Сайт-направленный мутагенез
Сайт-направленный мутагенез — это метод молекулярной биологии используется для внесения специфических и преднамеренных мутирующих изменений в последовательность ДНК гена , который и любых генных продуктов . Также называемый сайт-специфическим мутагенезом или олигонуклеотид-направленным мутагенезом , он используется для исследования структуры и биологической активности ДНК , РНК и белковых молекул, а также для белковой инженерии .
Сайт-направленный мутагенез — один из важнейших лабораторных методов создания библиотек ДНК путем введения мутаций в последовательности ДНК. Существует множество методов достижения сайт-направленного мутагенеза, но с уменьшением затрат на синтез олигонуклеотидов в настоящее время иногда используется синтез искусственных генов в качестве альтернативы сайт-направленному мутагенезу. С 2013 года развитие технологии CRISPR /Cas9, основанной на системе защиты прокариот от вирусов, также позволило редактировать геном проводить in vivo . , а мутагенез можно относительно легко [1]
История
[ редактировать ]Ранние попытки мутагенеза с использованием радиации или химических мутагенов были неспецифичными и приводили к случайным мутациям. [2] Аналоги нуклеотидов и другие химические вещества позже использовались для создания локализованных точковых мутаций . [3] примерами таких химических веществ являются аминопурин , [4] нитрозогуанидин , [5] и бисульфит . [6] Сайт-направленный мутагенез был достигнут в 1974 году в лаборатории Чарльза Вейсмана с использованием нуклеотидного аналога N. 4 -гидроксицитидин, индуцирующий переход ГЦ в АТ. [7] [8] Однако эти методы мутагенеза ограничены типом мутации, которую они могут достичь, и они не так специфичны, как более поздние методы сайт-направленного мутагенеза.
В 1971 году Клайд Хатчисон и Маршалл Эджелл показали, что можно получать мутанты с небольшими фрагментами фага φX174 и нуклеазами рестрикции . [9] [10] Позже Хатчисон вместе со своим коллегой Майклом Смитом в 1978 году разработал более гибкий подход к сайт-направленному мутагенезу с использованием олигонуклеотидов в методе удлинения праймера с помощью ДНК-полимеразы. [11] За участие в разработке этого процесса Майкл Смит позже разделил Нобелевскую премию по химии в октябре 1993 года с Кэри Б. Маллисом , который изобрел полимеразную цепную реакцию .
Основной механизм
[ редактировать ]Основная процедура требует синтеза короткого праймера ДНК. Этот синтетический праймер содержит желаемую мутацию и дополняет ДНК-матрицу вокруг сайта мутации, поэтому он может гибридизоваться с ДНК интересующего гена. Мутация может представлять собой одно изменение оснований ( точечная мутация ), множественные изменения оснований, удаление или вставку . Затем одноцепочечный праймер удлиняется с помощью ДНК-полимеразы , которая копирует остальную часть гена. Скопированный таким образом ген содержит мутированный сайт, затем его вводят в клетку-хозяина в векторе и клонируют . Наконец, мутанты отбираются путем секвенирования ДНК, чтобы проверить, содержат ли они желаемую мутацию.
Подходы
[ редактировать ]Исходный метод с использованием расширения с одним праймером оказался неэффективным из-за низкого выхода мутантов. Полученная смесь содержит как исходную немутантную матрицу, так и мутантную цепь, образуя смешанную популяцию мутантных и немутантных потомков. Кроме того, используемая матрица метилирована , в то время как мутантная цепь неметилирована, и мутанты могут быть подвергнуты встречному отбору из-за присутствия системы репарации ошибочных спариваний , которая благоприятствует метилированной ДНК-матрице, что приводит к меньшему количеству мутантов. С тех пор было разработано множество подходов для повышения эффективности мутагенеза.
Существует большое количество методов направленного мутагенеза. [12] хотя большинство из них редко использовались в лабораториях с начала 2000-х годов, поскольку новые методы позволяют использовать более простые и легкие способы введения сайт-специфической мутации в гены.
