Теропода
| Теропода | ||
|---|---|---|
  | ||
| Научная классификация | ||
| Домен: | Эукариота | |
| Королевство: | Животное | |
| Филум: | Chordata | |
| Клада : | Динозаурия | |
| Клада : | Сауриския | |
| Клада : | Теропода Марш , 1881 | |
| Подгруппы [ 1 ] | ||
| ||




Theropoda ( / θ ɒ ɒ P d / /; [ 2 ] от древнегреческого θηρίο- ποΔός [ θηρίον , ( терация ) "Дикий зверь"; πούς , ποΔός ( pous, podos ) "Foot"), члены которой известны как тероподы , является существующей динозавров кладкой , которая характеризуется пустыми костями и тремя пальцами ног и когтей на каждой конечности. Тероподы, как правило, классифицируются как группа сауришских динозавров. Они были в восточных количествах плотоядных , хотя ряд тероподных групп эволюционировали, чтобы стать травоядными и всеядными . Theropods впервые появился в возрасте карнианского позднего триаса периода 231,4 миллиона лет назад ( MA ) [ 3 ] и включал в себя большинство крупных наземных хищников от ранней юры, по крайней мере, до конца мела , около 66 млн. Лет. В юрском периоде птицы развивались из небольших специализированных колурозаврских теропод, и сегодня представлены около 11 000 живых видов.
Биология
[ редактировать ]Функции
[ редактировать ]Различные синапоморфии для Theropoda были предложены на основе того, какие таксоны включены в группу. Например, документ 1999 года, проведенный Полом Серено, предполагает, что тероподы характеризуются такими признаками, как эктоптеригоидная ямка (депрессия вокруг эктоптеригоидной кости), внутриманикулярного сустава, расположенного в нижней челюсти, и экстремальная внутренняя кавитация в костях. [ 4 ] Однако, поскольку таксоны, такие как Herrerasaurus, могут не быть тероподами, эти черты, возможно, были более широко распространены среди ранних сауришцев, а не уникальными для теропод.
Вместо этого таксоны с более высокой вероятностью в пределах тероподы могут иметь более специфические признаки, такие как выдающиеся перспективные фенестра, шейные позвонки с плеврокоэлями в передней части центра, ведущей к более пневматической шейке, пяти или более сакральным позвонкам, увеличению за заплеской кости и дистально вогнутой части голени, среди нескольких других признаков, найденных по всему скелету. Как и ранние сауроподоморфы, вторая цифра в руке теропода увеличена. Тероподы также имеют очень хорошо развитый шариковой и гнездо рядом с их шеей и головой. [ 5 ] [ 6 ]
Большинство теропод принадлежат к кладке Neotheropoda, характеризующемуся уменьшением нескольких костей фута, что оставляет три следов на землю на земле, когда они ходят (Tridactyl Feet). Цифр V была уменьшена до остатка в начале эволюции Theropod и исчезла по позднему триасу. Цифра I уменьшается и, как правило, не касается земли и значительно уменьшается в некоторых линиях. [ 7 ] Им также не хватает цифры V на руках, и они разработали фуркулу, которая в противном случае называется пошлин. [ 5 ] Ранние неотероподы, такие как колофизоиды, имеют заметный перевод в верхней челюсти, известный как субнарийский разрыв. Averosrans являются одними из наиболее полученных теропод и содержат столбняки и цератозаврию. В то время как некоторые использовались для того, чтобы рассматривать коелофизоиды и цератозавры в пределах одной и той же группы из -за таких признаков, как плавленое бедро, более поздние исследования показали, что более вероятно, что это были признаки, предпринимаемые для неотеропод, и были потеряны в базальных столбцах. [ 8 ] Averostrans и их близкие родственники объединены за счет полной потери любых остатков v цифры, меньше зубов в верхней челюсти, движения зубного ряда дальше вниз по верхней челюсти и слезного фенестра. Averostrans также делятся функциями в бедрах и зубах. [ 9 ]
Диета и зубы
[ редактировать ]
Тероподы демонстрируют широкий спектр диет, от насекомых до травоядных и плотоядных. Строгие плотоядные камни всегда считались диетой наследственной диеты для теропод в качестве группы, а более широкий спектр диет исторически считался характерным исключительным для птичьих теропод (птиц). Тем не менее, открытия в конце 20 -го и начале 21 -го веков показали, что различные диеты существовали даже в более базальных линиях. [ 10 ] Все ранние находки ископаемых теропод показали, что они в первую очередь плотоядные . Окаменелые образцы ранних теропод, известные ученым в 19 -м и начале 20 -го веков, обладали острыми зубами с зубчатыми краями для разрезания плоти, и некоторые образцы даже показали прямые доказательства хищного поведения. Например, был обнаружен ископаемый копсогнат -лонгипс с ящерицей в животе, а экземпляр Velociraptor mongoliensis был найден запертым в бою с протокератопс Эндрюси (тип орнитишского динозавра).
Первыми подтвержденными непрерывными ископаемыми тероподами были обнаружены теризинозавры , первоначально известные как «сегензавры». Впервые считали, что это просауроподы , эти загадочные динозавры позже оказались высокоспециализированными, травоядными тероподами. Теризинозавры обладали большими животными для переработки пищи растений и небольшими головами с клювами и зубами в форме листьев. Дальнейшее изучение манирапторовских теропод и их отношения показало, что теризинозавры были не единственными ранними членами этой группы, которые отказались от хитрости. Несколько других линий ранних манирапторов демонстрируют адаптацию для всеядную диету, в том числе сеяние семян (некоторые Troodontids ) и насекомые (многие авиаланцы и альварессавры ). Овираптозавры , орнитомимозавры и продвинутые troodontids, вероятно, также были всеядными, и некоторые ранние тероподы (такие как Masiakasaurus nopfleri и спинозавриды ), по -видимому, специализируются на ловке рыбы. [ 11 ] [ 12 ]
Диета в значительной степени выведена морфологией зубов , [ 13 ] Зубные знаки на костях добычи и содержание кишечника. Известно, что некоторые теропод, такие как Baryonyx , Lourinhanosaurus , Ornithomimosaurs и Birds, используют гастролиты или желудочные камни.
