Гаплодиплоиды
| Часть серии о |
| Секс |
|---|
 |
| Биологические термины |
| Половое размножение |
| Сексуальность |
| Сексуальная система |
Гаплодиплоидия — это система определения пола , при которой самцы развиваются из неоплодотворенных яиц и являются гаплоидными , а самки развиваются из оплодотворенных яиц и являются диплоидными . [ 1 ] Гаплодиплоидию иногда называют арренотокией .
Гаплодиплоидия определяет пол у всех представителей отрядов насекомых Hymenoptera ( пчёл , муравьев , ос ). [ 2 ] и Thysanoptera («трипсы»). [ 3 ] Система также спорадически встречается у некоторых паутинных клещей , полужесткокрылых , жесткокрылых ( короедов ) и коловраток .
В этой системе пол определяется количеством наборов хромосом, которые получает человек. Потомство, образовавшееся в результате слияния спермия и яйцеклетки , развивается как самка, а неоплодотворенная яйцеклетка — как самец. Это означает, что самцы имеют вдвое меньше хромосом, чем самки, и являются гаплоидными .
Гаплодиплоидная система определения пола имеет ряд особенностей. Например, у мужчины нет отца и он не может иметь сыновей, но у него есть дедушка и он может иметь внуков. Кроме того, если в эусоциальной колонии насекомых имеется только одна королева, и она спаривалась только один раз, то родство между рабочими (диплоидными самками) в улье или гнезде 3 ⁄ 4 . Это означает, что рабочие в таких моногамных колониях с одной маткой значительно более тесно связаны, чем в других системах определения пола, где родство братьев и сестер обычно не превышает 1 ⁄ 2 . Именно этот момент лежит в основе теории родственного отбора о том, как развивалась эусоциальность. [ 4 ] Действительно ли гаплодиплоидия открыла путь к эволюции эусоциальности, до сих пор остается предметом споров. [ 5 ] [ 6 ]
Другая особенность системы гаплодиплоидии заключается в том, что рецессивные летальные и вредные аллели будут быстро удаляться из популяции, поскольку они автоматически экспрессируются у самцов (доминантные летальные и вредные аллели удаляются из популяции каждый раз, когда они возникают, поскольку они убивают любую особь). они возникают в). [ 3 ]
Гаплодиплоидия — это не то же самое, что система определения пола X0 . При гаплодиплоидии мужчины получают половину хромосом, которые получают женщины, включая аутосомы . В системе определения пола X0 мужчины и женщины получают равное количество аутосом, но когда дело доходит до половых хромосом, женщины получат две Х-хромосомы, а мужчины получат только одну Х-хромосому.
Механизмы
[ редактировать ]Было предложено несколько моделей генетических механизмов гаплодиплоидного определения пола. Чаще всего упоминается модель комплементарной аллели . Согласно этой модели, если особь гетерозиготна по определенному локусу , она развивается в самку, тогда как гемизиготные и гомозиготные особи развиваются в самцов. Другими словами, диплоидное потомство развивается из оплодотворенных яиц и обычно является самкой, тогда как гаплоидное потомство развивается в самцов из неоплодотворенных яиц. Диплоидные самцы будут бесплодны, поскольку их клетки не подвергаются мейозу для образования сперматозоидов. Следовательно, сперма будет диплоидной, а это означает, что их потомство будет триплоидным . Поскольку мать и сыновья перепончатокрылых имеют одни и те же гены, они могут быть особенно чувствительны к инбридингу : инбридинг уменьшает количество аллелей разного пола, присутствующих в популяции, тем самым увеличивая появление диплоидных самцов.