метод Кункеля
[ редактировать ]В 1985 году Томас Канкель представил метод, который уменьшает необходимость отбора мутантов. [13] Фрагмент ДНК, подлежащий мутации, встраивают в фагмиду, такую как M13mp18/19, а затем трансформируют в штамм E. coli, дефицитный по двум ферментам: dUTPase ( dut ) и урацилдегликозидазе ( udg ). Оба фермента являются частью пути репарации ДНК , который защищает бактериальную хромосому от мутаций путем спонтанного дезаминирования dCTP в dUTP. Дефицит dUTPase предотвращает распад dUTP, что приводит к высокому уровню dUTP в клетке. Дефицит урацилдегликозидазы предотвращает удаление урацила из вновь синтезированной ДНК. с двойным мутантом Поскольку E. coli реплицирует фаговую ДНК, ее ферментативный механизм может, следовательно, неправильно включать dUTP вместо dTTP, в результате чего образуется одноцепочечная ДНК, содержащая некоторое количество урацилов (ssUDNA). ssUDNA экстрагируется из бактериофага, который высвобождается в среду, а затем используется в качестве матрицы для мутагенеза. Олигонуклеотид , содержащий желаемую мутацию, используется для удлинения праймера. Образующаяся гетеродуплексная ДНК состоит из одной родительской немутантной цепи, содержащей dUTP, и мутированной цепи, содержащей dTTP. Затем ДНК трансформируется в Штамм E. coli дикого типа , несущий гены dut и udg . Здесь урацил-содержащая родительская цепь ДНК разрушается, так что почти вся образующаяся ДНК состоит из мутировавшей цепи.
Кассетный мутагенез
[ редактировать ]В отличие от других методов, кассетный мутагенез не требует удлинения праймера с использованием ДНК-полимеразы. В этом методе фрагмент ДНК синтезируется, а затем вставляется в плазмиду. [14] Он включает расщепление ферментом рестрикции участка плазмиды и последующее лигирование пары комплементарных олигонуклеотидов, содержащих мутацию в гене, представляющем интерес для плазмиды. Обычно ферменты рестрикции, которые разрезают плазмиду и олигонуклеотид, одинаковы, что позволяет липким концам плазмиды и вставке лигироваться друг с другом. Этот метод может генерировать мутанты с эффективностью, близкой к 100%, но ограничен наличием подходящих сайтов рестрикции, фланкирующих сайт, который необходимо мутировать.
ПЦР-сайт-направленный мутагенез
[ редактировать ]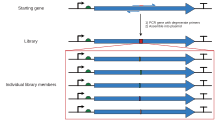
Ограничение сайтов рестрикции при кассетном мутагенезе можно преодолеть с помощью полимеразной цепной реакции с олигонуклеотидными « праймерами », так что может быть создан более крупный фрагмент, охватывающий два удобных сайта рестрикции. Экспоненциальная амплификация в ПЦР дает фрагмент, содержащий желаемую мутацию в достаточном количестве, чтобы его можно было отделить от исходной немутантной плазмиды с помощью гель-электрофореза , который затем можно вставить в исходный контекст с использованием стандартных методов рекомбинантной молекулярной биологии. Существует множество вариаций одной и той же техники. Самый простой метод размещает сайт мутации ближе к одному из концов фрагмента, при этом один из двух олигонуклеотидов, используемых для создания фрагмента, содержит мутацию. Это включает в себя один этап ПЦР, но все же возникает проблема, связанная с необходимостью наличия подходящего сайта рестрикции рядом с сайтом мутации, если только не используется очень длинный праймер. Поэтому в других вариантах используются три или четыре олигонуклеотида, два из которых могут быть немутагенными олигонуклеотидами, которые покрывают два удобных сайта рестрикции и образуют фрагмент, который можно расщепить и лигировать в плазмиду, тогда как мутагенный олигонуклеотид может быть комплементарен участку внутри этого фрагмента далеко от любого удобного сайта рестрикции. Эти методы требуют нескольких этапов ПЦР, чтобы последний лигируемый фрагмент мог содержать желаемую мутацию. Процесс проектирования для создания фрагмента с желаемой мутацией и соответствующими сайтами рестрикции может быть обременительным. Программные инструменты, такие как SDM-Assist [15] может упростить процесс.
