57 (7) 1817–1820. : 10.1002 /anie.201711710 . PMID 29237092 doi
- ^ Цинь, Кэ; Чжан, Хао; Чжао, Чжэньци; Чен, Син (20 мая 2020 г.). «S-гликомодификация белка посредством механизма исключения-добавления» . Журнал Американского химического общества . 142 (20): 9382–9388. дои : 10.1021/jacs.0c02110 . ISSN 0002-7863 . ПМИД 32339456 .
- ^ Перейти обратно: а б «Система ферментативной маркировки Click-IT™ O-GlcNAc» . www.thermofisher.com . Проверено 30 мая 2020 г.
- ^ Каррильо, LD; Кришнамурти, Л; Махал, ЛК (22 ноября 2006 г.). «Клеточный датчик на основе FRET для бета-O-GlcNAc, динамической модификации углеводов, участвующей в передаче сигналов». Журнал Американского химического общества . 128 (46): 14768–9. дои : 10.1021/ja065835+ . ПМИД 17105262 .
- ^ Каррильо, Луз Д.; Фромминг, Джошуа А.; Махал, Лара К. (25 февраля 2011 г.). «Датчики O-GlcNAc, предназначенные для применения в естественных условиях, выявляют дискретную динамику, специфичную для отдельных отсеков, во время передачи сигнала» . Журнал биологической химии . 286 (8): 6650–6658. дои : 10.1074/jbc.M110.191627 . ISSN 0021-9258 . ПМК 3057821 . ПМИД 21138847 .
- ^ Ма, Цзюньфэн; Харт, Джеральд В. (2 февраля 2017 г.). «Анализ белка O-GlcNAcylation методом масс-спектрометрии» . Современные протоколы в науке о белках . 87 : 24.10.1–24.10.16. дои : 10.1002/cpps.24 . ISSN 1934-3655 . ПМК 5300742 . ПМИД 28150883 .
- ^ Уэллс, Л; Восселер, К; Коул, Р.Н.; Кроншоу, Дж. М.; Матунис, МЮ; Харт, GW (октябрь 2002 г.). «Картирование сайтов модификации O-GlcNAc с использованием аффинных меток для посттрансляционных модификаций серина и треонина» . Молекулярная и клеточная протеомика . 1 (10): 791–804. дои : 10.1074/mcp.m200048-mcp200 . ПМИД 12438562 .
- ^ Чжао, Пэн; Винер, Роза; Тео, Чин Фен; Бунс, Герт-Ян; Хорн, Дэвид; Уэллс, Лэнс (2 сентября 2011 г.). «Сочетание высокоэнергетической диссоциации C-ловушки и диссоциации с переносом электрона для назначения сайта модификации белка O-GlcNAc» . Журнал исследований протеома . 10 (9): 4088–4104. дои : 10.1021/pr2002726 . ISSN 1535-3893 . ПМК 3172619 . ПМИД 21740066 .
- ^ Паланиаппан, Кришнан К.; Питчер, Остин А.; Смарт, Брайан П.; Спичарич, Дэвид Р.; Явароне, Энтони Т.; Бертоцци, Кэролин Р. (19 августа 2011 г.). «Перенос изотопной сигнатуры и прогнозирование массовой структуры (IsoStamp): метод, позволяющий использовать химически направленную протеомику» . АКС Химическая биология . 6 (8): 829–836. дои : 10.1021/cb100338x . ISSN 1554-8929 . ПМК 3220624 . ПМИД 21604797 .
- ^ Ву, СМ; Явароне, AT; Спичарич, доктор медицинских наук; Паланиаппан, КК; Бертоцци, ЧР (июнь 2015 г.). «Изотопно-ориентированная гликопротеомика (IsoTaG): независимая от массы платформа для открытия и анализа интактных N- и O-гликопептидов» . Природные методы . 12 (6): 561–7. дои : 10.1038/nmeth.3366 . ПМЦ 4599779 . ПМИД 25894945 .
- ^ Ву, Кристина М.; Феликс, Алехандра; Берд, Уильям Э.; Зюгель, Девон К.; Исихара, Маюми; Азади, Парастоо; Явароне, Энтони Т.; Питтери, Шэрон Дж.; Бертоцци, Кэролин Р. (07 апреля 2017 г.). «Разработка IsoTaG, метода химической гликопротеомики для определения профиля интактных N- и O-гликопептидов из цельноклеточных протеомов» . Журнал исследований протеома . 16 (4): 1706–1718. doi : 10.1021/acs.jproteome.6b01053 . ISSN 1535-3893 . ПМЦ 5507588 . ПМИД 28244757 .
- ^ Ву, Кристина М.; Феликс, Алехандра; Чжан, Личао; Элиас, Джошуа Э.; Бертоцци, Кэролин Р. (январь 2017 г.). «Изотопно-ориентированная гликопротеомика (IsoTaG) анализ сиалированных N- и O-гликопептидов на трибриде слияния с орбитальной ловушкой с использованием азидо- и алкиниловых сахаров» . Аналитическая и биоаналитическая химия . 409 (2): 579–588. дои : 10.1007/s00216-016-9934-9 . ISSN 1618-2642 . ПМЦ 5342897 . ПМИД 27695962 .
- ^ Перейти обратно: а б Ву, СМ; Лунд, П.Дж.; Хуанг, AC; Дэвис, ММ; Бертоцци, ЧР; Питтери, SJ (апрель 2018 г.). «Картирование и количественная оценка более 2000 O-связанных гликопептидов в активированных Т-клетках человека с помощью гликопротеомики, нацеленной на изотопы (изотаг)» . Молекулярная и клеточная протеомика . 17 (4): 764–775. дои : 10.1074/mcp.RA117.000261 . ПМК 5880114 . ПМИД 29351928 .
