Теропода
| Теропода | ||
|---|---|---|
   | ||
| Научная классификация | ||
| Домен: | Эукариоты | |
| Королевство: | животное | |
| Тип: | Хордовые | |
| Клэйд : | Динозаврия | |
| Клэйд : | Саурисхия | |
| Клэйд : | Евзаурисхия | |
| Клэйд : | Теропода Марш , 1881 г. | |
| Подгруппы [1] | ||
| ||



Теропода ( / θ ɪəˈr ɒ p ə də / ; [2] от древнегреческого θηρίο- ποδός [ θηρίον , ( терион ) «дикий зверь»; πούς , ποδός ( pous, podos ) «ступня»], представители которой известны как тероподы , представляют собой динозавров кладу , для которой характерны полые кости, три пальца и когти на каждой конечности. Тероподов обычно относят к группе ящеротазовых динозавров. Изначально они были плотоядными , хотя ряд групп теропод эволюционировали и стали травоядными и всеядными . Тероподы впервые появились в карнийский позднего триаса период 231,4 миллиона лет назад ( млн лет назад ). [3] и включал большинство крупных наземных хищников от ранней юры, по крайней мере, до конца мелового периода , около 66 млн лет назад. В юрском периоде птицы произошли от мелких специализированных целурозавровых теропод и сегодня представлены примерно 11 000 ныне живущими видами.
Биология
[ редактировать ]Черты
[ редактировать ]Были предложены различные синапоморфии Theropoda в зависимости от того, какие таксоны включены в группу. Например, в статье Пола Серено 1999 года предполагается, что тероподы характеризуются такими чертами, как эктоптеригоидная ямка (впадина вокруг эктоптеригоидной кости), внутричелюстной сустав, расположенный внутри нижней челюсти, и сильная внутренняя кавитация внутри костей. [4] Однако, поскольку такие таксоны, как Herrerasaurus, возможно, не являются тероподами, эти черты, возможно, были более широко распространены среди ранних ящеротазовых, а не были уникальными для теропод.
Вместо этого таксоны с более высокой вероятностью пребывания в составе Theropoda могут иметь более специфические черты, такие как выступающее переднечелюстное окно, шейные позвонки с плевроцелями в передней части центра, что приводит к более пневматической шее, пять или более крестцовых позвонков, увеличение запястной кости и дистально вогнутой части большеберцовой кости, среди нескольких других черт, обнаруженных по всему скелету. Как и у ранних зауроподоморфов, второй палец руки теропода увеличен. У тероподов также очень хорошо развиты шаровидные суставы возле шеи и головы. [5] [6]
Большинство теропод принадлежит к кладе Neotheropoda, для которой характерна редукция нескольких костей стопы, в результате чего при ходьбе оставляются трехпалые следы на земле (трехпалые стопы). Цифра V превратилась в остаток на ранних стадиях эволюции теропод и исчезла к концу триаса. Первый палец редуцирован и обычно не касается земли, а в некоторых линиях значительно редуцирован. [7] У них также отсутствует цифра V на руках, и у них есть вилка, также известная как поперечный рычаг. [5] Ранние неотероподы, такие как целофизоиды, имеют заметный изгиб верхней челюсти, известный как субнариальная щель. Авеространы являются одними из наиболее развитых теропод и содержат Tetanurae и Ceratosauria. Хотя некоторые раньше считали целофизоидов и цератозавров принадлежащими к одной группе из-за таких особенностей, как сросшееся бедро, более поздние исследования показали, что более вероятно, что эти особенности были предками неотеропод и были потеряны у базальных столбняков. [8] Авеространов и их близких родственников объединяет полная потеря остатков V пальца, меньшее количество зубов на верхней челюсти, перемещение зубного ряда дальше вниз по верхней челюсти и слезное окно. Авеространы также имеют общие черты бедер и зубов. [9]
Диета и зубы
[ редактировать ]
Тероподы имеют широкий спектр диеты: от насекомоядных до травоядных и плотоядных. Строгое хищничество всегда считалось наследственной диетой теропод как группы, а более широкое разнообразие диет исторически считалось характерной особенностью, свойственной исключительно птичьим тероподам (птицам). Однако открытия конца 20-го и начала 21-го веков показали, что разнообразие диет существовало даже в более базальных линиях. [10] Все ранние находки окаменелостей теропод показали, что они были преимущественно плотоядными . Ископаемые экземпляры ранних теропод, известные ученым в XIX и начале XX веков, обладали острыми зубами с зазубренными краями, способными разрезать плоть, а некоторые экземпляры даже демонстрировали прямые признаки хищнического поведения. Например, окаменелость Compsognathus longipes была найдена с ящерицей в желудке, а экземпляр Velociraptor mongoliensis был найден в бою с Protoceratops andrewsi (тип птицетазового динозавра).
Первыми подтвержденными ископаемыми тероподами, не являющимися плотоядными, были теризинозавры , первоначально известные как «сегнозавры». Эти загадочные динозавры, которых сначала считали прозауроподами , позже оказались узкоспециализированными травоядными тероподами. Теризинозавры обладали большим брюшком для переработки растительной пищи и маленькими головами с клювами и листовидными зубами. Дальнейшее изучение теропод- манирапторов и их родственных связей показало, что теризинозавры были не единственными ранними представителями этой группы, отказавшимися от хищничества. Несколько других линий ранних манирапторов демонстрируют адаптацию к всеядной диете, включая поедание семян (некоторые троодонтиды ) и насекомых (многие авиаланы и альваресзавры ). Овирапторозавры , орнитомимозавры и продвинутые троодонтиды, вероятно, также были всеядными, а некоторые ранние тероподы (такие как Masiakasaurus knopfleri и спинозавриды ), по-видимому, специализировались на ловле рыбы. [11] [12]
Диета во многом определяется морфологией зубов . [13] следы зубов на костях добычи и содержимом кишечника. Известно, что некоторые тероподы, такие как барионикс , лоуринханозавр , орнитомимозавры и птицы, используют гастролиты или желудочные камни.