Большинство тероподных зубов похожи на лезвие, с зубцами по краям, [ 14 ] называется Ziphodont. Другие - это Pachydont или Folidont в зависимости от формы зуба или зубчатых зубцов . [ 14 ] Морфология зубов достаточно отличная, чтобы отделить основные семьи отдельно, [ 13 ] которые указывают на разные стратегии диеты. Расследование в июле 2015 года обнаружило, что то, что казалось «трещинами» в их зубах, на самом деле были складками, которые помогли предотвратить разрушение зубов, укрепляя отдельные зубцы, когда они атаковали свою добычу. [ 15 ] Складки помогли зубам оставаться на месте дольше, особенно когда тероподы превратились в большие размеры и имели больше силы в их укусе. [ 16 ] [ 17 ]
Интюг (кожа, весы и перья)
[ редактировать ]
Мезозойские тероподы также были очень разнообразны с точки зрения текстуры кожи и покрытия. Перья или перья, похожие на конструкции (филаменты), подтверждаются в большинстве линий теропод (см. Перовой динозавр ). Однако за пределами коелурозавров перья могли быть ограничены молодыми, более мелкими видами или ограниченными частями животного. У многих более крупных теропод имели кожу, покрытую небольшими, ухабистыми чешуйками. У некоторых видов они были вкраплены с большими масштабами с костными ядрами или остеодермами . Этот тип кожи наиболее известен в Ceratosaur Carnotaurus , который был сохранен с обширными впечатлениями от кожи. [ 18 ]
Линии колурозавра, наиболее отдаленные от птиц, имели перья, которые были относительно короткие и состоящие из простых, возможно, ветвящихся нитей. [ 19 ] Простые филаменты также наблюдаются у теризинозавров, которые также обладали большими, жесткими «перо» перья. Более полные пернатые тероподы, такие как Dromaeosaurids , обычно сохраняют весы только на ногах. У некоторых видов могут быть смешанные перья в других местах тела. Scanoriopteryx сохранил весы возле нижней части хвоста, [ 20 ] и Juravenator , возможно, был преимущественно чешуйчатым с некоторыми простыми нитями, перемежающимися. [ 21 ] С другой стороны, некоторые тероподы были полностью покрыты перьями, такими как Troodontid anciornis , у которых даже были перья на ногах и ногах. [ 22 ]
Основываясь на взаимосвязи между размером зуба и длиной черепа, а также на сравнении степени износа зубов неавийских теропод и современных лепидозавров , делается вывод, что у теропод были губы, которые защищали их зубы снаружи. Визуально, у мордов таких теропод, как Daspletosaurus больше сходства с ящерицами, чем у крокодилов, в которых отсутствуют губы. [ 23 ]
Размер
[ редактировать ]

Тираннозавр был на протяжении многих десятилетий самым большим известным тероподом и наиболее известным широкой публике. Однако с момента его открытия было описано ряд других гигантских плотоядных динозавров, включая спинозавр , Carcharodontosaurus и Giganotosaurus . [ 24 ] Оригинальные образцы спинозавра (а также новые окаменелости, описанные в 2006 году), подтверждают идею о том, что спинозавра длиннее тиранозавра , показывая, что спинозавра , возможно, на 3 метра длиннее тиранозавра , хотя тираннозавр все еще может быть более массивным, чем спинозавр . [ 25 ] Образцы, такие как Сью и Скотти, как оцениваются самые тяжелые тероподы, известные науке. По -прежнему нет четкого объяснения того, почему эти животные стали такими тяжелыми и громоздкими по сравнению с сухопутными хищниками, которые появились до и после них.
Самым большим существующим тероподом является общий страус , высотой до 2,74 м (9 футов) и весом от 90 до 130 кг (200 - 290 фунтов). [ 26 ] Наименьшим тероподом без авиалан, известным у взрослых образцов, является Troodontid Anciornis Huxleyi , на 110 граммах веса и длиной 34 сантиметра (1 фут). [ 22 ] Когда входят современные птицы, пчелиная колибри ( Mellisuga Helenae ) наименьшая при длине 1,9 г и 5,5 см (2,2 дюйма). [ 27 ] [ 28 ]
Недавние теории предполагают, что размер теропода непрерывно сокращался в течение 50 миллионов лет, в среднем 163 килограмма (359 фунтов) до 0,8 килограмма (1,8 фунта), в конечном итоге превращаясь в более чем 11 000 видов современных птиц . Это было основано на доказательствах того, что тероподы были единственными динозаврами, которые стали постоянно меньше, и что их скелеты изменились в четыре раза быстрее, чем у других видов динозавров. [ 29 ] [ 30 ]
Темпы роста
[ редактировать ]Чтобы оценить темпы роста теропод, ученым необходимо рассчитать как возраст, так и массу тела динозавра. Обе эти меры могут быть рассчитаны только с помощью ископаемой кости и ткани , поэтому регрессионный анализ и существующие темпы роста животных, поскольку прокси используются для прогнозирования. Окаменелые кости демонстрируют рост кольца, которые появляются в результате роста или сезонных изменений, которые можно использовать для приблизительного возраста во время смерти. [ 31 ] Тем не менее, количество колец в скелете может варьироваться от кости к кости, а старые кольца также могут быть потеряны в позднем возрасте, поэтому ученым необходимо должным образом контролировать эти две, возможно, смешанные переменные.
Масса тела сложнее определить, поскольку костная масса представляет собой лишь небольшую долю общей массы тела животных. Один из методов состоит в том, чтобы измерить окружность бедра, которая, как было показано в неавийских динозаврах, относительно пропорциональными квадрупедным млекопитающим, млекопитающие, [ 32 ] и используйте это измерение как функцию массы тела, так как пропорции длинных костей, таких как бедренная кости, растут пропорционально массой тела. [ 32 ] Метод использования существующей кости животных, пропорциональных к соотношению массы тела для прогнозирования вымерших животных, известен как подход существующего масштаба (ES). [ 33 ] Второй метод, известный как подход объемной плотности (VD), использует полномасштабные модели скелетов для вывода о потенциальной массе. Подход ES лучше для исследований с широким диапазоном, включая множество образцов и не требует столько полного скелета, как подход VD, но подход VD позволяет ученым лучше отвечать на более физиологические вопросы о животном, таких как локомоция и центр гравитации. [ 33 ]
Текущий консенсус заключается в том, что неавийские тероподы не демонстрировали широкий рост группы, но вместо этого имели разнообразные скорости в зависимости от их размера. Тем не менее, все неавийские тероподы имели более высокие темпы роста, чем существующие рептилии, даже когда современные рептилии увеличиваются до большого размера некоторых неавийских теропод. По мере увеличения массы тела относительный темпы роста также увеличивается. Эта тенденция может быть связана с необходимостью достижения размера, необходимого для репродуктивной зрелости . [ 34 ] Например, одним из самых маленьких известных теропод был микрораптор Zhaoianus , который имел массу тела 200 граммов, вырос на скорости приблизительно 0,33 грамма в день. [ 35 ] Сопоставимая рептилия того же размера растет на половине этой скорости. Скорости роста неавийских теропод среднего размера (100–1000 кг) приблизились к скоростям прекочеальных птиц, которые намного медленнее, чем пешеходные птицы. Крупные теропод (1500–3500 кг) росли еще быстрее, аналогично тарифам, отображаемым эвритными млекопитающими. [ 35 ] Крупнейшие неавийские тероподы, такие как Tyrannosaurus Rex, имели аналогичную динамику роста с самым большим живым наземным животным сегодня, африканским слоном , который характеризуется быстрым периодом роста до зрелости, впоследствии сопровождается замедлением роста зрелого возраста. [ 36 ]
Позиция и походка
[ редактировать ]
Как чрезвычайно разнообразная группа животных, поза, принятая тероподами, вероятно, значительно различалась между различными линиями во времени. [ 37 ] Все известные тероподы являются двуноженными , с передними, уменьшенными в длину и специализированы для широкого спектра задач (см. Ниже). У современных птиц тело обычно удерживается в несколько вертикальном положении, а верхняя нога (бедренная кость) удерживается параллельно позвоночнику и с передней силой локомоции, созданной на колене. Ученые не уверены, как далеко в Тероподном семейном древе этот тип осанки и передвижения распространяется. [ 37 ]