- ^ Хидекель, Н; Фикарро, СБ; Кларк, премьер-министр; Брайан, MC; Суони, Д.Л.; Рексач, Дж. Э.; Солнце, Ю; Кун, Джей-Джей; Петерс, ЕС; Се-Уилсон, LC (июнь 2007 г.). «Исследование динамики гликозилирования O-GlcNAc в мозге с использованием количественной протеомики» (PDF) . Химическая биология природы . 3 (6): 339–48. дои : 10.1038/nchembio881 . ПМИД 17496889 .
- ^ Цинь, К; Чжу, Ю; Цинь, Вт; Гао, Дж; Шао, X; Ван, ЮЛ; Чжоу, Вт; Ван, К; Чен, X (17 августа 2018 г.). «Количественное профилирование сайтов O-GlcNAcylation белка с помощью расщепляемого линкера, меченного изотопом». АКС Химическая биология . 13 (8): 1983–1989. doi : 10.1021/acschembio.8b00414 . ПМИД 30059200 . S2CID 206528204 .
- ^ Ли, Дж; Ли, З; Дуань, X; Цинь, К; Данг, Л; Солнце, С; Кай, Л; Се-Уилсон, LC; Ву, Л; Йи, В (18 января 2019 г.). «Фоторасщепляемый зонд с изотопным кодированием для количественного определения профиля O-GlcNAcylation белка» (PDF) . АКС Химическая биология . 14 (1): 4–10. дои : 10.1021/acschembio.8b01052 . ПМИД 30620550 . S2CID 58620368 .
- ^ Лю, Тай-Вэй; Зандберг, Уэсли Ф.; Глостер, Трейси М.; Дэн, Лехуа; Мюррей, Келси Д.; Шань, Сяоян; Вокадло, Дэвид Дж. (25 июня 2018 г.). «Метаболические ингибиторы трансферазы O-GlcNAc, действующие in vivo, приводят к снижению уровней O-GlcNAc при опосредованном лептином распознавании питательных веществ» . Angewandte Chemie, международное издание . 57 (26): 7644–7648. дои : 10.1002/anie.201803254 . ISSN 1521-3773 . ПМК 6055616 . ПМИД 29756380 .
- ^ Мартин, Сара Э.С.; Тан, Чжи-Вэй; Итконен, Харри М.; Дюво, Дэмиен Ю.; Пауло, Жоао А.; Джанецко, Джон; Бутц, Пол Л.; Тёрк, Лиза; Мосс, Фредерик А.; Томас, Крейг Дж.; Гиги, Стивен П. (24 октября 2018 г.). «Структурная эволюция низконаномолярных ингибиторов трансферазы O-GlcNAc» . Журнал Американского химического общества . 140 (42): 13542–13545. дои : 10.1021/jacs.8b07328 . ISSN 1520-5126 . ПМК 6261342 . ПМИД 30285435 .
- ^ Дорфмюллер, Хельге К.; Бородкин Владимир С.; Шимпл, Марианна; Шеперд, Шэрон М.; Шпиро, Наталья Александровна; ван Алтен, Даан МФ (27 декабря 2006 г.). «GlcNAcstatin: пикомолярный селективный ингибитор O-GlcNAcase, который модулирует внутриклеточные уровни O-glcNAcylation» . Журнал Американского химического общества . 128 (51): 16484–16485. дои : 10.1021/ja066743n . ISSN 0002-7863 . ПМЦ 7116141 . ПМИД 17177381 .
- ^ Перейти обратно: а б Юзва, ЮАР; Маколи, Миссисипи; Хейнонен, Дж. Э.; Шан, X; Деннис, Р.Дж.; Привет; Уитворт, GE; Стаббс, штат Калифорния; МакИчерн, Э.Дж. (август 2008 г.). «Мощный ингибитор O-GlcNAcase, основанный на механизме, который блокирует фосфорилирование тау in vivo». Химическая биология природы . 4 (8): 483–90. дои : 10.1038/nchembio.96 . ПМИД 18587388 .
- ^ Акелла, Неха М.; Чираку, Лорела; Реджинато, Маурисио Х. (4 июля 2019 г.). «Разжигая огонь: новая роль пути биосинтеза гексозамина при раке» . БМК Биология . 17 (1): 52. дои : 10.1186/s12915-019-0671-3 . ISSN 1741-7007 . ПМК 6610925 . ПМИД 31272438 .
- ^ Перейти обратно: а б с д Таррант, МК; Ро, ХС; Се, Z; Цзян, ЮЛ; Гросс, К; Калхейн, Джей Си; Ян, Г; Цянь, Дж; Итикава, Ю (22 января 2012 г.). «Регуляция CK2 путем фосфорилирования и O-GlcNAcylation, выявленная с помощью полусинтеза» . Химическая биология природы . 8 (3): 262–9. дои : 10.1038/nchembio.771 . ПМЦ 3288285 . ПМИД 22267120 .
- ^ Перейти обратно: а б с д Маротта, Северная Каролина; Лин, Ю.Х.; Льюис, Ю.Е.; Амбросо, MR; Заро, BW; Рот, Монтана; Арнольд, Д.Б.; Ланген, Р; Пратт, MR (ноябрь 2015 г.). «Модификация O-GlcNAc блокирует агрегацию и токсичность белка α-синуклеина, связанного с болезнью Паркинсона» . Природная химия . 7 (11): 913–20. Бибкод : 2015НатЧ...7..913М . дои : 10.1038/nchem.2361 . ПМК 4618406 . ПМИД 26492012 .
- ^ Перейти обратно: а б Горелик, А; Бартуаль, СГ; Бородкин В.С.; Варгезе, Дж; Ференбах, АТ; ван Алтен, DMF (ноябрь 2019 г.). «Генетическая перекодировка для анализа роли сайт-специфического белка O-GlcNAcylation» . Структурная и молекулярная биология природы . 26 (11): 1071–1077. дои : 10.1038/s41594-019-0325-8 . ПМЦ 6858883 . ПМИД 31695185 .