Зубы теропод у большинства зубчатые, с зазубринами по краям. [14] называется зифодонтом. Другие — пахидонты или фолидонты, в зависимости от формы зуба или зубцов . [14] Морфология зубов достаточно различима, чтобы отличить основные семейства. [13] которые указывают на различные стратегии диеты. Расследование, проведенное в июле 2015 года, показало, что то, что казалось «трещинами» на их зубах, на самом деле было складками, которые помогали предотвратить поломку зубов за счет укрепления отдельных зазубрин во время нападения на добычу. [15] Складки помогли зубам дольше оставаться на месте, особенно когда тероподы стали более крупными и имели большую силу при укусе. [16] [17]
Покровы (кожа, чешуя и перья)
[ редактировать ]
Мезозойские тероподы также были весьма разнообразны по строению кожи и покрову. Перья или перьеподобные структуры (нити) встречаются у большинства родословных теропод (см. Пернатый динозавр ). Однако за пределами целурозавров перья могли быть присущи только молодым, более мелким видам или ограниченным частям животного. Кожа многих более крупных теропод была покрыта мелкими бугристыми чешуйками. У некоторых видов они перемежались более крупными чешуйками с костными ядрами или остеодермами . Этот тип кожи наиболее известен у цератозавра Carnotaurus , который сохранился с обширными отпечатками кожи. [18]
У представителей целурозавров, наиболее далеких от птиц, перья были относительно короткими и состояли из простых, возможно, ветвящихся нитей. [19] Простые нити также наблюдаются у теризинозавров, которые также обладали большими, жесткими перьями, похожими на иголки. У более полностью оперенных теропод, таких как дромеозавриды , чешуя обычно сохраняется только на ногах. Некоторые виды могут иметь смешанные перья и на других участках тела. У скансориоптерикса сохранилась чешуя возле нижней части хвоста. [20] а Джуравенатор , возможно, был преимущественно чешуйчатым с вкраплениями простых нитей. [21] С другой стороны, некоторые тероподы были полностью покрыты перьями, например троодонтид Anchiornis , у которого перья были даже на ступнях и пальцах ног. [22]
На основании взаимосвязи размера зубов и длины черепа, а также сравнения степени изношенности зубов нептичьих теропод и современных лепидозавров делается вывод о том, что тероподы имели губы, защищавшие их зубы снаружи. Визуально морды таких теропод, как дасплетозавр, имели больше сходства с ящерицами, чем с крокодилами, у которых нет губ. [23]
Размер
[ редактировать ]

Тираннозавр на протяжении многих десятилетий был крупнейшим известным тероподом и наиболее известным широкой публике. Однако с момента его открытия был описан ряд других гигантских хищных динозавров, в том числе спинозавр , кархародонтозавр и гиганотозавр . [24] Оригинальные образцы спинозавра (а также новые окаменелости, описанные в 2006 году) подтверждают идею о том, что спинозавр длиннее тираннозавра , показывая, что спинозавр , возможно, был на 3 метра длиннее тираннозавра , хотя тираннозавр все же мог быть более массивным, чем спинозавр . [25] Такие экземпляры, как Сью и Скотти, считаются самыми тяжелыми тероподами , известными науке. До сих пор нет четкого объяснения, почему эти животные стали такими тяжелыми и громоздкими по сравнению с наземными хищниками, существовавшими до них и после них.
Самый крупный из ныне живущих тероподов — обыкновенный страус , высотой до 2,74 м (9 футов) и весом от 90 до 130 кг (200–290 фунтов). [26] Самый маленький нептичий теропод, известный по взрослым особям, - это троодонтид Anchiornis huxleyi , его вес составляет 110 граммов, а длина - 34 сантиметра (1 фут). [22] Если включить современных птиц, колибри-пчелка ( Mellisuga helenae ) наименьшая: весит 1,9 г и длина 5,5 см (2,2 дюйма). [27] [28]
Недавние теории предполагают, что размер тела теропод постоянно уменьшался в течение 50 миллионов лет, в среднем со 163 кг (359 фунтов) до 0,8 кг (1,8 фунта), в конечном итоге превратившись в современных птиц . Это было основано на доказательствах того, что тероподы были единственными динозаврами, которые постоянно уменьшались в размерах, и что их скелеты менялись в четыре раза быстрее, чем у других видов динозавров. [29] [30]
Темпы роста
[ редактировать ]Чтобы оценить темпы роста теропод, ученым необходимо рассчитать возраст и массу тела динозавра. Оба эти показателя могут быть рассчитаны только на основе окаменелых костей и тканей , поэтому регрессионный анализ для прогнозирования используются и темпы роста современных животных в качестве косвенных показателей. Окаменелые кости имеют годичные кольца, которые появляются в результате роста или сезонных изменений, что можно использовать для приблизительного определения возраста на момент смерти. [31] Однако количество колец в скелете может варьироваться от кости к кости, а старые кольца также могут быть потеряны в пожилом возрасте, поэтому ученым необходимо должным образом контролировать эти две, возможно, сбивающие с толку переменные.
Массу тела определить труднее, поскольку костная масса составляет лишь небольшую часть от общей массы тела животных. Один из методов состоит в измерении окружности бедренной кости, которая, как было показано, у нептичьих тероподных динозавров относительно пропорциональна четвероногим млекопитающим. [32] и используйте это измерение как функцию веса тела, поскольку пропорции длинных костей, таких как бедренная кость, растут пропорционально массе тела. [32] Метод использования соотношения костей сохранившихся животных к массе тела для прогнозирования вымерших животных известен как подход масштабирования дошедших до нас (ES). [33] Второй метод, известный как подход объемной плотности (VD), использует полномасштабные модели скелетов, чтобы сделать выводы о потенциальной массе. Подход ES лучше подходит для широкомасштабных исследований, включающих множество образцов, и не требует такого большого количества полного скелета, как подход VD, но подход VD позволяет ученым лучше ответить на большее количество физиологических вопросов о животном, таких как передвижение и центрирование. гравитации. [33]
В настоящее время принято считать, что нептичьи тероподы не демонстрировали скорость роста всей группы, а вместо этого имели различные темпы роста в зависимости от их размера. Однако все нептичьи тероподы имели более высокие темпы роста, чем современные рептилии, даже если современные рептилии увеличены до больших размеров некоторых нептичьих теропод. По мере увеличения массы тела относительная скорость роста также увеличивается. Эта тенденция может быть связана с необходимостью достижения размеров, необходимых для репродуктивной зрелости . [34] Например, одним из самых маленьких известных теропод был Microraptor zhaoianus , который имел массу тела 200 граммов и рос со скоростью примерно 0,33 грамма в день. [35] Сопоставимая рептилия того же размера растет вдвое быстрее. Темпы роста нептичьих теропод среднего размера (100–1000 кг) приближались к темпам роста скороспелых птиц, которые значительно медленнее, чем альтрициальные птицы. Крупные тероподы (1500–3500 кг) росли еще быстрее, примерно так же, как и плакунообразные млекопитающие. [35] Крупнейшие нептичьи тероподы, такие как тираннозавр рекс, имели аналогичную динамику роста с крупнейшим ныне живущим наземным животным, африканским слоном , для которого характерен быстрый период роста до достижения зрелости, за которым следует замедление роста во взрослом возрасте. [36]
Стойка и походка
[ редактировать ]
Поскольку тероподы — чрезвычайно разнообразная группа животных, поза, которую принимали тероподы, вероятно, со временем значительно различалась между различными линиями. [37] Все известные тероподы двуногие , с уменьшенными в длине передними конечностями и специализированными для выполнения самых разных задач (см. ниже). У современных птиц тело обычно находится в несколько вертикальном положении, при этом верхняя часть ноги (бедренная кость) удерживается параллельно позвоночнику, а сила движения вперед создается в колене. Ученые не уверены, как далеко в генеалогическом древе теропод распространился этот тип позы и передвижения. [37]