Цельноплазмидный мутагенез
[ редактировать ]Для манипуляций с плазмидами другие методы сайт-направленного мутагенеза были в значительной степени вытеснены методами, которые являются высокоэффективными, но относительно простыми, легкими в использовании и коммерчески доступными в виде наборов. Примером таких техник является метод «Quikchange». [16] где пара комплементарных мутагенных праймеров используется для амплификации всей плазмиды в реакции термоциклирования с использованием высокоточной ДНК-полимеразы, не замещающей нити, такой как Pfu полимераза . В результате реакции образуется кольцевая ДНК с надрезами . ДНК-матрица должна быть удалена путем ферментативного расщепления с помощью фермента рестрикции , такого как Dpn I, который специфичен для метилированной ДНК. Вся ДНК, полученная из большинства штаммов Escherichia coli , будет метилирована; матричная плазмида, которая биосинтезируется в E. coli , следовательно, будет переварена, в то время как мутантная плазмида, которая образуется in vitro и, следовательно, неметилирована, останется непереваренной. Обратите внимание, что в этих методах мутагенеза двухцепочечных плазмид, хотя можно использовать реакцию термоциклирования, ДНК не экспоненциально амплифицируется, если два праймера сконструированы таким образом, что они связываются симметрично с одной и той же областью вокруг сайта мутагенеза, как описано в оригинальный протокол. В этом случае амплификация линейна, поэтому описывать процедуру как ПЦР неточно, поскольку цепная реакция отсутствует. Однако если праймеры сконструированы так, чтобы связываться смещенным образом, так что сайт мутагенеза находится близко к 5'-концу обоих праймеров, 3'-область праймеров может также связываться с амплифицированными продуктами и, таким образом, наблюдается экспоненциальное образование продукта. Название «Quikchange» происходит от зарегистрированного товарного знака «QuikChange мутагенез» Stratagene , теперь Agilent Technologies , для наборов для направленного мутагенеза. Метод разработали ученые, работающие в Stratagene. [16]
Обратите внимание, что полимераза Pfu может вытеснять цепь при более высокой температуре растяжения (≥70 ° C), что может привести к неудаче эксперимента, поэтому реакцию удлинения следует проводить при рекомендуемой температуре 68 ° C. Было замечено, что в некоторых приложениях этот метод приводит к вставке нескольких копий праймеров. [17] Вариант этого метода, названный SPRINP, предотвращает этот артефакт и используется в различных типах сайт-направленного мутагенеза. [17]
Другие методы, такие как сканирующий мутагенез олигонаправленных мишеней (SMOOT), могут полуслучайным образом комбинировать мутагенные олигонуклеотиды в мутагенезе плазмид. [18] Этот метод позволяет создавать библиотеки мутагенеза плазмид, начиная от одиночных мутаций и заканчивая комплексным мутагенезом кодонов по всему гену.
in vivo. Методы сайт-направленного мутагенеза
[ редактировать ]- Идеальное преступление [19]
- Трансзамен «всплывающее всплывающее окно»
- Прямая делеция гена и сайт-специфический мутагенез с помощью ПЦР и одного перерабатываемого маркера
- Прямая делеция гена и сайт-специфический мутагенез с помощью ПЦР и одного перерабатываемого маркера с использованием длинных гомологичных участков.