После спаривания каждая плодовитая самка перепончатокрылых хранит сперму во внутреннем мешочке, называемом сперматекой . Спаривающаяся самка контролирует высвобождение накопленной спермы изнутри органа: если она выпускает сперму, когда яйцеклетка проходит по ее яйцеводу , яйцеклетка оплодотворяется. [ 7 ] Социальные пчелы, осы и муравьи могут изменять соотношение полов внутри колоний, что максимизирует родство между членами и создает рабочую силу, соответствующую окружающим условиям. [ 8 ] У других одиночных перепончатокрылых самки откладывают неоплодотворенные яйца самцов на более бедные источники пищи, в то время как оплодотворенные яйца самок откладывают на более качественные источники пищи, возможно, потому, что на приспособленность самок будет более негативно влиять нехватка яиц в раннем возрасте. [ 9 ] [ 10 ] Манипулирование соотношением полов также практикуется гаплодиплоидными жуками-амброзиями , которые откладывают больше яиц самцов, когда у самцов больше шансов рассредоточиться и спариться с самками в разных местах. [ 11 ]
Определение пола у медоносных пчел
[ редактировать ]
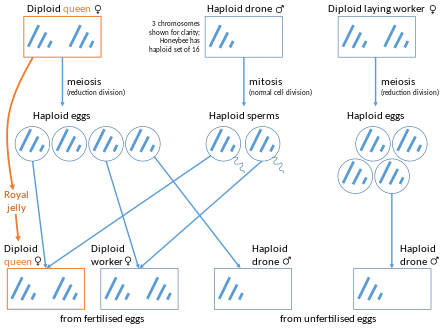
У медоносных пчел трутни (самцы) полностью происходят от королевы , их матери. Диплоидная королева имеет 32 хромосомы, а гаплоидные трутни - 16 хромосом. Дроны производят сперматозоиды , содержащие весь их геном, поэтому все сперматозоиды генетически идентичны, за исключением мутаций. Таким образом, генетический состав пчел-самцов полностью происходит от матери, тогда как генетический состав рабочих пчел-самок наполовину происходит от матери, а наполовину - от отца. [ 12 ] Таким образом, если пчелиная матка спаривается только с одним трутнем, любые две ее дочери в среднем будут делиться 3/4 своих генов . Геном диплоидной королевы рекомбинируется для ее дочерей, но геном гаплоидного отца наследуется его дочерьми «как есть». также может Рабочая пчела-несушка отложить неоплодотворенное яйцо, всегда мужского пола.
Встречаются редкие экземпляры диплоидных трутневых личинок. Это явление обычно возникает, когда происходит более двух поколений спаривания брата и сестры. [ 13 ] Определение пола у медоносных пчел изначально происходит за счет одного локуса, называемого комплементарным геном, определяющим пол ( csd ). У развивающихся пчел, если условия таковы, что особь гетерозиготна по гену csd , они разовьются в самок. Если условия таковы, что особь является гемизиготной или гомозиготной по гену csd , они разовьются в мужчин. Случаи, когда особь гомозиготна по этому гену, являются случаями диплоидных самцов. [ 14 ] Диплоидные самцы не доживают до взрослого возраста, так как рабочие пчелы-няньки съедают диплоидных самцов после вылупления. [ 15 ]
особи В то время как рабочие могут откладывать неоплодотворенные яйца, которые становятся их сыновьями, гаплодиплоидная система определения пола повышает приспособленность за счет непрямого отбора. Поскольку рабочий больше связан с дочерьми королевы (ее сестрами), чем со своим собственным потомством, помощь потомству королевы в выживании способствует распространению тех же генов, которыми обладает рабочий, более эффективно, чем прямое размножение. [ 16 ]
Партии рабочих пчел недолговечны и постоянно заменяются следующей партией, поэтому этот родственный отбор , возможно, является стратегией, обеспечивающей правильную работу улья. Однако, поскольку королевы обычно спариваются с дюжиной и более трутней, не все рабочие являются полнородными сестрами. Из-за отдельного хранения спермы трутней определенная партия выводка может быть более тесно связана, чем конкретная партия выводка, отложенная позже. Однако многие другие виды пчел, в том числе шмели, например Bombus terrestris , являются моноандрическими . [ 17 ] Это означает, что сестры почти всегда более связаны друг с другом, чем со своим собственным потомством, что устраняет конфликт изменчивого родства, присутствующий у медоносных пчел. [ 18 ]
Определение пола у хальцидоидных ос
[ редактировать ]У ос рода Nasonia документально подтвержден безCSD-метод определения пола. Самая последняя принятая модель для этой системы, не связанной с CSD, называется «Определение пола по геномному импринтингу по материнскому эффекту» (MEGISD). Эта модель включает ген маскулинизирующего/вирилизирующего материнского эффекта, который «импринтируется» на цитоплазматическом компоненте ооцитов, и «неимпринтированный» отцовский вклад (в потомстве женского пола), который обеспечивает противодействие вирилизации и позволяет происходить женскому развитию. Поскольку все диплоидные яйца становятся женскими (из-за фактора, обусловленного мужским генетическим вкладом, который предотвращает маскулинизацию), это отличается от CSD тем, что при CSD диплоидные яйца могут стать мужскими, если они гомозиготны или гемизиготны. [ 19 ]
Коэффициенты родства при гаплодиплоидии
[ редактировать ]Родство используется для расчета силы родственного отбора (по правилу Гамильтона ). [ 20 ] Гипотеза гаплодиплоидии предполагает, что необычное Коэффициент родства 3 ⁄ 4 среди полных гаплодиплоидных сестер отвечает за частоту эволюции эусоциального поведения у перепончатокрылых. [ 21 ] Эусоциальный работник, помогающий своей матери родить больше сестер, распространяет больше своих собственных генов, чем если бы она воспроизводила себя сама.