Нептичьи тероподы были впервые признаны двуногими в 19 веке, прежде чем их родство с птицами получило широкое признание. Считалось, что в этот период тероподы, такие как карнозавры и тираннозавриды, ходили с вертикальными бедрами и шипами в вертикальной, почти прямой позе, используя свои длинные мускулистые хвосты в качестве дополнительной опоры в стойке, подобной кенгуру. [37] Начиная с 1970-х годов биомеханические исследования вымерших гигантских теропод ставят под сомнение эту интерпретацию. Исследования сочленения костей конечностей и относительное отсутствие доказательств волочения хвоста по путям показали, что при ходьбе гигантские длиннохвостые тероподы принимали более горизонтальную позу с хвостом, удерживаемым параллельно земле. [37] [38] Однако ориентация ног этих видов при ходьбе остается спорной. Некоторые исследования подтверждают традиционную вертикально ориентированную бедренную кость, по крайней мере, у крупнейших длиннохвостых теропод. [38] в то время как другие предполагают, что колено обычно сильно сгибалось у всех теропод во время ходьбы, даже у таких гигантов, как тираннозавриды. [39] [40] Вполне вероятно, что у многих вымерших групп теропод существовал широкий диапазон поз тела, стоек и походок. [37] [41]
Нервная система и органы чувств
[ редактировать ]теропод, хотя и редко полные слепки эндокрания По окаменелостям известны . Эндокранию теропод также можно реконструировать по сохранившимся черепам мозга, не повреждая ценные образцы, с помощью компьютерной томографии и программного обеспечения для 3D-реконструкции . Эти находки имеют эволюционное значение, поскольку помогают документировать возникновение неврологии современных птиц из неврологии более ранних рептилий. Увеличение доли головного мозга, занимаемого головным мозгом, по-видимому, произошло с появлением целурозаврии и «продолжалось на протяжении всей эволюции манирапторов и ранних птиц». [42]
Исследования показывают, что тероподы имели очень чувствительную морду. Предполагается, что их могли использовать для определения температуры, пищевого поведения и обнаружения волн. [43] [44]
Морфология передних конечностей
[ редактировать ]
Укороченные передние конечности по отношению к задним были общей чертой среди теропод, особенно у абелизаврид (таких как Карнотавр ) и тираннозаврид (таких как Тираннозавр ). Однако эта черта не была универсальной: спинозавриды имели хорошо развитые передние конечности, как и многие целурозавры. Относительно крепкие передние конечности одного рода, Xuanhanosaurus , позволили Д. Чжимину предположить, что животное могло быть четвероногим. [45] Однако это уже не считается вероятным. [46]
Руки также сильно различаются у разных групп. Наиболее распространенной формой среди нептичьих теропод является придаток, состоящий из трех пальцев; пальцы I, II и III (или, возможно, II, III и IV ) с острыми когтями. Некоторые базальные тероподы, как и большинство цератозавров , имели четыре пальца, а также уменьшенную V пястную кость (например, Dilophosaurus ). У большинства столбняков было три, [а] [47] но у некоторых было еще меньше. [48]
Также считается, что сфера использования передних конечностей у разных семей была разной. Спинозавриды могли использовать свои мощные передние конечности для удержания рыбы. Считается, что некоторые мелкие манирапторы, такие как скансориоптеригиды, использовали свои передние конечности, чтобы лазить по деревьям . [20] Крылья современных птиц используются в основном для полета, хотя у некоторых групп они приспособлены и для других целей. Например, водные птицы, такие как пингвины, используют свои крылья как ласты.
Движение передних конечностей
[ редактировать ]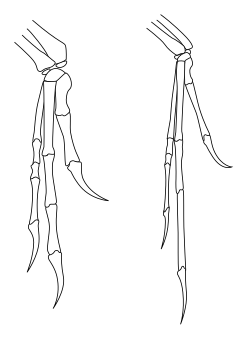
Вопреки тому, как тероподы часто реконструировались в искусстве и популярных средствах массовой информации, диапазон движений передних конечностей теропод был сильно ограничен, особенно по сравнению с ловкостью передних конечностей людей и других приматов . [49] В частности, тероподы и другие двуногие ящеротазовые динозавры (включая двуногих прозауроподов ) не могли пронационировать свои руки, то есть они не могли вращать предплечье так, чтобы ладони были обращены к земле или назад к ногам. У людей пронация достигается за счет движения лучевой кости относительно локтевой кости (двух костей предплечья). Однако у ящеротазовых динозавров конец лучевой кости возле локтя фактически был зафиксирован в борозде локтевой кости, предотвращая любое движение. Движение запястья также было ограничено у многих видов, из-за чего все предплечье и кисть двигались как единое целое с небольшой гибкостью. [50] У теропод и прозауроподов единственный способ, которым ладонь могла быть обращена к земле, - это разведение всей передней конечности вбок, как у птицы, поднимающей крыло. [49]
У карнозавров, подобных Acrocanthosaurus , сама рука сохраняла относительно высокую степень гибкости, с подвижными пальцами. Это справедливо и для более базальных теропод, таких как герреразавры . У целурозавров произошел сдвиг в использовании предплечья: большая гибкость плеча позволила поднять руку в горизонтальную плоскость, а у летающих птиц - еще в большей степени. Однако у целурозавров, таких как орнитомимозавры и особенно дромеозавриды, сама рука потеряла большую часть гибкости, а пальцы стали очень негибкими. Дромеозавриды и другие манирапторы также продемонстрировали повышенную подвижность запястья, не наблюдаемую у других теропод, благодаря наличию специальной кости запястья в форме полумесяца (полулунная запястья), которая позволяла всей кисти сгибаться назад по направлению к предплечью. образ современных птиц. [50]
Палеопатология
[ редактировать ]В 2001 году Ральф Э. Молнар опубликовал обзор патологий костей тероподных динозавров. Он обнаружил патологические особенности у 21 рода из 10 семейств. Патологии были обнаружены у теропод всех размеров тела, хотя в окаменелостях мелких теропод они встречались реже, хотя это может быть артефактом сохранности. Они очень широко представлены в различных частях анатомии теропод. Наиболее распространенными местами сохранившихся повреждений и заболеваний у тероподов являются ребра и хвостовые позвонки . Несмотря на обилие ребер и позвонков, травмы, похоже, «отсутствуют… или очень редки» на основных костях, поддерживающих вес тела, таких как крестец , бедренная кость и большеберцовая кость . Отсутствие сохранившихся повреждений в этих костях позволяет предположить, что они были выбраны эволюцией из-за устойчивости к переломам. Наименее распространенными местами сохранившихся повреждений являются череп и передние конечности, при этом травмы на каждом участке встречаются примерно с одинаковой частотой. Большинство патологий, сохранившихся в окаменелостях теропод, представляют собой остатки травм, таких как переломы, ямки и проколы, часто, вероятно, возникшие в результате укусов. Некоторые палеопатологии теропод, по-видимому, являются свидетельством инфекции , которые, как правило, ограничивались лишь небольшими участками тела животного. Доказательства врожденных пороков развития также были обнаружены в останках теропод. Подобные открытия могут дать информацию, полезную для понимания эволюционной истории процессов биологического развития. Необычные слияния элементов черепа или их асимметрия , вероятно, свидетельствуют о том, что мы исследуем окаменелости чрезвычайно старого человека, а не больного. [51]
Плавание
[ редактировать ]След плавающего теропода, первого в Китае ихногена по имени Characichnos , был обнаружен в формации Фэйтяньшань в провинции Сычуань. [52] Эти новые плавательные следы подтверждают гипотезу о том, что тероподы были приспособлены к плаванию и способны преодолевать умеренно глубокие воды. Следы плавания динозавров считаются редкими окаменелостями и относятся к классу плавательных следов позвоночных, к которому также относятся следы птерозавров и крокодиломорфов . В ходе исследования были описаны и проанализированы четыре полных естественных слепка отпечатков стоп теропод, которые сейчас хранятся в Центре исследований и разработок следов динозавров Хуася (HDT). Эти следы динозавров на самом деле были следами когтей, что позволяет предположить, что этот теропод плавал у поверхности реки и только кончики его пальцев и когтей могли касаться дна. Следы указывают на скоординированное движение влево-вправо и влево-вправо, что подтверждает предположение о том, что тероподы были хорошо скоординированными пловцами. [52]
Эволюционная история
[ редактировать ]
В позднем триасе ряд примитивных прототеропод и тероподов существовали и развивались бок о бок друг с другом.
