Протоцератопс
| Протоцератопс Временной диапазон: поздний мел , ( кампан ) ~
| |
|---|---|
| |
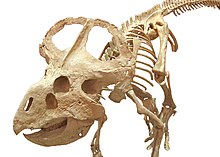
| Установленный скелет P. andrewsi , Центр динозавров Вайоминга. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | † Орнитишия |
| Клэйд : | † Неорнитишия |
| Клэйд : | † Цератопсия |
| Клэйд : | † Коронозаврия |
| Семья: | † Протоцератопсиды |
| Род: | † Протоцератопс Грейнджер и Грегори , 1923 год. |
| Типовой вид | |
| † Протоцератопс Эндрюси Грейнджер и Грегори, 1923 год.
| |
| Другие виды | |
| |
Протоцератопс ( / ˌ p r oʊ t oʊ ˈ s ɛr ə t ɒ p s / ; букв. « первое рогатое лицо » ) [ 1 ] — род небольших протоцератопсидных динозавров , живших в Азии в позднем меловом периоде , около 75–71 миллиона лет назад. Род Protoceratops включает два вида: P. andrewsi и более крупный P. hellenikorhinus . Первый был описан в 1923 году на окаменелостях из монгольской формации Джадохта , а второй — в 2001 году на окаменелостях из китайской формации Баян Мандаху . протоцератопс Первоначально считалось, что является предком анкилозавров и более крупных цератопсов, таких как трицератопс и его родственники, до открытия других протоцератопсид. Популяции P. andrewsi , возможно, превратились в Bagaceratops в результате анагенеза .
Протоцератопсы были маленькими цератопсами, длиной до 2–2,5 м (6,6–8,2 футов) и массой тела около 62–104 кг (137–229 фунтов). В то время как взрослые особи были в основном четвероногими , подростки могли при необходимости передвигаться на двух ногах . Они характеризовались пропорционально большим черепом , короткой и жесткой шеей и воротником на шее . Оборка, вероятно, использовалась для демонстрации или внутривидового боя , а также для защиты шеи и фиксации мышц челюсти. Над носом присутствовала роговидная структура, которая варьировалась от одиночной структуры у P. andrewsi до двойной парной структуры у P. hellenikorhinus . нет «Рог» и оборка сильно различались по форме и размеру у особей одного и того же вида, но признаков полового диморфизма . У них был выдающийся клюв, похожий на попугайский , на кончике челюсти. У P. andrewsi была пара цилиндрических тупых зубов на кончике верхней челюсти. На передних конечностях было пять пальцев , из которых только первые три имели широкие и плоские когти . Ступни для были широкими, с четырьмя пальцами и уплощенными лопатообразными когтями, которые пригодились бы копаясь в песке. Задние конечности были длиннее передних. Хвост был длинным и имел загадочную структуру, похожую на парус , которая, возможно, использовалась для демонстрации, плавания или метаболических целей.
Протоцератопсы , как и многие другие цератопсы, были травоядными животными, оснащенными выступающими челюстями и зубами, подходящими для измельчения листвы и другого растительного материала. Считается, что они жили очень общительными группами разного возраста. Похоже, они заботились о своих детенышах . с мягкой скорлупой Они откладывали яйца , что является редкостью для динозавров. Во время взросления череп и шейный воротник быстро росли. На протоцератопсов охотился велоцираптор , и один особенно известный экземпляр (« Боевые динозавры ») сохранил пару из них, сражавшихся в бою. Раньше протоцератопсов считали ночными из-за большого склеротического кольца вокруг глаза, но теперь считается, что они вели катемерный образ жизни (активны на рассвете и в сумерках).
История открытия
[ редактировать ]
В 1900 году Генри Фэйрфилд Осборн предположил, что Центральная Азия могла быть центром происхождения большинства видов животных, включая человека , что привлекло внимание исследователя и зоолога Роя Чепмена Эндрюса . Эта идея позже породила Первую (1916–1917), Вторую (1919) и Третью (1921–1930) среднеазиатские экспедиции в Китай и Монголию , организованные Американским музеем естественной истории под руководством Осборна и полевого руководства Эндрюс. Команда третьей экспедиции прибыла в Пекин в 1921 году для последних приготовлений и приступила к полевым работам в 1922 году. В конце 1922 года экспедиция исследовала знаменитые Пылающие скалы района Шабарах-Усу формации Джадохта , пустыни Гоби , ныне известной как Район Байн-Дзак. 2 сентября фотограф Джеймс Б. Шакелфорд частичный череп молодого человека, который впоследствии стал образцом голотипа (AMNH 6251) протоцератопса обнаружил в красноватых песчаниках . Впоследствии он был проанализирован палеонтологом. Уолтер В. Грейнджер, который определил его как рептилию . 21 сентября экспедиция вернулась в Пекин, и хотя она была создана для поиска останков человеческих предков, команда собрала множество динозавров окаменелостей и, таким образом, дала представление о богатой летописи окаменелостей Азии. Вернувшись в Пекин, череп, найденный Шакелфордом, был отправлен в Американский музей естественной истории для дальнейшего изучения, после чего Осборн связался с Эндрюсом и командой по телеграмме, уведомив их о важности экземпляра. [ 2 ] [ 3 ]
В 1923 году экспедиция снова исследовала Пылающие скалы, на этот раз обнаружив еще больше экземпляров протоцератопса , а также первые останки овираптора , заурорнитоидеса и велоцираптора . В частности, команда обнаружила первые окаменелые яйца динозавров рядом с голотипом Овираптора, и, учитывая, насколько многочисленны были протоцератопсы , гнездо было отнесено к этому таксону . [ 3 ] Позже это привело к интерпретации Овираптора как похитителя яиц . [ 4 ] В том же году Грейнджер и Уильям К. Грегори официально описали новый род и вид Protoceratops andrewsi на основе голотипа черепа. Видовое имя , andrewsi , дано в честь Эндрюса за его выдающееся руководство во время экспедиций. Они идентифицировали протоцератопса как птицетазового динозавра, тесно связанного с цератопсами, представляющего возможного общего предка анкилозавров и цератопсов . Поскольку протоцератопс был более примитивным, чем любой другой известный в то время цератопс, Грейнджер и Грегори создали новое семейство Protoceratopsidae , в основном характеризующееся отсутствием рогов. Соавторы также согласились с Осборном в том, что Азия, если ее более тщательно исследовать, сможет устранить многие серьезные эволюционные пробелы в летописи окаменелостей. [ 2 ] Хотя это и не указано в оригинальном описании, родовое название Протоцератопс представляет означает «первое рогатое лицо», поскольку считалось, что Протоцератопс собой раннего предка цератопсид . [ 5 ] Другие исследователи сразу же отметили важность находок протоцератопсов , и этот род был провозглашен «долгожданным предком трицератопса ». Большинство окаменелостей находились в отличной сохранности, у некоторых экземпляров сохранились даже склеротические кольца (тонкие глазные кости), что быстро сделало протоцератопса одним из самых известных динозавров Азии. [ 3 ] [ 6 ]

Проведя большую часть 1924 года за планированием следующих сезонов полевых работ, в 1925 году Эндрюс и его команда снова исследовали Пылающие скалы. В этом году было собрано больше яиц и гнезд, а также хорошо сохранившиеся и полные экземпляры протоцератопсов . К этому времени протоцератопс стал одним из самых многочисленных динозавров в регионе: известно более 100 экземпляров, включая черепа и скелеты нескольких особей на разных стадиях роста. больше останков протоцератопса , наибольшая их численность наблюдалась в сезоны 1922–1925 годов. Хотя в последующие годы экспедиций было собрано [ 3 ] [ 6 ] Грегори и Чарльз К. Мук опубликовали еще одно описание протоцератопса в 1925 году, обсуждая его анатомию и взаимоотношения. Благодаря большой коллекции черепов, найденных в экспедициях, они пришли к выводу, что протоцератопс представлял собой цератопса, более примитивного, чем цератопсиды, а не предка анкилозавра-цератопса. [ 7 ] В 1940 году Барнум Браун и Эрих Марен Шлайкьер описали анатомию P. andrewsi , используя недавно подготовленные экземпляры из азиатских экспедиций. подробно [ 6 ]
В 1963 году монгольский палеонтолог Демберэлин Дашзевег сообщил об открытии нового ископаемого местонахождения джадохтинской свиты: Тугрикен Шире. Как и соседний Байн-Дзак, это новое местонахождение содержало множество окаменелостей протоцератопсов . [ 8 ] В течение 1960-1970 х - - монгольские годов польско и российско -монгольские палеонтологические экспедиции собрали в этом месте новые, от частичных до полных экземпляров протоцератопса , что сделало этот вид динозавров обычным явлением в Тугрикен Шире. [ 9 ] [ 10 ] [ 11 ] С момента своего открытия в местности Тугрикен Шире были обнаружены некоторые из наиболее значительных экземпляров протоцератопсов , таких как боевые динозавры , [ 9 ] особи in situ - условие сохранения, в некоторых случаях также известное как «стоящие» особи или образцы -, [ 12 ] настоящие гнезда, [ 13 ] и небольшие стадные группы. [ 14 ] Экземпляры из этого местонахождения обычно встречаются в сочленении, что позволяет предположить возможную массовую гибель. [ 12 ]
Стефан Н. Ф. Спикман и его коллеги сообщили о частичном черепе P. andrewsi (RGM 818207) в коллекциях Центра биоразнообразия Naturalis , Нидерланды , в 2015 году. Поскольку окаменелости протоцератопса встречаются только в пустыне Гоби в Монголии, и этот образец, вероятно, был обнаружен во время Центральной Команда Asiatic Expeditions пришла к выводу, что этот череп, вероятно, был приобретен Делфтским университетом между 1940 и 1972 годами в рамках передачи коллекции. [ 15 ]
Виды и синонимы
[ редактировать ]
Останки протоцератопсид были обнаружены в 1970-х годах в местонахождении Хулсан формации Барун-Гойот в Монголии в ходе работы нескольких польско-монгольских палеонтологических экспедиций. В 1975 году польские палеонтологи Тереза Марьянска и Гальшка Осмольска описали второй вид протоцератопсов , который они назвали P. kozlowski . Этот новый вид был основан на материале Хулсана, состоящем в основном из молодых образцов черепов. Видовое название « Козловский » дано в честь польского палеонтолога Романа Козловского . Они также дали название новому роду и виду протоцератопсида Bagaceratops rozhdestvenskyi , известному по экземплярам из близлежащего местонахождения Гермиин Цав. [ 10 ] В 1990 году российский палеонтолог Сергей Михайлович Курзанов дополнительный материал Гермиина Цава передал П. Козловскому . имеется достаточно различий Однако он отметил, что между P. andrewsi и P. kozlowski , и выделил новый род и комбинацию Breviceratops kozlowskie . [ 16 ] Хотя Бревицератопс считается синонимом ювенильной стадии Багацератопса . [ 17 ] [ 18 ] Лукаш Чепинский в 2019 году пришел к выводу, что первый имеет достаточно анатомических различий, чтобы его можно было рассматривать как отдельный таксон . [ 19 ]
В 2001 году Оливер Ламберт с коллегами назвал новый и особый вид протоцератопса — P. hellenikorhinus . Первые известные останки P. hellenikorhinus были собраны в местонахождении Баян-Мандаху формации Баян-Мандаху , Внутренняя Монголия , в 1995 и 1996 годах во время китайско - бельгийских палеонтологических экспедиций. Экземпляры голотипа (ИММ 95ВМ1/1) и паратипа (ИММ 96ВМ1/4) состоят из крупных черепов без остатков тела. Череп голотипа был обнаружен обращенным вверх — поза, о которой сообщалось у экземпляров протоцератопса из Тугрикен Шире. Видовое название hellenikorhinus происходит от греческих слов hellenikos (что означает греческий) и rhis (что означает нос) в связи с его широкой и угловатой мордой, которая напоминает прямые профили греческих скульптур . [ 20 ] В 2017 году обильное количество протоцератопсид было обнаружено в Альхе возле Баян-Мандаху. [ 21 ] и он может быть предпочтительнее P. hellenikorhinus . [ 19 ]
Виктор Терещенко и Владимир Р. Алифанов в 2003 году назвали нового протоцератопсидного динозавра из местонахождения Байн-Дзак — Bainoceratops efremovi . Этот род был основан на нескольких спинных позвонках, которые, как утверждалось, отличались от позвонков протоцератопса . [ 22 ] В 2006 году североамериканские палеонтологи Питер Маковицки и Марк А. Норелл предположили, что байноцератопс может быть синонимом протоцератопса, поскольку большинство признаков, используемых для отделения первого от второго, были зарегистрированы у других цератопсов, включая самого протоцератопса , и они с большей вероятностью выпадут. в пределах широкого диапазона внутривидовой изменчивости конкурирующего P. andrewsi . [ 23 ] Авторы Бренда Дж. Чиннери и Джон Р. Хорнер в 2007 году во время описания Cerasinops заявили, что Bainoceratops , наряду с другими сомнительными родами, был определен как вариант или незрелый экземпляр других родов. На основании этого они исключили Bainoceratops из своего филогенетического анализа. [ 24 ]
Яйца и гнезда
[ редактировать ]
В рамках Третьей центральноазиатской экспедиции 1923 года Эндрюс и его команда обнаружили голотип овираптора вместе с некоторыми из первых известных окаменевших яиц динозавра (гнездо AMNH 6508) в формации Джадохта. Каждое яйцо было удлиненным и имело твердую скорлупу, а из-за близости и большого количества протоцератопсов в формации в то время считалось, что эти яйца принадлежали этому динозавру. Это привело к интерпретации современного овираптора как животного-хищника, что также отражено в его родовом названии. [ 25 ] [ 4 ] В 1975 году китайский палеонтолог Чжао Цзикуй назвал новые оогенеры Elongatoolithus и Macroolithus , включив их в новое семейство : Elongatoolithidae . Как следует из названия, они представляют собой удлиненные яйца динозавров, в том числе некоторые из которых относят к протоцератопсам . [ 26 ]
В 1994 году российский палеонтолог Константин Евгеньевич Михайлов назвал новый ооген Protoceratopsidovum из формаций Барун-Гойот и Джадохта с типовым видом P. sinrum и дополнительными P. fluxuosum и P.minimum . Этот оотаксон был твердо установлен как принадлежащий протоцератопсидным динозаврам, поскольку они были преобладающими динозаврами, у которых были обнаружены яйца, а некоторые скелеты протоцератопсов были обнаружены в непосредственной близости от яиц Protoceratopsidovum . Точнее, Михайлов заявил, что P.iserum и P.minimum были заложены протоцератопсом , а P.fluxuosum — бревицератопсом . [ 27 ]