- ^ Льюис, Ю.Е.; Галесич, А; Левин, премьер-министр; Де Леон, Калифорния; Ламири, Н; Бреннан, КК; Пратт, MR (21 апреля 2017 г.). «O-GlcNAcylation α-синуклеина по серину 87 уменьшает агрегацию, не влияя на мембранное связывание» . АКС Химическая биология . 12 (4): 1020–1027. дои : 10.1021/acschembio.7b00113 . ПМК 5607117 . ПМИД 28195695 .
- ^ Перейти обратно: а б Чу, Келли Н.; Батт, Анна Р.; Заро, Балин В.; Дарабедян, Нарек; Маротта, Николас П.; Бреннан, Кэролайн К.; Амирхекмат, Арья; Пратт, Мэтью Р. (14 июня 2017 г.). «Новый химический репортер 6-алкинил-6-дезокси-GlcNAc раскрывает модификацию O-GlcNAc апоптотических каспаз, которая может блокировать расщепление / активацию каспазы-8» . Журнал Американского химического общества . 139 (23): 7872–7885. дои : 10.1021/jacs.7b02213 . ISSN 0002-7863 . ПМК 6225779 . ПМИД 28528544 .
- ^ Мейнард, Дж. К.; Берлингейм, Алабама; Медзиградский, К.Ф. (ноябрь 2016 г.). «Цистеин S-связанный N-ацетилглюкозамин (S-GlcNAcylation), новая посттрансляционная модификация у млекопитающих» . Молекулярная и клеточная протеомика . 15 (11): 3405–3411. дои : 10.1074/mcp.M116.061549 . ПМК 5098038 . ПМИД 27558639 .
- ^ Маколи, Миссисипи; Стаббс, штат Калифорния; Вокадло, диджей (14 декабря 2005 г.). «O-GlcNAcase катализирует расщепление тиогликозидов без общего кислотного катализа» . Журнал Американского химического общества . 127 (49): 17202–3. дои : 10.1021/ja0567687 . ПМИД 16332065 .
- ^ Мехта, AY; Вирайя, РХ; Дутта, С; Гот, СК; Ханес, М.С.; Гао, К; Ставенхаген, К; Кардиш, Р; Мацумото, Ю; Хаймбург-Молинаро, Дж; Бойс, М; Пол, НЛБ; Каммингс, Р.Д. (17 сентября 2020 г.). «Параллельный синтез гликопептидных библиотек Glyco-SPOT» . Клеточная химическая биология . 27 (9): 1207–1219.e9. doi : 10.1016/j.chembiol.2020.06.007 . ПМЦ 7556346 . ПМИД 32610041 .
- ^ Де Леон, Калифорния; Левин, премьер-министр; Крэйвен, ТВ; Пратт, MR (11 июля 2017 г.). «Связанный с серой аналог O-GlcNAc (S-GlcNAc) является ферментативно стабильным и разумным структурным заменителем O-GlcNAc на уровне пептидов и белков» . Биохимия . 56 (27): 3507–3517. doi : 10.1021/acs.biochem.7b00268 . ПМК 5598463 . ПМИД 28627871 .
- ^ Рамирес, Д.Х.; Аонбангхен, К; Ву, HY; Нафталий, Ж.А.; Тан, С; О'Мира, TR; Ву, СМ (17 апреля 2020 г.). «Разработка трансферазы O-GlcNAc, направленной на близость, для селективного O-GlcNAcylation белка в клетках» . АКС Химическая биология . 15 (4): 1059–1066. doi : 10.1021/acschembio.0c00074 . ПМЦ 7296736 . ПМИД 32119511 .
- ^ Перейти обратно: а б Феррер, Кристина М.; Линч, Томас П.; Соди, Валери Л.; Фальконе, Джон Н.; Шваб, Лусиана П.; Пикок, Даниэль Л.; Вокадло, Дэвид Дж.; Сигровс, Тиффани Н.; Реджинато, Маурисио Х. (5 июня 2014 г.). «O-GlcNAcylation регулирует метаболизм рака и передачу сигналов стресса выживания посредством регуляции пути HIF-1» . Молекулярная клетка . 54 (5): 820–831. doi : 10.1016/j.molcel.2014.04.026 . ISSN 1097-4164 . ПМК 4104413 . ПМИД 24857547 .
- ^ Перейти обратно: а б Ма, З; Вокадло, диджей; Восселер, К. (24 мая 2013 г.). «Гипер-O-GlcNAcylation является антиапоптотическим и поддерживает конститутивную активность NF-κB в клетках рака поджелудочной железы» . Журнал биологической химии . 288 (21): 15121–30. дои : 10.1074/jbc.M113.470047 . ПМЦ 3663532 . ПМИД 23592772 .
- ^ Торрес, ИО; Фухимори, генеральный директор (декабрь 2015 г.). «Функциональная связь между авторами, ластиками и читателями гистонов и метилирования ДНК» . Современное мнение в области структурной биологии . 35 : 68–75. дои : 10.1016/j.sbi.2015.09.007 . ПМЦ 4688207 . ПМИД 26496625 .
- ^ Перейти обратно: а б Чу, CS; Ло, ПВ; Да, ЮХ; Сюй, PH; Пэн, SH; Тенг, ЮК; Канг, ML; Вонг, CH; Хуан, ЖЖ (28 января 2014 г.). «O-GlcNAcylation регулирует стабильность и функцию белка EZH2» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (4): 1355–60. Бибкод : 2014PNAS..111.1355C . дои : 10.1073/pnas.1323226111 . ПМЦ 3910655 . ПМИД 24474760 .
- ^ Ло, ПВ; Ши, Джей-Джей; ЧЧен, Швейцария; Ву, CY; Сюй, ТЛ; Вонг, Швейцария (10 июля 2018 г.). «O-GlcNAcylation регулирует стабильность и ферментативную активность гистон-метилтрансферазы EZH2» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (28): 7302–7307. Бибкод : 2018PNAS..115.7302L . дои : 10.1073/pnas.1801850115 . ПМК 6048490 . ПМИД 29941599 .