Самыми ранними и примитивными из тероподовых динозавров были плотоядные и , возможно, герреразавриды Аргентины Eodromaeus . Герреразавры существовали в начале позднего триаса (от позднего карна до раннего нория ). Они были найдены в Северной Америке и Южной Америке, а также, возможно, в Индии и Южной Африке. Герреразавры характеризовались мозаикой примитивных и продвинутых черт. Некоторые палеонтологи в прошлом считали герреразавров представителями Theropoda, в то время как другие предполагали, что эта группа была базальными ящеротазами и, возможно, даже эволюционировала до разделения ящеротазовых и птицетазовых. Кладистический анализ, проведенный после открытия Тавы , еще одного триасового динозавра, предполагает, что герреразавры, вероятно, были ранними тероподами. [53]
Самыми ранними и примитивными однозначными тероподами являются Coelophysoidea . Целофизоиды представляли собой группу широко распространенных, легко сложенных и потенциально стадных животных. Среди них были мелкие охотники, такие как Целофиз и Кампозавр . Эти успешные животные продолжались с позднего карна (начало позднего триаса) до тоара (конец ранней юры ). Хотя в ранних кладистических классификациях они были включены в состав Ceratosauria и считались боковой ветвью более продвинутых теропод. [54] они могли быть предками всех других теропод (что делало их парафилетической группой). [55] [56]
Neotheropoda что означает «новые тероподы») — клада , в которую входят целофизоиды и более развитые тероподы ( , и единственная группа теропод, пережившая триасово-юрское вымирание . Neotheropoda была названа RT Bakker в 1986 году как группа, включающая относительно производные подгруппы теропод Ceratosauria и Tetanurae и исключающая целофизоиды . [57] Однако большинство более поздних исследователей использовали его для обозначения более широкой группы. Neotheropoda была впервые определена Полом Серено как 1998 клада в году как Coelophys плюс современные птицы , которая включает почти всех теропод, за исключением самых примитивных видов. [58] Dilophosauridae ранее считались небольшой кладой внутри Neotheropoda, но позже стали считаться парафилетическими . К ранней юре все неавеространские неотероподы вымерли. [59]
Аверостра (или «птичьи морды») — это клада неотеропод, включающая большинство тероподных динозавров , а именно цератозавров и тетанурий . Это единственная группа постраннеюрских теропод. Важным диагностическим признаком Аверостры является отсутствие пятой пястной кости. Другие ящеротазовые сохранили эту кость, хотя и в значительно уменьшенной форме. [60]
Несколько более продвинутые цератозавры (в том числе цератозавр и карнотавр ) появились в ранней юре и продолжались вплоть до поздней юры в Лавразии . Они соревновались вместе со своими более анатомически развитыми родственниками-столбняками и — в форме линии абелизавров — просуществовали до конца мелового периода в Гондване .
Тетануры снова более специализированы , чем цератозавры. Они подразделяются на базальных Megalosauroidea (поочередно Spinosauroidea ) и более производных Avetheropoda . Мегалозавриды были в основном хищниками от средней юры до раннего мела, а останки их родственников-спинозаврид в основном происходят из пород раннего и среднего мела. Аветероподы, как указывает их название, были более тесно связаны с птицами и снова делятся на Allosauroidea ( разнообразные кархародонтозавры ) и Coelurosauria (очень большая и разнообразная группа динозавров, включая птиц).
Таким образом, в позднеюрском периоде существовало не менее четырех различных линий теропод — цератозавров, мегалозавров, аллозавров и целурозавров, — охотившихся на множество мелких и крупных травоядных динозавров. Все четыре группы дожили до мелового периода, а три из них — цератозавры, целурозавры и аллозавры — дожили до конца периода, когда они были географически разделены: цератозавры и аллозавры в Гондване и целурозавры в Лавразии.
Из всех групп теропод целурозавры были самыми разнообразными. Некоторыми группами целурозавров, которые процветали в меловой период, были тираннозавриды (включая тираннозавра ), дромеозавриды (включая велоцираптора и дейнониха , которые удивительно похожи по форме на старейшую из известных птиц, археоптерикса ), [61] [62] птицеподобные троодонтиды и овирапторозавры, орнитомимозавры (или «страусиные динозавры»), странные травоядные теризинозавры с гигантскими когтями и авиаланы, которые включают современных птиц и являются единственной линией динозавров, пережившей мел -палеогеновое вымирание . [63] Хотя корни этих различных групп находятся в средней юре, они стали многочисленными только в раннем мелу. Некоторые палеонтологи, такие как Грегори С. Пол , предположили, что некоторые или все эти продвинутые тероподы на самом деле произошли от летающих динозавров или протоптиц, таких как археоптерикс , которые потеряли способность летать и вернулись в наземную среду обитания. [64]
Сообщалось также об эволюции птиц от других динозавров-теропод, при этом некоторыми связующими чертами являются и ( по крайней мере , вилка (поперечный рычаг), пневматизированные кости, высиживание яиц у целурозавров) перья . [29] [30] [65]
Классификация
[ редактировать ]История классификации
[ редактировать ]
О. С. Марш придумал название Theropoda (что означает «ноги зверя») в 1881 году. [66] Первоначально Марш назвал Theropoda подотрядом , включив в него семейство Allosauridae , но позже расширил его сферу охвата, переклассифицировав его как отряд, включивший в него широкий спектр «плотоядных» семейств динозавров, включая Megalosauridae , Compsognathidae , Ornithomimidae , Plateosauridae и Anchisauridae (теперь известны как травоядные зауроподоморфы ) и Hallopodidae (впоследствии выявленные как родственники крокодилов). Из-за масштабов отряда Марша Theropoda он пришел на смену предыдущей таксономической группе, которую соперник Марша Э.Д. Коуп создал в 1866 году для хищных динозавров: Goniopoda («наклонные лапы»). [46]
К началу 20 века некоторые палеонтологи, такие как Фридрих фон Хюне , больше не считали плотоядных динозавров естественной группой. Хюэне отказался от названия «Theropoda», вместо этого используя Гарри Сили отряд Saurischia , который Хюэне разделил на подотряды Coelurosauria и Pachypodosauria . Хюне отнес большинство мелких групп теропод к целурозаврии, а крупных теропод и прозауроподов к пахиподозаврии, которую он считал предком завропод ( в то время прозауроподы все еще считались плотоядными из-за неправильной ассоциации рауизуховых черепов и зубов с тела прозауроподов у таких животных, как Тератозавр ). [46] Описывая первого известного дромеозавра ( Dromaeosaurus albertensis ) в 1922 году, [67] У. Д. Мэтью и Барнум Браун стали первыми палеонтологами, исключившими прозауроподов из числа хищных динозавров, и попытались возродить для этой группы название «гониоподы», но другие ученые не приняли ни одно из этих предложений. [46]

В 1956 году термин «Theropoda» снова стал использоваться как таксон , включающий хищных динозавров и их потомков, когда Альфред Ромер реклассифицировал отряд Saurischia на два подотряда: Theropoda и Sauropoda. Это основное разделение сохранилось в современной палеонтологии, за исключением, опять же, Prosauropoda, которого Ромер включил в инфраотряд теропод. Ромер также придерживался разделения на Coelurosauria и Carnosauria (которые он также причислял к инфраотрядам). Эта дихотомия была нарушена открытием в 1969 году дейнониха и дейнохейруса , ни одного из которых нельзя было легко классифицировать как «карнозавров» или «целурозавров». В свете этих и других открытий к концу 1970-х годов Ринчен Барсболд создал новую серию инфраотрядов теропод: Coelurosauria, Deinonychosauria , Oviraptorosauria , Carnosauria, Ornithomimosauria и Deinocheirosauria . [46]
С появлением кладистики и филогенетической номенклатуры в 1980-х годах и их развитием в 1990-х и 2000-х годах стала формироваться более ясная картина взаимоотношений теропод. В 1986 году Жак Готье назвал несколько основных групп теропод, включая кладу Tetanurae , обозначающую одну ветвь основного теропода, разделившуюся с другой группой, Ceratosauria. По мере того как появлялась дополнительная информация о связи между динозаврами и птицами, более похожие на птиц тероподы были сгруппированы в кладу Манираптора (также названную Готье в 1986 году). [54] ). Эти новые разработки также привели к признанию большинством ученых того, что птицы произошли непосредственно от теропод-манирапторов, и, после отказа от рангов в кладистической классификации, к переоценке птиц как подгруппы динозавров-теропод, которые пережили мезозойское вымирание и жили в настоящее. [46]
Основные группы
[ редактировать ]




Ниже приводится упрощенная классификация групп теропод, основанная на их эволюционных взаимоотношениях и организованная на основе списка мезозойских видов динозавров, предоставленного Хольцем. [1] Более подробную версию можно найти в классификации динозавров .Кинжал (†) используется для обозначения групп, в которых нет живых членов.