- in vivo с использованием синтетических олигонуклеотидов. Сайт-направленный мутагенез [20]
КРИСПР
[ редактировать ]С 2013 года развитие технологии CRISPR -Cas9 позволило эффективно внедрять различные мутации в геном самых разных организмов. Этот метод не требует места вставки транспозона, не оставляет маркеров, а его эффективность и простота сделали его предпочтительным методом редактирования генома . [21] [22]
Приложения
[ редактировать ]
Сайт-направленный мутагенез используется для создания мутаций, которые могут привести к образованию рационально спроектированного белка с улучшенными или особыми свойствами (т.е. белковой инженерии).
Инструменты исследования – специфические мутации в ДНК позволяют исследовать функцию и свойства последовательности ДНК или белка рациональным подходом. Более того, изменения отдельных аминокислот в белках путем сайт-направленного мутагенеза могут помочь понять важность посттрансляционных модификаций. Например, замена определенного серина (фосфоакцептора) на аланин (фосфо-неакцептор) в белке-субстрате блокирует присоединение фосфатной группы, тем самым позволяя исследовать фосфорилирование. Этот подход был использован для обнаружения фосфорилирования белка CBP киназой HIPK2. [23] Другим комплексным подходом является сайт -насыщенный мутагенез, при котором один кодон или набор кодонов можно заменить всеми возможными аминокислотами в определенных положениях. [24]
Коммерческое применение . Белки можно сконструировать для получения мутантных форм, адаптированных для конкретного применения. Например, обычно используемые стиральные порошки могут содержать субтилизин , форма дикого типа которого содержит метионин, который может окисляться отбеливателем, что значительно снижает активность белка в процессе. [25] Этот метионин может быть заменен аланином или другими остатками, что делает его устойчивым к окислению, тем самым сохраняя активность белка в присутствии отбеливателя. [26]
Синтез генов
[ редактировать ]Поскольку стоимость синтеза ДНК-олигонуклеотидов падает, искусственный синтез полного гена теперь является жизнеспособным методом внесения мутаций в ген. Этот метод позволяет провести обширный мутагенез на нескольких сайтах, включая полную переработку использования кодонов гена для оптимизации его для конкретного организма. [27]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Сюй П.Д., Ландер Э.С., Чжан Ф. (июнь 2014 г.). «Разработка и применение CRISPR-Cas9 для генной инженерии» . Клетка . 157 (6): 1262–78. дои : 10.1016/j.cell.2014.05.010 . ПМЦ 4343198 . ПМИД 24906146 .
- ^ Килби, Би Джей (1995). «Шарлотта Ауэрбах (1899–1994)» . Генетика . 141 (1): 1–5. дои : 10.1093/генетика/141.1.1 . ПМК 1206709 . ПМИД 8536959 .
- ^ Шортл, Д.; Димайо, Д.; Натанс, Д. (1981). «Направленный мутагенез». Ежегодный обзор генетики . 15 : 265–294. дои : 10.1146/annurev.ge.15.120181.001405 . ПМИД 6279018 .
- ^ Карас, И.В.; Макиннес, Массачусетс; Персинг, Д.Х.; Коффино, П.; Мартин-младший, DW (1982). «Механизм мутагенеза 2-аминопурина в клетках Т-лимфосаркомы мыши» . Молекулярная и клеточная биология . 2 (9): 1096–1103. дои : 10.1128/MCB.2.9.1096 . ПМК 369902 . ПМИД 6983647 .
- ^ Макхью, GL; Миллер, CG (1974). «Выделение и характеристика мутантов пролинпептидазы Salmonella typhimurium» . Журнал бактериологии . 120 (1): 364–371. дои : 10.1128/JB.120.1.364-371.1974 . ПМЦ 245771 . ПМИД 4607625 .
- ^ Д. Шортл и Д. Натанс (1978). «Локальный мутагенез: метод создания вирусных мутантов с заменами оснований в заранее выбранных участках вирусного генома» . Труды Национальной академии наук . 75 (5): 2170–2174. Бибкод : 1978PNAS...75.2170S . дои : 10.1073/pnas.75.5.2170 . ПМЦ 392513 . ПМИД 209457 .