- ^ Чжан, Вопрос; Лю, Х; Гао, Вт; Ли, П; Хоу, Дж; Ли, Дж; Вонг, Дж. (28 февраля 2014 г.). «Дифференциальная регуляция семейства диоксигеназ Ten-Eleven Translocation (TET) с помощью O-связанной β-N-ацетилглюкозаминтрансферазы (OGT)» . Журнал биологической химии . 289 (9): 5986–96. дои : 10.1074/jbc.M113.524140 . ПМЦ 3937666 . ПМИД 24394411 .
- ^ Чжан, Цяо; Лю, Сяогуан; Гао, Вэньци; Ли, Пишунь; Хоу, Джингли; Ли, Дживэнь; Вонг, Цземин (28 февраля 2014 г.). «Дифференциальная регуляция семейства диоксигеназ Ten-Eleven Translocation (TET) с помощью O-связанной β-N-ацетилглюкозаминтрансферазы (OGT)» . Журнал биологической химии . 289 (9): 5986–5996. дои : 10.1074/jbc.M113.524140 . ISSN 0021-9258 . ПМЦ 3937666 . ПМИД 24394411 .
- ^ Перейти обратно: а б с Чжу, Гуйчжоу; Чжан, Лю, Сяоцзюань; Сюй, Чивэй; Шэнь, Айгуо (август 2016 г.) . деацетилазы 1 при гепатоцеллюлярной карциноме способствуют прогрессированию рака» . Гликобиология . 26 (8): 820–833. doi : 10.1093/гликоб/cww025 . ISSN 1460-2423 . PMID 27060025 .
- ^ Фонг, Джерри Дж.; Нгуен, Бренда Л.; Бриджер, Роберт; Медрано, Эстела Э.; Уэллс, Лэнс; Пан, Шуцзюань; Сиферс, Ричард Н. (6 апреля 2012 г.). «β-N-ацетилглюкозамин (O-GlcNAc) является новым регулятором митоз-специфического фосфорилирования гистона H3» . Журнал биологической химии . 287 (15): 12195–12203. дои : 10.1074/jbc.M111.315804 . ISSN 0021-9258 . ПМК 3320971 . ПМИД 22371497 .
- ^ Перейти обратно: а б с Фуджики, Р; Хашиба, Ж; Секин, Х; Ёкояма, А; Чиканиши, Т; Ито, С; Имаи, Ю; Ким, Дж; Он, HH (27 ноября 2011 г.). «GlcNAcylation гистона H2B облегчает его моноубиквитинирование» . Природа . 480 (7378): 557–60. Бибкод : 2011Natur.480..557F . дои : 10.1038/nature10656 . ПМЦ 7289526 . ПМИД 22121020 .
- ^ Чен, Кью; Чен, Ю; Биан, К; Фуджики, Р; Ю, Х (24 января 2013 г.). «TET2 способствует O-GlcNAcylation гистонов во время транскрипции гена» . Природа . 493 (7433): 561–4. Бибкод : 2013Natur.493..561C . дои : 10.1038/nature11742 . ПМЦ 3684361 . ПМИД 23222540 .
- ^ Перейти обратно: а б с д Сюй, Цюран; Чэнь, Яли, Хайлун; Чжан, Лэй; Лю, Цингуан (01.05.2014). 9 «AMPK регулирует O-GlcNAcylation гистона» ( 42 . ) 5594–5604 10.1093 / gku236 . ISSN 0305-1048 . : nar . doi : /
- ^ Креппель, ЛК; Харт, GW (5 ноября 1999 г.). «Регуляция цитозольной и ядерной трансферазы O-GlcNAc. Роль тетратрикопептидных повторов» . Журнал биологической химии . 274 (45): 32015–32022. дои : 10.1074/jbc.274.45.32015 . ISSN 0021-9258 . ПМИД 10542233 .
- ^ Перейти обратно: а б Чунг, Вин Д.; Харт, Джеральд В. (9 мая 2008 г.). «AMP-активируемая протеинкиназа и p38 MAPK активируют O-GlcNAcylation нейрональных белков во время дефицита глюкозы» . Журнал биологической химии . 283 (19): 13009–13020. дои : 10.1074/jbc.M801222200 . ISSN 0021-9258 . ПМЦ 2435304 . ПМИД 18353774 .
- ^ Цзоу, Луюнь; Чжу-Маулдин, Сяоюань; Марчес, Ричард Б.; Патерсон, Эндрю Дж.; Лю, Цзянь; Ян, Цинлинь; Чатем, Джон К. (05 октября 2012 г.). «Вызванное лишением глюкозы увеличение белка O-GlcNAcylation в кардиомиоцитах зависит от кальция» . Журнал биологической химии . 287 (41): 34419–34431. дои : 10.1074/jbc.M112.393207 . ISSN 1083-351X . ПМЦ 3464547 . ПМИД 22908225 .
- ^ Тейлор, Родрик П.; Паркер, Глендон Дж.; Хейзел, Марк В.; Соесанто, Юди; Фуллер, Уильям; Яззи, Марла Дж.; Макклейн, Дональд А. (7 марта 2008 г.). «Депривация глюкозы стимулирует модификацию белков O-GlcNAc посредством активации O-связанной N-ацетилглюкозаминилтрансферазы» . Журнал биологической химии . 283 (10): 6050–6057. дои : 10.1074/jbc.M707328200 . ISSN 0021-9258 . ПМИД 18174169 .
- ^ Чен, PH; Смит, Ти Джей; Ву, Дж; Сиссер, П.Дж.; Биснетт, Би Джей; Хан, Ф; Хог, М; Содерблом, Э; Тан, Ф; Маркс, младший; Майор, МБ; Свартс, Б.М.; Бойс, М; Чи, Джен-Цан (01 августа 2017 г.). «Гликозилирование KEAP1 связывает восприятие питательных веществ с передачей сигналов окислительно-восстановительного стресса» . Журнал ЭМБО . 36 (15): 2233–2250. дои : 10.15252/embj.201696113 . ПМЦ 5538768 . ПМИД 28663241 .