- † Coelophysoidea (небольшие ранние тероподы; включает Coelophysis и близких родственников)
- † Цератозаврия (как правило, с развитыми рогами, доминирующие южные хищники мелового периода; включает карнотавра и его близких родственников, таких как майюнгазавр и ченанизавр )
- Тетанура («жесткие хвосты»; включает большинство теропод)
- † Megalosauroidea (ранняя группа крупных хищников, включая полуводных спинозавридов)
- † Allosauroidea ( аллозавр и близкие родственники, такие как кархародонтозавр )
- † Megaraptora (группа средних и крупных орионид с неизвестным родством, довольно распространенная в южном полушарии)
- Целурозаврия (пернатые тероподы с разными размерами тела и нишами)
- † Compsognathidae (ранние целурозавры с редуцированными передними конечностями)
- † Tyrannosauridae ( тираннозавр и его близкие родственники; передние конечности были уменьшены)
- † Орнитомимозаврия (« имитаторы страуса »; в основном беззубые; от плотоядных до возможных травоядных)
- Манираптора («похитители рук»; имели длинные тонкие руки и пальцы)
- † Alvarezsauroidea (маленькие насекомоядные с уменьшенными передними конечностями, на каждой из которых имеется по одному увеличенному когтю)
- † Теризинозаврия (двуногие травоядные животные с большими когтями на руках и маленькой головой)
- † Scansoriopterygidae (маленькие древесные манирапторы с длинными третьими пальцами)
- † Овирапторозаврия (в основном беззубые; диета и образ жизни неизвестны)
- † Archaeopterygidae (маленькие крылатые протоптицы)
- † Dromaeosauridae (тероподы малого и среднего размера)
- † Troodontidae (маленькие изящные тероподы)
- Avialae (птицы и вымершие родственники)
- † Omnivoropterygidae (крупные ранние короткохвостые птички)
- † Confuciusornithidae (маленькие беззубые птицы)
- † Энантиорниты (примитивные древесные, летающие птицы)
- Эуорниты (продвинутые летающие птицы)
- † Yanornithiformes (зубистые китайские птицы мелового периода)
- † Hesperornithes (специализированные водоплавающие птицы)
- Аве (современные клюворылые птицы и их вымершие родственники)
Отношения
[ редактировать ]Следующее генеалогическое древо иллюстрирует синтез взаимоотношений основных групп теропод на основе различных исследований, проведенных в 2010-х годах. [69]
Аверостра была названа Г. С. Полом , основанная на апоморфии, в 2002 году как клада определенная как группа, включающая Dromaeosauridae и другие Avepoda с (предком) верхнечелюстным окном ( fenestra promaxillaris ), которое также можно назвать верхнечелюстным окном. [70] дополнительное отверстие в передней внешней стороне верхней челюсти , кости, из которой состоит верхняя челюсть. [71] в 2007 году переопределили его Позже Мартин Эскурра и Жиль Кюни как кладу на основе узлов, содержащую Ceratosaurus nasicornis , Allosaurus fragilis , их последнего общего предка и всех его потомков. [72] Микки Мортимер прокомментировал, что первоначальное определение Пола, основанное на апоморфии, может сделать Аверостру гораздо более широкой кладой, чем узел Цератозавр + Аллозавр , потенциально включая всю Авеподу или более. [73]
Большое исследование ранних динозавров, проведенное доктором Мэтью Г. Бароном, Дэвидом Норманом и Полом М. Барреттом (2017), опубликованное в журнале Nature, показало, что Theropoda на самом деле более тесно связана с Ornithischia , с которой она образовала сестринскую группу в кладе Ornithoscelida. . Эта новая гипотеза также выявила Herrerasauridae как сестринскую группу Sauropodomorpha в обновленном Saurischia и предположила, что морфология гиперплотоядных животных, наблюдаемая у экземпляров теропод и герреразавридов, была приобретена конвергентно . [74] [75] Однако эта филогения остается спорной, и проводится дополнительная работа для выяснения этих взаимоотношений. [76]
Сноски
[ редактировать ]- ^ некоторые роды внутри Avetheropoda имели четыре пальца, см. Однако « Теропода I» на Аветероподах » . Департамент геологии. Университет Мэриленда . 14 июля 2006 г.
См. также
[ редактировать ]- Пернатые динозавры
- Происхождение птиц
- Другие основные клады динозавров
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Хольц, Томас Р. младший (зима 2011 г.). «Приложение» (PDF) . Динозавры: самая полная и актуальная энциклопедия для любителей динозавров всех возрастов (опубликовано в 2012 г.).
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ «Терапода» . Словарь Merriam-Webster.com .
- ^ Мартинес, Рикардо Н.; Серено, Пол С.; Алькобер, Оскар А.; Коломби, Карина Э.; Ренне, Пол Р.; Монтаньес, Изабель П.; Карри, Брайан С. (2011). «Базальный динозавр с начала эры динозавров на юго-западе Пангеи». Наука . 331 (6014): 206–210. Бибкод : 2011Sci...331..206M . дои : 10.1126/science.1198467 . hdl : 11336/69202 . ПМИД 21233386 . S2CID 33506648 .
- ^ Серено, Пол К. (25 июня 1999 г.). «Эволюция динозавров» . Наука . 284 (5423): 2137–2147. дои : 10.1126/science.284.5423.2137 . ISSN 0036-8075 . ПМИД 10381873 .
- ^ Перейти обратно: а б Новас, Фернандо Э.; Аньолин, Федерико Л.; Эскурра, Мартин Д.; Темп Мюллер, Родриго; Мартинелли, Агустин Г.; Лангер, Макс К. (1 октября 2021 г.). «Обзор окаменелостей ранних динозавров из Южной Америки и его филогенетические последствия» . Журнал южноамериканских наук о Земле . 110 : 103341. Бибкод : 2021JSAES.11003341N . дои : 10.1016/j.jsames.2021.103341 . ISSN 0895-9811 .