Однако также в 1994 году Норелл и его коллеги сообщили и кратко описали окаменелый теропода эмбрион внутри яйца (MPC-D 100/971) из джадохтинской свиты. Они идентифицировали этот эмбрион как динозавра -овираптора , и при внимательном рассмотрении яичная скорлупа оказалась скорлупой яиц элонгатоолитид, и, таким образом, был сделан вывод, что о-семейство Elongatoolithidae представляет яйца овирапторидов. Эта находка доказала, что гнездо AMNH 6508 принадлежало Овираптору , а не похитителю яиц, а голотипом на самом деле была зрелая особь, которая погибла, высиживая яйца. [ 28 ] Более того, филогенетический анализ, опубликованный в 2008 году Дарлой К. Зеленицкой и Франсуа Террьеном, показал, что Protoceratopsidovum представляет собой яйца манираптора , более производные, чем овирапториды, а не протоцератопсы . [ 29 ] Описание яичной скорлупы Protoceratopsidovum дополнительно подтвердило, что они на самом деле принадлежат к манирапторану, возможно, таксону дейнонихозавров . [ 30 ]
подлинное гнездо протоцератопса Тем не менее, в 2011 году Дэвид Э. Фастовский и его коллеги сообщили и описали . Гнездо (MPC-D 100/530) с 15 членистоногими молодью было собрано в местонахождении Тугрикен Шире джадохтинской свиты в ходе работы монголо- японских палеонтологических экспедиций. [ 13 ] Грегори М. Эриксон и его команда в 2017 году сообщили о кладке яиц с эмбрионами (MPC-D 100/1021) протоцератопсов из также ископаемого местонахождения Ухаа Толгод, обнаруженной во время палеонтологических экспедиций Американского музея естественной истории и Монгольской академии наук . Эта кладка состоит как минимум из 12 яиц и эмбрионов, и только у 6 эмбрионов сохранились почти полные скелеты. [ 31 ] Норелл с коллегами в 2020 году исследовали окаменелые останки вокруг яиц этой кладки, что указывает на их мягкую скорлупу. [ 32 ]
Борьба с динозаврами
[ редактировать ]
Образец «Боевые динозавры» сохраняет протоцератопса (MPC-D 100/512) и велоцираптора (MPC-D 100/25), окаменевших в бою, и дает важное окно в отношении прямых доказательств поведения хищник-жертва у нептичьих динозавров. [ 9 ] [ 33 ] В 1960-х и начале 1970-х годов в пустыню Гоби было проведено множество польско-монгольских палеонтологических экспедиций с целью обнаружения окаменелостей. В 1971 году экспедиция исследовала несколько участков джадохтинской и немегтинской свит. В ходе полевых работ 3 августа в местонахождении Тугрикен Шир (формация Джадохта) было обнаружено несколько окаменелостей протоцератопса и велоцираптора , включая блок, содержащий по одному экземпляру каждого из них. Особи в этом блоке были идентифицированы как P. andrewsi и V. mongoliensis . Хотя условия их захоронения до конца не изучены, было ясно, что они погибли одновременно в борьбе. [ 9 ]
Экземпляр, получивший прозвище «Боевые динозавры», был исследован и изучен многочисленными исследователями и палеонтологами, и существуют различные мнения о том, как животные были захоронены и вообще сохранены. Хотя Барсболд предложил сценарий утопления, [ 33 ] такая гипотеза считается маловероятной, учитывая аридную палеообстановку джадохтинской свиты. Обычно считается, что они были заживо погребены песчаной бурей или обрушившейся дюной . [ 34 ] [ 35 ] [ 36 ]
Отпечатки кожи и следы
[ редактировать ]
Во время Третьей Центрально-Азиатской экспедиции в 1923 году протоцератопса в Пылающих скалах был собран почти полный скелет (экземпляр AMNH 6418). В отличие от других экземпляров, он был обнаружен в свернутом положении, а на его черепе сохранился тонкий, твердый и морщинистый слой матрицы (окружающие отложения ). Этот образец был позже описан в 1940 году Брауном и Шлайкьером, которые обсуждали природу части матрицы. Они заявили, что этот слой имел текстуру, очень похожую на кожу , и покрывал в основном левую сторону черепа от морды до воротника на шее . Браун и Шлайкьер отвергли идею возможных отпечатков кожи, поскольку этот похожий на кожу слой, вероятно, был продуктом разложения и захоронения человека, в результате чего отложения сильно прикреплялись к черепу. [ 6 ]
Потенциальная важность этих останков не была признана и не учтена, и к 2020 году образец уже был полностью подготовлен, потеряв все следы этого похожего на кожу слоя. Некоторые элементы были повреждены в процессе, например, трибуна . [ 37 ] В 2022 году Фил Р. Белл и его коллеги кратко описали эти потенциальные мягкие ткани на основе фотографий, предоставленных Брауном и Шлайкьером, а также других мягких тканей цератопсов. [ 38 ] Однако, хотя первоначально предполагалось, что весь похожий на кожу слой был удален, фотографии, которыми поделился Чепинский в том же году, показали, что правая сторона черепа осталась нетронутой, сохранив большую часть этого слоя и ожидая дальнейшего анализа. [ 37 ]
Также в рамках польско-монгольских палеонтологических экспедиций в 1965 г. полувзрослого протоцератопса из местонахождения Байн-Дзак джадохтинской свиты был собран сочлененный скелет (экз. ZPAL Mg D-II/3). В 2000-х годах во время подготовки образца . под тазовым поясом был обнаружен окаменелый слепок четырехпалого пальцевого следа Этот след был описан в 2012 году Гжегожем Недзведцким и его коллегами, которые считали его одной из первых зарегистрированных находок следа динозавра в сочетании с сочлененным скелетом, а также первой обнаруженной находкой протоцератопса . [ 39 ] Элементы конечностей скелета ZPAL Mg D-II/3 были описаны в 2019 году палеонтологами Юстиной Словяк, Виктором Терещенко и Люцией Фостович-Фрелик. [ 40 ] Терещенко в 2021 г. полностью описал осевой скелет этого экземпляра. [ 41 ]
Описание
[ редактировать ]
Протоцератопс относительно небольшого размера был цератопсом , длина P. andrewsi и P. hellenikorhinus оценивалась в 2–2,5 м (6,6–8,2 фута). [ 42 ] [ 43 ] и масса тела около 62–104 кг (137–229 фунтов). [ 44 ] Несмотря на схожий общий размер тела, последний имел относительно большую длину черепа. [ 20 ] Оба вида можно отличить по следующим признакам:
- P. andrewsi – на предчелюстной кости имелось два зуба; морда была низкая и длинная; носовой рог представлял собой единую заостренную структуру; нижний край зубной кости слегка изогнут. [ 6 ] [ 20 ]
- P. hellenikorhinus – отсутствие предчелюстных зубов; морда была высокая и широкая; носовой рог был разделен на два заостренных гребня; нижний край зубной кости был прямым. [ 20 ]
Череп
[ редактировать ]
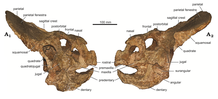
Череп был относительно большим по протоцератопса . сравнению с его телом и крепко сложен Череп типового вида P. andrewsi имел среднюю общую длину около 50 см (500 мм). С другой стороны, P. hellenikorhinus общая длина черепа составляла около 70 см (700 мм). Задняя часть черепа имела форму выраженного шейного оборка (также известного как «теменная оборка»), состоящего в основном из теменных и чешуйчатых костей. Точный размер и форма оборки варьировались в зависимости от человека; у некоторых были короткие компактные оборки, а у других — почти до половины длины черепа. Чешуйчатая кость касалась скуловой кости , была очень увеличенной и высокой, с изогнутым концом, образующим границы оборки. Теменные кости представляли собой самые задние кости черепа и основные элементы оборки. На виде сверху они имели треугольную форму и соединялись лобными костями (костями крыши черепа ). Обе теменные кости были сращены (слиты), образуя длинный гребень в центре оборки. Скуловая кость была глубокой и резко развитой, наряду с квадратно-скуловые они образовывали роговидное расширение, направленное вниз на боковых сторонах черепа. Эпижугалия (конечная область скуловой кости) была отделена от скуловой кости выступающим швом ; этот шов был более заметен у взрослых. Поверхности вокруг надсудной кости были шероховатыми, что указывало на то, что она была покрыта роговой оболочкой. В отличие от многих производных цератопсид , лобные и заглазничные кости протоцератопсов были плоскими и не имели роговых ядер или надглазничных рогов. ( Пальпебральная кость маленькая шпорообразная кость) присоединялась к префронтальной кости над передней частью глазницы ( глазницы). У P. hellenikorhinus глазные кости выступали вверх из префронтальной кости , чуть выше глазницы и слегка соприкасались с лобной, образуя небольшую роговидную структуру. кость Слёзная представляла собой кость почти прямоугольной формы, расположенную перед глазницей, что способствовало форме последней. Склеротическое кольцо (структура, поддерживающая глазное яблоко ), обнаруженное внутри орбиты, имело круглую форму и образовано последовательными костными пластинками. [ 6 ] [ 20 ]
Рыло . образовано носовой , верхнечелюстной , предчелюстной и ростральной костями Нос, как правило, был закругленным, но у некоторых особей был острый носовой выступ (особенность, получившая название «носовой рог»). У P. hellenikorhinus этот выступ был разделен на два острых и длинных гребня. Верхняя челюсть была очень глубокой и имела до 15 альвеол ( зубных лунок) на нижней стороне или опорной поверхности зубов. Предчелюстная кость имела две альвеолы на нижнем крае — признак, который присутствовал, по крайней мере, у P. andrewsi . Ростральная кость была лишена зубов, высокая, треугольной формы. У него был острый конец и шероховатая текстура, что указывает на рамфотеки (рогового клюва наличие ). В целом череп имел четыре пары фенестров (отверстий в черепе). Самое переднее отверстие, ноздри (ноздри), имело овальную форму и было значительно меньше, чем ноздри, наблюдаемые у цератопсид. У протоцератопсов были большие орбиты диаметром около 5 см (50 мм) и неправильной формы в зависимости от человека. Обращенные вперед и близко расположенные глазницы в сочетании с узкой мордой давали У протоцератопса хорошо развито бинокулярное зрение . Позади глаза было окно немного меньшего размера, известное как подвисочное окно , образованное изгибами скуловой и чешуйчатой кости. Последними отверстиями черепа были два теменных окна (отверстия в оборке). [ 6 ] [ 20 ]


Нижняя челюсть протоцератопса представляла собой крупный элемент, состоящий из предзубной , зубной , венечной , угловой и надугольной частей . Преддентарная (передняя кость) была очень заостренной и удлиненной, с V-образной симфизарной областью (сращением костей) спереди. Зубная кость (кость, несущая зубы) была прочной, глубокой, слегка изогнутой и сросшейся с угловой и надугловой. По латеральной поверхности зубной кости проходил большой и толстый гребень, соединявший венечный отросток — костный выступ, отходящий вверх от верхней поверхности нижней челюсти за зубным рядом, — и надугольный. На верхнем крае имело до 12-14 альвеол. И преддентарий, и дентарий имели ряд отверстий (небольших ямок), причем последние преимущественно на переднем конце. Венечный отросток (самая высокая точка нижней челюсти) имел тупую форму и касался венечного отростка зубной кости и был скрыт скуловой костью. Надугольный отросток имел форму, близкую к треугольной, и у старых особей он сросся с венечным отростком. Угловая часть располагалась ниже двух последних костей и позади зубной кости. Это была большая и несколько округлая кость, дополнявшая кривизну зубной кости. На своей внутренней поверхности он крепился к суставной . Суставная кость представляла собой меньшую по размеру кость и имела на внутренней поверхности вогнутость для сочленения с квадратной костью. [ 6 ] [ 20 ]
У протоцератопсов были зубные и верхнечелюстные зубы листовидной формы с несколькими зубчиками (зубцами) на соответствующих краях. Коронки (верхняя открытая часть) имели две грани или доли , которые были разделены центральной гребнеобразной структурой (также называемой «первичным гребнем»). Зубья были упакованы в один ряд, что создавало режущую поверхность. Как зубные, так и верхнечелюстные зубы имели выраженную гомодонтию — состояние зубов, при котором зубы имеют одинаковую форму и размер. P. andrewsi имел два маленьких зуба, похожих на шипы, которые располагались на нижней стороне каждой предчелюстной кости. Второй предчелюстной зуб был крупнее первого. В отличие от зубных и верхнечелюстных зубов предчелюстной зубной ряд лишен зубчиков и имеет относительно гладкую поверхность. Все зубы имели один корень (нижняя часть вросла в альвеолы). [ 6 ] [ 45 ] [ 46 ]
Посткраниальный скелет
[ редактировать ]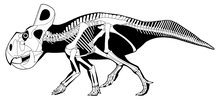
В позвоночнике протоцератопса . было девять шейных (шея), 12 спинных (спина), восемь крестцовых (тазовых) и более 40 хвостовых (хвостовых) позвонков Центры ( центр ; тела позвонков) первых трех шейных позвонков были объединены вместе ( атлант , ось и третий шейный отдел соответственно), создавая жесткую структуру. Шея была довольно короткой и имела плохую гибкость. Атлант был самым маленьким шейным отделом и состоял в основном из центра, поскольку нервная дуга (верхняя и заостренная область позвонка) представляла собой тонкий, узкий стержень кости, который простирался вверх и назад до основания оси нервного отростка . Капитулярная фасетка (место прикрепления шевронов ; также известная как шейные ребра) образована низким выступом, расположенным вблизи основания нервной дуги. Передняя фасетка центрального атласа была сильно вогнутой и обеспечивала сочленение затылочного мыщелка черепа. Нервная дуга и ость оси были заметно больше, чем сам атлас и любая другая шейная часть. Осевой нервный отросток был широким, развитым назад и слегка соединенным с отростком третьего шейного отдела. С четвертого по девятый все шейки матки были относительно равны по размеру и пропорциям. Их нервные отростки были меньше первых трех позвонков, а развитие головчатой фасетки уменьшалось, начиная с четвертого шейного отдела. [ 6 ] [ 47 ] [ 48 ]








Спинные позвонки были схожи по форме и размеру. Их нервные отростки имели удлиненную субпрямоугольную форму с тенденцией к более удлинению в задних позвонках. Центры были большими и преимущественно амфиплатными (плоскими с обеих сторон) и круглыми, если смотреть спереди. Иногда у старых особей последний спинной позвонок несколько сращен с первым крестцовым. Крестцовые позвонки прочно срослись, придав форму крестцу, который соединялся с внутренними сторонами обеих подвздошных костей. Их нервные отростки были широкими, не сросшимися и довольно одинаковой длины. Центры преимущественно опистоцельные (вогнутые на задней фасетке и выпуклые на передней), их размеры к концу уменьшаются. Хвостовые позвонки постепенно уменьшались в размерах к концу и имели очень удлиненные нервные отростки в середине ряда, образуя парусоподобную структуру. Это удлинение началось с первого по четырнадцатый хвостовой отдел. Центры гетероцельные (седловидные с обеих сторон). На передних хвостовых отделах они были широкими, однако начиная с двадцать пятого центра центры удлинялись вдоль нервных отростков. На нижней стороне хвостовых позвонков был прикреплен ряд шевронов, придающих форму нижней части хвоста. Первый шеврон располагался на месте соединения третьего и четвертого каудальных элементов. Шевроны с третьего по девятый были самыми большими, а начиная с десятого они становились меньше. [ 6 ] [ 47 ] [ 48 ] [ 49 ]
Все позвонки протоцератопса имели ребра, прикрепленные на боковых сторонах, за исключением ряда хвостовых. Первые пять шейных ребер (иногда называемые шевронами) были одними из самых коротких ребер, причем среди них первые два были длиннее остальных. С третьего по шестое спинные (грудные) ребра были самыми длинными ребрами в скелете протоцератопса , последующие ребра становились меньше по размеру по мере продвижения к концу позвоночного столба. Два последних спинных ребра были самыми маленькими, причем последнее из них соприкасалось с внутренними поверхностями подвздошной кости. Большинство крестцовых ребер были сращены с крестцом и имели довольно изогнутую форму. [ 6 ]


Грудной пояс протоцератопса . образован скапулокоракоидом ( слиянием клювовидного отростка и лопатки) и ключицы Лопатки ( лопатки ) были относительно большими и закругленными с внутренней стороны. В верхней части лопатки были широкими. В нижней части лопатки встречаются с коракоидами. Коракоиды были относительно эллиптическими, а иногда и сращенными (сросшимися) с лопатками. Ключица протоцератопса представляла собой элемент U-образной или слегка V-образной формы, который соединялся с верхним краем лопатокоракоида. В общем виде передние конечности протоцератопса были короче задних и состояли из плечевой, лучевой и локтевой костей. ( Плечевая кость кость плеча) была большой и тонкой, а в нижней части соединялась как с лучевой, так и с локтевой костью. Лучевая кость имела слегка изогнутую форму и была длиннее локтевой кости. В верхней части ее имелась вогнутость, служащая соединением с плечевой костью и образующая локтевой сустав . кость Локтевая представляла собой довольно короткую кость прямой формы. manus (рука) протоцератопса имела пять пальцев (пальцев). Первые три пальца имели когти (когтевые кости) и были самыми большими пальцами. Последние два были лишены когтей и имели небольшие размеры, в основном рудиментарные (сохранились, но без важной функции). Когтевые кости рук и ног были плоскими, тупыми и напоминали копыта. [ 6 ] [ 40 ]
Тазовый пояс образован подвздошной , лобковой и седалищной костями . Подвздошная кость представляла собой крупный элемент, имеющий узкий предвертлужный отросток (передний конец) и широкий поствертлужный отросток (задний конец). Лобковая кость была самым маленьким элементом тазового пояса и имела неправильную форму, хотя ее нижний конец был развит в заостренный костный выступ вниз. Седалищная кость была самой длинной костью тазового пояса. Он имел удлиненное древко с несколько широким нижним концом. Задние конечности протоцератопса были довольно длинными, с немного более длинной большеберцовой костью (кость голени), чем бедренной костью (бедренная кость). ( Бедренная кость бедренная кость) была крепкой и имела довольно округлый и выраженный большой вертел , слегка загнутый внутрь. Большеберцовая кость (голень) была длинной и тонкой, с широким нижним концом. В ее верхней части образовалась вогнутость для соединения с меньшей малоберцовой костью . Пес и четырех пальцев, на (стопа) состоял из четырех плюсневых костей которых были лопатообразные когтевые кости. Первая плюсневая кость и палец были самыми маленькими, остальные элементы имели аналогичную форму и длину. [ 6 ] [ 40 ]
Классификация
[ редактировать ]
протоцератопса В 1923 году Грейнджер и Грегори включили в недавно названное семейство Protoceratopsidae в качестве репрезентативного вида. Это семейство характеризовалось своей общей примитивной морфологией по сравнению с более развитыми Ceratopsidae , такой как отсутствие хорошо развитых роговых ядер и относительно меньший размер тела. Сам протоцератопс авторы считали каким-то образом связанным с анкилозаврами по особенностям черепа, в более усиленной степени с трицератопсом и его родственниками. [ 2 ] Грегори и Чарльз К. Мук в 1925 году после более глубокого анализа протоцератопса и его общей морфологии пришли к выводу, что этот таксон представляет собой цератопса, более примитивного, чем цератопсиды, а не предка анкилозавра-цератопса. [ 7 ] В 1951 году Эдвин Х. Колберт считал протоцератопса ключевым предком линии цератопсидов, предполагая, что это в конечном итоге привело к эволюции крупных цератопсов, таких как стиракозавр и трицератопс . Было высказано предположение, что такая линия произошла от примитивного цератопсового пситтакозавра . Он также считал протоцератопса одним из первых цератопсов с жабо, появившихся в летописи окаменелостей. [ 50 ]
Однако в 1975 году Марьянска и Осмольска утверждали, что очень маловероятно, что протоцератопсиды произошли от пситтакозаврид , а также маловероятно, что они дали начало высокоразвитым (продвинутым) цератопсидам. Первое положение было подтверждено многочисленными анатомическими различиями между протоцератопсидами и пситтакозавридами, в первую очередь крайней редукцией некоторых пальцев рук в последней группе — черта, гораздо менее выраженная у протоцератопсид. Второй момент был объяснен на основе уже сложившейся анатомии протоцератопсид, таких как Багацератопс или Протоцератопс (например, морфологии челюстей). Марианска и Осмольска также подчеркнули, что некоторые ранние представители Ceratopsidae отражают гораздо более древнюю эволюционную историю. [ 10 ] В 1998 году палеонтолог Пол Серено официально определил Protoceratopsidae как ветвящуюся , кладу включающую всех коронозавров, более близких к протоцератопсу , чем к трицератопсу . [ 51 ]