При нормальном половом размножении отец имеет два набора хромосом, и кроссинговер происходит , между хроматидами каждой пары во время мейоза , в результате которого образуется сперма . Следовательно, сперматозоиды не идентичны , поскольку в каждой хромосоме пары будут разные аллели во многих локусах . Но когда отец гаплоиден, все сперматозоиды идентичны (за исключением небольшого количества, где генные мутации произошли в зародышевой линии ). Таким образом, все потомки женского пола наследуют хромосомы самца на 100% в неизменном виде. Пока самка спарилась только с одним самцом , все ее дочери имеют полный набор хромосом этого самца. У перепончатокрылых самцы обычно производят достаточно спермы, чтобы хватило самке на всю ее жизнь после единственного спаривания с этим самцом. [ 20 ]
Коэффициенты родства у гаплодиплоидных организмов следующие, если предположить, что самка спаривалась только один раз. Эти соотношения применяются, например, ко всему пчелиному улью, если только некоторые рабочие-несушки не производят потомство, которое все будет самцами из неоплодотворенных яиц: в этом случае среднее родство будет ниже, чем показано.
Общие пропорции генов в гапло-диплоидных отношениях системы определения пола Секс Женский Мужской Дочь 1 ⁄ 2 1 Сын 1 ⁄ 2 — Мать 1 ⁄ 2 1 Отец 1 ⁄ 2 — Сестра 3 ⁄ 4 1 ⁄ 2 Брат 1 ⁄ 4 1 ⁄ 2 тетя по материнской линии 3 ⁄ 8 3 ⁄ 4 Дядя по материнской линии 1 ⁄ 8 1 ⁄ 4 тетя по отцовской линии 1 ⁄ 4 — Дядя по отцовской линии 1 ⁄ 4 — Племянница (дочь сестры) 3 ⁄ 8 1 ⁄ 4 Племянница (дочь брата) 1 ⁄ 4 1 ⁄ 2 Племянник (сын сестры) 3 ⁄ 8 1 ⁄ 4
Если исходить из того, что матери спариваются только один раз, сестры более тесно связаны друг с другом, чем со своими дочерьми. Этот факт был использован для объяснения эволюции эусоциальности у многих перепончатокрылых . Однако в колониях, в которых есть рабочие особи от нескольких маток или маток, которые спаривались несколько раз, родство между рабочими и рабочими будет меньше, чем родство между рабочими и дочерьми, как, например, у Melipona scutellaris .
См. также
[ редактировать ]- Эффект зеленой бороды
- Плоидность
- Псевдоарренотокия
- Половая дифференциация
- Рабочая полиция
- Х-хромосома
- Y-хромосома
Ссылки
[ редактировать ]- ^ Кинг, RC; Стэнсфилд, штат Вашингтон; Маллиган, ПК (2006). Генетический словарь (7-е изд.). Издательство Оксфордского университета. п. 194 . ISBN 978-0-19-530761-0 .
- ^ Гримальди, Д.; Энгель М.С. (2005). Эволюция насекомых . Издательство Кембриджского университета. п. 408 . ISBN 978-0-521-82149-0 .
- ^ Jump up to: а б Уайт, Майкл Джей Ди (1984). «Хромосомные механизмы размножения животных». Боллеттино ди Зоология . 51 (1–2): 1–23. дои : 10.1080/11250008409439455 . ISSN 0373-4137 .
- ^ Гримальди, Д.; Энгель М.С. (2005). Эволюция насекомых . Издательство Кембриджского университета. п. 465 . ISBN 978-0-521-82149-0 .
- ^ Хьюз, WOH; и др. (2008). «Наследственная моногамия показывает, что родственный отбор является ключом к эволюции эусоциальности». Наука . 320 (5880): 1213–1216. Бибкод : 2008Sci...320.1213H . дои : 10.1126/science.1156108 . ПМИД 18511689 . S2CID 20388889 .
- ^ Эдвард О. Уилсон (2005). «Родственный отбор как ключ к альтруизму: его взлет и падение». Социальные исследования . 72 (1): 159–166. дои : 10.1353/sor.2005.0012 . JSTOR 40972006 . S2CID 142713581 .
- ^ ван Вильгенбург, Эллен; Дриссен, Джерард и Бьюкебум, Лео В. Однолокусное комплементарное определение пола у перепончатокрылых: «неразумный» замысел? Границы в зоологии 2006, 3:1
- ^ Маховальд, Майкл; Веттберг, Эрик Определение пола в Суортморском колледже перепончатокрылых (1999 г.)
- ^ Чоу, А.; МакКауэр, М. (1996). «Последовательное распределение полов потомства у гиперпаразитоидной осы Dendrocerus carpenteri». Поведение животных . 51 (4): 859–870. дои : 10.1006/anbe.1996.0090 . S2CID 53192398 .