Неавийские тероподы были впервые признаны двуногими в течение 19-го века, прежде чем их отношения с птицами были широко приняты. , что тероподы, такие как карнозавры и тиранозавриды, ходили с вертикальными бедрами и шипами в вертикальной, почти прямой осанке, используя свои длинные мускулистые хвосты в качестве дополнительной поддержки в кангуру, похожих на триподальную позицию. В течение этого периода считалось [ 37 ] Начиная с 1970 -х годов, биомеханические исследования вымерших гигантских теропод ставят под сомнение эту интерпретацию. Исследования артикуляции костей конечностей и относительного отсутствия доказательств трека для перетаскивания хвоста показали, что при ходьбе гигантские, длиннохвостые тероподы приняли бы более горизонтальную осанку с хвостом, удерживаемым параллельным земле. [ 37 ] [ 38 ] Однако ориентация ног у этих видов во время ходьбы остается спорной. Некоторые исследования поддерживают традиционную вертикально ориентированную бедру, по крайней мере, в крупнейших длиннохвостых тероподах, [ 38 ] В то время как другие предполагают, что колено обычно было сильно сгибается во всех тероподах во время ходьбы, даже гиганты, такие как тиранозавриды. [ 39 ] [ 40 ] Вполне вероятно, что широкий спектр позов, позиций и походки существовал во многих вымерших группах теропод. [ 37 ] [ 41 ]
Нервная система и чувства
[ редактировать ]Несмотря на то, что редкие, полные составы тероподской эндокрании известны из окаменелостей. Эндокрания Theropod также может быть реконструирована из сохраненных случаев мозга без повреждения ценных образцов с использованием компьютерного томографического сканирования и программного обеспечения для реконструкции 3D . Эти находки имеют эволюционное значение, потому что они помогают документировать появление неврологии современных птиц из более ранних рептилий. Увеличение доли мозга, занятого мозгом, по -видимому, произошло с появлением Coelurosauria и «продолжалось на протяжении всей эволюции манирапторов и ранних птиц». [ 42 ]
Исследования показывают, что у теропод были очень чувствительными мордами. Предполагается, что они могли быть использованы для обнаружения температуры, поведения питания и обнаружения волн. [ 43 ] [ 44 ]
Морфология передней основы
[ редактировать ]
Укороченные передние конечности по отношению к задним ногам были общей признаком среди теропод, особенно в абелизавридах (таких как карнотавр ) и тиранозавридах (таких как тираннозавр ). Эта черта, однако, была не универсальной: спинозавриды имели хорошо разработанные передние мысли, как и многие колурозавры. Относительно надежные передники одного рода, Сюанханозавр , привели Д. Жиминга, чтобы предположить, что животное могло быть четвероногим. [ 45 ] Тем не менее, это больше не считается вероятным. [ 46 ]
Руки также очень разные среди разных групп. Наиболее распространенной формой среди неавийских теропод является придаток, состоящий из трех пальцев; цифры I, II и III (или, возможно, II, III и IV ), с острыми когтями. Некоторые базальные тероподы, как и большинство церотозавров , имели четыре цифры, а также уменьшенный метакарпал V (например, Dilophosaurus ). У большинства тетануран было три, [ А ] [ 47 ] Но у некоторых было еще меньше. [ 48 ]
Считается, что объем использования передней части передних манец также отличался среди разных семей. Спинозавриды могли бы использовать свои мощные передние конечности, чтобы держать рыбу. Считается, что некоторые маленькие манирапторы, такие как сканериоптеригиды, использовали свои передники для подъема на деревьях . [ 20 ] Крылья современных птиц используются в основном для полета, хотя они адаптированы для других целей в определенных группах. Например, водные птицы, такие как пингвины, используют свои крылья в качестве ластаков.
Передалка
[ редактировать ]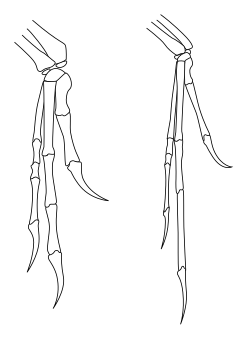
В отличие от того, как тероподы часто реконструированы в искусстве и популярных средах, диапазон движений тероподных передних марок был серьезно ограничен, особенно по сравнению с ловкостью людей и других приматов . [ 49 ] В частности, теропод и другие двуногих сауришских динозавров (включая двуногие просауроподы ) не могли проиграть их руки, то есть они не могли повернуть предплечье, чтобы пальмы обращались к земле или назад к ногам. У людей пронация достигается движением радиуса относительно локтевой кости (две кости предплечья). Однако у сауришских динозавров конец радиуса возле локтя был фактически заперт в канавке локтевой кости, предотвращая любое движение. Движение на запястье также было ограничено во многих видах, заставляя все предплечье и руку перемещаться как единое целое с небольшой гибкостью. [ 50 ] У Theropods и Prosauropods единственным способом для ладонь было лицом к земле, которое было бы путем бокового расщепления всей передней основы, как в птице, поднимающей свое крыло. [ 49 ]
У карнозавров, таких как AcroCanthosaurus , сама рука сохранила относительно высокую степень гибкости с подвижными пальцами. Это также относилось к более базальным тероподам, таким как эрреразавры . Коелурозавры показали сдвиг в использовании предплечья, с большей гибкостью у плеча, позволяя поднять руку к горизонтальной плоскости, и, в более высокой степени у летающих птиц. Тем не менее, у коелурозавров, таких как орнитомимозавры и особенно дромозавриды, сама рука потеряла наибольшую гибкость, с очень негибкими пальцами. Dromaeosaurids и другие манираптоны также показали повышенную подвижность на запястье, не наблюдаемое в других тероподах, благодаря наличию специализированной запястье в полуминке (полуза-заплеска), которая позволила всей руке складываться назад к предплечью в манера современных птиц. [ 50 ]
Палеопатология
[ редактировать ]В 2001 году Ральф Э. Молнар опубликовал обследование патологий в кости динозавра теропод. Он нашел патологические особенности в 21 родах из 10 семей. Патологии были обнаружены в тероподах всего размера тела, хотя они были менее распространены у окаменелостей мелких теропод, хотя это может быть артефактом сохранения. Они очень широко представлены в разных частях анатомии теропод. Наиболее распространенными участками сохранившихся травм и заболеваний у динозавров теропода являются ребра и хвостовые позвонки . Несмотря на то, что они обильны в ребрах и позвонках, травмы, по -видимому, «отсутствуют ... или очень редко» на первичном весе тела, поддерживающих кости, такие как крестцы , бедренная кость и большеберцовая кость . Отсутствие сохранившихся травм в этих костях предполагает, что они были выбраны эволюцией для сопротивления поломке. Наименее распространенными участками сохранившейся травмы являются череп и передняя часть передней основы, причем травмы происходят примерно на одинаковой частоте на каждом участке. Большинство патологий, сохранившихся в ископаемых тероподах, являются остатками травм, таких как переломы, ямы и проколы, часто вероятно, возникающие с укусами. Некоторые тероподные палеопатологии, по -видимому, свидетельствуют инфекции , которые имели тенденцию ограничиваться только небольшими областями тела животного. Свидетельство о врожденных пороках развития также были обнаружены в останках теропод. Такие открытия могут предоставить информацию, полезную для понимания эволюционной истории процессов биологического развития. Необычные слияния в черепных элементах или асимметриях в том же, вероятно, свидетельствуют о том, что кто -то изучает ископаемые чрезвычайно старого человека, а не больного. [ 51 ]
Плавание
[ редактировать ]Трасса плавательного теропода, первого в Китае Ichnogenus, названной Harxichnos , была обнаружена на формировании Фейтианшан в Сычуани. [ 52 ] Эти новые плавающие трассы подтверждают гипотезу о том, что тероподы были адаптированы к плаванию и способны пройти умеренно глубокую воду. Плава динозавров считаются редкими окаменелостями и являются одними из классов плавания позвоночных, которые также включают в себя птерозавры и крокодиломорфы . В исследовании описаны и проанализировали четыре полных природных форм тероподных отпечатков ног, которые в настоящее время хранятся в Центре исследований и разработок в области исследований и разработок Huaxia (HDT). Эти следы динозавров были на самом деле следамить, которые предполагают, что этот теропод плавал возле поверхности реки, и только кончики ее пальцев ног и когтей могли коснуться дна. Треки указывают на скоординированную, слева правую, слева правую прогрессию, которая подтверждает предположение, что тероподы были хорошо скоординированными пловцами. [ 52 ]
Эволюционная история
[ редактировать ]
Во время позднего триаса существовали и эволюционировали ряд примитивных прото-летопод и динозавров с тероподом.