- ^ Дингус, Лоуэлл; Роу, Тимоти (1998). Ошибочное вымирание: эволюция динозавров и происхождение птиц . Нью-Йорк: WH Freeman. ISBN 978-0-7167-2944-0 .
- ^ Харрис, доктор юридических наук (1997). «Следы четырехпалых теропод и палеомагнитный возраст из водопада Уэтстоун, члена формации Харбелл (верхний мел: маастрихт), северо-западный Вайоминг» (PDF) . Меловые исследования . 18 (1): 139. doi : 10.1006/cres.1996.0053 – через Государственный университет Дикси , Сент-Джордж, Юта.
( Saurexallopus lovei )
- ^ Хольц, Томас Р.; Рей, Луис В. (2007). Динозавры: самая полная и актуальная энциклопедия для любителей динозавров всех возрастов (1-е изд.). Нью-Йорк: Рэндом Хаус. ISBN 978-0-375-82419-7 .
- ^ СМИТ, НАТАН Д.; МАКОВИЦКИ, ПИТЕР Дж.; ХАММЕР, УИЛЬЯМ Р.; КАРРИ, ФИЛИПП Дж. (октябрь 2007 г.). «Остеология Cryolophosaurus ellioti (Dinosauria: Theropoda) из ранней юры Антарктиды и последствия для ранней эволюции теропод» . Зоологический журнал Линнеевского общества . 151 (2): 377–421. дои : 10.1111/j.1096-3642.2007.00325.x . ISSN 1096-3642 .
- ^ Занно, Линдси Э.; Джилетт, Дэвид Д.; Олбрайт, Л. Барри; Титус, Алан Л. (25 августа 2010 г.). «Новый североамериканский теризинозавр и роль травоядных в эволюции хищных динозавров» . Труды Королевского общества Б. 276 (1672): 3505–3511. дои : 10.1098/rspb.2009.1029 . ПМК 2817200 . ПМИД 19605396 .
- ^ Лонгрич, Николас Р.; Карри, Филип Дж. (февраль 2009 г.). « Alvarezsauridae , новый альвареззавр (Dinosauria: Theropoda) из раннего маастрихта Альберты, Канада: значение для систематики и экологии Alvarezsauridae». Меловые исследования . 30 (1): 239–252. Бибкод : 2009CrRes..30..239L . дои : 10.1016/j.cretres.2008.07.005 .
- ^ Хольц, Т.Р. младший; Бринкман, Д.Л.; Чендлер, CL (1998). «Дентальная морфометрия и, возможно, всеядность питания тероподного динозавра Троодона ». ГАЙЯ . 15 : 159–166.
- ^ Перейти обратно: а б Хендрикс, К; Матеус, О (2014). «Abelisauridae (Dinosauria: Theropoda) из поздней юры Португалии и филогения на основе зубных рядов как вклад в идентификацию изолированных зубов теропод». Зоотакса . 3759 : 1–74. дои : 10.11646/zootaxa.3759.1.1 . ПМИД 24869965 . S2CID 12650231 .
- ^ Перейти обратно: а б Хендрикс, Кристоф; Матеус, Октавио; Араужо, Рикардо (2015). «Предлагаемая терминология зубов теропод (Dinosauria, Saurischia)» . Журнал палеонтологии позвоночных (представленная рукопись). 35 (е982797): е982797. Бибкод : 2015JVPal..35E2797H . дои : 10.1080/02724634.2015.982797 . S2CID 85774247 .
- ^ Геггель, Лаура (28 июля 2015 г.). «Один сильный укус: в зубах тираннозавра было секретное оружие» . Фокс Ньюс . Проверено 1 августа 2015 г.
- ^ «Особые зубцы дали хищным динозаврам эволюционное преимущество» . 17 августа 2015 г. Архивировано из оригинала 18 августа 2015 г.
- ^ Бринк, Канзас; Рейс, РР; Леблан, ARH; Чанг, РС; Ли, ЮК; Чан, CC; Хуанг, Т.; Эванс, округ Колумбия (2015). «Развитие и эволюционная новизна зазубренных зубов тероподовых динозавров» . Научные отчеты . 5 : 12338. Бибкод : 2015NatSR...512338B . дои : 10.1038/srep12338 . ПМЦ 4648475 . ПМИД 26216577 . S2CID 17247595 .
- ^ Бонапарт; Новас; Кориа (1990). Carnotaurus sastrei ( Бонапарт ), рогатый, легкого телосложения карнозавр из среднего мела Патагонии (Отчет). Вклад в науку. Том. 416. Округ Лос-Анджелес, Калифорния: Музей естественной истории.
- ^ Гёлих, UB; Кьяппе, LM (16 марта 2006 г.). «Новый плотоядный динозавр с позднеюрского архипелага Зольнхофен» (PDF) . Природа . 440 (7082): 329–332. Бибкод : 2006Natur.440..329G . дои : 10.1038/nature04579 . ПМИД 16541071 . S2CID 4427002 – через doc.rero.ch.
- ^ Перейти обратно: а б Черкас, С.А.; Юань, К. (2002). «Древесный манирапторан из северо-восточного Китая» (PDF) . В Черкасе, SJ (ред.). Пернатые динозавры и происхождение полета . Журнал Музея динозавров. Том. 1. Бландинг, Юта: Музей динозавров. стр. 63–95.
- ^ Гёлих, UB; Тишлингер, Х.; Кьяппе, LM (2006). « Juraventaor starki (Reptilia, Theropoda) новый хищный динозавр из верхней юры Южного Франконского Альба (Южная Германия): находки по анатомии скелета и взвешиванию частей». Археоптерикс . 24 :1–26.
- ^ Перейти обратно: а б Сюй, Х.; Чжао, К.; Норелл, М.; Салливан, К.; Хон, Д.; Эриксон, Г.; Ван, X.; Хан Ф. и Го Ю. (февраль 2009 г.). «Новая окаменелость пернатого динозавра-манираптора, которая заполняет морфологический пробел в птичьем происхождении» . Китайский научный бюллетень . 54 (3): 430–435. Бибкод : 2009SciBu..54..430X . дои : 10.1007/s11434-009-0009-6 . Абстрактный
- ^ Каллен, Томас М.; Ларсон, Дерек Уильям; Виттон, Марк П.; Скотт, Дайан; Махо, Чай; Бринк, Кирстин С.; и др. (30 марта 2023 г.). «Реконструкция лица динозавра-теропода и важность мягких тканей в палеобиологии» . Наука . 379 (6639): 1348–1352. Бибкод : 2023Sci...379.1348C . дои : 10.1126/science.abo7877 . ПМИД 36996202 . S2CID 257836765 .
- ^ Терриен, Ф.; Хендерсон, DM (2007). «Мой теропод больше вашего… или нет: оцениваем размер тела теропод по длине черепа». Журнал палеонтологии позвоночных . 27 (1): 108–115. doi : 10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2 . S2CID 86025320 .
- ^ даль Сассо, К.; Магануко, С.; Бюффето, Э.; Мендес, Массачусетс (2005). «Новая информация о черепе загадочного теропода спинозавра с замечаниями о его размерах и сходстве». Журнал палеонтологии позвоночных (представленная рукопись). 25 (4): 888–896. doi : 10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2 . S2CID 85702490 .