- ^ Р. А. Флавелл; Д.Л. Сабо; Э. Ф. Бандл и К. Вайсманн (1975). «Сайт-направленный мутагенез: влияние экстрацистронной мутации на размножение in vitro РНК бактериофага Qbeta» . Proc Natl Acad Sci США . 72 (1): 367–371. Бибкод : 1975PNAS...72..367F . дои : 10.1073/pnas.72.1.367 . ПМК 432306 . ПМИД 47176 .
- ^ Вилли Мюллер; Ганс Вебер; Франсуа Мейер; Чарльз Вайсманн (1978). «Сайт-направленный мутагенез в ДНК: создание точечных мутаций в клонированной ДНК, комплементарной β-глобину, в положениях, соответствующих аминокислотам со 121 по 123». Журнал молекулярной биологии . 124 (2): 343–358. дои : 10.1016/0022-2836(78)90303-0 . ПМИД 712841 .
- ^ Хатчисон III, Калифорния; Эджелл, Миннесота (1971). «Генетический анализ небольших фрагментов дезоксирибонуклеиновой кислоты бактериофага φX174» . Журнал вирусологии . 8 (2): 181–189. doi : 10.1128/JVI.8.2.181-189.1971 . ПМК 356229 . ПМИД 4940243 .
- ^ Маршалл Х. Эджелл, Клайд А. Хатчисон III и Мортон Склер (1972). «Специфические фрагменты эндонуклеазы R дезоксирибонуклеиновой кислоты бактериофага X174» . Журнал вирусологии . 9 (4): 574–582. doi : 10.1128/JVI.9.4.574-582.1972 . ПМК 356341 . ПМИД 4553678 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Хатчисон К.А., Филлипс С., Эджелл М.Х., Гиллам С., Янке П., Смит М. (сентябрь 1978 г.). «Мутагенез в определенном положении последовательности ДНК» (PDF) . Ж. Биол. Хим . 253 (18): 6551–60. дои : 10.1016/S0021-9258(19)46967-6 . ПМИД 681366 .
- ^ Браман, Джефф, изд. (2002). Протоколы мутагенеза in vitro . Методы молекулярной биологии. Том. 182 (2-е изд.). Хумана Пресс. ISBN 978-0896039100 .
- ^ Кункель Т.А. (1985). «Быстрый и эффективный сайт-специфический мутагенез без фенотипического отбора» . Труды Национальной академии наук . 82 (2): 488–92. Бибкод : 1985PNAS...82..488K . дои : 10.1073/pnas.82.2.488 . ПМК 397064 . ПМИД 3881765 .
- ^ Уэллс, Дж.А.; Эстелл, Д.А. (1988). «Субтилизин - фермент, созданный для инженерии». Тенденции биохимических наук . 13 (8): 291–297. дои : 10.1016/0968-0004(88)90121-1 . ПМИД 3154281 .
- ^ Карник, Абхиджит; Карник, Руча; Грефен, Кристофер (2013). «Программное обеспечение SDM-Assist для разработки праймеров для сайт-направленного мутагенеза, вводящих «тихие» сайты рестрикции» . БМК Биоинформатика . 14 (1): 105. дои : 10.1186/1471-2105-14-105 . ISSN 1471-2105 . ПМЦ 3644487 . ПМИД 23522286 .
- ^ Jump up to: а б Папворт, К., Бауэр, Дж. К., Браман, Дж. и Райт, Д. А. (1996). «Сайт-направленный мутагенез за один день с эффективностью> 80%». Стратегии . 9 (3): 3–4.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Эдельхейт, О; Ханукоглу, А; Ханукоглу, я (2009). «Простой и эффективный сайт-направленный мутагенез с использованием двух параллельных реакций с одним праймером для создания мутантов для исследований структуры и функции белка» . БМК Биотехнология . 9:61 . дои : 10.1186/1472-6750-9-61 . ПМК 2711942 . ПМИД 19566935 .