Более того, после повторных исследований Turanoceratops в 2009 году и Zuniceratops — двух важнейших таксонов цератопсов с точки зрения эволюционной истории цератопсид — в 2010 году был сделан вывод, что происхождение цератопсид не связано с летописью окаменелостей протоцератопсов и их родственников и старше их. [ 52 ] [ 53 ] В самых последних/современных филогенетических анализах протоцератопсы и багацератопсы обычно выделяются как сестринские таксоны , оставляя без подтверждения интерпретации, предполагающие прямые связи с более производными цератопсами. [ 54 ]
В 2019 году Чепинский проанализировал подавляющее большинство экземпляров, отнесенных к цератопсам Bagaceratops и Breviceratops , и пришел к выводу, что большинство из них на самом деле были экземплярами первых. Хотя роды Gobiceratops , Lamaceratops , Magnirostris и Platyceratops долгое время считались действительными и отдельными таксонами и иногда помещались в состав Protoceratopsidae, Чепинский обнаружил, что диагностические (идентификаторы) признаки, используемые для различения этих таксонов, в значительной степени присутствуют у Bagaceratops и, таким образом, становятся синонимами этот род. Согласно этому рассуждению, Protoceratopsidae состоит из Bagaceratops , Breviceratops и Protoceratops . Ниже приведены предполагаемые Чепинским отношения между Protoceratopsidae: [ 19 ]
В 2019 году Битнара Ким и коллеги описали относительно хорошо сохранившийся скелет Багацератопса из формации Барун-Гойот , отметив многочисленные сходства с протоцератопсом . Несмотря на то, что анатомия их черепа имела существенные различия, их посткраниальный скелет был практически одинаковым. Филогенетический анализ, проведенный командой, обнаружил, что оба протоцератопсиды являются сестринскими таксонами, что указывает на то, что Bagaceratops и Protoceratops были анатомически и систематически связаны. Ниже представлена полученная кладограмма , показывающая положение протоцератопса и багацератопса : [ 55 ]

Эволюция
[ редактировать ]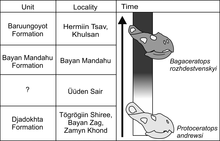
Лонгрич и его команда в 2010 году указали, что высокоразвитая морфология P. hellenikorhinus — по сравнению с P. andrewsi — указывает на то, что этот вид может представлять собой линию протоцератопсов , имевшую более длительную эволюционную историю по сравнению с P. andrewsi , или просто прямой потомок П. Эндрюси . Разница в морфологии протоцератопсов также позволяет предположить, что близлежащая формация Баян-Мандаху немного моложе формации Джадохта. [ 56 ]
В 2020 году Чепинский проанализировал несколько давно неописанных экземпляров протоцератопсидов из местонахождений Удын-Сайр и Замын-Хондт джадохтинской свиты. Было показано, что один экземпляр (MPC-D 100/551B) имеет черты черепа, которые являются промежуточными между Bagaceratops rozhdestvenskyi (который является родным для соседних Баян-Мандаху и Барун-Гойот ) и P. andrewsi . Образец происходит из местонахождения Удын-Сайр, где доминируют останки протоцератопса , и, учитывая отсутствие более убедительных анатомических особенностей, Чепинский присвоил экземпляру вид Bagaceratops sp. Он объяснил, что присутствие этого экземпляра Bagaceratops в такой необычной местности может быть решено путем: (1) сосуществования и симпатрической (в целом) эволюции как Bagaceratops , так и Protoceratops в этом одном месте; (2) появление B. rozhdestvenskyi в другом регионе и возможная миграция в Удын-Сайр; (3) гибридизация между двумя протоцератопсидами, учитывая близкое расположение Баяна Мандаху и Джадохты; (4) анагенетический (прогрессивная эволюция) эволюционный переход от П. Андревси — Б. Рождественский . Среди сценариев анагенетический переход лучше всего поддержал Чепинский, учитывая тот факт, что B. rozhdestvenskyi в Удын-Сайре не обнаружено окончательных окаменелостей , как и ожидалось от события гибридизации; У MPC-D 100/551B отсутствует хорошо развитое дополнительное анторбитальное окно (отверстие позади ноздрей), признак, который, как ожидается, будет присутствовать, если B. rozhdestvenskyi мигрировал в этот район; и многие экземпляры P. andrewsi , обнаруженные в Удын-Сайре, уже характеризуются уменьшением количества примитивных предчелюстных зубов, что подтверждает растущее изменение популяций. [ 57 ]
Палеобиология
[ редактировать ]Кормление
[ редактировать ]В 1955 году палеонтолог Георг Хаас исследовал общую форму черепа протоцератопса и попытался реконструировать мускулатуру его челюстей . Он предположил, что большой воротник на шее , вероятно, был местом прикрепления жевательных мышц. Такое расположение мышц, возможно, помогло закрепить нижние челюсти, необходимые для кормления. [ 58 ] Яннике Дофин и его коллеги в 1988 году описали эмали микроструктуру протоцератопса , наблюдая непризматический внешний слой. Они пришли к выводу, что форма эмали не связана с рационом питания или функцией зубов , поскольку большинство животных не обязательно используют зубы для переработки пищи. Верхнечелюстные зубы цератопсов обычно были упакованы в зубную батарею , образующую вертикальные режущие лезвия, которые, вероятно, рубили листья . Этот метод питания, вероятно, был более эффективным для протоцератопсид, поскольку поверхность эмали протоцератопсов имела грубую текстуру, а кончики микрозубцов развивались на основании зубов, что, вероятно, помогало крошить растительность. Основываясь на их колышкообразной форме и уменьшении микроорнаментации, Дофин и его коллеги предположили, что предчелюстные зубы протоцератопса не имели специфической функции. [ 45 ]
В 1991 году палеонтолог Грегори С. Пол заявил, что вопреки популярному мнению о птицетазовых как облигатных травоядных , некоторые группы, возможно, были оппортунистическими мясоедами , включая представителей Ceratopsidae и Protoceratopsidae. Он отметил, что их выдающиеся, как у попугаев, клювы и острые зубы, а также мощные мышцы челюстей предполагают всеядную диету, во многом похожую на диету свиней , кабанов , кабанов и энтелодонтов . Такой сценарий указывает на возможную конкуренцию с более хищными тероподами за туши , однако, поскольку поедание тканей животных было случайным, а не основной частью их рациона, поток энергии в экосистемах был относительно простым. [ 59 ] Ю Хайлу и Питер Додсон в 2004 году предположили, что предчелюстные зубы протоцератопса могли быть полезны для выборочного сбора урожая и кормления. [ 60 ]
В 2009 году Кё Танке и его команда предположили, что базальные цератопсы, такие как протоцератопсиды, скорее всего, были низкими браузерами из-за их относительно небольшого размера тела. Этот метод с низким уровнем просмотра позволил бы питаться листвой и фруктами в пределах досягаемости, а крупные базальные цератопсы могли потреблять более жесткие семена или растительный материал, недоступный более мелким базальным цератопсам. [ 61 ]
Дэвид Дж. Баттон и Линдси Э. Занно в 2019 году провели масштабный филогенетический анализ, основанный на биомеханических характеристиках черепа, предоставленных 160 видами мезозойских динозавров, чтобы проанализировать множественные проявления травоядных среди нептичьих динозавров. Их результаты показали, что травоядные динозавры в основном придерживались двух различных способов питания: либо переработка пищи в кишечнике, характеризующемся относительно изящными черепами и низкой силой укуса , либо рот, который характеризовался особенностями, связанными с интенсивной переработкой, такими как высокая сила укуса и крепкая челюстная мускулатура. Было обнаружено , что цератопсы (включая протоцератопсидов), наряду с динозаврами Euoplocephalus , Hungarosaurus , паркозавридами , орнитоподами и гетеродонтозавринами , относятся к первой категории, что указывает на то, что протоцератопсы и их родственники обладали сильной силой укуса и полагались в основном на свои челюсти для переработки пищи. [ 62 ]
Онтогенез
[ редактировать ]

Браун и Шлайкьер в 1940 году после обширного описания и пересмотра протоцератопса заметили, что орбиты, лобные и слезные кости уменьшаются в относительных размерах по мере старения животного; верхняя граница ноздрей стала более вертикальной; носовые кости постепенно удлинялись и сужались; и оборка на шее в целом тоже увеличивается с возрастом. В частности, оборка на шее претерпела резкие изменения: от маленькой, плоской и почти округлой структуры у молодых особей до большой, веерообразной у полностью зрелых протоцератопса . особей [ 6 ] В 2001 году Ламберт и его коллеги считали развитие двух носовых «рогов» P. hellenikorhinus признаком, который задерживается по сравнению с появлением полово -дискриминантных признаков. Это было основано на том факте, что один небольшой экземпляр (IMM 96BM2/1) имеет размер черепа немного больше, чем череп предполагаемого половозрелого P. andrewsi (AMNH 6409), но при этом у него отсутствуют двойные носовые рога, присутствующие у полностью зрелого P. hellenikorhinus. . [ 20 ]
Маковицки и его команда в 2007 году провели гистологический анализ нескольких образцов протоцератопса из коллекций Американского музея естественной истории , чтобы получить представление об истории жизни протоцератопса . Исследованные ископаемые кости показали, что протоцератопс замедлил свой онтогенез (рост) примерно на 9–10 годах жизни и прекратился примерно на 11–13 годах. Они также заметили, что максимальная или последняя стадия развития шейного воротника и носового рога наблюдалась у самых старых особей протоцератопса , что указывает на то, что такие черты были онтогенно изменчивыми (то есть они менялись с возрастом). Маковицкий и его команда также заявили, что, поскольку максимальные/радикальные изменения на шее и роге носа присутствовали у большинства взрослых особей, попытка дифференцировать половой диморфизм (анатомические различия между полами) у взрослых протоцератопсов может оказаться неэффективной практикой. [ 63 ]
Дэвид Хоун и его коллеги в 2016 году, проанализировав шейные оборки P. andrewsi , обнаружили, что оборка протоцератопса была непропорционально меньше у молодых особей, росла быстрее, чем у остального животного в онтогенезе, и достигала значительных размеров только у молодых особей. крупные взрослые особи. Другие изменения в ходе онтогенеза включают удлинение предчелюстных зубов, которые меньше у молодых особей и увеличиваются у взрослых, а также увеличение средних нервных отростков в хвостовых или хвостовых позвонках, которые, по-видимому, становятся намного выше по мере приближения к взрослой жизни . [ 64 ]

В 2017 году Мототака Санеоши с командой проанализировал несколько экземпляров протоцератопсов из формации Джадохта , отметив, что от перинатальных /ювенильных до полувзрослых особей теменные и чешуйчатые кости увеличивали свои стороны до задних сторон черепа. От полувзрослых до взрослых особей чешуйчатая кость увеличивалась в размерах больше, чем теменная кость, а оборка расширялась в верхнем направлении. Команда пришла к выводу, что оборка протоцератопса может характеризоваться этими онтогенетическими изменениями. [ 65 ]
В 2018 году палеонтологи Люция Фостович-Фрелик и Юстина Словяк изучили гистологию костей нескольких экземпляров P. andrewsi на поперечных срезах, чтобы проанализировать изменения роста этого динозавра. Отобранные элементы состояли из шейного воротника, бедренной кости, большеберцовой кости, малоберцовой кости, ребер, плечевой кости и лучевой кости и показали, что гистология протоцератопсов оставалась довольно однородной на протяжении всего онтогенеза. Он характеризовался простой фиброламеллярной костью — костной тканью с неправильной, волокнистой текстурой и заполненной кровеносными сосудами — с выраженной плетено -волокнистой костью и низким уровнем ремоделирования кости . Большинство костей протоцератопса сохраняют большое количество костных волокон (включая волокна Шарпи ), которые, вероятно, придавали органу прочность и повышали его эластичность. Команда также обнаружила, что скорость роста бедренной кости увеличилась на подростковой стадии, что указывает на изменения в пропорциях костей, такие как удлинение задних конечностей. Эта скорость роста во многом аналогична скорости роста других мелких травоядных динозавров, таких как примитивные Пситтакозавр или Скутеллозавр . [ 66 ]
Движение
[ редактировать ]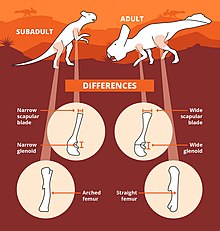
В 1996 году Терещенко реконструировал модель ходьбы протоцератопса , полагая, что наиболее вероятным сценарием будет протоцератопс как обязательное четвероногое животное, учитывая пропорции его конечностей. Основная походка протоцератопса, вероятно, была рысью , в основном с использованием задних конечностей, и маловероятно, что он использовал асимметричную походку. оказался в определенной ситуации (например, в опасности или в поисках пищи), Если бы Протоцератопс он мог бы использовать быстрое факультативное прямохождение . Он также отметил, что плоские и широкие когти протоцератопса, возможно, позволяли эффективно передвигаться по рыхлой местности, например, по песку , который был обычным явлением в его окрестностях. Терещенко с помощью скорости уравнений также оценил среднюю максимальную скорость ходьбы протоцератопса примерно в 3 км/ч ( километров в час ). [ 67 ]
Проанализировав передние конечности нескольких цератопсов, Фил Сентер в 2007 году предположил, что руки протоцератопсов могли достигать земли, когда задние конечности находились в вертикальном положении, а общая морфология передних конечностей и диапазон движений могут отражать, что это было, по крайней мере, факультативное (необязательное). ) четвероногий. Передние конечности протоцератопса могли растягиваться в стороны, но не для передвижения на четвероногих, что достигалось при локтях . поджатых [ 68 ] В 2010 году Александр Кузнецов и Терещенко проанализировали несколько серий позвонков протоцератопса , чтобы оценить общую подвижность, и пришли к выводу, что протоцератопс обладает большей боковой подвижностью в пресакральной (предтазовой) серии позвонков и снижает вертикальную подвижность в шейном (шейном) отделе. [ 48 ] Окаменелый след, связанный с образцом ZPAL Mg D-II/3, описанным Недзведским в 2012 году, указывает на то, что протоцератопс был пальцевидным , то есть он ходил, опираясь на пальцы ног, поддерживая вес тела. [ 39 ]
Однако в 2019 году Словяк и его команда описали элементы конечностей ZPAL Mg D-II/3, который представляет собой полувзрослую особь, и отметили сочетание признаков, типичных для двуногих цератопсов, таких как узкая суставная мышца с лопаточной пластинкой и изогнутая бедренная кость. . Отсутствие этих признаков у зрелых особей указывает на то, что молодые протоцератопсы были способны к факультативному прямохождению, а взрослые особи имели облигатную четвероногие позиции. Несмотря на то, что взрослые протоцератопсы были коренастыми и четвероногими, соотношение длины их большеберцовой и бедренной костей (голень длиннее бедра, что характерно для двуногих цератопсов) предполагает способность иногда стоять на задних конечностях. Словяк и его команда также предположили, что плоские и широкие когти рук (когтевая кость) протоцератопса могли быть полезны для передвижения по рыхлой местности (например, по песку), не погружаясь при этом. [ 40 ]
Копание поведения
[ редактировать ]
Лонгрич в 2010 году предположил, что протоцератопс , возможно, использовал свои задние конечности, чтобы рыть норы или укрываться под кустами и/или царапинами, чтобы избежать самых температур дневных жарких . Роющим действиям задних конечностей, вероятно, способствовала сильная хвостобедренная мышца и большие ступни, снабженные плоскими лопатообразными когтями. Поскольку такое поведение было обычным для протоцератопсов , оно предрасполагало людей к погребению заживо во время внезапного обрушения их нор и высокоэнергетических песчаных событий, таких как песчаные бури , и, таким образом, объясняло положение на месте некоторых экземпляров . Кроме того, Лонгрич предположил, что рытье нор назад может объяснить сохранение некоторых экземпляров, направленных вперед с изогнутыми хвостами. [ 69 ]
В 2019 году Виктория М. Арбор и Дэвид К. Эванс назвали прочность локтевой кости Феррисавра полезной особенностью для рытья, что, возможно, было справедливо и для протоцератопса . [ 70 ]
Хвостовая функция
[ редактировать ]
Грегори и Мук в 1925 году предположили, что протоцератопс был частично водным из-за его больших ступней (которые были больше рук) и очень длинных нервных отростков, обнаруженных в хвостовых (хвостовых) позвонках. [ 7 ] Браун и Шлайкьер в 1940 году указали, что расширение дистального (нижнего) седалищного конца может отражать сильную седалищно-каудальную мышцу, которая вместе с высокими хвостовыми нервными отростками использовалась для плавания . [ 6 ] Барсболд в своем кратком описании экземпляра боевых динозавров в 1974 году принял эту гипотезу и предположил, что протоцератопс был земноводным (адаптированным к воде) и имел хорошо развитые способности к плаванию, основанные на его сплющенном из стороны в сторону хвосте с очень высокими нервными отростками. [ 33 ]
Джек Боуман Бэйли в 1997 году не согласился с предыдущими водными гипотезами и указал, что высокие хвостовые нервные отростки вместо этого больше напоминают луковицеобразные хвосты некоторых видов пустынных ящериц (таких как Heloderma или Uromastyx ), которые связаны с накоплением жира с метаболической водой в хвосте. . Он считал адаптацию к плаванию маловероятной, учитывая засушливые условия джадохтинской свиты. [ 71 ]
В 2008 году на основании нахождения некоторых экземпляров протоцератопсов в речных (речных отложениях) отложениях джадохтинской свиты и гетероцельных (седлообразных на обоих концах) хвостовых позвонков протоцератопсид Терещенко пришел к выводу, что приподнятые хвостовые шипы приспособление для плавания. Он предположил, что протоцератопсиды перемещались по воде, используя свои сплющенные с боков хвосты в качестве весла, помогающего плавать. По словам Терещенко, Багацератопс был полностью водным, а Протоцератопс — лишь частично водным. [ 72 ] Лонгрич в 2010 году утверждал, что высокий хвост и оборка протоцератопса, возможно, помогали ему отводить избыточное тепло в течение дня (действуя как структуры с большой поверхностью), когда животное было активным, чтобы выжить в относительно засушливых условиях джадохтинской свиты без высокоразвитые механизмы охлаждения . [ 69 ]