- ^ «Страус» . Африканский фонд дикой природы . Проверено 28 октября 2020 г.
- ^ Брусатте С.Л., О'Коннор Дж.К., Джарвис Э.Д. (октябрь 2015 г.). «Происхождение и разнообразие птиц» . Современная биология . 25 (19): 888–98 рэндов. Бибкод : 2015CBio...25.R888B . дои : 10.1016/j.cub.2015.08.003 . hdl : 10161/11144 . ПМИД 26439352 . S2CID 3099017 .
- ^ Хендри, Лиза (2023). «Являются ли птицы единственными выжившими динозаврами?» . Попечители Музея естественной истории в Лондоне . Проверено 14 апреля 2023 г.
- ^ Перейти обратно: а б Боренштейн, Сет (31 июля 2014 г.). «Исследование прослеживает эволюцию динозавров до ранних птиц» . АП Новости . Проверено 3 августа 2014 г.
- ^ Перейти обратно: а б Зои Гоф (31 июля 2014 г.). «Динозавры регулярно «сжимались» и превращались в птиц» . Би-би-си .
- ^ Падиан, Кевин; де Риклес, Арманд Ж.; Хорнер, Джон Р. (июль 2001 г.). «Темпы роста динозавров и происхождение птиц» . Природа . 412 (6845): 405–408. Бибкод : 2001Natur.412..405P . дои : 10.1038/35086500 . ISSN 1476-4687 . ПМИД 11473307 . S2CID 4335246 .
- ^ Перейти обратно: а б Андерсон, Дж. Ф.; Холл-Мартин, А.; Рассел, Д.А. (1985). «Окружность и вес длинных костей у млекопитающих, птиц и динозавров» . Журнал зоологии . 207 (1): 53–61. дои : 10.1111/j.1469-7998.1985.tb04915.x . ISSN 1469-7998 .
- ^ Перейти обратно: а б Кампионе, Николас Э.; Эванс, Дэвид С. (2020). «Точность и точность оценки массы тела нептичьих динозавров» . Биологические обзоры . 95 (6): 1759–1797. дои : 10.1111/brv.12638 . ISSN 1469-185Х . ПМИД 32869488 . S2CID 221404013 .
- ^ Ли, Эндрю Х.; Вернинг, Сара (15 января 2008 г.). «Половая зрелость растущих динозавров не соответствует моделям роста рептилий» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (2): 582–587. дои : 10.1073/pnas.0708903105 . ISSN 1091-6490 . ПМК 2206579 . ПМИД 18195356 .
- ^ Перейти обратно: а б Эриксон, Грегори М.; Роджерс, Кристина Карри; Йерби, Скотт А. (июль 2001 г.). «Модели роста динозавров и быстрые темпы роста птиц» . Природа . 412 (6845): 429–433. Бибкод : 2001Natur.412..429E . дои : 10.1038/35086558 . ISSN 1476-4687 . ПМИД 11473315 . S2CID 4319534 . (Ошибка: дои : 10.1038/nature16488 , PMID 26675731 , Часы втягивания )
- ^ Хорнер, Джон Р.; Падиан, Кевин (22 сентября 2004 г.). «Возраст и динамика роста тираннозавра рекса» . Труды Королевского общества . Б: Биологические науки. 271 (1551): 1875–1880. дои : 10.1098/rspb.2004.2829 . ISSN 0962-8452 . ПМК 1691809 . ПМИД 15347508 .
- ^ Перейти обратно: а б с д и Хатчинсон, младший (март – апрель 2006 г.). «Эволюция передвижения архозавров» (PDF) . Comptes Рендус Палевол . 5 (3–4): 519–530. Бибкод : 2006CRPal...5..519H . дои : 10.1016/j.crpv.2005.09.002 .
- ^ Перейти обратно: а б Ньюман, Б.Х. (1970). «Стойка и походка плотоядного тираннозавра » . Биологический журнал Линнеевского общества . 2 (2): 119–123. дои : 10.1111/j.1095-8312.1970.tb01707.x .
- ^ Падиан, К.; Олсен, ЧП (1989). «Следы ратитов, поза и походка мезозойских теропод». В Джилетте, округ Колумбия; Локли, М.Г. (ред.). Следы и следы динозавров . Кембридж, Великобритания: Издательство Кембриджского университета. стр. 231–241.
- ^ Пол, GS (1998). «Дизайн, функции и беговые качества конечностей у страусов-имитаторов и тираннозавров». Гайя . 15 : 257–270.
- ^ Фарлоу, Джо; Гейтси, С.М.; Хольц, Т.Р. младший; Хатчинсон-младший; Робинсон, Дж. М. (2000). «Передвижение теропода» . Являюсь. Зоол . 40 (4): 640–663. дои : 10.1093/icb/40.4.640 .
- ^ «Аннотация», в Чуре (2001). Стр. 19. [ нужна полная цитата ]
- ^ Баркер, Крис Тиджани; Нэйш, Даррен; Ньюхэм, Элис; Кацаменис, Орестис Л.; Дайк, Гарет (16 июня 2017 г.). «Сложная нейроанатомия рострума теропода Neovenator salerii с острова Уайт» . Научные отчеты . 7 (1): 3749. Бибкод : 2017НатСР...7.3749Б . дои : 10.1038/s41598-017-03671-3 . ISSN 2045-2322 . ПМК 5473926 . ПМИД 28623335 .
- ^ Кавабе, Соитиро; Хаттори, Соки (3 июля 2022 г.). «Сложная сосудисто-нервная система в зубной кости тираннозавра» . Историческая биология . 34 (7): 1137–1145. Бибкод : 2022HBio...34.1137K . дои : 10.1080/08912963.2021.1965137 . ISSN 0891-2963 . S2CID 238728702 .
- ^ Донг, З. (1984). «Новый динозавр-теропод из средней юры бассейна Сычуани». Позвоночные Палазиатские . 22 (3): 213–218.
- ^ Перейти обратно: а б с д и ж Раухут, Огайо (2003). Взаимосвязь и эволюция базальных тероподов динозавров . Издательство Блэквелл. ISBN 0-901702-79-Х .
- ^ Барта, Делавэр; Несбитт, С.Дж.; Норелл, Массачусетс (2017). «Эволюция кисти ранних тероподных динозавров характеризуется высокой меж- и внутривидовой изменчивостью» . Журнал анатомии . 232 (1): 80–104. дои : 10.1111/joa.12719 . ПМК 5735062 . ПМИД 29114853 .
- ^ «Первый однопалый динозавр» . физ.орг . Январь 2011.
- ^ Перейти обратно: а б Карпентер, К. (2002). «Биомеханика передних конечностей нептичьих динозавров-теропод в условиях хищничества». Сенкенбергиана Летея . 82 (1): 59–76. дои : 10.1007/BF03043773 . S2CID 84702973 .
- ^ Перейти обратно: а б Сентер, П.; Робинс, Дж. Х. (июль 2005 г.). «Амплитуда движений передней конечности динозавра-теропода Acrocanthosaurus atokensis и последствия для хищнического поведения» . Журнал зоологии . 266 (3). Лондон, Великобритания: 307–318. дои : 10.1017/S0952836905006989 .
- ^ Мольнар, Р.Э. (2001). «Палеопатология теропод: обзор литературы». В Танке, DH; Карпентер, К. (ред.). Жизнь мезозойских позвоночных . Издательство Университета Индианы. п. 337-363.