- ^ МакГрил, СР; Бхушан, Б; Валески, К; МакГилл, MR; Лебофски, М; Кандел, SE; Уайнфилд, Род-Айленд; Йешке, Х; Захара, штат Невада; Чжан, З; Тан, EP; Слоусон, К; Апте, Ю (01 апреля 2018 г.). «Модуляция уровней O-GlcNAc в печени влияет на повреждение печени, вызванное ацетаминофеном, влияя на образование белковых аддуктов и синтез глутатиона» . Токсикологические науки . 162 (2): 599–610. дои : 10.1093/toxsci/kfy002 . ПМК 6012490 . ПМИД 29325178 .
- ^ Перейти обратно: а б Юзва, ЮАР; Шан, X; Маколи, Миссисипи; Кларк, Т; Скоробогатько Ю.; Восселер, К; Вокадло, диджей (26 февраля 2012 г.). «Увеличение количества O-GlcNAc замедляет нейродегенерацию и стабилизирует тау против агрегации». Химическая биология природы . 8 (4): 393–9. дои : 10.1038/nchembio.797 . ПМИД 22366723 .
- ^ Ченг, X.; Коул, Р.Н.; Зая, Дж.; Харт, GW (26 сентября 2000 г.). «Альтернативное О-гликозилирование/О-фосфорилирование бета-рецептора эстрогена мыши». Биохимия . 39 (38): 11609–11620. дои : 10.1021/bi000755i . ISSN 0006-2960 . ПМИД 10995228 .
- ^ Комер, Финляндия; Харт, GW (3 июля 2001 г.). «Взаимность между O-GlcNAc и O-фосфатом на карбоксильном концевом домене РНК-полимеразы II». Биохимия . 40 (26): 7845–7852. дои : 10.1021/bi0027480 . ISSN 0006-2960 . ПМИД 11425311 .
- ^ Перейти обратно: а б Лю, Фэй; Икбал, Халид; Грундке-Икбал, Инге; Харт, Джеральд В.; Гонг, Чэн-Синь (20 июля 2004 г.). «O-GlcNAcylation регулирует фосфорилирование тау: механизм, участвующий в болезни Альцгеймера» . Труды Национальной академии наук Соединенных Штатов Америки . 101 (29): 10804–10809. Бибкод : 2004PNAS..10110804L . дои : 10.1073/pnas.0400348101 . ISSN 0027-8424 . ПМК 490015 . ПМИД 15249677 .
- ^ Перейти обратно: а б Ян, WH; Ким, Дж. Э.; Нам, HW; Джу, JW; Ким, HS; Ким, Ю.С.; Чо, JW (октябрь 2006 г.). «Модификация р53 О-связанным N-ацетилглюкозамином регулирует активность и стабильность р53». Природная клеточная биология . 8 (10): 1074–83. дои : 10.1038/ncb1470 . ПМИД 16964247 . S2CID 12326082 .
- ^ Перейти обратно: а б Диас, ВБ; Чунг, штат Вирджиния; Ван, З; Харт, GW (7 августа 2009 г.). «Регуляция кальций/кальмодулин-зависимой киназы IV путем модификации O-GlcNAc» . Журнал биологической химии . 284 (32): 21327–37. дои : 10.1074/jbc.M109.007310 . ПМЦ 2755857 . ПМИД 19506079 .
- ^ Перейти обратно: а б с Ма, З; Чокли, Р.Дж.; Восселер, К. (2 июня 2017 г.). «Гипер-O-GlcNAcylation активирует ядерный фактор κ-усилитель легкой цепи активированных B-клеток (NF-κB) посредством взаимодействия с фосфорилированием и ацетилированием» . Журнал биологической химии . 292 (22): 9150–9163. дои : 10.1074/jbc.M116.766568 . ПМЦ 5454098 . ПМИД 28416608 .
- ^ Перейти обратно: а б с Оливье-Ван Стихелен, Стефани; Деэннаут, Ванесса; Бюзи, Армель; Захаюс, Жан-Люк; Гинес, Селин; Мир, Анн-Мари; Эль-Езиди-Белкура, Икрам; Копен, Мари-Кристин; Бурем, Дидье; Лойо, Дени; Феррара, Паскуаль (август 2014 г.). «O-GlcNAcylation стабилизирует β-катенин посредством прямой конкуренции с фосфорилированием треонина 41» . Журнал ФАСЭБ . 28 (8): 3325–3338. дои : 10.1096/fj.13-243535 . ISSN 1530-6860 . ПМК 4101651 . ПМИД 24744147 .
- ^ Перейти обратно: а б Хуан, Сюнь; Пан, Цюмин; Сан, Дэнни; Чен, Вэй; Шен, Айджун; Хуан, Мин; Дин, Цзянь; Гэн, Мэйю (20 декабря 2013 г.). «O-GlcNAcylation кофилина способствует инвазии клеток рака молочной железы» . Журнал биологической химии . 288 (51): 36418–36425. дои : 10.1074/jbc.M113.495713 . ISSN 0021-9258 . ПМЦ 3868755 . ПМИД 24214978 .
- ^ Перейти обратно: а б с Селник, Гарольд Г.; Хесс, Дж. Фред; Тан, Куюэ; Лю, Кун; Шахтер, Джоэл Б.; Баллард, Джанин Э.; Маркус, Джейкоб; Кляйн, Дэниел Дж.; Ван, Сяохай; Пирсон, Мишель; Сэвидж, Мэри Дж.; Каул, Рамеш; Ли, Тонг-Шуан; Вокадло, Дэвид Дж.; Чжоу, Юаньси; Чжу, Юнбао; Му, Чанвэй; Ван, Яодэ; Вэй, Чжунъюн; Бай, Чанг; Даффи, Джозеф Л.; Макихерн, Эрнест Дж. (ноябрь 2019 г.). «Открытие MK-8719, мощного ингибитора O-GlcNAcase как потенциального средства лечения таупатий» . Журнал медицинской химии . 62 (22): 10062–10097. doi : 10.1021/acs.jmedchem.9b01090 . ISSN 1520-4804 . ПМИД 31487175 .