примитивными из динозавров теропода были плотоядные эодромея и, возможно, эррерасавриды Аргентины Самыми ранними и самыми . Эрреразавры существовали во время раннего позднего триаса (поздний карнейский до раннего ) . Они были найдены в Северной Америке и Южной Америке, а также, возможно, также Индия и Южная Африка. Эрреразавры характеризовались мозаикой примитивных и передовых особенностей. Некоторые палеонтологи в прошлом считали, что эррерасавры являются членами Theropoda, в то время как другие теоретизировали, что группа является базальными саурисами, и, возможно, даже развивались до сауриша-эрнитишского раскола. Кладистический анализ после открытия Тавы , еще одного триасового динозавра, предполагает, что эрреразавры, вероятно, были ранними тероподами. [ 53 ]
Самыми ранними и самыми примитивными однозначными тероподами являются Coelophysoidea . Коелофизоиды были группой широко распространенных, слегка построенных и потенциально общительных животных. Они включали маленьких охотников, таких как колофиз и Кампозавр . Эти успешные животные продолжали от покойного карна (раннего позднего триаса) до Toarcian (поздний ранний юрский период ). Хотя в ранних кладистических классификациях они были включены в Ceratosauria и считались боковой ветвью более продвинутых теропод, [ 54 ] Возможно, они были наследственными для всех других теропод (что сделало бы их парафилетической группой). [ 55 ] [ 56 ]
Neotheropoda (что означает «новые теропод») - это клада , которая включает в себя колофизоиды и более продвинутые теропод динозавры , и является единственной группой теропод, которые пережили триасик -джурассуя . Neotheropoda была названа RT Bakker в 1986 году как группу, включая относительно полученные подгруппы Theropod Ceratosauria и Tetanurae , и исключая колофизоиды . [ 57 ] Тем не менее, большинство более поздних исследователей использовали его для обозначения более широкой группы. Neotheropoda был впервые определен как клада Полом Серено в 1998 году как Coelophysis Plus Modern Birds , который включает в себя почти все тероподы, за исключением наиболее примитивных видов. [ 58 ] Dilophosauridae ранее считалась небольшой кладой в Neotheropoda, но впоследствии считался парафилетическим . В ранней юре все неотеропод не авертрана вымерли. [ 59 ]
Averostra (или «Птичья морда») - это клада в Neotheropoda, которая включает в себя большинство тероподных динозавров , а именно Ceratosauria и Tetanurae . Он представляет собой единственную группу пост-ранга юрских теропод. Одной из важных диагностических особенностей Averostra является отсутствие пятого метакарпала. Другие сауришцы сохранили эту кость, хотя и в значительно уменьшенной форме. [ 60 ]
Несколько более продвинутые цератозавры (включая Ceratosaurus и Carnotaurus ) появились во время ранней юры и продолжались до поздней юры в Лауразии . Они соревновались вместе со своими более анатомически продвинутыми родственниками из тетанурана и - в форме линии Абелисавра , закрепленной до конца меловой в Гондване .
Tetanurae снова более специализированы , чем цератозавры. Они подразделяются на базальную мегалозавроиду (альтернативно спинозавроида ) и более полученную avetheropoda . Megalosauridae были в первую очередь средней юрской и ранней мела -хищники, а останки их родственников спинозавридов в основном из ранних и средних мела. Avetheropoda, как указывает их название, были более тесно связаны с птицами и снова разделены на Allosauroidea ( разнообразные карчародотозавры ) и Coelurosauria (очень большая и разнообразная группа динозавров, включая птиц).
Таким образом, во время поздней юры было не менее четырех отдельных линий теропод - сератозавров, мегалозавров, аллозавров и коелурозавров - на изобилии мелких и крупных травоядных динозавров. Все четыре группы выжили в меловом виде, и три из них - цератозавры, колурозавры и аллозавры - заставили конец периода, когда они были географически отделены, цератозавры и аллозавры в Гондване и коелурозавра в Лауразии.
Из всех тероподных групп коелурозавры были самыми разнообразными. Некоторые группы колурозавров, которые процветали во время меловых, были тиранозавридами (включая тиранозавр ), Дромаозавриды (включая велоцираптор и деиноних , которые удивительно похожи по форме с самой старой известной птицей, археоптерикс ), [ 61 ] [ 62 ] Птицы, похожие на Troodontids и Oviraptorosaurs, орнитомимозавры (или «страусные динозавры»), странные гигантские травоядные теризинозавры и авиаланы, которые включают современные птицы и являются единственным происхождением динозавров для выживших к креетуско-палеогенскому исчезновению события . [ 63 ] В то время как корни этих различных групп встречаются в средней юре, они стали обильными только во время раннего мелового. Несколько палеонтологов, таких как Грегори С. Пол , предположили, что некоторые или все эти продвинутые тероподы фактически произошли от летающих динозавров или прото-птиц, таких как Archeopteryx , которые потеряли способность летать и вернулись к наземной среде обитания. [ 64 ]
Также сообщалось о эволюции птиц из других динозавров теропода, причем некоторые из связующих особенностей являются фуркула (протекание), пневматизированные кости, размышления яиц ) и (по крайней мере, в колурозаврах, по крайней мере перьями . [ 29 ] [ 30 ] [ 65 ]
Классификация
[ редактировать ]История классификации
[ редактировать ]
OC Marsh придумал имя Theropoda (то есть «ноги зверя») в 1881 году. [ 66 ] Марш изначально назвал Theropoda в качестве подборка , включающего семейство Allosauridae , но впоследствии расширил свои масштабы, переоценив его как приказ, включив в себя широкий спектр «плотоядных» семейств динозавров, включая мегалозауриды , Compsognathidae , Ornithomimidae , Titalosauridae и Anchisaurae (теперь известно как травоядные сауроподоморфы ) и Hallopodidae (впоследствии выявлены как родственники крокодилов). Из -за масштаба приказа Марша Theropoda он заменил предыдущую таксономическую группу, которую соперник Марша Эд Коуп создал в 1866 году для плотоядных динозавров: Goniopoda («Угловые ноги»). [ 46 ]
К началу 20 -го века некоторые палеонтологи, такие как Фридрих фон Хуен , больше не считали плотоядными динозаврами, которые сформировали естественную группу. Хуен отказался от названия «Theropoda», вместо этого, используя Гарри Сили приказ Сауришиа , который Хуэн разделил на подчиненные Coelurosauria и Pachypodosauria . Хуен поместил большинство небольших тероподных групп в колурозаврию, а также крупные тероподы и просоуроподы в пахиподозаврию, которые он считал наследственным для ( в то время просоуроподы все еще считались плотоядными, из -за неконтроли Сауроподы Просауроподские тела, у животных, таких как Teratosaurus ). [ 46 ] Описывая первую известную Dromaeosaurid ( Dromaeosaurus albertensis ) в 1922 году, [ 67 ] В.Д. Мэтью и Барнум Браун стали первыми палеонтологами, которые исключили просауропод из плотоядных динозавров, и попытались возродить имя «Гониопода» для этой группы, но другие ученые не приняли ни одного из этих предложений. [ 46 ]