В 2011 году во время описания Koreaceratops Юонг-Нам Ли и его коллеги обнаружили, что вышеупомянутые гипотезы плавания трудно доказать, поскольку они основаны на обилии Protoceratops в эоловых (отложенных ветром) отложениях, которые отложились в заметных засушливых средах. Они также отметили, что, хотя такие таксоны, как Leptoceratops и Montanoceratops, извлекаются из речных отложений, они, по оценкам, являются одними из самых плохих пловцов. Ли и его коллеги пришли к выводу, что, хотя морфология хвоста Koreaceratops — и других базальных цератопсов — не свидетельствует против привычки плавать, приведенных доказательств этого недостаточно. [ 73 ]
Терещенко в 2013 году исследовал строение позвоночника протоцератопса и пришел к выводу, что он имел приспособления к наземному и водному образу жизни. Проведенные наблюдения показали, что большое количество хвостовых позвонков могло быть полезно для плавания и использования хвоста для уравновешивания веса. Он также указал, что передние хвостовые отростки были лишены высоких нервных отростков и обладали повышенной подвижностью — подвижность, которая начинает уменьшаться по направлению к высоким нервным отросткам, — что позволяет предположить, что хвост мог быть значительно приподнят от основания. Вполне вероятно, что протоцератопс поднимал хвост в качестве сигнала ( показки ) или самки могли использовать этот метод во время откладки яиц , чтобы расширить и расслабить клоаку . [ 49 ]
В 2016 году Хоун и его команда указали, что хвост протоцератопса , особенно средняя часть с приподнятыми нервными отростками, мог использоваться для демонстрации, чтобы произвести впечатление на потенциальных партнеров и/или для распознавания видов. Хвост мог быть связан с такими структурами, как оборка, для демонстрации поведения. [ 64 ]
Ким и его команда в 2019 году отметили, что удлиненные хвостовые шипы хорошо подходят для плавания. Они указали, что и Багацератопс , и Протоцератопс, возможно, использовали свои хвосты одинаковым образом в одинаковых ситуациях, например, при плавании, учитывая, насколько похожими были их посткраниальные скелеты. Команда также предположила, что адаптация к плаванию могла бы быть полезна, чтобы избежать водных хищников, таких как крокодиломорфы . [ 55 ]
Социальное поведение
[ редактировать ]

Томаш Ежикевич в 1993 г. сообщил о нескольких моновидовых (содержащих только один доминирующий вид) комплексах гибели протоцератопсов из формаций Баян-Мандаху и Джадохта. Группа из пяти средних и взрослых протоцератопсов наблюдалась в местонахождении Баян-Мандаху. Особи в этом скоплении лежали на животе, головой вверх, бок о бок, параллельно и под углом примерно 21 градус к горизонтальной плоскости. Две другие группы были обнаружены в местонахождении Тугрикен Шире; одна группа, состоящая из шести человек, и другая группа, состоящая примерно из 12 скелетов. [ 12 ]
В 2014 году Дэвид В.Е. Хон и его коллеги сообщили и описали два блока, содержащие трупные скопления P. andrewsi из Тугрикен Шире. Первый блок (MPC-D 100/526) состоит из четырех молодых особей, находящихся в непосредственной близости с головами вверх, а второй блок (MPC-D 100/534) состоит из двух полувзрослых особей с горизонтальной ориентацией. Основываясь на предыдущих сборках и двух блоках, команда определила, что протоцератопс был социальным динозавром , который формировал стада на протяжении всей своей жизни, и такие стада могли различаться по составу, включая взрослых особей, полувзрослых особей, братьев и сестер из одного гнезда или местных членов. стада, присоединяющегося вскоре после вылупления. Однако, поскольку в группе могли быть потери членов из-за хищничества или других факторов, оставшиеся особи объединялись в более крупные группы, чтобы увеличить свою выживаемость. Хон и его коллеги, в частности, предположили, что молодые особи объединяются в первую очередь для защиты от хищников и повышенной защиты от нескольких взрослых особей внутри группы. Группа также отметила, что, хотя Протоцератопс является прямым свидетельством формирования одиночных когортных скоплений на протяжении всей жизни, нельзя исключать возможность того, что некоторые протоцератопсы были одиночными. [ 14 ]
Половой диморфизм и проявление
[ редактировать ]
Браун и Шлайкьер в 1940 году после обширного анализа протоцератопсов отметили потенциальное наличие полового диморфизма среди экземпляров P. andrewsi , заключив, что это состояние может быть полностью субъективным или отражать реальные различия между полами . Лица с высоким носовым рогом, массивными префронтальными костями и лобно-теменной впадиной предварительно определялись как мужчины . Для самок преимущественно характерно отсутствие хорошо развитых носовых рогов. [ 6 ] В 1972 году Курзанов сравнил черепа P. andrewsi из Байн Дзак и Тугрикен Шире, отметив различия в носовом роге внутри популяций. [ 74 ]
Питер Додсон в 1996 году использовал анатомические особенности черепа P. andrewsi для количественной оценки областей, подверженных онтогенным изменениям и половому диморфизму. Всего было измерено и сравнено 40 символов черепа, включая такие области, как оборка и носовой рог. Додсон обнаружил, что большинство этих признаков сильно различаются у разных экземпляров, особенно оборка, которая, как он интерпретировал, играла более важную роль в отображении поведения, чем просто служила местом расположения жевательных мышц. Он считал такую интерпретацию маловероятной, учитывая относительную хрупкость некоторых костей жабо и большие индивидуальные различия, которые могли повлиять на развитие этих мышц. Додсон обнаружил, что длина оборки у экземпляров имеет довольно неравномерный рост, поскольку у молоди AMNH 6419 длина оборки была меньше, чем у других молодых особей. Он согласился с Брауном и Шлайкьером в том, что высокий, хорошо развитый носовой рог представляет собой мужской признак, а противоположный — женский. Кроме того, Додсон предположил, что такие черты, как носовой рог и оборка, у мужчин Протоцератопсы , возможно, были важным визуальным средством привлечения самок и отпугивания других самцов или даже хищников. что как самцы, так и самки не имели существенных различий в размерах тела и что половую зрелость протоцератопсов Наконец, он отметил , можно было распознать в тот момент, когда можно было отличить самцов от самок. [ 75 ]
В 2001 году Ламберт и его команда после описания P. hellenikorhinus также отметили различия внутри отдельных особей. Так, у некоторых экземпляров (например, голотип IMM 95BM1/1) сохраняются высокие носовые кости с парой рогов; относительно короткая анторбитальная длина; и вертикально ориентированные ноздри. Такие черты считались характерными для самца P. hellenikorhinus . Для другой группы черепов характерны низкие носовые кости с неразвитыми рогами; относительно большая длина анторбитальной области; и более косые ноздри. Эти особи считались самками. Однако команда не смогла провести более глубокий анализ полового диморфизма P. hellenikorhinus из-за отсутствия полных образцов. [ 20 ] Также в 2001 г. Терещенко проанализировал несколько экземпляров P. andrewsi с целью оценки полового диморфизма. Он обнаружил 19 анатомических различий в позвоночнике и тазовой области рассматриваемых особей мужского и женского пола , которые, по его мнению, представляют собой настоящие половые признаки. [ 76 ]
В 2012 г. Наото Ханда с коллегами описали четыре экземпляра P. andrewsi из местонахождения Удын-Сайр джадохтинской свиты. Они указали, что половой диморфизм в этой популяции отмечен выступающим носовым рогом у самцов (признак, также отмеченный другими авторами), относительно более широкими ноздрями у самок и более широкими оборками на шее у самцов. Несмотря на сохранение морфологии черепа большинства экземпляров протоцератопсов (например, предчелюстных зубов), оборка шеи в этой популяции была более прямой и имела почти треугольную форму. Ханда и его команда также обнаружили различия в выборке Удын Сайра и классифицировали их на три группы. К первой группе относятся особи с хорошо развитым костным валиком на латеральной поверхности чешуйчатой кости, причем задний край чешуйчатой кости обращен назад. Вторая группа имела довольно округлый задний край чешуйчатой кости и длинный и хорошо развитый костный гребень на заднем крае теменной кости. Наконец, третья группа характеризовалась изогнутым задним краем чешуйчатой кости и известной морщинистой текстурой верхней поверхности теменной кости. Такие черты черепа считались выраженными. внутривидовые вариации внутри протоцератопсов , и они отличаются от других популяций формации Джадохта (например, Тугрикен Шире), будучи уникальными для региона Удын-Сайр. Эти морфологии шейных оборок отличаются от таковых у протоцератопсов из джадохтинской свиты соседнего местонахождения динозавров Тугрикинского Шира. Морфологические различия среди экземпляров Удын Сайра могут указывать на внутривидовую изменчивость протоцератопсов . [ 77 ] Большой и хорошо развитый костный гребень на теменной кости наблюдался у другого экземпляра P. andrewsi , MPC-D 100/551, также из Удын-Сайра. [ 57 ]

Тем не менее, Леонардо Майорино и его команда в 2015 году провели большой геометрический морфометрический анализ, используя 29 черепов P. andrewsi, чтобы оценить фактический половой диморфизм. Полученные результаты показали, что, за исключением носового рога, который оставался единственной чертой черепа с потенциальным половым диморфизмом, все ранее предложенные признаки для дифференциации гипотетических самцов от самок были в большей степени связаны с онтогенными изменениями и внутривидовыми вариациями, независимыми от пола, особенно шейный воротник. . Геометрические данные не выявили последовательных морфологических различий между особями, которых предыдущие авторы считали самцами и самками, но также слегка подтвердили различия в роструме по всей выборке. Тем не менее Майорино и его команда отметили, что типичный рассматриваемый самец протоцератопса , AMNH 6438, во многом напоминает морфологию рострума AMNH 6466, типичной рассматриваемой самки. Однако они предположили, что подлинные различия между полами все еще могут присутствовать в посткраниальном скелете. Хотя ранее предлагалось для P. hellenikorhinus , команда утверждала, что выборка, использованная для этого вида, была недостаточной, а учитывая, что половой диморфизм не был обнаружен у P. andrewsi , маловероятно, что он имел место у P. hellenikorhinus . [ 78 ]
В 2016 году Хоун и его коллеги проанализировали 37 черепов P. andrewsi и обнаружили, что воротник шеи протоцератопса (как по длине, так и по ширине) претерпел положительную аллометрию во время онтонгении, то есть более быстрый рост/развитие этой области, чем остальная часть черепа. животное. Скуловые кости также имели тенденцию к увеличению относительного размера. Эти результаты позволяют предположить, что они функционировали как сигналы социально-сексуального доминирования или в основном использовались для демонстрации. Использование оборки в качестве демонстрационной структуры может быть связано с другими анатомическими особенностями протоцератопсов, такими как предчелюстные зубы (по крайней мере, у P. andrewsi ), которые могли использоваться в демонстрационных или внутривидовых боях , или высокие нервные отростки хвоста. С другой стороны, Хоун и его команда утверждали, что если бы вместо этого оборки на шее использовались в защитных целях, большие оборки могли бы послужить апосематическим (предупреждающим) сигналом для хищников. Однако такие стратегии наиболее эффективны, когда таксон редок в общей среде обитания, в отличие от Протоцератопс , который выглядит чрезвычайно многочисленным динозавром среднего размера. [ 64 ]
Терещенко в 2018 г. исследовал серию шейных позвонков шести экземпляров P. andrewsi . У большинства из них были различия в одном и том же позвонке, такие как форма и пропорции центров позвонков и ориентация нервных дуг. В соответствии с этими различиями были идентифицированы четыре группы, что позволило сделать вывод, что индивидуальные вариации распространились на позвоночник протоцератопса . [ 79 ]
Тем не менее, в 2020 году Эндрю К. Кнапп и его команда провели морфометрический анализ большой выборки экземпляров P. andrewsi , в первую очередь запутавшись в том, что воротник на шее протоцератопса не имеет индикаторов или доказательств полового диморфизма. Вместо этого полученные результаты показали, что несколько областей черепа протоцератопса независимо различались по скорости роста, онтогенетической форме и морфологии; высокий рост оборки в онтогенезе по отношению к другим участкам тела; и большая вариативность оборки на шее независимо от размера. Кнапп и его команда отметили, что результаты исследования оборки указывают на то, что эта структура играла важную роль в передаче сигналов внутри вида, что согласуется с выбором потенциальных партнеров с качественными украшениями и, следовательно, репродуктивным успехом или передачей сигналов доминирования . Такое использование оборки может свидетельствовать о том, что внутривидовое социальное поведение было очень важно для протоцератопса . Результаты также подтверждают общую гипотезу о том, что оборка на шее цератопсов функционировала как структура социально-сексуальных сигналов. [ 80 ]
Воспроизведение
[ редактировать ]
В 1989 году Уолтер П. Кумбс пришел к выводу, что крокодилы , бескилевые и многоногие птицы являются подходящими современными аналогами гнездового поведения динозавров . Он в основном считал яйца элонгатоолитид принадлежащими протоцератопсам, поскольку скелеты взрослых особей были найдены в непосредственной близости от гнезд , интерпретируя это как свидетельство родительской заботы . Более того, Кумбс считал большую концентрацию яиц протоцератопсов в небольших регионах показателем выраженного филопатрического гнездования (гнездования на одной и той же территории). Гнездо протоцератопса должно было быть выкопано задними конечностями и построено в виде насыпи в форме кратера в центре с яйцами, расположенными полукругом. [ 81 ] Ричард А. Талборн в 1992 году проанализировал различные типы яиц и гнезд — большинство из них на самом деле являются элонгатоолитидами — относящимися к протоцератопсам и их строение. Он выделил типы А и Б, оба имеют вытянутую форму. Яйца типа А отличались от яиц типа Б наличием защемленного конца. На основании сравнений с другими птицетазовыми динозаврами, такими как майазавра и ородромеус , известными по более полным гнездам, Талборн пришел к выводу, что большинство изображений гнезд протоцератопса основано на не полностью сохранившихся кладках и в основном на яйцах типа А, которые, скорее всего, были отложены орнитопод. Он пришел к выводу, что гнезда строились в неглубоких холмах, а яйца откладывались радиально, в отличие от популярных реставраций кратерообразных гнезд протоцератопсов . [ 82 ]

первое подлинное гнездо протоцератопса В 2011 году Дэвид Э. Фастовский и его команда описали (MPC-D 100/530) из местонахождения Тугрикен Шире. Поскольку некоторые особи плотно прижаты к четко очерченному краю гнезда, оно могло иметь круглую или полукруглую форму, как предполагалось ранее, диаметром 70 см (700 мм). Большинство особей в гнезде имели почти одинаковый возраст, размер и рост, что позволяет предположить, что они принадлежали к одному гнезду, а не к совокупности особей. Фастовский и его команда также предположили, что, хотя особи были молодыми, они не были перинатами из-за отсутствия фрагментов яичной скорлупы и их большого размера по сравнению с еще более мелкими молодыми особями из этого местонахождения. Тот факт, что особи, вероятно, проводят некоторое время в гнезде после вылупления для роста, предполагает, что родители протоцератопсов могли заботиться о своих детенышах в гнездах, по крайней мере, на ранних стадиях жизни. Поскольку протоцератопс был относительно базальным (примитивным) цератопсом, это открытие может означать, что другие цератопсы также заботились о своих детенышах. [ 13 ]
В 2017 году Грегори М. Эриксон и его коллеги определили инкубационные периоды P. andrewsi и Hypacrosaurus , используя линии задержки роста (LAGS; линии роста) зубов в образцах эмбрионов ( кладка яиц протоцератопса MPC-D 100/1021). Результаты показывают, что средний период замены эмбриональных зубов составляет 30,68 дней и относительно плезиоморфно (общий по наследству) длительный инкубационный период для P. andrewsi с минимальным инкубационным временем 83,16 дней. [ 31 ] Норелл и его команда в 2020 году снова проанализировали эту кладку и пришли к выводу, что протоцератопс откладывает яйца с мягкой скорлупой. Большинство эмбрионов в этой кладке имеют согнутое положение, а также присутствуют очертания яиц, что позволяет предположить, что они были похоронены в яйце (в яйце). Очертания яиц и эмбрионов указывают на эллипсоидную форму яиц при жизни с размерами около 12 см (120 мм) в длину и 6 см (60 мм) в ширину. Некоторые эмбрионы были связаны с черно-белым ореолом (окружностью). Норелл и его команда провели гистологическое исследование его химического состава , обнаружив следы белковой яичной скорлупы, и по сравнению с другими зауропсидами команда пришла к выводу, что они не были биоминерализованы при жизни и, следовательно, имели мягкую скорлупу. Учитывая, что яйца с мягкой скорлупой более уязвимы к обезвоживанию и раздавливанию, протоцератопс, возможно, закопал свои яйца во влажный песок или почву . Поэтому растущие эмбрионы полагались на внешнее тепло и родительскую заботу. [ 32 ]
Палеопатология
[ редактировать ]В 2018 г. Терещенко исследовал и описал несколько сочлененных шейных позвонков P. andrewsi и сообщил о наличии двух аномально сросшихся позвонков (экз. PIN 3143/9). Слияние позвонков, вероятно, было результатом болезни или внешнего повреждения . [ 79 ]
Взаимодействие хищник-жертва
[ редактировать ]