- ^ Перейти обратно: а б Син, Л.Д.; Локли, МГ; Чжан, Япония; и др. (2013). «Новый комплекс следов динозавров раннего мела и первый определенный путь плавания нептичьих тероподов из Китая» . Чин Научный Бык . 58 (19): 2370–2378. Бибкод : 2013ЧСБу..58.2370Х . дои : 10.1007/s11434-013-5802-6 .
- ^ Несбитт, С.Дж.; Смит, Северная Дакота; Ирмис, РБ; Тернер, А.Х.; Даунс, А. и Норелл, Массачусетс (11 декабря 2009 г.). «Полный скелет ящера позднего триаса и ранняя эволюция динозавров». Наука . 326 (5959): 1530–1533. Бибкод : 2009Sci...326.1530N . дои : 10.1126/science.1180350 . ПМИД 20007898 . S2CID 8349110 . .
- ^ Перейти обратно: а б Роу, Т. и Готье, Дж. (1990). «Цератозаврия». В Вейшампеле, Д.Б.; Додсон П. и Осмольска Х. (ред.). Динозаврия . Беркли, Калифорния / Лос-Анджелес, Калифорния / Оксфорд, Великобритания: Издательство Калифорнийского университета. стр. 151–168.
- ^ Мортимер, М. (4 июля 2001 г.). «Диссертация Раухута» . dml.cmnh.org . Архив списков рассылки динозавров.
- ^ Каррано, Монтана; Сэмпсон, SD; Форстер, Калифорния (2002). «Остеология Masiakasaurus knopfleri , небольшого абелизавроида (Dinosauria: Theropoda) из позднего мела Мадагаскара». Журнал палеонтологии позвоночных . 22 (3): 510–534. doi : 10.1671/0272-4634(2002)022[0510:TOOMKA]2.0.CO;2 . S2CID 85655323 .
- ^ Баккер, RT (1986). Ересь динозавров (PDF) . Нью-Йорк, штат Нью-Йорк: Уильям Морроу – через doc.rero.ch.
- ^ Серено, П. (1998). «Обоснование филогенетических определений с применением к таксономии динозавров более высокого уровня». Трактаты. Новогодний альбом по геологии и палеонтологии (ежегодный). 210 : 41–83. дои : 10.1127/njgpa/210/1998/41 .
- ^ Марш, Адам Д.; Роу, Тимоти Б. (2020). «Всесторонняя анатомическая и филогенетическая оценка Dilophosaurus wetherilli (Dinosauria, Theropoda) с описаниями новых экземпляров из формации Кайента в северной Аризоне» . Журнал палеонтологии . 94 (С78): 1–103. Бибкод : 2020JPal...94S...1M . дои : 10.1017/jpa.2020.14 . S2CID 220601744 .
- ^ Сассо, Криштиану Даль; Магануко, Симоне; Кау, Андреа (19 декабря 2018 г.). «Старейший цератозавр (Dinosauria: Theropoda) из нижней юры Италии проливает свет на эволюцию трехпалой руки птиц» . ПерДж . 6 : е5976. дои : 10.7717/peerj.5976 . ISSN 2167-8359 . ПМК 6304160 . ПМИД 30588396 .
- ^ Остром, Дж. Х. (1969). «Остеология Deinonychus antirhopus , необычного теропода из нижнего мела Монтаны». Бюллетень естественной истории Музея Пибоди . 30 :1–165.
- ^ Пол, GS (1988). Хищные динозавры мира . Нью-Йорк, штат Нью-Йорк: ISBN Simon and Schuster Co. 0-671-61946-2 .
- ^ Дингус, Л.; Роу, Т. (1998). Ошибочное вымирание: эволюция динозавров и происхождение птиц . Фриман.
- ^ Пол, GS (2002). Воздушные динозавры: эволюция и потеря способности летать у динозавров и птиц . Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса. ISBN 0-8018-6763-0 .
- ^ Ли, MichaelS.Y.; Кау, Андреа; Нэйш, Даррен; Дайк, Гарет Дж. (1 августа 2014 г.). «Постоянная миниатюризация и анатомические инновации у динозавровых предков птиц». Наука . 345 (6196): 562–566. Бибкод : 2014Sci...345..562L . дои : 10.1126/science.1252243 . ПМИД 25082702 . S2CID 37866029 .
- ^ Марш, О.К. (1881 г.). «Главные персонажи американских юрских динозавров. Часть V.». Американский журнал науки и искусства . 3. 21 (125): 417–423. Бибкод : 1881AmJS...21..417M . дои : 10.2475/ajs.s3-21.125.417 . S2CID 219234316 .
- ^ Мэтью, Вашингтон ; Браун, Б. (1922). «Семейство Deinodontidae с уведомлением о новом роде из мелового периода Альберты». Бюллетень Американского музея естественной истории . 46 : 367–385.
- ^ Андерсон, Тед Р. (2006). Биология вездесущего домового воробья: от генов к популяциям. Оксфорд: Издательство Оксфордского университета. ISBN 0-19-530411-X .
- ^ Хендрикс, К.; Хартман, SA; Матеус, О. (2015). «Обзор открытий и классификации нептичьих теропод». Журнал палеонтологии позвоночных PalArch . 12 (1): 1–73.
- ^ Карр, Томас (22 октября 2013 г.). «Остеология VIII: Анторбитальная полость» . Тираннозавроида центральная . Проверено 2 апреля 2024 г.
- ^ Пол, GS (2002). Динозавры воздуха . Балтимор, Мэриленд / Лондон, Великобритания: Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-6763-7 – через книги Google.
- ^ Эскурра, доктор медицины; Куни, Г. (2007). «Целофизоид Lophostropheus airelensis , gen. nov.: Обзор систематики «Liliensternus» airelensis из триасово-юрских обнажений Нормандии (Франция)» . Журнал палеонтологии позвоночных . 27 (1): 73–86. doi : 10.1671/0272-4634(2007)27[73:TCLAGN]2.0.CO;2 – через Researchgate.net.
- ^ Мортимер, Микки (15 ноября 2010 г.). «Аверостра и Авепода» . Блог базы данных Theropod . Проверено 6 сентября 2020 г.
- ^ Барон, МГ; Норман, Д.Б.; Барретт, премьер-министр (2017). «Новая гипотеза взаимоотношений динозавров и ранней эволюции динозавров» (PDF) . Природа . 543 (7646): 501–506. Бибкод : 2017Natur.543..501B . дои : 10.1038/nature21700 . ПМИД 28332513 . S2CID 205254710 .
- ^ «Новое исследование потрясает корни генеалогического древа динозавров» . cam.ac.uk. Кембридж, Великобритания: Кембриджский университет . 22 марта 2017 г.
- ^ Лангер, MC; Эскурра, доктор медицины; Раухут, OWM; Бентон, MJ; Нолл, Ф.; Макфи, BW; и др. (2017). «Распутывание генеалогического древа динозавров» . Природа . 551 (7678): Е1–Е3. Бибкод : 2017Natur.551E...1L . дои : 10.1038/nature24011 . hdl : 1983/d088dae2-c7fa-4d41-9fa2-aeebbfcd2fa3 . ПМИД 29094688 . Архивировано из оригинала 9 июля 2023 года . Проверено 8 июля 2023 г.
Внешние ссылки
[ редактировать ]

















| Базы данных органов управления : Национальные |
|---|