В 1956 году «Theropoda» вернулась к использованию-как таксон , содержащий плотоядных динозавров и их потомков-когда Альфред Ромер повторно классифицировал приказ Сауришию в два подчинения: Теропода и Сауропода. Это базовое разделение выжило в современную палеонтологию, за исключением, опять же, Prosauropoda, которую Ромер включил в качестве недостатка теропод. Ромер также сохранил разделение между Coelurosauria и Carnosauria (которое он также считал инфрадориями). Эта дихотомия была расстроена открытием Deinonychus и Deinocheirus в 1969 году, ни один из которых не мог быть легко классифицирован как «карнозавры» или «Coelurosaurs». В свете этих и других открытий, к концу 1970 -х годов Ринчен Барсболд создал новую серию тероподных инфрадартов: Coelurosauria, Deinonychosauria , Овираптозаурия , Карнозаурия, Орнитомимозаврия и Deinocheorosauria . [ 46 ]
С появлением кладистики и филогенетической номенклатуры в 1980 -х годах и их развитием в 1990 -х и 2000 -х годах началась более четкая картина отношений теропод. Жак Готье назвал несколько основных групп теропод в 1986 году, в том числе Clade Tetanurae для одной ветви базового разделения Theropod с другой группой, Ceratosauria. По мере того, как больше информации о связи между динозаврами и птицами выявилось, более похожие на птицы тероподы были сгруппированы в Кладе Манираптора (также названным Готье в 1986 году. [ 54 ] ) Эти новые события также поступили с признанием среди большинства ученых, что птицы возникли непосредственно из манирапторанских теропод и, по отказу от рядов в кладистической классификации, с переоценкой птиц в качестве подмножества динозавров, которые пережили мезозои настоящий. [ 46 ]
Основные группы
[ редактировать ]




Ниже приводится упрощенная классификация тероподных групп на основе их эволюционных отношений и организованная на основе списка видов мезозойских динозавров, предоставленных Holtz. [ 1 ] Более подробную версию можно найти в классификации динозавров . Кинжал (†) используется для обозначения групп без живых членов.
- † Coelophysoidea (малые, ранние тероподы; включает в себя коелофиз и близких родственников)
- † Ceratosauria (как правило, искаженно рогается, доминирующие южные плотоядные животные мела; включает Carnotaurus и близких родственников, таких как Majungasaurus и Chenanisaurus )
- Tetanurae («Жесткие хвосты»; включает в себя большинство теропод)
- † Megalosauroidea (ранняя группа крупных плотоядных животных, включая полу-квадратные спинозавриды)
- † Allosauroidea ( Allosaurus и близкие родственники, такие как Carcharodontosaurus )
- † Megaraptora (группа средних и крупных орионидов с неизвестным сродством, довольно распространенная в южном полушарии)
- Coelurosauria (пернатые тероподы, с различными размерами тела и нишами)
- † Compsognathidae (Ранние колурозавры с уменьшенными передниками)
- † Tyrannosauridae ( Tyrannosaurus и близкие родственники; имели уменьшенные передние конечности)
- † Ornithomimosauria (" страус -мимимика"; в основном беззубые; плотоядные к возможным травоядным животным)
- Манираптора («Ручные захватыватели»; имели длинные, тонкие руки и пальцы)
- † Alvarezsauroidea (небольшие насекомоядные с уменьшенными передним мажок, каждая из которых имеет один увеличенный коготь)
- † Therizinosauria (двуногие травоядные животные с большими когтями рук и небольшими головами)
- † Scanoriopterygidae (маленькие, древесные манирапторы с длинными третьими пальцами)
- † Oviraptorosauria (в основном беззубая; их диета и образ жизни неясны)
- † Archaeopterygidae (маленькие, крылатые протобирды)
- † DROMAEOSAURIDAE (тероподы из малых до средних размеров)
- † Troodontidae (маленькие, графильные тероподы)
- Avialae (птицы и вымершие родственники)
- † Omnivoropterygidae (крупные, ранние короткохвостые авиаляны)
- † Confuciusornithidae (маленькие беззубые птицы)
- † Enantiornithes (примитивные деревья, летающие птицы)
- Euornithes (продвинутые летающие птицы)
- † Yanornithiformes (зубчатые меловые китайские птицы)
- † Hesperornithes (специализированные водные птицы)
- Авес (современные, черновые птицы и их вымершие родственники)
Отношения
[ редактировать ]Следующее семейное древо иллюстрирует синтез взаимосвязей основных тероподных групп на основе различных исследований, проведенных в 2010 -х годах. [ 69 ]
Аверостра была названа Г.С. Полом в 2002 году в качестве апоморфической клады, определяемой как группа, включающая Dromaeosauridae и другие Avepoda , с (предком с) промиксиллярным фенестрам ( Fenestra Promaxillaris ), который также может быть назван в качестве верхнего фенестры, фенестра, фенестра, Fenestra Promaxillaris), который также можно назвать челюстным фенестром, Fenestra) [ 70 ] Дополнительное отверстие в передней внешней стороне верхней челюсти , кости, которая составляет верхнюю челюсть. [ 71 ] Позже он был изменен Мартином Эзкуррой и Жилем Куни в 2007 году в качестве клады на основе узлов, содержащей Ceratosaurus nasicornis , Allosaurus fragilis , их последний общий предок и все его потомки. [ 72 ] Микки Мортимер прокомментировал, что оригинальное определение апоморфии Павла может сделать Averostra гораздо более широкой кладкой, чем Ceratosaurus + узел Allosaurus , что потенциально включает в себя все Avepoda или более. [ 73 ]
Большое исследование ранних динозавров доктора Мэтью Дж. Барона, Дэвида Нормана и Пола М. Барретта (2017), опубликованных в журнале Nature, предположил, что Theropoda фактически более тесно связана с Ornithischia , с которой она сформировала родственную группу в рамках Clade Ornithoscelida Полем Эта новая гипотеза также восстановила Herrerasauridae в качестве сестринской группы в Sauropodomorpha в переосмысленной Saurischia и предположила, что морфологии гиперкарнивора, которые наблюдаются в образцах теропод и эрреразавридов, были приобретены . [ 74 ] [ 75 ] Тем не менее, эта филогения остается спорной, и выполняется дополнительная работа, чтобы прояснить эти отношения. [ 76 ]
Сноски
[ редактировать ]- ^ Некоторые роды в Avetheropoda , однако, имели четыре цифры, см. " Theropoda I" на avetheropoda " . Отдел геологии. Университет Мэриленда . 14 июля 2006 г.
Смотрите также
[ редактировать ]- Пернатые динозавры
- Происхождение птиц
- Другие крупные клады динозавра
Ссылки
[ редактировать ]- ^ Jump up to: а беременный Хольц, Томас Р., младший (зима 2011). «Приложение» (PDF) . Динозавры: самая полная, современная энциклопедия для любителей динозавров всех возрастов (опубликовано 2012).
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ «Теропода» . Merriam-Webster.com Словарь . Мерриам-Уэбстер.
- ^ Мартинес, Рикардо Н.; Серено, Пол С.; Alcober, Oscar A.; Коломби, Карина Э.; Ренне, Пол Р.; Монтаньес, Изабель П.; Керри, Брайан С. (2011). «Базальный динозавр с рассвета эпохи динозавра на юго -западе Пангея». Наука . 331 (6014): 206–210. Bibcode : 2011sci ... 331..206M . doi : 10.1126/science.1198467 . HDL : 11336/69202 . PMID 21233386 . S2CID 33506648 .