- ^ Швейн, Пол А; Ву, Кристина М (20 марта 2020 г.). «Модификация O-GlcNAc киназ» . АКС Химическая биология . 15 (3): 602–617. дои : 10.1021/acschembio.9b01015 . ПМЦ 7253032 . ПМИД 32155042 .
- ^ Перейти обратно: а б Буллен, Дж.В.; Болсбо, Дж.Л.; Чанда, Д; Шабановиц, Дж; Хант, DF; Нойманн, Д; Харт, GW (11 апреля 2014 г.). «Взаимодействие между двумя незаменимыми чувствительными к питательным веществам ферментами: O-GlcNAc трансферазой (OGT) и AMP-активируемой протеинкиназой (AMPK)» . Журнал биологической химии . 289 (15): 10592–606. дои : 10.1074/jbc.M113.523068 . ПМК 4036179 . ПМИД 24563466 .
- ^ Перейти обратно: а б Ло, Бай; Паркер, Глендон Дж.; Кукси, Роберт С.; Соесанто, Юди; Эванс, Марк; Джонс, Дебора; Макклейн, Дональд А. (9 марта 2007 г.). «Хронический поток гексозамина стимулирует окисление жирных кислот путем активации АМФ-активируемой протеинкиназы в адипоцитах» . Журнал биологической химии . 282 (10): 7172–7180. дои : 10.1074/jbc.M607362200 . ISSN 0021-9258 . ПМИД 17227772 .
- ^ Перейти обратно: а б Соди, ВЛ; Бачигалупа, ЗА; Феррер, CM; Ли, СП; Гокал, Вашингтон; Мухопадхьяй, Д; Веллен, Кентукки; Иван, М; Реджинато, MJ (15 февраля 2018 г.). «Сенсор питательных веществ трансфераза O-GlcNAc контролирует метаболизм раковых липидов посредством регуляции SREBP-1» . Онкоген . 37 (7): 924–934. дои : 10.1038/onc.2017.395 . ПМЦ 5814337 . ПМИД 29059153 .
- ^ Уэллс, Лэнс; Креппель, Лиза К.; Комер, Фрэнк И.; Вадзински, Брайан Э.; Харт, Джеральд В. (10 сентября 2004 г.). «O-GlcNAc-трансфераза находится в функциональном комплексе с каталитическими субъединицами протеинфосфатазы 1» . Журнал биологической химии . 279 (37): 38466–38470. дои : 10.1074/jbc.M406481200 . ISSN 0021-9258 . ПМИД 15247246 .
- ^ Чунг, Вин Д.; Сакабе, Каору; Хаусли, Майкл П.; Диас, Вагнер Б.; Харт, Джеральд В. (5 декабря 2008 г.). «Специфичность субстрата O-связанной бета-N-ацетилглюкозаминилтрансферазы регулируется нацеливанием на миозинфосфатазу и другие взаимодействующие белки» . Журнал биологической химии . 283 (49): 33935–33941. дои : 10.1074/jbc.M806199200 . ISSN 0021-9258 . ПМК 2590692 . ПМИД 18840611 .
- ^ Ян, X.; Су, К.; Роос, доктор медицины; Чанг, К.; Патерсон, Эй Джей; Кадлоу, Дж. Э. (5 июня 2001 г.). «О-связывание N-ацетилглюкозамина с доменом активации Sp1 ингибирует его транскрипционную способность» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (12): 6611–6616. Бибкод : 2001PNAS...98.6611Y . дои : 10.1073/pnas.111099998 . ISSN 0027-8424 . ПМК 34401 . ПМИД 11371615 .
- ^ Ламарр-Винсент, Натан; Се-Уилсон, Линда К. (4 июня 2003 г.). «Динамическое гликозилирование фактора транскрипции CREB: потенциальная роль в регуляции генов» (PDF) . Журнал Американского химического общества . 125 (22): 6612–6613. дои : 10.1021/ja028200t . ISSN 0002-7863 . ПМИД 12769553 .
- ^ Рексач, Джессика Э.; Кларк, Питер М.; Мейсон, Дэниел Э.; Нив, Рэйчел Л.; Питерс, Эрик С.; Се-Уилсон, Линда К. (22 января 2012 г.). «Динамическая модификация O-GlcNAc регулирует CREB-опосредованную экспрессию генов и формирование памяти» . Химическая биология природы . 8 (3): 253–261. дои : 10.1038/nchembio.770 . ISSN 1552-4469 . ПМЦ 3288555 . ПМИД 22267118 .
- ^ Тоулман, Клиффорд А.; Шумахер, Мария А.; Ю, Сок-Хо; Цзэн, Вэньцзе; Кокс, Натан Дж.; Смит, Тимоти Дж.; Содерблом, Эрик Дж.; Уандс, Эмберлин М.; Колер, Дженнифер Дж.; Бойс, Майкл (5 июня 2018 г.). «Структурные основы распознавания O-GlcNAc белками 14-3-3 млекопитающих» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (23): 5956–5961. Бибкод : 2018PNAS..115.5956T . дои : 10.1073/pnas.1722437115 . ISSN 0027-8424 . ПМК 6003352 . ПМИД 29784830 .
- ^ Гинес, Селин; Лемуан, Жером; Михальский, Жан-Клод; Лефевр, Тони (18 июня 2004 г.). «Белок теплового шока 70 кДа обладает регулируемой лектиновой активностью по отношению к O-связанному N-ацетилглюкозамину». Связь с биохимическими и биофизическими исследованиями . 319 (1): 21–26. дои : 10.1016/j.bbrc.2004.04.144 . ISSN 0006-291X . ПМИД 15158436 .