Барсболд в 1974 году кратко описал экземпляр «Сражающихся динозавров» и обсудил возможные сценарии. Правая нога велоцираптора , прижата к телу протоцератопса а левый серповидный коготь ориентирован в область горла. Протоцератопс . укусил хищника за правую руку, подразумевая, что тот не смог спастись Барсболд предположил, что оба животных утонули, когда упали в болото похожий на водоем, , или похожее на зыбучий песок дно озера, , могло удержать их вместе в последние моменты их боя. [ 33 ]
Осмольска в 1993 году предложила еще две гипотезы, чтобы объяснить их сохранение. Во время смертельной борьбы большая дюна , возможно, обрушилась одновременно, похоронив и Протоцератопса , и Велоцираптора . Другое предположение состоит в том, что велоцираптор собирал мусор уже мертвого протоцератопса , когда тот был похоронен и в конечном итоге убит при неопределенных обстоятельствах. [ 34 ]
В 1995 году Дэвид М. Анвин и его коллеги поставили под сомнение предыдущие объяснения, особенно гипотезу об мусоре, поскольку существовали многочисленные признаки одновременного случая смерти. Например, протоцератопс имеет полупрямую позицию, а его череп почти горизонтален, что было бы невозможно, если бы животное уже было мертво. Правая рука велоцираптора а зажата в челюстях протоцератопса , левая сжимает череп протоцератопса . Более того, он лежит на полу, ноги направлены к животу и горлу жертвы, что указывает на то, что этот велоцираптор не падал. Анвин и его коллеги исследовали отложения, окружающие образец, и предположили, что оба они были заживо погребены мощной песчаной бурей . Они интерпретировали это взаимодействие как то, что протоцератопса схватили и ударили ногой, нанесенный низколежащим велоцираптором . Они также считали возможным, что популяции велоцирапторов знали о приседании протоцератопсов во время мощных песчаных бурь и использовали это для успешной охоты. [ 35 ]

Кеннет Карпентер в 1998 году считал образец боевых динозавров убедительным доказательством того, что тероподы являются активными хищниками , а не падальщиками. Он предложил другой сценарий, в котором множественные раны , нанесенные велоцираптором горлу протоцератопса , к смерти последнего животного привели . В качестве последней попытки Протоцератопс укусил хищника за правую руку и поймал ее под собственным весом, что привело к гибели высыханию велоцираптора и . Отсутствующие конечности протоцератопса впоследствии забрали падальщики. Наконец, обоих животных засыпало песком. Учитывая, что велоцираптор относительно целостный, Карпентер предположил, что он мог быть полностью или частично погребен под песком. [ 83 ]
В 2010 году Дэвид Хоун и его команда сообщили о новом взаимодействии между велоцираптором и протоцератопсом, основанном на следах зубов . В 2008 году в местонахождении Гейт формации Баян-Мандаху было собрано несколько окаменелостей , включая остатки зубов и тел протоцератопсидных и велоцирапторовых динозавров. Команда отнесла эти элементы к протоцератопсу и велоцираптору, главным образом, на основании их распространенности по всей единице, хотя они признали, что обнаруженные останки могут представлять собой разные, но родственные таксоны (в данном случае Linheraptor вместо велоцираптора ). По крайней мере, на восьми окаменелостях тела протоцератопсов имеются следы активных зубов, которые были интерпретированы как следы питания. В отличие от образца боевых динозавров, предполагается, что следы зубов были оставлены дромеозавридами во время поедания туш на поздней стадии, либо во время сбора мусора, либо после группового убийства . Команда заявила, что питание велоцираптора протоцератопсом , вероятно, было относительно распространенным явлением в этих средах и что этот цератопс активно составлял часть рациона протоцератопсов. Велоцираптор . [ 84 ]
В 2016 году Барсболд повторно исследовал экземпляр «Боевых динозавров» и обнаружил несколько аномалий у особи протоцератопса : у обоих коракоидов есть небольшие фрагменты костей, свидетельствующие о переломе грудного пояса; правая передняя конечность и лопатка оторваны влево и назад относительно туловища . Он пришел к выводу, что заметное смещение грудных элементов и правой передней конечности было вызвано внешней силой, которая пыталась их вырвать. Поскольку это событие, вероятно, произошло после смерти обоих животных или в момент, когда движение было невозможно, а у протоцератопса отсутствуют другие элементы тела, Барсболд предположил, что наиболее вероятными виновниками были падальщики. Поскольку протоцератопс считается пастушеским животным , другая гипотеза состоит в том, что члены стада пытались вытащить уже похороненного протоцератопса , что привело к вывиху суставов конечностей. Однако Барсболд отметил, что внутри всего образца нет связанных следов, подтверждающих эту последнюю интерпретацию. Наконец, он восстановил ход боя с Протоцератопс нанес мощный удар по велоцираптору , который использовал когти на ногах, чтобы повредить области горла и живота, а когти рук, чтобы схватить голову травоядного. Перед их похоронами смертельный поединок закончился на земле, а Велоцираптор лежал на спине прямо под Протоцератопсом . После захоронения либо стадо протоцератопсов , либо падальщики оторвали захороненного протоцератопса влево и назад, в результате чего хищник и жертва оказались слегка разделены. [ 36 ]
Ежедневная активность
[ редактировать ]
В 2010 году Ник Лонгрич исследовал относительно большое соотношение орбит и склеротическое кольцо протоцератопса , что он предположил в качестве доказательства ночного образа жизни. Судя по размеру склеротического кольца, протоцератопс имел необычно большое среди протоцератопсид глазное яблоко . У птиц склеротическое кольцо среднего размера указывает на то, что животное является хищником, большое склеротическое кольцо указывает на то, что оно ведет ночной образ жизни, а самый большой размер кольца указывает на то, что оно является активным ночным хищником. Размер глаз является важной адаптацией у хищников и ночных животных, поскольку большее соотношение глаз обеспечивает более высокую чувствительность и разрешение. Из-за энергии, необходимой для поддержания большего размера глазного яблока, и слабости черепа, соответствующей большей орбите, Лонгрич утверждает, что эта структура могла быть адаптацией к ночному образу жизни. Морфология челюстей протоцератопса — более подходящего для переработки растительного материала — и его чрезвычайная численность указывают на то, что он не был хищником, поэтому, если он вел дневной образ жизни животное, то можно было бы ожидать, что оно будет иметь гораздо меньший размер склеротического кольца. [ 69 ]
Однако в 2011 году Ларс Шмитц и Рёсуке Мотани измерили размеры склеротического кольца и глазницы у ископаемых экземпляров динозавров и птерозавров, а также некоторых ныне живущих видов. Они отметили, что в то время как фотопические (дневные) животные имеют меньшие склеротические кольца, скотопические (ночные) животные, как правило, имеют более крупные кольца. Между этими двумя ареалами находятся мезопические ( катемерные ) животные, которые нерегулярно активны в течение дня и ночи. Шмитц и Мотани разделили экологические и филогенетические факторы и, исследовав 164 ныне живущих вида, заметили, что измерения глаз довольно точны при выводе о дневном, катемеральном или ночном образе жизни у вымерших четвероногих . Результаты показали, что протоцератопс был катемерным травоядным, а велоцираптор в основном вел ночной образ жизни, что позволяет предположить, что смертельная схватка сражающихся динозавров могла происходить в сумерках или в условиях низкой освещенности. Наконец, Шмитц и Мотани пришли к выводу, что экологическая ниша является потенциальной главной движущей силой развития повседневной деятельности. [ 85 ] Однако последующее исследование, проведенное в 2021 году, показало, что протоцератопс обладал большей способностью к ночному зрению, чем велоцираптор . [ 86 ]
Палеосреда
[ редактировать ]После формирования Мандаху
[ редактировать ]
На основании общего сходства между фауной позвоночных и отложениями формации Баян-Мандаху и формации Джадохта, формация Баян-Мандаху считается позднемеловой по возрасту, примерно кампанской . Преобладающая литология - красновато-коричневый, плохо сцементированный, мелкозернистый песчаник с небольшим количеством конгломерата и калише . Другие фации включают аллювиальные ( отложившиеся ручьями ) и эоловые ( отложившиеся ветром ) отложения . Вполне вероятно, что отложения в Баян-Мандаху были отложены недолговечными реками и озерами на аллювиальной равнине (равнина, состоящая из отложений, отложенных горными реками) с сочетанием дюнных палеосреды полей в полузасушливом климате . Это образование известно своими окаменелостями позвоночных в позах, напоминающих живые, большая часть которых сохранилась в неструктурированном песчанике, что указывает на катастрофическое быстрое захоронение. [ 12 ] [ 87 ]
Палеофауна Баян-Мандаху по составу очень похожа на близлежащую формацию Джадохта, причем обе формации имеют несколько одних и тех же родов, но различаются конкретными видами. В этой формации P. hellenikorhinus представительным видом является , который разделял свою палеосреду с многочисленными динозаврами , такими как дромеозавриды Linheraptor и Velociraptor osmolskae ; [ 88 ] [ 89 ] овирапториды Machairasaurus и Wulatelong ; [ 56 ] [ 90 ] и троодонтиды Линхевенатор , Папилиовенатор и Филовенатор . [ 91 ] Другие представители динозавров включают альваресзаврид Linhenykus ; [ 92 ] анкилозавр Pinacosaurus mephistocephalus ; [ 93 ] [ 94 ] и близкородственный протоцератопсид Bagaceratops . [ 19 ] Дополнительная фауна этого подразделения включает черепах наньсюнчелид , [ 95 ] и множество чешуйчатых и млекопитающих . [ 96 ] [ 97 ]
Джадохтинская свита
[ редактировать ]
Протоцератопс известен из большинства местонахождений формации Джадохта в Монголии , которая восходит к позднему мелу примерно 71–75 миллионов лет назад и отложилась во время быстрой последовательности изменений полярности в поздней части кампанского этапа. [ 98 ] и песчаники от средне- до мелкозернистых Преобладающие отложения в Джадохте включают преобладающие красновато-оранжевые и бледно-оранжевые до светло-серых пески , калише и редкие речные (речные отложения) отложения. На основании этих компонентов палеообстановка джадохтинской свиты интерпретируется как имеющая жаркий полузасушливый климат с большими полями дюн/песчаными дюнами и несколькими недолговечными водоемами , подобными современной пустыне Гоби . Подсчитано, что в конце кампанского периода и в Маастрихте климат сместится к более мезическим (влажным/влажным) условиям, наблюдаемым в формации Немегт . [ 99 ] [ 100 ] [ 101 ]
Джадохтинская свита разделена на нижнюю байн-дзакскую и верхнюю тургругынскую пачки. Протоцератопс широко известен от обоих представителей, причем P. andrewsi является доминирующим и репрезентативным видом в общей формации. [ 98 ] [ 100 ] В пачке Байн-Дзак (в основном местонахождение Байн-Дзак) обнаружены дромеозавриды Halszkaraptor и Velociraptor mongoliensis ; [ 102 ] [ 103 ] овирапторид Овираптор ; [ 4 ] анкилозавр Pinacosaurus grangeri ; [ 94 ] и троодонтид Saurornithoides . [ 104 ] В состав пачки Байн-Дзак также входит местонахождение Ухаа Толгод, богатое ископаемыми. [ 100 ] палеофауна динозавров состоит из альваресзаврид Коль и Шувууя ; [ 105 ] [ 106 ] анкилозаврид Минотавразавр ; [ 107 ] птицы Апсаравис и Гобиптерикс ; [ 108 ] [ 109 ] дромеозаврид Цааган ; [ 110 ] овирапториды Citipati и Khaan ; [ 111 ] троодонтиды Алмас и Байронозавр ; [ 112 ] [ 113 ] и новый, безымянный протоцератопсид, тесно связанный с Protoceratops . [ 114 ] В пачке Тургругьин (главным образом местонахождение Тугрикен Шире) P. andrewsi делил свою палеосреду с птицей Elsornis ; [ 115 ] дромеозавриды Mahakala и Velociraptor mongoliensis ; [ 102 ] [ 116 ] и орнитомимид Aepyornithomimus . [ 101 ] P. andrewsi также в изобилии встречается в Удын-Сайре. [ 77 ] [ 57 ] где Авимимус и Уданоцератопс . были обнаружены [ 117 ] [ 118 ]
Относительно низкое палеоразнообразие динозавров, небольшие размеры тела большинства динозавров и засушливые условия джадохтинской свиты по сравнению с нэмэгтинской свитой позволяют предположить, что протоцератопс и современная биота жили в стрессовой палеосреде (физические факторы, оказывающие неблагоприятное воздействие на экосистему). . [ 69 ] Кроме того, высокая распространенность окаменелостей протоцератопсидов в засушливых отложениях указывает на то, что эти цератопсы предпочитали теплую среду. [ 56 ] [ 69 ] Хотя P. andrewsi был преобладающим протоцератопсидом в этой формации, предварительные остатки P. hellenikorhinus были обнаружены в местонахождениях Удын-Сайр и Бор-Толгой, что позволяет предположить, что оба вида сосуществовали. Если P. andrewsi встречается в эоловых отложениях (Байн Дзак или Тугрикен Шире), то P. hellenikorhinus встречается в эолово-речных отложениях. Поскольку последний тип отложений также встречается в формации Баян-Мандаху, вполне вероятно, что P. hellenikorhinus предпочитал среду, сочетающую влажные и засушливые условия. [ 119 ]
Тафономия
[ редактировать ]

В 1993 году Ежикевич предположил, что многие особи протоцератопсов с сочлененными суставами погибли, пытаясь освободиться от массивных песчаных тел, которые захватили их во время песчаных бурь и не были перенесены факторами окружающей среды. Он указал на характерную позу некоторых протоцератопсов, Сражающихся динозавров». заключающуюся в том, что тело и голова выгнуты вверх, а передние конечности прижаты по бокам (состояние, известное в отдельных случаях как «стояние»), отсутствие осадочных структур в отложениях, сохраняющих особей, и тафономику « сама история как свидетельство этого катастрофического сохранения. Учитывая, что такая поза свойственна популяциям формаций Баян-Мандаху и Джадохта, Ежикевич указал, что такое поведение не является уникальным для какой-либо местности. Он также считал маловероятным, что эти особи протоцератопсов умерли после того, как закопались в песок, поскольку эти экземпляры встречаются только в бесструктурных песчаниках; изогнутая поза будет представлять трудность условия дыхания ; а роющие , как известно, копают головой вперед и субгоризонтально. [ 12 ]
Фастовский в 1997 году исследовал геологию Тугрикен Шире, что позволило получить представление о тафономии протоцератопса . Он согласился с тем, что сохранение образцов протоцератопсов указывает на то, что они подверглись катастрофическому событию, такому как бури в пустыне, и трупы не были перемещены падальщиками или факторами окружающей среды. Сообщалось также, что несколько изолированных нор, обнаруженных в отложениях в этом месте, проникли в поверхность костей некоторых захороненных особей протоцератопса . Фастовский отметил, что эти два фактора в совокупности указывают на то, что это место было местом высокой биотической активности, в основном состоящей из членистоногих- падальщиков, которые также участвовали в переработке протоцератопсов туш . Согнутое положение большинства захороненных протоцератопсов указывает на высыхание и сморщивание связок / сухожилий ног, шеи и хвоста после смерти. [ 120 ]

В 1998 году во время доклада на конференции в Обществе палеонтологии позвоночных Джеймс И. Киркланд и его команда сообщили о множественных слепках куколок членистоногих и бурениях (туннелях) на образце протоцератопса печально известное количество куколок. с хорошо сочлененными суставами из Тугрикен Шире, найденном в 1997 году. Было обнаружено группами и поодиночке вдоль поверхности костей, преимущественно в области суставов, где следообразователи питались высушенными связками, сухожилиями и хрящами . Рассмотренные куколки из экземпляра представляют собой более цилиндрические структуры с закругленными концами. Сообщалось, что куколки, обнаруженные у этой особи протоцератопса, имели длину 2,5 см (25 мм) и ширину 1 см (10 мм) и лучше всего сравнивались с куколками, приписываемыми одиночным осам . Кроме того, обнаруженные отверстия имеют структуру, отличную от следов, оставленных жуками-кожами . Команда отметила, что и куколки, и следы бурения отражают заметную экологическую связь между тушами динозавров и относительно крупным некрофагов таксоном насекомых- . [ 121 ]
Позже в 2010 году Киркланд и Кеннет Бейдер переописали и обсудили многочисленные следы питания этого экземпляра протоцератопса Фокса» , которого они назвали «Протоцератопс . У этой особи они обнаружили как минимум три типа следов питания; почти круглые отверстия диаметром 0,6–1 см (6,0–10,0 мм), которые, как они обнаружили, лучше всего коррелируют со следами питания, оставленными жуками-кожами; выемки полукруглой формы по краям костей; и разрушение суставных поверхностей, преимущественно в суставах конечностей. Коллеги также отметили, что Протоцератопс на территории Фокса сохраняет связанные с ним следы в осадочных отложениях, что указывает на деятельность некрофагов после того, как животное было захоронено. Киркланд и Бадер пришли к выводу, что взрослые особи большого таксона жуков обнаруживали разлагающиеся трупы, погребенные под песком, и копали их, чтобы накормиться и отложить яйца. Выйдя из яиц, личинки питались трупом перед окукливанием. Последние появившиеся личинки питались высушенными сухожилиями и хрящами в области суставов, что объясняет пресловутую плохую сохранность этих областей в образце, а затем пережевывали сами кости перед окукливанием. Достигнув полной зрелости, взрослые жуки выкапывались обратно на поверхность, скорее всего, оставляя отверстия в костях, и, наконец, начинали искать новые трупы и, таким образом, продолжали перерабатывать остатки. Трупы протоцератопса . [ 122 ]