- ^ Серено, Пол С. (25 июня 1999 г.). «Эволюция динозавров» . Наука . 284 (5423): 2137–2147. doi : 10.1126/science.284.5423.2137 . ISSN 0036-8075 . PMID 10381873 .
- ^ Jump up to: а беременный Novas, Fernando E.; Agnolin, Federico L.; Эзкурра, Мартин Д.; Temp Müller, Rodrigo; Martinelli, Agustín G.; Лангер, Макс С. (1 октября 2021 г.). Полем Американская земля 110 103341. Bibcode : : doi : 10.1016/j.jsames . ISSN 0895-9811 .
- ^ Динг, Лоуэлл; Роу, Тимоти (1998). Ошибочное вымирание: эволюция динозавра и происхождение птиц . Нью -Йорк: WH Freeman. ISBN 978-0-7167-2944-0 .
- ^ Харрис, JD (1997). «Четырехногенные тероподные следы и палеомагнитный век от члена Whetstone Falls Formation (Верхний меловой: маастрихтский), северо-западный Вайоминг» (PDF) . Мерашные исследования . 18 (1): 139. doi : 10.1006/cres.1996.0053 - Виа Государственного университета Дикси , Сент -Джордж, Юта.
( Saurexallopus lovei )
- ^ Хольц, Томас Р.; Рей, Луис В. (2007). Динозавры: самая полная, современная энциклопедия для любителей динозавров всех возрастов (1-е изд.). Нью -Йорк: Рэндом Хаус. ISBN 978-0-375-82419-7 .
- ^ Смит, Натан Д.; Маковицки, Питер Дж.; Хаммер, Уильям Р.; Керри, Филипп Дж. (Октябрь 2007 г.). «Остеология Cryolophosaurus ellioti (Dinosauria: Theropoda) из ранней юры Антарктиды и последствия для ранней эволюции теропод» . Зоологический журнал Линневого общества . 151 (2): 377–421. doi : 10.1111/j.1096-3642.2007.00325.x . ISSN 1096-3642 .
- ^ Занно, Линдси Э.; Джилетт, Дэвид Д.; Олбрайт, Л. Барри; Титус, Алан Л. (25 августа 2010 г.). «Новый североамериканский теризинозаврид и роль травоядных в« хищнической »эволюции динозавров» . Труды Королевского общества б . 276 (1672): 3505–3511. doi : 10.1098/rspb.2009.1029 . PMC 2817200 . PMID 19605396 .
- ^ Лонгрич, Николас Р.; Керри, Филипп Дж. (Февраль 2009 г.). « Albertonykus borealis , новый Alvarezsaur (Dinosauria: Theropoda) из раннего маастрихта Альберты, Канада: последствия для систематики и экологии Alvarezsauridae». Мерашные исследования . 30 (1): 239–252. BIBCODE : 2009CRRES..30..239L . doi : 10.1016/j.cretres.2008.07.005 .
- ^ Holtz, Tr Jr.; Бринкман, DL; Чендлер, Кл (1998). «Стоматологическая морфометрия и, возможно, всеядающая привычка кормления для динозавра Theropod Troodon ». Гайя . 15 : 159–166.
- ^ Jump up to: а беременный Hendrickx, C; Матеус, О (2014). «Abelisauridae (Dinosauria: Theropoda) из поздней юры филогения на основе зубного узора в качестве вклада для идентификации изолированных тероподных зубов». Zootaxa . 3759 : 1–74. doi : 10.11646/Zootaxa.3759.1.1 . PMID 24869965 . S2CID 12650231 .
- ^ Jump up to: а беременный Хендрикс, Кристоф; Матеус, Октавио; Araújo, Ricardo (2015). «Предложенная терминология зубов теропод (Dinosauria, Saurischia)» . Журнал палеонтологии позвоночных (представленная рукопись). 35 (E982797): E982797. Bibcode : 2015jvpal..35e2797h . doi : 10.1080/02724634.2015.982797 . S2CID 85774247 .
- ^ Geggel, Laura (28 июля 2015 г.). «Один жесткий укус: зубы Т. Рекса имели секретное оружие» . Fox News . Получено 1 августа 2015 года .
- ^ «Специальные зубцы дали плотоядным динозаврам эволюционное преимущество» . 17 августа 2015 года. Архивировано с оригинала 18 августа 2015 года.
- ^ Бринк, KS; Рейш, RR; Лебланк, Арх; Чанг, RS; Ли, YC; Чиан, CC; Huang, T.; Эванс, округ Колумбия (2015). «Развитие и эволюционная новинка в зубчатых зубах динозавров теропод» . Научные отчеты . 5 : 12338. Bibcode : 2015natsr ... 512338b . doi : 10.1038/srep12338 . PMC 4648475 . PMID 26216577 . S2CID 17247595 .
- ^ Бонапарт; Новас; Кория (1990). Carnotaurus sastrei ( Bonaparte ), рогатый, слегка построенный карнозавр из среднего мела Патагонии (отчет). Вклад в науку. Тол. 416. Округ Лос -Анджелес, Калифорния: Музей естественной истории.
- ^ Гелих, UB; Chiappe, LM (16 марта 2006 г.). «Новый хищный динозавр из покойного юрского архипелага» (PDF) . Природа . 440 (7082): 329–332. Bibcode : 2006natur.440..329G . doi : 10.1038/nature04579 . PMID 16541071 . S2CID 4427002 - через Doc.Rero.ch.
- ^ Jump up to: а беременный Черкас, SA; Юань, С. (2002). «Случайный манираптан с северо -восточного Китая» (PDF) . В Черкасе, SJ (ред.). Пернатые динозавры и происхождение полета . Журнал музея динозавров. Тол. 1. Бландинг, UT: Музей динозавров. С. 63–95.
- ^ Голич, UB; Tischlinger, H.; Chiappe, LM (2006). « Juraventaor Starki (Reptilia, Theropoda) хищные динозавры Nuer из верхней южной южной Франкэльб (Sueddeutschland): скелеттанатомия и выводы для частиц». Archeopteryx . 24 : 1–26.
- ^ Jump up to: а беременный Сюй, х.; Чжао, Q.; Norell, M.; Салливан, C.; Hone, D.; Эриксон, Г.; Ван, х.; Хан, Ф. и Го, Ю. (февраль 2009 г.). «Новая пернатая манирапторанская ископаемость динозавров, которая заполняет морфологический разрыв в птичьем происхождении» . Китайский научный бюллетень . 54 (3): 430–435. Bibcode : 2009scibu..54..430x . doi : 10.1007/s11434-009-0009-6 . Абстрактный
- ^ Каллен, Томас М.; Ларсон, Дерек Уильям; Виттон, Марк П.; Скотт, Дайан; Махо, чай; Бринк, Кирстин С.; и др. (30 марта 2023 г.). «Реконструкция лицевого лицера Theropod и важность мягких тканей в палеобиологии» . Наука . 379 (6639): 1348–1352. Bibcode : 2023sci ... 379.1348c . doi : 10.1126/science.abo7877 . PMID 36996202 . S2CID 257836765 .
- ^ Терриен, Ф.; Хендерсон, Д.М. (2007). «Мой теропод больше вашего ... или нет: оценка размера тела по длине черепа в тероподах». Журнал палеонтологии позвоночных . 27 (1): 108–115. doi : 10.1671/0272-4634 (2007) 27 [108: mtibty] 2.0.co; 2 . S2CID 86025320 .