- ^ Чжан, Ф; Су, К; Ян, Х; Боу, Д.Б.; Патерсон, Эй Джей; Кадлоу, Дж. Э. (12 декабря 2003 г.). «Модификация O-GlcNAc является эндогенным ингибитором протеасомы» . Клетка . 115 (6): 715–25. дои : 10.1016/s0092-8674(03)00974-7 . ПМИД 14675536 . S2CID 8221476 .
- ^ Чжан, Фэнсюэ; Ху, Юн; Хуан, Пин; Тоулман, Клиффорд А.; Патерсон, Эндрю Дж.; Кадлоу, Джеффри Э. (3 августа 2007 г.). «Функция протеасомы регулируется циклической AMP-зависимой протеинкиназой посредством фосфорилирования Rpt6» . Журнал биологической химии . 282 (31): 22460–22471. дои : 10.1074/jbc.M702439200 . ISSN 0021-9258 . ПМИД 17565987 .
- ^ Кимбиехетти, Читра Н.; Кржеслак, Анна; С любовью, Дона К.; Ганновер, Джон А. (15 августа 2011 г.). «Изоформа O-GlcNAcase, нацеленная на липид-капли, является ключевым регулятором протеасомы» . Журнал клеточной науки . 124 (Часть 16): 2851–2860. дои : 10.1242/jcs.083287 . ISSN 1477-9137 . ПМК 3148132 . ПМИД 21807949 .
- ^ Захара, Наташа Е.; О'Доннелл, Найл; Чунг, Вин Д.; Мерсер, Джессика Дж.; Март, Джейми Д.; Харт, Джеральд В. (16 июля 2004 г.). «Динамическая модификация нуклеоцитоплазматических белков O-GlcNAc в ответ на стресс. Реакция выживания клеток млекопитающих» . Журнал биологической химии . 279 (29): 30133–30142. дои : 10.1074/jbc.M403773200 . ISSN 0021-9258 . ПМИД 15138254 .
- ^ Икбал, Халид; Лю, Фэй; Гонг, Чэн-Синь; Грундке-Икбал, Инге (декабрь 2010 г.). «Тау при болезни Альцгеймера и связанных с ней таупатиях» . Текущие исследования болезни Альцгеймера . 7 (8): 656–664. дои : 10.2174/156720510793611592 . ISSN 1567-2050 . ПМК 3090074 . ПМИД 20678074 .
- ^ Арнольд, CS; Джонсон, ГВ; Коул, Р.Н.; Донг, DL; Ли, М; Харт, GW (15 ноября 1996 г.). «Связанный с микротрубочками белок тау сильно модифицирован О-связанным N-ацетилглюкозамином» . Журнал биологической химии . 271 (46): 28741–4. дои : 10.1074/jbc.271.46.28741 . ПМИД 8910513 .
- ^ О'Доннелл, Н., Захара, Н., Харт, Г.В., Март Дж.Д. (2004). Ogt-зависимое гликозилирование белка, связанного с Х-хромосомой, является необходимой модификацией функции соматических клеток и жизнеспособности эмбриона. Молекулярная и клеточная биология. 24: 1680-1690. Диол.
- ^ Сандху, Пунам; Ли, Чонхун; Баллард, Джанин; Уокер, Бретань; Эллис, Джоан; Маркус, Джейкоб; Тулан, Рассвет; Дрейер, Дэниел; МакЭвой, Томас; Даффи, Джозеф; Миченер, Мария (июль 2016 г.). «P4-036: Фармакокинетика и фармакодинамика для поддержки клинических исследований MK-8719: ингибитора O-GlcNAcase при прогрессирующем супрануклеарном параличе». Болезнь Альцгеймера и деменция . 12 (7S_Part_21): P1028. дои : 10.1016/j.jalz.2016.06.2125 . S2CID 54229492 .
- ^ Медина, Мигель (11 апреля 2018 г.). «Обзор клинического развития терапии на основе тау» . Международный журнал молекулярных наук . 19 (4): 1160. doi : 10.3390/ijms19041160 . ISSN 1422-0067 . ПМК 5979300 . ПМИД 29641484 .
- ^ Йи, Вэнь; Кларк, Питер М.; Мейсон, Дэниел Э.; Кинан, Мари К.; Хилл, Коллин; Годдард, Уильям А.; Питерс, Эрик С.; Дриггерс, Эдвард М.; Се-Уилсон, Линда К. (24 августа 2012 г.). «Гликозилирование PFK1 является ключевым регулятором роста раковых клеток и центральных метаболических путей» . Наука . 337 (6097): 975–980. дои : 10.1126/science.1222278 . ISSN 0036-8075 . ПМЦ 3534962 . ПМИД 22923583 .
- ^ Колдуэлл, ЮАР; Джексон, СР; Шахриари, Канзас; Линч, ТП; Сетхи, Дж; Уокер, С; Восселер, К; Реджинато, MJ (13 мая 2010 г.). «Сенсор питательных веществ O-GlcNAc Трансфераза регулирует онкогенез рака молочной железы путем воздействия на онкогенный транскрипционный фактор FoxM1» . Онкоген . 29 (19): 2831–42. дои : 10.1038/onc.2010.41 . ПМИД 20190804 . S2CID 25957261 .
- ^ Линч, ТП; Феррер, CM; Джексон, СР; Шахриари, Канзас; Восселер, К; Реджинато, MJ (30 марта 2012 г.). «Критическая роль O-связанной β-N-ацетилглюкозаминтрансферазы в инвазии рака простаты, ангиогенезе и метастазировании» . Журнал биологической химии . 287 (14): 11070–81. дои : 10.1074/jbc.M111.302547 . ПМЦ 3322861 . ПМИД 22275356 .