В 2010 году палеонтологи Юкихидэ Мацумото и Мототака Санеёси сообщили о множественных отверстиях и следах укусов на участках суставов сочлененных экземпляров багацератопса и протоцератопса из местонахождения Тугрикен Шире формации Джадохта и местонахождения Хермиин Цав формации Барун Гойот соответственно. Они интерпретировали поврежденные участки экземпляра Protoceratops как результат активного питания роющих членистоногих, скорее всего, насекомых. [ 123 ] Эти образцы были официально описаны и обсуждены в 2011 году Санеёси и его командой, включая окаменелости велоцираптора и анкилозаврид . Сообщаемые следы были идентифицированы как ямки, выемки, отверстия и каналы на скелетах, особенно в области суставов конечностей. Команда отметила, что весьма вероятно, что они были сделаны насекомыми-падальщиками, однако относительно большие отверстия (около 3 см (30 мм) в ширину) в ребрах и лопатках одного экземпляра протоцератопса (MPC-D100/534) указывают на то, что насекомые В повреждении костей участвовали не только падальщики, но и млекопитающие . Учитывая засушливые и суровые палеоэкологические условия таких единиц, как джадохтинская свита, туши динозавров среднего и крупного размера могли быть важным источником питания для мелких животных. Санеёси и его команда подчеркнули, что высокая частота следов питания на суставах конечностей многочисленных экземпляров и отчеты о предыдущих исследованиях указывают на то, что мелкие животные, возможно, использовали коллаген, обнаруженный в суставном хряще высушенных туш динозавров, в качестве источника азота , которого было мало в засушливых условиях пустыни этих окаменелостей динозавров. [ 124 ]

В 2011 году Фастовский с коллегами пришли к выводу, что молодые особи в гнезде MPC-D 100/530 были быстро захвачены сильным песчаным потоком и погребены заживо. Отложения гнезда предполагают, что отложения образовались в результате смещения дюн или сильных песчаных бурь, а ориентация особей указывает на то, что отложения были принесены преобладающим западно-юго-западным ветром . У большинства особей сохранились растопыренные передние конечности и вытянутые задние конечности, что позволяет предположить, что молодые протоцератопсы пытались противостоять мощному воздушному потоку в первоначально рыхлом песке. До или во время захоронения некоторые, возможно, пытались залезть на других. Поскольку общепризнано, что большинство ископаемых экземпляров в Тугрикен Шире сохранилось в результате быстро мигрирующих дюн и песчаных бурь, Фастовский с коллегами предположил, что подветренные границы гнезда были местом, где воздух был свободен от песка и, следовательно, все молодые протоцератопсы возможно, изо всех сил пытались добраться до этого места, что привело к их окончательному захоронению и возможной смерти. [ 13 ]
Хон и его коллеги в 2014 году указали, что две скопления протоцератопсов в Тугрикен Шире (MPC-D 100/526 и 100/534) предполагают, что отдельные особи умерли одновременно, а не накапливались с течением времени. Так, в блоке из четырех молодых особей сохранились особи с почти одинаковыми позами, пространственным положением, и все они обращены головами вверх, что свидетельствует о том, что они были живы на момент захоронения. При захоронении животные, скорее всего, вообще не были полностью ограничены в движениях, учитывая, что особи МРС-Д 100/526 находятся в относительно нормальном жизненном положении и их никто не беспокоил. Внутри этого блока сохранились как минимум два человека с руками на уровне выше ног, что наводит на мысль о попытках подняться вверх с целью освободиться. Команда также отметила наличие отверстий на черепах и скелетах обоих скоплений, которые, возможно, были оставлены личинками насекомых после смерти животных. [ 14 ]
В 2016 году Мегуру Такеучи и его команда сообщили о многочисленных окаменелых следах питания, сохранившихся на скелетах протоцератопсов из местонахождений Байн Дзак, Тугрикен Шире и Удин Сайр, а также от других динозавров. Сообщалось, что сохранившиеся следы представляли собой ямы, выемки, скважины и туннели, которые они приписывали падальщикам. Диаметр следов питания, сохранившихся на черепе протоцератопса из Байн-Дзака, был больше, чем следы, обнаруженные среди других образцов, что указывает на то, что падальщики, ответственные за эти следы, резко отличались от других создателей следов, сохранившихся на образцах. [ 125 ]
Культурное значение
[ редактировать ]Возможное влияние на легенду о Грифоне
[ редактировать ]Фольклорист и историк науки Адриенн Мэйор из Стэнфордского университета предположила, что прекрасно сохранившиеся ископаемые скелеты протоцератопса , пситтакозавра и других клювастых динозавров, найденные древними скифскими кочевниками, добывавшими золото в горах Тянь-Шаня и Алтая в Центральной Азии , возможно, сыграли роль роль в образе мифического существа, известного как грифон . Грифонов описывали как четвероногих размером с волка или льва с большими когтями и клювом, похожим на хищную птицу; они откладывали яйца в гнезда на земле. [ 126 ]
Додсон в 1996 году отметил, что греческие писатели начали описывать грифона около 675 г. до н.э., в то время, когда появились первые греческие сочинения о скифских кочевниках, хотя контакты со скифскими кочевниками могли произойти раньше, в бронзовом веке, когда греки импортировали олово из Афганистана, перевозившееся по караванные пути через Гоби и другие пустыни. Грифоны описывались как «охраняющие» месторождения золота на засушливых холмах и образованиях из красного песчаника в дикой местности под горами Тянь-Шаня и Алтая. Регион Монголии и Китая , где найдено множество протоцератопсов и других окаменелостей динозавров, богат россыпными золотыми стоками с соседних гор, что придает некоторую достоверность теории о том, что эти окаменелости сыграли роль в описаниях грифонов в седьмом веке до нашей эры, сделанных римлянами. раз. [ 127 ]
Мэр в 2001 и 2011 годах уточнил гипотезу о том, что Протоцератопс оказал влияние на легенду о грифоне, проанализировав письменные детали и художественные образы. Она также процитировала некоторые другие греческие истории о мифологических существах, на которые, возможно, повлияли открытия окаменелостей древних людей, таких как циклопы и гиганты . [ 128 ] [ 129 ]
В 2016 году эта гипотеза подверглась критике со стороны британского палеонтолога и палеохудожника Марка П. Уиттона , поскольку она игнорирует догреческое «искусство и описания грифонов». (Никаких письменных упоминаний о грифонах не известно примерно до 675 г. до н. э., когда впервые упоминается слово gryps/грифон.) Далее Уиттон указывает, что крылья традиционных грифонов расположены над лопатками, а не за шеей, как оборки. Протоцератопса ископаемых , что тела грифонов гораздо больше напоминают тела современных больших кошек, чем тела протоцератопса , и что месторождения золота в Центральной Азии находятся в сотнях километров от известных останков протоцератопса , среди многих других несоответствий. Он утверждает, что проще понять грифона как мифическую комбинацию хорошо известных современных видов животных, чем как древнее заблуждение, связанное с коллекциями окаменелых костей. [ 130 ] Позже Уиттон совместно с Ричардом Хингом опубликовал в 2024 году статью, в которой подробно изложил свои взгляды на тонкую связь между грифонами и протоцератопсами . [ 131 ]
-
 Традиционное изображение грифона
Традиционное изображение грифона -
 Адриенн Мэр предположила, что открытие окаменелостей протоцератопса могло вдохновить или повлиять на истории о грифонах.
Адриенн Мэр предположила, что открытие окаменелостей протоцератопса могло вдохновить или повлиять на истории о грифонах.