- ^ Ван Альфен, JJM; Тунниссен, И. (1982). «Выбор хозяев и распределение пола Pachycrepoideus Vindemiae Rondani (Pteromalidae) как факультативный гиперпаразитоид Asobara Tabida Nees (Braconidae; Alysiinae) и Leptopilina Heterotoma (Cynipoidea; Eucoilidae)» Нидерландский журнал зоологии . 33 (4): 497–514. дои : 10.1163/002829683X00228 .
- ^ Пер, К.; Таборский, М. (2004). «Самки жуков-амброзии корректируют соотношение полов в своем потомстве в соответствии с возможностями аутбридинга для своих сыновей» . Журнал эволюционной биологии . 17 (2): 257–264. дои : 10.1111/j.1420-9101.2003.00687.x . ПМИД 15009259 .
- ^ Синерво, Отбор Барри Кина и гаплодиплоидия у социальных перепончатокрылых. Архивировано 16 июля 2011 г. в Wayback Machine , 1997 г.
- ^ Войка, Дж.; Пщельництва, Заклад; Личинки трутней из оплодотворенных яиц медоносной пчелы. Архивировано 22 октября 2014 г. в Wayback Machine журнале исследований пчеловодства Journal , (1963), страницы 19–24.
- ^ Вайнсток, Джордж М.; Робинсон, Джин Э. и Консорциум по секвенированию генома медоносных пчел. Взгляд на социальных насекомых на основе генома медоносной пчелы Apis mellifera Nature , том «443» (2006), страницы 931-949.
- ^ Сантомауро, Джулия; Олдхэм, Нил Дж.; Боланд, Вильгельм; Энгельс Вольф; Каннибализм диплоидных личинок трутней у медоносных пчел (Apis mellifera) опубликован в журнале Odd Pattern of Circular Substance Journal of Apicultural Research , том «43» (2004), страницы 69-74
- ^ Фостер, Кевин Р.; Ратниекс, Фрэнсис Л.В. (2001). «Влияние предвзятого отношения к полу на эволюцию работы полиции в обществах перепончатокрылых» (PDF) . Американский натуралист . 158 (6): 615–623. дои : 10.1086/323588 . ПМИД 18707355 . S2CID 46052954 .
- ^ Баер, Б.; П. Шмид-Хемпель (2001). «Неожиданные последствия полиандрии для паразитизма и приспособленности шмеля Bombus terrestris ». Эволюция . 55 (8): 1639–1643. doi : 10.1554/0014-3820(2001)055[1639:ucopfp]2.0.co;2 . ПМИД 11580023 .
- ^ Дэвис, Николас Б., Джон Р. Кребс и Стюарт А. Уэст. (2012). Введение в поведенческую экологию . Уайли-Блэквелл. стр. 371–375.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Цзоу, Юань и др. «Химерный ген по отцовской линии инструктирует определение женского пола у гаплодиплоидной осы Nasonia». Наука 370 (2020): 1115–1118.
- ^ Jump up to: а б Гамильтон, WD (1996). Узкие дороги генной страны: собрание статей У. Д. Гамильтона . Оксфорд, Нью-Йорк: WH Freeman/Spektrum. ISBN 978-0-7167-4530-3 .
- ^ Кевин Р. Фостер; Том Венселерс; Фрэнсис Л.В. Ратниекс (2006). «Родственный отбор – ключ к альтруизму» . Тенденции в экологии и эволюции . 21 (2): 57–60. дои : 10.1016/j.tree.2005.11.020 . ПМИД 16701471 .
Библиография
[ редактировать ]- Бей, Мартин; и др. (1999). «Необычайно высокая скорость рекомбинации обнаружена в области полового локуса медоносной пчелы ( Apis mellifera )» . Генетика . 153 (4): 1701–1708. дои : 10.1093/генетика/153.4.1701 . ПМЦ 1460844 . ПМИД 10581277 .
- Ву, З.; и др. (2005). «Однолокусное комплементарное определение пола отсутствует у Heterospilus prosopidis (Hymenoptera: Braconidae)» . Наследственность . 95 (3): 228–234. дои : 10.1038/sj.hdy.6800720 . ПМИД 16077738 .
- Ратниекс, Фрэнсис (1988). «Репродуктивная гармония посредством взаимного контроля со стороны рабочих эусоциальных перепончатокрылых». Американский натуралист . 132 (2): 217–236. дои : 10.1086/284846 . JSTOR 2461867 . S2CID 84742198 .

пола Определение и дифференциация | |
|---|---|
| Обзор | |
| Генетическая основа |
|
| См. также | |