- ^ Dal Sasso, C.; Maganuco, S.; Buffetaut, E.; Мендес, М.А. (2005). «Новая информация о черепе загадочного спинозавра Theropod , с замечаниями о его размерах и сродстве». Журнал палеонтологии позвоночных (представленная рукопись). 25 (4): 888–896. doi : 10.1671/0272-4634 (2005) 025 [0888: niotso] 2.0.co; 2 . S2CID 85702490 .
- ^ "Острич" . Африканский фонд дикой природы . Получено 28 октября 2020 года .
- ^ Brusatte SL, O'Connor JK, Jarvis ED (октябрь 2015). «Происхождение и диверсификация птиц» . Текущая биология . 25 (19): R888–98. BIBCODE : 2015CBIO ... 25.R888B . doi : 10.1016/j.cub.2015.08.003 . HDL : 10161/11144 . PMID 26439352 . S2CID 3099017 .
- ^ Хендри, Лиза (2023). "Птицы являются единственными выжившими динозаврами?" Полем Попечители Музея естественной истории, Лондон . Получено 14 апреля 2023 года .
- ^ Jump up to: а беременный Боренштейн, Сет (31 июля 2014 г.). «Исследование прослеживает эволюцию динозавров в ранних птиц» . AP News . Получено 3 августа 2014 года .
- ^ Jump up to: а беременный Зои Гоф (31 июля 2014 г.). «Динозавры сжимаются« регулярно, чтобы стать птицами » . Би -би -си .
- ^ Падиан, Кевин; De Ricqlès, Armand J.; Хорнер, Джон Р. (июль 2001 г.). «Динозаврские темпы роста и происхождение птиц» . Природа . 412 (6845): 405–408. Bibcode : 2001natur.412..405p . doi : 10.1038/35086500 . ISSN 1476-4687 . PMID 11473307 . S2CID 4335246 .
- ^ Jump up to: а беременный Андерсон, JF; Hall-Martin, A.; Рассел, Д.А. (1985). «Окружность и вес с длинными костями у млекопитающих, птиц и динозавров» . Журнал зоологии . 207 (1): 53–61. doi : 10.1111/j.1469-7998.1985.tb04915.x . ISSN 1469-7998 .
- ^ Jump up to: а беременный Campione, Nicolás E.; Эванс, Дэвид С. (2020). «Точность и точность оценки массы тела у неавийских динозавров» . Биологические обзоры . 95 (6): 1759–1797. doi : 10.1111/brv.12638 . ISSN 1469-185x . PMID 32869488 . S2CID 221404013 .
- ^ Ли, Эндрю Х.; Werning, Сара (15 января 2008 г.). «Сексуальная зрелость в растущих динозаврах не соответствует моделям роста рептилии» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (2): 582–587. doi : 10.1073/pnas.0708903105 . ISSN 1091-6490 . PMC 2206579 . PMID 18195356 .
- ^ Jump up to: а беременный Эриксон, Грегори М.; Роджерс, Кристина Карри; Еерби, Скотт А. (июль 2001 г.). «Модели роста динозавров и быстрые темпы роста птиц» . Природа . 412 (6845): 429–433. Bibcode : 2001natur.412..429e . doi : 10.1038/35086558 . ISSN 1476-4687 . PMID 11473315 . S2CID 4319534 . (Ошибка: doi : 10.1038/nature16488 , PMID 26675731 , реверкция Watch )
- ^ Хорнер, Джон Р.; Падиан, Кевин (22 сентября 2004 г.). «Динамика возраста и роста тиранозавра Рекса» . Труды Королевского общества . Б: Биологические науки. 271 (1551): 1875–1880. doi : 10.1098/rspb.2004.2829 . ISSN 0962-8452 . PMC 1691809 . PMID 15347508 .
- ^ Jump up to: а беременный в дюймовый и Хатчинсон, младший (март -апрель 2006 г.). PDF ( ) Рендус учетные записи Palevol 5 (3–4): 519–530. Bibcode : 2006crpal . два 10.1016/j.crpv.2005.09.002:
- ^ Jump up to: а беременный Ньюман, BH (1970). , едящем плоть «Стоя и походка в тиранозавре » . Биологический журнал Линневого общества . 2 (2): 119–123. doi : 10.1111/j.1095-8312.1970.tb01707.x .
- ^ Падиан, К.; Olsen, PE (1989). «Следы ратита, а также позиция и походка мезозойских теропод». В Гилетте, DD; Локли, мг (ред.). Треки и следы динозавров . Кембридж, Великобритания: издательство Кембриджского университета. С. 231–241.
- ^ Пол, Г.С. (1998). «Дизайн конечностей, функционирование и выполнение производительности в страусе-мимике и тиранозаврах». Гайя . 15 : 257–270.
- ^ Фарлоу, Джо; Гейтси, С.М.; Holtz, Tr Jr.; Хатчинсон, младший; Робинсон, JM (2000). «Теропод локомоция» . Являюсь. Zool . 40 (4): 640–663. doi : 10.1093/icb/40.4.640 .
- ^ «Аннотация», в Chure (2001). Пг 19 [ Полная цитата необходима ]
- ^ Баркер, Крис Тиджани; Найш, Даррен; Ньюхэм, Элис; Katsamenis, Oress L.; Дайк, Гарет (16 июня 2017 г.). «Сложная нейронаатомия в ростроме острова Уайт Теропод Невейтор Салерии » Научные отчеты 7 (1): 3749. Bibcode : 2017natsr ... 7.3749b Doi : 10.1038/ s41598-017-03671-3 ISSN 2045-2 5473926PMC 28623335PMID
- ^ Kawabe, Soichiro; Хаттори, Соки (3 июля 2022 года). «Сложная нейросулярная система в зубном состоянии Тиранозавра» . Историческая биология . 34 (7): 1137–1145. Bibcode : 2022hbio ... 34.1137K . doi : 10.1080/08912963.2021.1965137 . ISSN 0891-2963 . S2CID 238728702 .
- ^ Донг, З. (1984). «Новый динозавр Theropod из средней юры в бассейне Сычуань». Повертеблат Паласиатика . 22 (3): 213–218.
- ^ Jump up to: а беременный в дюймовый и фон Раухут, Оу (2003). Взаимосвязи и эволюция базальных динозавров . Blackwell Publishing. ISBN 0-901702-79-х .
- ^ Барта, де; Nesbitt, SJ; Norell, MA (2017). «Эволюция манусов ранних динозавров теропода характеризуется высокими и внутривидовыми вариациями» . Журнал анатомии . 232 (1): 80–104. doi : 10.1111/joa.12719 . PMC 5735062 . PMID 29114853 .
- ^ «Первый динозавр с одним пальцем» . Phys.org . Январь 2011.
- ^ Jump up to: а беременный Карпентер К. (2002). «Биомеханика передней основы неавийских динозавров в хищнике». Senckenbergiana Lethaea . 82 (1): 59–76. doi : 10.1007/bf03043773 . S2CID 84702973 .
- ^ Jump up to: а беременный Senter, P.; Робинс, JH (июль 2005 г.). «Диапазон движения в передней основе тероподного динозавра акрокантозавра атокенсиса и последствия для хищного поведения» . Журнал зоологии . 266 (3). Лондон, Великобритания: 307–318. doi : 10.1017/s0952836905006989 .