См. также
[ редактировать ]- Список образцов динозавров с задокументированной тафономической историей
- Хронология исследований цератопсов
Ссылки
[ редактировать ]- ^ Кольбер, Эдвин Х. (Эдвин Харрис); Найт, Чарльз Роберт (1951). Книга динозавров: правящие рептилии и их родственники . Нью-Йорк: МакГроу-Хилл. п. 153.
- ^ Jump up to: а б с Грейнджер, WW; Грегори, WK (1923). «Protoceratops andrewsi, доцератопсовый динозавр из Монголии» (PDF) . Американский музей естественной истории Novitates (72): 1–9. HDL : 2246/4670 .
- ^ Jump up to: а б с д Эндрюс, Р.К. (1932). Ридс, Калифорния (ред.). Новое завоевание Центральной Азии: рассказ об исследованиях среднеазиатских экспедиций в Монголии и Китае, 1921-1930 гг. (PDF) . Том. 1 (1-е изд.). Нью-Йорк: Американский музей естественной истории. стр. 1–549. OCLC 766770 .
- ^ Jump up to: а б с Осборн, HF (1924). «Три новых теропода, зона протоцератопса, центральная Монголия» (PDF) . Новитаты Американского музея (144): 1–12. HDL : 2246/3223 . OCLC 40272928 .
- ^ Грегори, WK (1927). «Пробелы в биографии монголов». Научный ежемесячник . 24 (2): 169–181. Бибкод : 1927SciMo..24..169G . JSTOR 7818 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Браун, Б.; Шлайкьер, Э.М. (1940). «Строение и взаимоотношения протоцератопсов» . Анналы Нью-Йоркской академии наук . 40 (3): 133–266. Бибкод : 1940NYASA..40..133B . дои : 10.1111/j.2164-0947.1940.tb00068.x . OCLC 1673730 .
- ^ Jump up to: а б с Грегори, ВК; Мук, CC (1925). «О протоцератопсе, примитивном цератопсовом динозавре из нижнего мела Монголии» (PDF) . Новитаты Американского музея (156): 1–10. hdl : 2246/4515 .
- ^ Dashzeveg, D. (1963). "Яйца динозавров" [Dinosaur eggs]. Priroda (in Russian). 9 : 100.
- ^ Jump up to: а б с д Килан-Яворовска, З.; Барсболд, Р. (1972). «Рассказ о польско-монгольских палеонтологических экспедициях 1967–1971 годов» (PDF) . Польская палеонтология . 27 : 1−12.
- ^ Jump up to: а б с Марьянская, Т.; Осмольска, Х. (1975). «Протоцератопсиды (Динозаврии) Азии» (PDF) . Польская палеонтология . 33 : 134–143. Архивировано из оригинала (PDF) 21 сентября 2018 года . Проверено 10 июня 2022 г.
- ^ Курочкин Е.Н.; Барсболд, Р. (2000). «Российско-Монгольские экспедиции и исследования в области палеонтологии позвоночных» (PDF) . В Бентоне, MJ; Шишкин, М.А.; Анвин, DM; Курочкин Е.Н. (ред.). Эпоха динозавров в России и Монголии . Издательство Кембриджского университета. п. 235−255.
- ^ Jump up to: а б с д и ж Ежикевич, Т.; Карри, Пи Джей; Эберт, Д.А.; Джонстон, Пенсильвания; Костер, Э.Х.; Чжэн, Ж.-Ж. (1993). «Корреляционные толщи джадохтинской свиты во Внутренней Монголии Китая: обзор стратиграфии, осадочной геологии и палеонтологии и сравнение с типовым местонахождением в Предалтайской Гоби» . Канадский журнал наук о Земле . 30 (10): 2180–2195. Бибкод : 1993CaJES..30.2180J . дои : 10.1139/e93-190 .
- ^ Jump up to: а б с д Фастовский, Д.Э.; Вейшампель, Д.Б.; Ватабе, М.; Барсболд, Р.; Цогтбаатар, К.; Нармандах, П. (2011). «Гнездо Protoceratops andrewsi (Dinosauria, Ornithischia)» . Журнал палеонтологии . 85 (6): 1035–1041. дои : 10.1666/11-008.1 . JSTOR 41409110 . S2CID 129085129 .
- ^ Jump up to: а б с Хон, DWE; Фарке, А.А.; Ватабе, М.; Сигеру, С.; Цогтбаатар, К. (2014). «Новая массовая смертность молодых протоцератопсов и поведение агрегаций с разделением по размеру у молодых нептичьих динозавров» . ПЛОС ОДИН . 9 (11): е113306. Бибкод : 2014PLoSO...9k3306H . дои : 10.1371/journal.pone.0113306 . ПМК 4245121 . ПМИД 25426957 .
- ^ Спикман, СНФ; Бастианс, Д.; Шульп, А.С. (2015). Частичный череп Protoceratops andrewsi из Среднеазиатской экспедиции в коллекциях Naturalis (Лейден, Нидерланды) . Европейская ассоциация палеонтологов позвоночных.
- ^ Kurzanov, S. M. (1990). "Новый род протоцератопсид из позднего мела Монголии" [A new Late Cretaceous protoceratopsid genus from Mongolia] (PDF) . Paleontological Journal (in Russian) (4): 91−97.
- ^ Серено, ПК (2000). «Летопись окаменелостей, систематика и эволюция пахицефалозавров и цератопсов Азии» (PDF) . В Бентоне, MJ; Шишкин, М.А.; Анвин, DM; Курочкин Е.Н. (ред.). Эпоха динозавров в России и Монголии . Издательство Кембриджского университета. п. 489−492.
- ^ Маковицкий, П.Дж. (2001). «Мозговой череп Montanoceratops cerorhynchus (Dinosauria: Ceratopsia) из формации Каньон Подкова в Альберте» . В Танке, DH; Карпентер, К. (ред.). Жизнь мезозойских позвоночных . Жизнь прошлого. Издательство Университета Индианы. стр. 243−262. ISBN 978-0-253-33907-2 .
- ^ Jump up to: а б с д Чепинский, Л. (2019). «Онтогенез и изменчивость протоцератопсидного динозавра Bagaceratops rozhdestvenskyi из позднего мела пустыни Гоби» (PDF) . Историческая биология . 32 (10): 1394–1421. дои : 10.1080/08912963.2019.1593404 . S2CID 132780322 . Архивировано из оригинала (PDF) 8 июля 2021 года . Проверено 10 июня 2022 г.
- ^ Jump up to: а б с д и ж г час я Ламберт, О.; Годфруа, П.; Ли, Х.; Шан, Южная Корея; Донг, З. (2001). «Новый вид протоцератопсов (Dinosauria, Neoceratopsia) из позднего мела Внутренней Монголии (ННР Китай)» (PDF) . Бюллетень Королевского института естественных наук Бельгии, наук о Земле . 71 :5−28.
- ^ Джи, С.; Чжан, Л.; Лу, Л.; Хао (2017). «Первое открытие позднемеловой фауны протоцератопсид из Алхи, Внутренняя Монголия, Китай» . Acta Geologica Sinica (английское издание) . 91 (5): 1908–1909. Бибкод : 2017AcGlS..91.1908J . дои : 10.1111/1755-6724.13421 . S2CID 134276217 .
- ^ Терещенко В.; Алифанов, В.Р. (2003). «Bainoceratops efremovi, новый протоцератопидный динозавр (Protoceratopidae, Neoceratopsia) из местонахождения Баин-Дзак (Южная Монголия)» . Палеонтологический журнал . 37 (3): 293–302.
- ^ Маковицкий, П.Дж.; Норелл, Массачусетс (2006). «Yamaceratops dorngobiensis, новый примитивный цератопс (Dinosauria: Ornithischia) из мела Монголии» (PDF) . Американский музей Novitates (3530): 1–42. doi : 10.1206/0003-0082(2006)3530[1:YDANPC]2.0.CO;2 . hdl : 2246/5808 .
- ^ Чиннери, Би Джей; Хорнер, младший (2007). «Новый неоцератопсический динозавр, связывающий североамериканские и азиатские таксоны» . Журнал палеонтологии позвоночных . 27 (3): 625–641. doi : 10.1671/0272-4634(2007)27[625:ANNDLN]2.0.CO;2 . S2CID 86091277 .
- ^ Осборн, HF (1924). «Открытие неизвестного континента». Естественная история . 24 (2): 133–149.
- ^ Чжао, ZK (1975). «Микроструктура яичной скорлупы динозавров бассейна Наньсюн, провинция Гуандун. К классификации яиц динозавров» (PDF) . Vertebrata PalAsiatica (на китайском языке). 13 (2): 105–117.
- ^ Михайлов, К.Э. (1994). «Яйца теропод и протоцератопсов динозавров из мела Монголии и Казахстана» . Палеонтологический журнал . 28 (2): 101–120.
- ^ Норелл, Массачусетс; Кларк, Дж. М.; Дашевег, Д.; Барсболд, Р.; Кьяппе, LM; Дэвидсон, Арканзас; Маккенна, MC; Альтангерель, П.; Новачек, MJ (1994). «Эмбрион тероподного динозавра и сходство яиц динозавра из Пылающих скал» . Наука . 266 (5186): 779–782. Бибкод : 1994Sci...266..779N . дои : 10.1126/science.266.5186.779 . JSTOR 2885545 . ПМИД 17730398 . S2CID 22333224 .
- ^ Зеленицкий, ДК; Терриен, Ф. (2008). «Филогенетический анализ репродуктивных особенностей теропод манирапторанов и его значение для паратаксономии яиц». Палеонтология . 51 (4): 807–816. Бибкод : 2008Palgy..51..807Z . дои : 10.1111/J.1475-4983.2008.00770.x . S2CID 84859809 .
- ^ Цой, С.; Барта, Делавэр; Морено-Азанза, М.; Ким, Нью-Хэмпшир; Шоу, Калифорния; Варриккио, диджей (2022). «Микроструктурное описание яйца манираптора Protoceratopsidovum» Статьи по палеонтологии 8 (2): e1430. Бибкод : 2022PPal....8E1430C . дои : 10.1002/spp2.1430 . S2CID 248337010 .
- ^ Jump up to: а б Эриксон, генеральный директор; Зеленицкий, ДК; Кей, ДИ; Норрелл, Массачусетс (2017). «Инкубационные периоды динозавров, определенные непосредственно по подсчету линий роста в эмбриональных зубах, демонстрируют развитие уровня рептилий» . Труды Национальной академии наук . 114 (3): 540–545. Бибкод : 2017PNAS..114..540E . дои : 10.1073/pnas.1613716114 . ПМК 5255600 . ПМИД 28049837 .
- ^ Jump up to: а б Норелл, Массачусетс; Виманн, Дж.; Фаббри, М.; Ты, К.; Марсикано, Калифорния; Мур-Налл, А.; Варриккио, диджей; Пол, Д.; Зеленицкий, ДК (2020). «Первое яйцо динозавра было мягким» (PDF ) Природа 583 (7816): 406–410. Бибкод : 2020Nature.583..406N . дои : 10.1038/ s41586-020-2412-8 ПМИД 32555457 . S2CID 219730449 .
- ^ Jump up to: а б с д Barsbold, R. (1974). "Поединок динозавров" [Dueling dinosaurs]. Priroda (in Russian). 2 : 81−83.
- ^ Jump up to: а б Осмольска, Х. (1993). «Были ли монгольские боевые динозавры действительно сражались?». Преподобный Палеобиол . 7 : 161–162.
- ^ Jump up to: а б Анвин, DM; Перл, А.; Труман, К. (1995). «Протоцератопс и велоцираптор сохранились вместе: данные хищнического поведения хищных динозавров?». Журнал палеонтологии позвоночных . 15 (доп. 003): 57А. дои : 10.1080/02724634.1995.10011277 .
- ^ Jump up to: а б Барсболд, Р. (2016). «Боевые динозавры: положение их тел до и после смерти». Палеонтологический журнал . 50 (12): 1412–1417. Бибкод : 2016PalJ...50.1412B . дои : 10.1134/S0031030116120042 . S2CID 90811750 .
- ^ Jump up to: а б Гринфилд, Т. (2022). «Потерянная мумия Протоцератопса — Приложение» . Неопределенные места WordPress.
- ^ Белл, PR; Хендрикс, К.; Питтман, М.; Кэй, Т.Г.; Майр, Г. (2022). «Изысканно сохранившиеся покровы пситтакозавра и чешуйчатая кожа цератопсовых динозавров» . Коммуникационная биология . 5 (809): 809. дои : 10.1038/s42003-022-03749-3 . ПМЦ 9374759 . ПМИД 35962036 .
- ^ Jump up to: а б Недзведский, Г.; Сингер, Т.; Герлинский, Г.Д.; Локли, МГ (2012). «Скелет протоцератопсида с соответствующим следом из верхнего мела Монголии» (PDF) . Меловые исследования . 33 (1): 7−10. Бибкод : 2012CrRes..33....7N . дои : 10.1016/j.cretres.2011.07.001 .
- ^ Jump up to: а б с д Славяк, Ю.; Терещенко В.С.; Фостович-Фрелик, Л. (2019). «Аппендикулярный скелет Protoceratops andrewsi (Dinosauria, Ornithischia): сравнительная морфология, онтогенетические изменения и значение для передвижения цератопсов, не являющихся цератопсидами» . ПерДж . 7 : е7324. дои : 10.7717/peerj.7324 . ПМЦ 6657679 . ПМИД 31367485 .
- ^ Терещенко, В.С. (2021). «Осевой скелет полувзрослого Protoceratops andrewsi из джадохтинской свиты (верхний мел, Монголия)». Палеонтологический журнал . 55 (7): 1408–1457. Бибкод : 2021PalJ...55.1408T . дои : 10.1134/S0031030121120030 . S2CID 247387644 .
- ^ Хольц, ТР; Рей, Л.В. (2007). Динозавры: самая полная и современная энциклопедия для любителей динозавров всех возрастов . Случайный дом. ISBN 9780375824197 . Список родов Holtz 2012 Информация о весе
- ^ Пол, GS (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. п. 282. ИСБН 9780691167664 .
- ^ Кампионе, Невада; Эванс, округ Колумбия (2020). «Точность и точность оценки массы тела нептичьих динозавров» . Биологические обзоры . 95 (6): 1759–1797. дои : 10.1111/brv.12638 . ПМИД 32869488 . S2CID 221404013 . Дополнительная информация
- ^ Jump up to: а б Дофин, Ю.; Йегер, Ж.-Ж.; Осмольска, Х. (1988). «Микроструктура эмали зубов цератопсов (Reptilia, Archosauria)». Геобиос . 21 (3): 319–327. Бибкод : 1988Geobi..21..319D . дои : 10.1016/S0016-6995(88)80056-1 .
- ^ Тануэ, К.; Вы, Х.-Л.; Додсон, П. (2009). «Сравнительная анатомия избранных базальных зубных рядов цератопсов». Канадский журнал наук о Земле . 46 (6): 425–439. Бибкод : 2009CaJES..46..425S . дои : 10.1139/E09-030 . S2CID 58910055 .
- ^ Jump up to: а б Терещенко, В.С. (2007). «Определитель протоцератопоидных позвонков (Ceratopsia, Dinosauria) из Монголии» . Палеонтологический журнал . 41 (2): 175–188. Бибкод : 2007PalJ...41..175T . дои : 10.1134/S0031030107020086 . S2CID 84954199 .
- ^ Jump up to: а б с Кузнецов А.Н.; Терещенко, В.С. (2010). «Метод оценки латеральной и вертикальной подвижности платикоэлозных позвонков четвероногих» . Палеонтологический журнал . 44 (2): 209–225. Бибкод : 2010PalJ...44..209K . дои : 10.1134/S0031030110020139 . S2CID 84321442 .
- ^ Jump up to: а б Терещенко В.С.; Сингер, Т. (2013). «Особенности строения нервных отростков хвостовых позвонков протоцератопоидов (Ornithischia: Neoceratopsia)» . Палеонтологический журнал . 47 (6): 618–630. Бибкод : 2013PalJ...47..618T . дои : 10.1134/S0031030113060105 . S2CID 84639150 .
- ^ Кольбер, Э.Х. (1951). «Виды динозавров» . Книга о динозаврах: правящие рептилии и их родственники . McGraw-Hill Book Company Inc., стр. 79–83.
- ^ Серено, ПК (1998). «Обоснование филогенетических определений с применением к таксономии динозавров более высокого уровня» (PDF) . Новогодний ежегодник геологии и палеонтологии - Трактаты . 210 (1): 41–83. дои : 10.1127/njgpa/210/1998/41 .
- ^ Сьюс, Х.-К.; Аверьянов, А. (2009). «Turanoceratops tardabilis — первый цератопсидный динозавр из Азии» . Naturwissenschaften . 96 (5): 645–652. Бибкод : 2009NW.....96..645S . дои : 10.1007/s00114-009-0518-9 . ПМИД 19277598 . S2CID 21951969 .
- ^ Вульф, генеральный директор; Киркланд, Дж.И.; Смит, Д.; Пул, К.; Чиннери-Альгейер, Б.; Макдональд, А. (2010). «Zuniceratops christopheri: североамериканский сестринский таксон цератопсид, реконструированный на основе новых данных» . В Райане, MJ; Чиннери-Альгейер, Би Джей; Эберт, Д.А. (ред.). Новые взгляды на рогатых динозавров: Симпозиум цератопсов Королевского музея Тиррелла . Издательство Университета Индианы. стр. 91−98. ISBN 978-0-253-35358-0 .
- ^ Имин Хэ; Питер Дж. Маковицки; Кебай Ван; Шуцин Чен; Корвин Салливан; Фэнлу Хан; Син Сюй; Майкл Дж. Райан; Дэвид К. Эванс; Филип Дж. Карри; Калеб М. Браун; Дон Бринкман (2015). «Новый лептоцератопсид (Ornithischia, Ceratopsia) с уникальной седалищной костью из верхнего мела провинции Шаньдун, Китай» . ПЛОС ОДИН . 10 (12): e0144148. Бибкод : 2015PLoSO..1044148H . дои : 10.1371/journal.pone.0144148 . ПМЦ 4689537 . ПМИД 26701114 .
- ^ Jump up to: а б Ким, Б.; Юн, Х.; Ли, Ю.-Н. (2019). «Посткраниальный скелет багацератопса (Ornithischia: Neoceratopsia) из формации Баруунгойот (верхний мел) в Хермиин-Цаве на юго-западе Гоби, Монголия » Журнал Геологического общества Кореи . 55 (2): 179–190. дои : 10.14770/jgsk.2019.55.2.179 . S2CID 150321203 .
- ^ Jump up to: а б с Лонгрич, Северная Каролина; Карри, Пи Джей; Донг, З. (2010). «Новый овирапторид (Dinosauria: Theropoda) из верхнего мела Баян Мандаху, Внутренняя Монголия» . Палеонтология . 53 (5): 945–960. Бибкод : 2010Palgy..53..945L . дои : 10.1111/j.1475-4983.2010.00968.x .
- ^ Jump up to: а б с Чепинский, Л. (2020). «Новые образцы протоцератопсид улучшают возрастную корреляцию слоев пустыни Гоби верхнего мела» (PDF) . Acta Palaeontologica Polonica . 65 (3): 481–497. дои : 10.4202/app.00701.2019 .
- ^ Хаас, Г. (1955). «Челюстная мускулатура у протоцератопсов и других цератопсов» (PDF) . Новитаты Американского музея (1729 г.): 1–24. hdl : 2246/2444 .
- ^ Пол, GS (1991). «Многие мифы динозаврологии, некоторые старые, некоторые новые» (PDF) . Современная геология . 16 :69−99.
- ^ Хайлу, Ю.; Додсон, П. (2004). «Базальная цератопсия» (PDF) . В Вейшампеле, Д.Б.; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Издательство Калифорнийского университета. п. 493. ИСБН 9780520941434 .
- ^ Тануэ, К.; Грандстафф, бакалавр наук; Вы, Х.-Л.; Додсон, П. (2009). «Механика челюстей при базальной цератопсии (орнитишии, динозаврии)» . Анатомическая запись . 292 (9): 1352–1369. дои : 10.1002/ar.20979 . ПМИД 19711460 .
- ^ Баттон, диджей; Занно, LE (2019). «Повторяющаяся эволюция дивергентных способов травоядности у нептичьих динозавров» (PDF) . Современная биология . 30 (1): 158–168. дои : 10.1016/j.cub.2019.10.050 . ПМИД 31813611 . S2CID 208652510 .
- ^ Маковицкий, П.Дж.; Сэдлер, Р.; Додсон, П.; Эриксон, генеральный директор; Норелл, Массачусетс (2007). «История жизни протоцератопса андрюси из Байн Зага, Монголия». Журнал палеонтологии позвоночных . 27 (доп. 003): 109А. дои : 10.1080/02724634.2007.10010458 . S2CID 220411226 .
- ^ Jump up to: а б с Хон, DWE; Вуд, Д.; Нелл, Р.Дж. (2016). «Положительная аллометрия преувеличенных структур цератопсового динозавра Protoceratops andrewsi поддерживает передачу социально-сексуальных сигналов» (PDF) . Электронная палеонтология (19.1.5A): 1–13. дои : 10.26879/591 .
- ^ Санеоши, М.; Мисима, С.; Цогтбаатар, К.; Майнбаяр, Б. (2017). «Морфологические изменения черепа Protoceratops andrewsi с онтогенетическими процессами» (PDF) . Naturalistae (на японском языке) (21): 1–6.
- ^ Фостович-Фрелик, Л.; Славяк, Ю. (2018). «Гистология костей Protoceratops andrewsi из позднего мела Монголии и ее биологическое значение» (PDF) . Acta Palaeontologica Polonica . 63 (3): 503–517. дои : 10.4202/app.00463.2018 .
- ^ Терещенко В.С. (1996). «Реконструкция передвижения протоцератопса» . Палеонтологический журнал . 30 (2): 232–245.
- ^ Сентер, П. (2007). «Анализ функции передних конечностей у базальных цератопсов». Журнал зоологии . 273 (3): 305–314. дои : 10.1111/j.1469-7998.2007.00329.x .