- ^ Лю, К; Патерсон, Эй Джей; Чин, Е; Кадлоу, Дж. Э. (14 марта 2000 г.). «Глюкоза стимулирует модификацию белка с помощью O-связанного GlcNAc в бета-клетках поджелудочной железы: связь O-связанного GlcNAc с гибелью бета-клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (6): 2820–5. Бибкод : 2000PNAS...97.2820L . дои : 10.1073/pnas.97.6.2820 . ПМК 16013 . ПМИД 10717000 .
- ^ Патхак, Шалини; Дорфмюллер, Хельге К.; Бородкин Владимир С.; ван Аалтен, Даан МФ (25 августа 2008 г.). «Химическое исследование связи между стрептозотоцином, O-GlcNAc и гибелью клеток поджелудочной железы» . Химия и биология . 15 (8): 799–807. doi : 10.1016/j.chembiol.2008.06.010 . ISSN 1074-5521 . ПМЦ 2568864 . ПМИД 18721751 .
- ^ Перейти обратно: а б Восселер, К; Уэллс, Л; Лейн, доктор медицины; Харт, GW (16 апреля 2002 г.). «Повышенное нуклеоцитоплазматическое гликозилирование с помощью O-GlcNAc приводит к резистентности к инсулину, связанной с дефектами активации Akt в адипоцитах 3T3-L1» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (8): 5313–8. Бибкод : 2002PNAS...99.5313V . дои : 10.1073/pnas.072072399 . ПМЦ 122766 . ПМИД 11959983 .
- ^ Ян, Х; Онгусаха, ПП; Майлз, PD; Хавстад, JC; Чжан, Ф; Итак, ВВ; Кадлоу, Дж. Э.; Мичелл, Р.Х.; Олефски, Х.М.; Филд, SJ; Эванс, RMdate=21 февраля 2008 г. (2008 г.). «Фосфоинозитидная сигнальная связь трансферазы O-GlcNAc с резистентностью к инсулину». Природа . 451 (7181): 964–9. Бибкод : 2008Natur.451..964Y . дои : 10.1038/nature06668 . ПМИД 18288188 . S2CID 18459576 .
{{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Уилан, Стивен А.; Лейн, М. Дэниел; Харт, Джеральд В. (1 августа 2008 г.). «Регуляция O-связанной β-N-ацетилглюкозаминтрансферазы посредством передачи сигналов инсулина» . Журнал биологической химии . 283 (31): 21411–21417. дои : 10.1074/jbc.M800677200 . ISSN 0021-9258 . ПМК 2490780 . ПМИД 18519567 .
- ^ Маколи, Миссисипи; Бабб, АК; Мартинес-Флейтес, центральный; Дэвис, Дж.Дж.; Вокадло, диджей (12 декабря 2008 г.). «Повышение глобальных уровней O-GlcNAc в адипоцитах 3T3-L1 путем избирательного ингибирования O-GlcNAcase не вызывает резистентности к инсулину» . Журнал биологической химии . 283 (50): 34687–95. дои : 10.1074/jbc.M804525200 . ПМК 3259902 . ПМИД 18842583 .
- ^ Стефанис, Леонидас (февраль 2012 г.). «А-синуклеин при болезни Паркинсона» . Перспективы Колд-Спринг-Харбора в медицине . 2 (2): а009399. doi : 10.1101/cshperspect.a009399 . ISSN 2157-1422 . ПМЦ 3281589 . ПМИД 22355802 .
- ^ «Гликозилирование как ингибитор агрегации альфа-синуклеина» . Фонд Майкла Дж. Фокса по исследованию болезни Паркинсона | Болезнь Паркинсона . Проверено 5 июня 2020 г.
- ^ Рю, Ин Хён; До, Су-Ил (29 апреля 2011 г.). «Денитрозилирование S-нитрозилированного OGT запускается при LPS-стимулированном врожденном иммунном ответе». Связь с биохимическими и биофизическими исследованиями . 408 (1): 52–57. дои : 10.1016/j.bbrc.2011.03.115 . ISSN 1090-2104 . ПМИД 21453677 .
- ^ Ван, Цимин; Фанг, Пейнинг; Он, Руи; Ли, Мэнци; Ю, Хайшэн; Чжоу, Ли; Йи, Ю; Ван, Фубинг; Ронг, Юань; Чжан, И; Чен, Айдун (15 апреля 2020 г.). «O-GlcNAc-трансфераза способствует цитокиновому шторму, вызванному вирусом гриппа А, воздействуя на регуляторный фактор интерферона-5» . Достижения науки . 6 (16): eaaz7086. Бибкод : 2020SciA....6.7086W . дои : 10.1126/sciadv.aaz7086 . ISSN 2375-2548 . ПМЦ 7159909 . ПМИД 32494619 .
- ^ Каспар А.А.; Райхерт, Дж. М. (сентябрь 2013 г.). «Будущие направления развития пептидной терапии». Открытие наркотиков сегодня . 18 (17–18): 807–17. дои : 10.1016/j.drudis.2013.05.011 . ПМИД 23726889 .
- ^ Левин, премьер-министр; Балана, АТ; Штерхлер, Э; Кул, К; Нода, Х; Зажицка, Б; Дейли, Э.Дж.; Труонг, ТТ; Катрич, В; Гарделла, Ти Джей; Вуттен, Д; Секстон, премьер-министр; Макдональд, П; Пратт, MR (11 сентября 2019 г.). «Инженерия O-GlcNAc пептидов-агонистов GPCR улучшает их стабильность и активность in vivo» . Журнал Американского химического общества . 141 (36): 14210–14219. дои : 10.1021/jacs.9b05365 . ПМК 6860926 . ПМИД 31418572 .
Дальнейшее чтение
[ редактировать ]- Захара, Наташа; Акимото, Ёсихиро; Харт, Джеральд В. (2015), Варки, Аджит; Каммингс, Ричард Д.; Эско, Джеффри Д.; Стэнли, Памела (ред.), « Модификация O-GlcNAc », Основы гликобиологии (3-е изд.), Cold Spring Harbor Laboratory Press, PMID 28876858 .