- ^ Черчионе, Дерек; Лавлак, Кэтрин; Тиллотсон, Эрик Л.; Харбински, Фред; ДаСильва, Джен; Келли, Чейз П.; Кестон-Смит, Элиза; Фернандес, Сесилия А.; Майер, Вик Э.; Джаярам, Харихаран; Стейнберг, Барретт Э. (16 апреля 2020 г.). «Библиотеки SMOOT и индуцированная фагами направленная эволюция Cas9 для снижения нецелевой активности» . ПЛОС ОДИН . 15 (4): e0231716. Бибкод : 2020PLoSO..1531716C . дои : 10.1371/journal.pone.0231716 . ISSN 1932-6203 . ПМК 7161989 . ПМИД 32298334 .
- ^ Сторич Ф.; Резник М.А. (2006). «Подход Delitto Perfetto к сайт-направленному мутагенезу и хромосомным перестройкам in vivo с использованием синтетических олигонуклеотидов в дрожжах». Восстановление ДНК, Часть Б. Методы энзимологии. Том. 409. стр. 329–45. дои : 10.1016/S0076-6879(05)09019-1 . ISBN 9780121828141 . ПМИД 16793410 .
- ^ Сторич Ф.; Резник М.А. (2003). «Направленный мутагенез Delitto perfetto в дрожжах с помощью олигонуклеотидов». Генная инженерия . 25 : 189–207. ПМИД 15260239 .
- ^ Дамьен Био-Пеллетье; Винсент Джей Джей Мартин (2016). «Бесшовный сайт-направленный мутагенез генома Saccharomyces cerevisiae с использованием CRISPR-Cas9» . Журнал биологической инженерии . 10 :6. дои : 10.1186/s13036-016-0028-1 . ПМЦ 4850645 . ПМИД 27134651 .
- ^ Сюй С (20 августа 2015 г.). «Применение редактирования генома CRISPR-Cas9 у Caenorhabditis elegans» . Дж Генет Геномикс . 42 (8): 413–21. дои : 10.1016/j.jgg.2015.06.005 . ПМЦ 4560834 . ПМИД 26336798 .
- ^ Ковач К.А., Штайнманн М., Халфон О., Маджистретти П.Дж., Кардино Дж.Р. (ноябрь 2015 г.). «Комплексная регуляция CREB-связывающего белка с помощью протеинкиназы 2, взаимодействующей с гомеодоменом» (PDF) . Сотовая сигнализация . 27 (11): 2252–60. doi : 10.1016/j.cellsig.2015.08.001 . hdl : 10754/594104 . ПМИД 26247811 .
- ^ Ритц, Монтана; Карбалейра Джей Ди (2007). «Итеративный насыщающий мутагенез (ISM) для быстрой направленной эволюции функциональных ферментов». Протоколы природы . 2 (4): 891–903. дои : 10.1038/nprot.2007.72 . ПМИД 17446890 . S2CID 37361631 .
- ^ Стауффер CE, Этсон Д. (10 октября 1969 г.). «Влияние на активность субтилизина окисления остатка метионина» . Журнал биологической химии . 244 (19): 5333–8. дои : 10.1016/S0021-9258(18)63664-6 . ПМИД 5344139 .
- ^ Эстелл Д.А., Грейкар Т.П., Уэллс Дж.А. (10 июня 1985 г.). «Создание фермента с помощью сайт-направленного мутагенеза, устойчивого к химическому окислению» . Журнал биологической химии . 260 (11): 6518–21. дои : 10.1016/S0021-9258(18)88811-1 . ПМИД 3922976 .
- ^ Юрий Худяков, Говард А. Филдс, изд. (25 сентября 2002 г.). Искусственная ДНК: методы и приложения . ЦРК Пресс. п. 13. ISBN 9781420040166 .