- ^ Jump up to: а б с д и Лонгрич, Северная Каролина (2010). «Функция больших глаз у протоцератопса: ночной цератопс?» . В Райане, MJ; Чиннери-Альгейер, Би Джей; Эберт, Д.А. (ред.). Новые взгляды на рогатых динозавров: Симпозиум цератопсов Королевского музея Тиррелла . Издательство Университета Индианы. стр. 308−327. ISBN 978-0-253-35358-0 .
- ^ Арбур, ВМ; Эванс, округ Колумбия (2019). «Новый динозавр-лептоцератопсид из отложений маастрихтского возраста в бассейне Сустут, север Британской Колумбии, Канада» . ПерДж . 7 : е7926. дои : 10.7717/peerj.7926 . ПМЦ 6842559 . ПМИД 31720103 .
- ^ Бейли, Дж. Б. (1997). «Удлинение нервного отдела позвоночника у динозавров: парусники или буйволы?» . Журнал палеонтологии . 71 (6): 1124–1146. Бибкод : 1997JPal...71.1124B . дои : 10.1017/S0022336000036076 . JSTOR 1306608 . S2CID 130861276 .
- ^ Терещенко, В.С. (2008). «Адаптивные особенности протоцератопсид (Ornithischia: Neoceratopsia)» . Палеонтологический журнал . 42 (3): 50–64. Бибкод : 2008PalJ...42..273T . дои : 10.1134/S003103010803009X . S2CID 84366476 .
- ^ Ли, Ю.-Н.; Райан, MJ; Кобаяши, Ю. (2011). «Первый цератопсовый динозавр из Южной Кореи» (PDF) . Naturwissenschaften . 98 (1): 39−49. Бибкод : 2011NW.....98...39L . дои : 10.1007/s00114-010-0739-y . ПМИД 21085924 . S2CID 23743082 .
- ^ Курзанов, С.М. (1972). «Половой диморфизм у протоцератопсов». Палеонтологический журнал (1): 91−97.
- ^ Додсон, П. (1976). «Количественные аспекты относительного роста и полового диморфизма у протоцератопсов». Журнал палеонтологии . 50 (5): 929–940. JSTOR 1303590 .
- ^ Терещенко, В.С. (2001). «Половой диморфизм посткраниального скелета протоцератопсид (Neoceratopsia, Protoceratopsidae) из Монголии» . Палеонтологический журнал . 35 (4): 415–425. hdl : 123456789/25744 .
- ^ Jump up to: а б Ханда, Н.; Ватабе, М.; Цогтбаатар, К. (2012). «Новые экземпляры протоцератопсов (Dinosauria: Neoceratopsia) из верхнего мела в Удын-Сайре, Южная Гоби, Монголия». Палеонтологические исследования . 16 (3): 179–198. дои : 10.2517/1342-8144-16.3.179 . S2CID 130903035 .
- ^ Майорино, Л.; Фарке, А.А.; Коцакис, Т.; Пирас, П. (2015). «Самцы напоминают самок: переоценка полового диморфизма у Protoceratops andrewsi (Neoceratopsia, Protoceratopsidae)» . ПЛОС ОДИН . 10 (5): e0126464. дои : 10.1371/journal.pone.0126464 . ПМЦ 4423778 . ПМИД 25951329 .
- ^ Jump up to: а б Терещенко, В.С. (2018). «О полиморфизме протоцератопсов Эндрюси Грейнджер и Грегори, 1923 (Protoceratopidae, Neoceratopsia)» . Палеонтологический журнал . 52 (4): 429–444. Бибкод : 2018PalJ...52..429T . дои : 10.1134/S0031030118040135 . S2CID 92796229 .
- ^ Кнапп, AC; Нелл, Р.Дж.; Хон, DWE (2021). «Трехмерный геометрический морфометрический анализ черепа Protoceratops andrewsi подтверждает социально-сексуальную сигнальную роль цератопсового воротника» . Труды Королевского общества B: Биологические науки . 288 (1944). дои : 10.1098/rspb.2020.2938 . ПМЦ 7893235 . ПМИД 33529562 .
- ^ Кумбс, WP (1989). «Современные аналоги гнездования динозавров и родительского поведения». В Фарлоу, Джо (ред.). Палеобиология динозавров . Специальные статьи Геологического общества Америки. Том. Специальный доклад Геологического общества Америки 238. Колорадо: Боулдер. стр. 21−54. дои : 10.1130/SPE238-p21 . ISBN 0-8137-2238-1 .
- ^ Талборн, РА (1992). «Гнездо динозавра Протоцератопса» . Летайя . 25 (2): 145–149. Бибкод : 1992Лета..25..145Т . дои : 10.1111/j.1502-3931.1992.tb01379.x .
- ^ Карпентер, К. (1998). «Свидетельства хищнического поведения хищных динозавров» (PDF) . Гайя . 15 : 135–144.
- ^ Хон, Д.; Шуаньер, Ж.; Салливан, К.; Сюй, Х.; Питтман, М.; Тан, К. (2010). «Новые доказательства трофических взаимоотношений между динозаврами велоцираптором и протоцератопсом». Палеогеография, Палеоклиматология, Палеоэкология . 291 (3–4): 488–492. Бибкод : 2010PPP...291..488H . дои : 10.1016/j.palaeo.2010.03.028 .
- ^ Шмитц, Л.; Мотани, Р. (2011). «Ночной образ жизни динозавров, выведенный на основании морфологии склерального кольца и орбиты». Наука . 332 (6030): 705–708. Бибкод : 2011Sci...332..705S . дои : 10.1126/science.1200043 . ПМИД 21493820 . S2CID 33253407 .
- ^ Шуаньер, Иона Н.; Нинан, Джеймс М.; Шмитц, Ларс; Форд, Дэвид П.; Шапель, Кимберли Э.Дж.; Баланофф, Эми М.; Сипла, Джастин С.; Джорджи, Джастин А.; Уолш, Стиг А.; Норелл, Марк А.; Сюй, Син; Кларк, Джеймс М.; Бенсон, Роджер Би Джей (7 мая 2021 г.). «Эволюция зрения и слуха у тероподовых динозавров» . Наука . 372 (6542): 610–613. Бибкод : 2021Sci...372..610C . дои : 10.1126/science.abe7941 . ISSN 0036-8075 . ПМИД 33958472 . S2CID 233872840 .
- ^ Эберт, Д.А. (1993). «Обстановка осадконакопления и фациальные переходы красноцветных отложений верхнего мела, содержащих динозавров, в Баян Мандаху (Внутренняя Монголия, Китайская Народная Республика)». Канадский журнал наук о Земле . 30 (10): 2196–2213. Бибкод : 1993CaJES..30.2196E . дои : 10.1139/e93-191 .
- ^ Годфруа, П.; Карри, Пи Джей; Ли, Х.; Шан, CY; Донг, З.-М. (2008). «Новый вид велоцираптора (Dinosauria: Dromaeosauridae) из верхнего мела северного Китая». Журнал палеонтологии позвоночных . 28 (2): 432–438. doi : 10.1671/0272-4634(2008)28[432:ANSOVD]2.0.CO;2 . JSTOR 20490961 . S2CID 129414074 .
- ^ Син, X .; Шуанере, JN; Питтман, М.; Тан, Квинсленд; Сяо, Д.; Ли, ZQ; Тан, Л.; Кларк, Дж. М.; Норелл, Массачусетс; Хоне, DW E; Салливан, К. (2010). «Новый дромеозаврид (Dinosauria: Theropoda) из верхнемеловой формации Улансухай во Внутренней Монголии, Китай» (PDF) . Зоотакса . 2403 (1): 1–9. дои : 10.11646/zootaxa.2403.1.1 .
- ^ Син, X .; Цин-Вэй, Т.; Шуо, В.; Салливан, К.; Хон, DWE; Фэн-Лу, Х.; Цин-Ю, М.; Лин, Т.; Донг, Т. (2013). «Новый овирапторид из верхнего мела Ней-Монгола, Китай, и его стратиграфические последствия» (PDF) . Позвоночные Палазиатские . 51 (2): 85–101.
- ^ Пей, Р.; Цинь, Юин; Вэнь, Айшу; Чжао, К.; Ван, З.; Лю, З.; Го, В.; Лю, П.; Йе, В.; Ван, Л.; Инь, З.; Дай, Р.; Сюй, X. (2022). «Новый троодонтид из верхнемелового бассейна Гоби во внутренней Монголии, Китай». Меловые исследования . 130 (105052): 105052. Бибкод : 2022CrRes.13005052P . дои : 10.1016/j.cretres.2021.105052 . S2CID 244186762 .
- ^ Син, X .; Салливан, Корвин; Питтман, М.; Шуаньер, Ж.Н.; Хон, DWE; Апчерч, П.; Тан, К.; Сяо, Донг; Лин, Тан; Хан, Ф. (2011). «Монодактильный нептичий динозавр и сложная эволюция альваресзавроидной руки» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (6): 2338–2342. Бибкод : 2011PNAS..108.2338X . дои : 10.1073/pnas.1011052108 . ПМК 3038769 . ПМИД 21262806 .
- ^ Годфруа, П.; Переда-Субервиола, X.; Ли, Х.; Донг, З.М. (1999). «Новый вид анкилозавридного динозавра Pinacosaurus из позднего мела Внутренней Монголии (КНР)» (PDF) . Бюллетень Королевского института естественных наук Бельгии, Науки о Земле . 69 (приложение Б): 17–36.
- ^ Jump up to: а б Карри, Пи Джей; Бадамгарав, Д.; Коппельхус, Э.Б.; Сиссонс, Р.; Викариус, МК (2011). «Руки, ноги и поведение пинакозавра (Dinosauria: Ankylosauridae)» (PDF ) Польский палеонтологический акт . 56 (3): 489–504. дои : 10.4202/app.2010.0055 . S2CID 129291148 .
- ^ Бринкман, Д.Б.; Тонг, Х.-Ю.; Ли, Х.; Сан, Ю.; Чжан, Ж.-С.; Годфруа, П.; Чжан, З.-М. (2015). «Новые исключительно хорошо сохранившиеся экземпляры «Zangerlia» neimongolensis из Баяна Мандаху, Внутренняя Монголия, и их таксономическое значение» . Comptes Рендус Палевол . 14 (6–7): 577–587. Бибкод : 2015CRPal..14..577B . дои : 10.1016/j.crpv.2014.12.005 .
- ^ Гао, К.; Хоу, Л. (1996). «Систематика и таксономическое разнообразие чешуек верхнемеловой формации Джадохта, Баян Мандаху, пустыня Гоби, Китайская Народная Республика». Канадский журнал наук о Земле . 33 (4): 578–598. Бибкод : 1996CaJES..33..578G . дои : 10.1139/e96-043 .
- ^ Уибл, младший; Шелли, СЛ; Би, С. (2019). «Новый род и вид Djadochtatheriid Multituberculate (Allotheria, Mammalia) из верхнемеловой формации Баян Мандаху Внутренней Монголии» . Анналы музея Карнеги . 85 (4): 285–327. дои : 10.2992/007.085.0401 . S2CID 210840006 .
- ^ Jump up to: а б Дашевег, Д.; Дингус, Л.; Петля, Д.Б.; Свишер III, CC; Дулам, Т.; Суини, MR (2005). «Новое стратиграфическое подразделение, условия осадконакопления и оценка возраста верхнемеловой джадохтинской свиты, южный бассейн Улан-Нура, Монголия» (PDF) . Новитаты Американского музея (3498): 1–31. doi : 10.1206/0003-0082(2005)498[0001:NSSDEA]2.0.CO;2 . hdl : 2246/5667 . S2CID 55836458 .
- ^ Ежикевич, Т. (1997). «Джадохтинская свита». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 188 −191. ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б с Дингус, Л.; Петля, БД; Дашевег, Д.; Свишер III, CC; Минджин, К.; Новачек, MJ; Норелл, Массачусетс (2008). «Геология Ухаа Толгода (джадохтинская свита, верхний мел, бассейн Немегта, Монголия)» (PDF ) Новитаты Американского музея (3616 г.): 1–40. дои : 10.1206/442.1 . hdl : 2246/5916 . S2CID 129735494 . [ постоянная мертвая ссылка ]
- ^ Jump up to: а б Чинзориг, Т.; Кобаяши, Ю.; Цогтбаатар, К.; Карри, Пи Джей; Ватабе, М.; Барсболд, Р. (2017). «Первые орнитомимиды (Theropoda, Ornithomimosauria) из верхнемеловой джадохтинской свиты Тёгрогийн Ширэ, Монголия» . Научные отчеты 7 (5835): 5835. Бибкод : 2017NatSR...7.5835C . дои : 10.1038/ s41598-017-05272-6 ПМЦ 5517598 . ПМИД 28724887 .
- ^ Jump up to: а б Норелл, Массачусетс; Маковицкий, П.Дж. (1999). «Важные особенности скелета дромеозаврида II: информация из недавно собранных экземпляров Velociraptor mongoliensis». Новитаты Американского музея (3282): 1–45. HDL : 2246/3025 . OCLC 802169086 .
- ^ Кау, А.; Бейранд, В.; Воетен, DFAE; Фернандес, В.; Таффоро, П.; Штейн, К.; Барсболд, Р.; Цогтбаатар, К.; Карри, Пи Джей; Годфруа, П. (2017). «Синхротронное сканирование выявило экоморфологию земноводных в новой кладе птицеподобных динозавров» . Природа . 552 (7685): 395–399. Бибкод : 2017Natur.552..395C . дои : 10.1038/nature24679 . ПМИД 29211712 . S2CID 4471941 .
- ^ Норелл, Массачусетс; Маковицкий, П.Дж.; Бевер, Г.С.; Баланов, А.М.; Кларк, Дж. М.; Барсболд, Р.; Роу, Т. (2009). «Обзор монгольского мелового динозавра Saurornithoides (Troodontidae, Theropoda)» . Новитаты Американского музея (3654 г.): 1–63. дои : 10.1206/648.1 . hdl : 2246/5973 .
- ^ Сузуки, С.; Кьяппе, LM; Дайк, Дж.Дж.; Ватабе, М.; Барсболд, Р.; Цогтбаатар, К. (2002). «Новый экземпляр Shuvuuia Deserti Chiappe et al., 1998, из позднего мела Монголии с обсуждением взаимоотношений альваресзаврид с другими тероподами-динозаврами» . Вклад в науку . 494 : 1–18. дои : 10.5962/стр.226791 . S2CID 135344028 .
- ^ Тернер, А.Х.; Несбитт, С.Дж.; Норелл, Массачусетс (2009). «Крупный альваресзаурид из позднего мела Монголии» (PDF) . Новитаты Американского музея (3648): 1–14. дои : 10.1206/639.1 . hdl : 2246/5967 . S2CID 59459861 .
- ^ Алиса, Дж.; Лоуэн, М. (2013). «Новый материал минотавразавра из формации Джодокта устанавливает новые таксономические и стратиграфические критерии для этого таксона» . Журнал палеонтологии позвоночных . Программа и тезисы: 76.
- ^ Кьяппе, LM; Норелл, Массачусетс; Кларк, Дж. (2001). «Новый череп Gobipteryx minuta (Aves: Enantiornithes) из мела пустыни Гоби» (PDF) . Новитаты Американского музея (3346): 1–15. doi : 10.1206/0003-0082(2001)346<0001:ANSOGM>2.0.CO;2 . hdl : 2246/2899 . S2CID 51857603 . .
- ^ Кларк, Дж.А.; Норелл, Массачусетс (2002). «Морфология и филогенетическое положение Apsaravis ukhaana из позднего мела Монголии» (PDF) . Новитаты Американского музея (3387): 1–46. doi : 10.1206/0003-0082(2002)387<0001:TMAPPO>2.0.CO;2 . HDL : 2246/2876 . S2CID 52971055 .
- ^ Норелл, Массачусетс; Кларк, Дж. М.; Тернер, АХ; Маковицкий, П.Дж.; Барсболд, Р.; Роу, Т. (2006). «Новый теропод-дромеозавр из Ухаа Толгод (Омнёгов, Монголия) » Американский музей Novitates (3545): 1–51. doi : 10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2 . hdl : 2246/5823 .
- ^ Кларк, Дж. М.; Норелл, Массачусетс; Барсболд, Р. (2001). «Два новых овирапторида (Theropoda: Oviraptorosauria) из позднемеловой джадоктинской свиты, Ухаа Толгод» . Журнал палеонтологии позвоночных . 21 (2): 209–213. doi : 10.1671/0272-4634(2001)021[0209:TNOTOU]2.0.CO;2 . JSTOR 20061948 . S2CID 86076568 .
- ^ Маковицкий, П.Дж.; Норелл, Массачусетс; Кларк, Дж. М.; Роу, TE (2003). «Остеология и взаимоотношения Byronosaurus jaffei (Theropoda: Troodontidae)» (PDF) . Новитаты Американского музея (3402): 1–32. doi : 10.1206/0003-0082(2003)402<0001:oarobj>2.0.co;2 . hdl : 2246/2828 . S2CID 51824767 .
- ^ Пей, Р.; Норелл, Массачусетс; Барта, Д.Э.; Бевер, Г.С.; Питтман, М.; Сюй, X. (2017). «Остеология нового экземпляра троодонтида позднего мела из Ухаа Толгода, Омногови аймака, Монголия» (PDF ) Новитаты Американского музея (3889): 1–47. дои : 10.1206/3889.1 . hdl : 2246/6818 . S2CID 90883541 .
- ^ Прието-Маркес, А.; Гарсиа-Порта, Дж.; Джоши, Ш.; Норелл, Массачусетс; Маковицкий, П.Дж. (2020). «Модульность и гетерохрония в эволюции оборки цератопсовых динозавров» . Экология и эволюция . 10 (13): 6288–6309. Бибкод : 2020EcoEv..10.6288P . дои : 10.1002/ece3.6361 . ПМЦ 7381594 . ПМИД 32724514 .
- ^ Кьяппе, LM; Сузуки, С.; Дайк, Дж.Дж.; Ватабе, М.; Цогтбаатар, К.; Барсболд, Р. (2007). «Новая энантиорнитиновая птица из позднего мела пустыни Гоби». Журнал систематической палеонтологии . 5 (2): 193–208. Бибкод : 2007JSPal...5..193C . дои : 10.1017/S1477201906001969 . S2CID 85391743 .
- ^ Тернер, А.Х.; Пол, Д.; Кларк, Дж.А.; Эриксон, генеральный директор; Норелл, Массачусетс (2007). «Базальный дромеозаврид и эволюция размеров, предшествовавшая полету птиц» . Наука . 317 (5843): 1378–1381. Бибкод : 2007Sci...317.1378T . дои : 10.1126/science.1144066 . ПМИД 17823350 .
- ^ Курзанов, С.М. (1992). «Гигантский протоцератопсид из верхнего мела Монголии». Палеонтологический журнал : 81−93.
- ^ Ватабе, М.; Сузуки, С.; Цогтбаатар, К. (2006). «Геологическое и географическое распространение птицеподобного теропода Авимимуса в Монголии». Журнал палеонтологии позвоночных . 26 (доп. 003): 136А-137А. дои : 10.1080/02724634.2006.10010069 . S2CID 220413406 .
- ^ Чиба, К.; Райан, MJ; Санеоши, М.; Кониши, С.; Ямамото, Ю.; Майнбаяр, Б.; Цогтбаатар, К. (12–16 октября 2020 г.). Таксономическая переоценка образцов протоцератопсов (Dinosauria: Ceratopsia) из Удын-Сайра, Монголия (PDF) . 80-е ежегодное собрание Общества палеонтологии позвоночных. п. 104.
- ^ Фастовский, Д.Э. (1997). «Палеообстановка Тугрикин-Шире (пустыня Гоби, Монголия) и аспекты тафономии и палеоэкологии протоцератопсов (Dinosauria: Ornithishichia)». ПАЛЕОС . 12 (1): 59−70. Бибкод : 1997Palai..12...59F . дои : 10.2307/3515294 . JSTOR 3515294 .
- ^ Киркланд, Дж.И.; Дельгадо, ЧР; Чимедцерен, А.; Хасиотис, СТ; Фокс, Э.Дж. (1998). «Насекомое? скученные скелеты динозавров и связанные с ними куколки из джадохтинской свиты (мел, кампан), Монголия» . Журнал палеонтологии позвоночных . 18 (доп. 003): 56А. дои : 10.1080/02724634.1998.10011116 . JSTOR i406883 .
- ^ Киркланд, Дж.И.; Бадер, К. (2010). «Окаменелости насекомых, связанные с тушами протоцератопсов в формации Джадохта (верхний мел), Монголия: судебно-медицинская энтомология в верхнем мелу» . В Райане, MJ; Чиннери-Альгейер, Би Джей; Эберт, Д.А. (ред.). Новые взгляды на рогатых динозавров: Симпозиум цератопсов Королевского музея Тиррелла . Издательство Университета Индианы. стр. 509–519. ISBN 9780253353580 . JSTOR j.ctt16gzgng .
- ^ Мацумото, Ю.; Санеоши, М. (2010). «Скучающие скелеты динозавров» . Журнал Геологического общества Японии . 116 (1): I – II. дои : 10.5575/geosoc.116.1.I_II .
- ^ Санеоши, М.; Ватабе, М.; Сузуки, С.; Цогтбаатар, К. (2011). «Следы окаменелостей на костях динозавров из эоловых отложений верхнего мела в Монголии: тафономическая интерпретация палеоэкосистем в древних пустынных условиях» . Палеогеография, Палеоклиматология, Палеоэкология . 311 (1–2): 38–47. Бибкод : 2011PPP...311...38S . дои : 10.1016/j.palaeo.2011.07.024 .
- ^ Такеучи, М.; Санеоши, М.; Цогтбаатар, К.; Майнбаяр, Б.; Ульзийтерен, С. (2016). «Отслеживание окаменелостей на скелетах динозавров из верхнего мела пустыни Гоби, Монголия» . Бюллетень Научно-исследовательского института естественных наук Научного университета Окаямы (46): 1–6.
- ^ Мэр А. (1994). «Хранители золота». Археология . 47 (6): 53–58.
- ^ Додсон, П. (1996). Рогатые динозавры . Издательство Принстонского университета, Принстон, Нью-Джерси. стр. 200−234 . ISBN 978-0-691-05900-6 .
- ^ Мэр А. (2000). Первые охотники за ископаемыми: палеонтология в греческие и римские времена (1-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. стр. 1–384. ISBN 978-0-691-05863-4 . JSTOR j.ctt7s6mm .
- ^ Мэр А. (2011). Первые охотники за ископаемыми: палеонтология в греческие и римские времена (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. стр. 1−400. ISBN 978-0-691-15013-0 .
- ^ Виттон, член парламента (4 апреля 2016 г.). «Почему Протоцератопс почти наверняка не был источником вдохновения для легенды о грифоне» . Блог Марка Уиттона . Блогер.
- ^ Виттон, Марк П.; Хинг, Ричард А. (20 июня 2024 г.). «Рогатый динозавр Протоцератопс вдохновил грифона?» . Междисциплинарные научные обзоры . дои : 10.1177/03080188241255543 . ISSN 0308-0188 .