- ^ Molnar, Re (2001). «Теропод палеопатология: литературное обследование». В Танке, DH; Карпентер, К. (ред.). Мезозойская жизнь позвоночных . Издательство Университета Индианы. С. 337–363.
- ^ Jump up to: а беременный Xing, Ld; Локли, мг; Zhang, JP; и др. (2013). «Новая ранняя меловая трасса динозавров и первая определенная неавийская тероподная плавательная трасса из Китая» . Чин SCI Bull . 58 (19): 2370–2378. Bibcode : 2013Chsbu..58.2370x . doi : 10.1007/s11434-013-5802-6 .
- ^ Nesbitt, SJ; Смит, ND; Ирмис, РБ; Тернер, ах; Downs, A. & Norell, MA (11 декабря 2009 г.). «Полный скелет позднего триасового сауришяна и ранняя эволюция динозавров». Наука . 326 (5959): 1530–1533. Bibcode : 2009Sci ... 326.1530n . doi : 10.1126/science.1180350 . PMID 20007898 . S2CID 8349110 . Полем
- ^ Jump up to: а беременный Rowe, T. & Gauthier, J. (1990). "Ceratosauria". В Вайшампеле, дБ; Dodson, P. & Osmólska, H. (Eds.). Динозаврия . Беркли, Калифорния / Лос -Анджелес, Калифорния / Оксфорд, Великобритания: издательство Университета Калифорнии. С. 151–168.
- ^ Мортимер М. (4 июля 2001 г.). «Тезис Раухута» . dml.cmnh.org . Архив списка рассылки динозавров.
- ^ Каррано, MT; Сэмпсон, SD; Forster, CA (2002). «Остеология масиакасаура Кнопфлери , небольшого авелизавра (Dinosauria: Theropoda) от позднего мела Мадагаскара». Журнал палеонтологии позвоночных . 22 (3): 510–534. doi : 10.1671/0272-4634 (2002) 022 [0510: toomka] 2.0.co; 2 . S2CID 85655323 .
- ^ Bakker, RT (1986). Ереси динозавра (PDF) . Нью -Йорк, Нью -Йорк: Уильям Морроу - через Doc.Rero.ch.
- ^ Серено П. (1998). «Рациональный для филогенетических определений, с применением к таксономии динозаврии более высокого уровня». Трактаты. Новый ежегодник для геологии и палеонтологии (ежегодный). 210 : 41–83. Doi : 10.1127/njgpa/210/998/41 .
- ^ Марш, Адам Д.; Роу, Тимоти Б. (2020). «Комплексная анатомическая и филогенетическая оценка Dilophosaurus wetherilli (Dinosauria, Theropoda) с описаниями новых образцов из формирования кайенты северной Аризоны» . Журнал палеонтологии . 94 (S78): 1–103. Bibcode : 2020jpal ... 94S ... 1M . doi : 10.1017/jpa.2020.14 . S2CID 220601744 .
- ^ Сассо, Криштиану Дал; Магануко, Симона; Кау, Андреа (19 декабря 2018 г.). «Самый старый цератозавр (Dinosauria: Theropoda), из нижней юры в Италии, проливает свет на эволюцию трехкоренной руки птиц» . ПЕРЕЙ . 6 : E5976. doi : 10.7717/peerj.5976 . ISSN 2167-8359 . PMC 6304160 . PMID 30588396 .
- ^ Ostrom, JH (1969). «Остеология Deinonychus antirrhopus , необычный теропод из нижнего мела Монтаны». Бюллетень музея Пибоди естественной истории . 30 : 1–165.
- ^ Пол, Г.С. (1988). Хищные динозавры мира . Нью -Йорк, Нью -Йорк: Simon and Schuster Co. ISBN 0-671-61946-2 .
- ^ Dingus, L.; Роу Т. (1998). Ошибочное вымирание: эволюция динозавра и происхождение птиц . Фриман.
- ^ Пол, Г.С. (2002). Динозавры воздуха: эволюция и потеря полета у динозавров и птиц . Балтимор, доктор медицинских наук: издательство Университета Джона Хопкинса. ISBN 0-8018-6763-0 .
- ^ Lee, Michaels.Y.; Кау, Андреа; Найш, Даррен; Дайк, Гарет Дж. (1 августа 2014 г.). «Постоянная миниатюризация и анатомические инновации в динозаврских предках птиц». Наука . 345 (6196): 562–566. Bibcode : 2014sci ... 345..562L . doi : 10.1126/science.1252243 . PMID 25082702 . S2CID 37866029 .
- ^ Марш, OC (1881). «Основные персонажи американской юрской динозавров. Часть V.». Американский журнал науки и искусства . 3. 21 (125): 417–423. Bibcode : 1881AMJS ... 21..417M . doi : 10.2475/ajs.s3-21.125.417 . S2CID 219234316 .
- ^ Мэтью, WD ; Браун, Б. (1922). «Семейство Deinodontidae, с уведомлением о новом роде из мела Альберты». Бюллетень Американского музея естественной истории . 46 : 367–385.
- ^ Андерсон, Тед Р. (2006). Биология повсеместного дома воробья: от генов до популяций. Оксфорд: издательство Оксфордского университета. ISBN 0-19-530411-X .
- ^ Hendrickx, C.; Хартман, SA; Матеус, О. (2015). «Обзор неавийских обнаружений и классификации». Паларх журнал о палеонтологии позвоночных . 12 (1): 1–73.
- ^ Карр, Томас (22 октября 2013 г.). «Остеология VIII: анторбитальная полость» . Tyrannosauroidea Central . Получено 2 апреля 2024 года .
- ^ Пол, Г.С. (2002). Динозавры воздуха . Балтимор, MD / Лондон, Великобритания: издательство Университета Джона Хопкинса. ISBN 978-0-8018-6763-7 - через Google Books.
- ^ Ezcurra, MD; Cuny, G. (2007). «Coelophysoid Lophostropheus airelensis , Gen. Nov.: Обзор систематики « Liliensernus »Airelensis из триасовых выходов в Нормандию (Франция)» . Журнал палеонтологии позвоночных . 27 (1): 73–86. doi : 10.1671/0272-4634 (2007) 27 [73: tclagn] 2.0.co; 2 -через Researchgate.net.
- ^ Мортимер, Микки (15 ноября 2010 г.). «Аверостра и Авепода» . Блог базы данных Theropod . Получено 6 сентября 2020 года .
- ^ Барон, мг; Норман, дБ; Барретт, PM (2017). «Новая гипотеза о отношениях динозавров и ранней эволюции динозавров» (PDF) . Природа . 543 (7646): 501–506. Bibcode : 2017natur.543..501b . doi : 10.1038/nature21700 . PMID 28332513 . S2CID 205254710 .
- ^ «Новое исследование встряхивает корни семейного древа динозавров» . cam.ac.uk. Кембридж, Великобритания: Кембриджский университет . 22 марта 2017 года.
- ^ Лангер, MC; Ezcurra, MD; Раухут, Оум; Бентон, MJ; Knoll, F.; Макфи, BW; и др. (2017). «Распутывание семейного древа динозавров» . Природа . 551 (7678): E1 - E3. Bibcode : 2017natur.551e ... 1L . doi : 10.1038/nature24011 . HDL : 1983/D088DAE2-C7FA-4D41-9FA2-AEEBBFCD2FA3 . PMID 29094688 . Архивировано из оригинала 9 июля 2023 года . Получено 8 июля 2023 года .
Внешние ссылки
[ редактировать ]

















| Базы данных управления авторитетом : национальный |
|---|